Capítulo 14. SISTEMAS DIGESTIVO
Y RESPIRATORIO
Y CAVIDADES CORPORALES
HISTOGÉNESIS DEL TRACTO INTESTINAL
GLÁNDULAS DEL SISTEMA DIGESTIVO
Formación del hígado
DUPLICACIONES, DIVERTÍCULOS Y ATRESIA
(ENFERMEDAD DE HIRSCHSPRUNG)
FÍSTULAS DEL INTESTINO POSTERIOR
MALFORMACIONES MACROSCÓPICAS DE LOS PULMONES
SÍNDROME DE DIFICULTAD RESPIRATORIA
(ENFERMEDAD DE MEMBRANA HALINA PULMONAR)
CASO CLÍNICO
El primitivo sistema digestivo debido al plegamiento lateral del endodermo se transforma en una estructura tubular que se ha descrito en el capítulo 5. A partir de un intestino tubular sencillo, el desarrollo del sistema digestivo puede analizarse a varios niveles, desde el alargamiento, las incurvaciones y los pliegues microscópicos del propio tubo, hasta la serie de inducciones e interacciones tisulares que constituyen la base para el desarrollo de las glándulas digestivas, hasta la maduración bioquímica de los epitelios secretor y de absorción asociados y presentes en el tracto digestivo. En las correlaciones clínicas 14-1, 14_2 y 14-3 se discuten las malformaciones asociadas con el sistema digestivo.
La formación del sistema respiratorio comienza con una discreta evaginación del intestino anterior. Sin embargo, en muy poco tiempo esta evaginación se determina en desarrollo distinto pero todavía seguirá algunos de los patrones básicos de las interacciones epitelio-mesénquima características de otras glándulas del intestino. Al comienzo, los sistemas digestivo y respiratorio se forman en una cavidad corporal común, más adelante se produce la división de esta cavidad primitiva por características distintas entre ambos sistemas. La correlación clínica 14_4 presenta las malformaciones asociadas con el sistema respiratorio y en la 14_5 se examinan las malformaciones relacionadas con otras cavidades corporales.
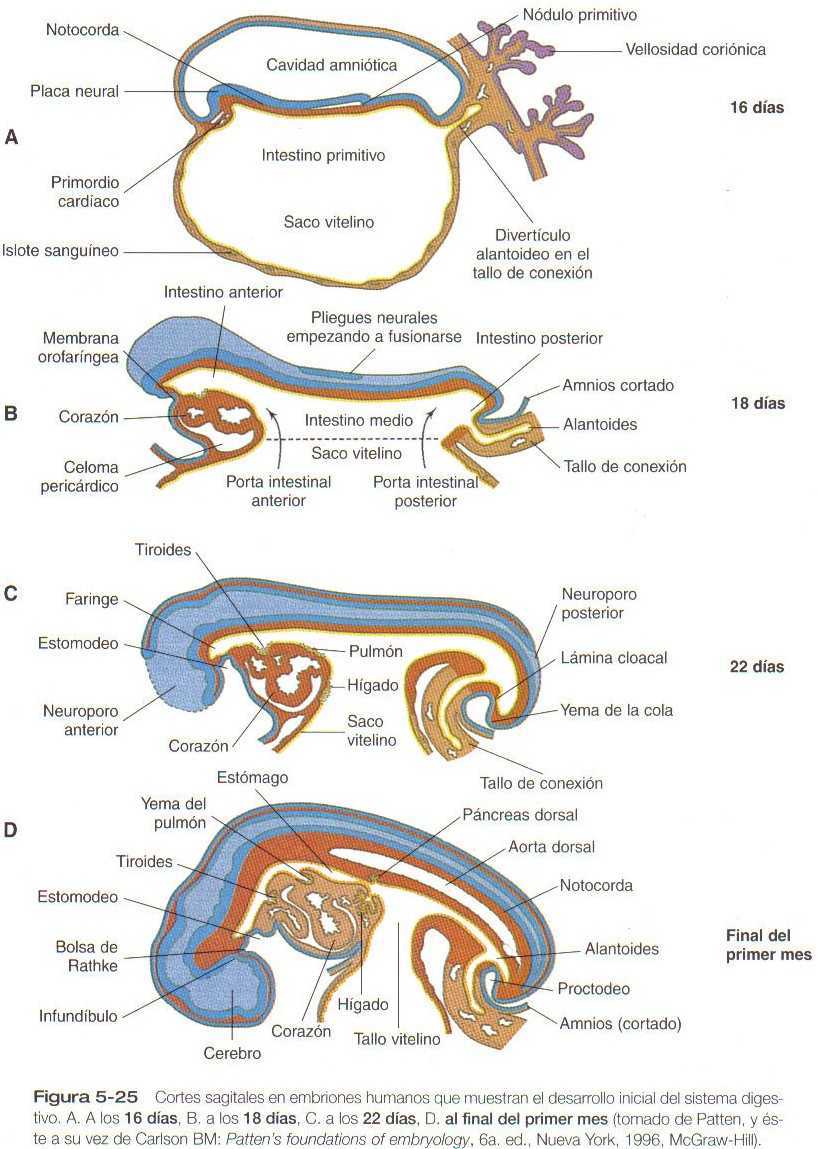
En el capítulo 5 se describe la formación del tubo digestivo endodérmico primitivo, delimitado en su extremo cefálico por la membrana orofaríngea y en su extremo caudal por la placa cloacal (figura 5_25). Debido a su íntima relación con el saco vitelino a través del pedículo vitelino (onfaloentérico) , puede dividirse el intestino en varias porciones: una anterior, una media comunicada con él, y una posterior. Al final del primer mes puede identificarse unos pequeños divertículos endodérmicos que representan los primordios de las principales glándulas digestivas (figura 14_1). El desarrollo de la faringe y sus derivados glandulares se explica en el capítulo 13.
La formación del tubo intestinal propiamente dicho comprende la elongación continua, la herniación fuera de la pared y la cavidad corporal, la rotación y el plegamiento para que quede bien alojado en la cavidad corporal, la histogénesis y la maduración funcional. Paralelas a estos procesos, las glándulas digestivas y las estructuras respiratorias están creciendo de acuerdo con complejos patrones de ramificación como resultado de las continuas interacciones epitelio – mesénquima. Estas interacciones también se producen en el tubo digestivo en desarrollo propiamente dicho, ya que allí, unas influencias mesenquimatosas específicas locales determinan el carácter del epitelio que tapiza esa parte del tracto digestivo.
Inmediatamente caudal a las bolsas faríngeas más caudales del embrión de cuatro semanas de vida, la faringe sufre un brusco estrechamiento y aparece una pequeña evaginación ventral (la yema del pulmón), según se ilustra en la figura 5_25. La región del intestino anterior que queda inmediatamente caudal a la yema del pulmón, es el esófago. Este segmento al comienzo es muy corto, y parece como si el estómago llegara casi hasta la faringe. En el segundo mes del desarrollo, durante el cual el intestino se alarga de forma considerable, el esófago adopta unas proporciones similares a las postnatales en relación con la localización del estómago.
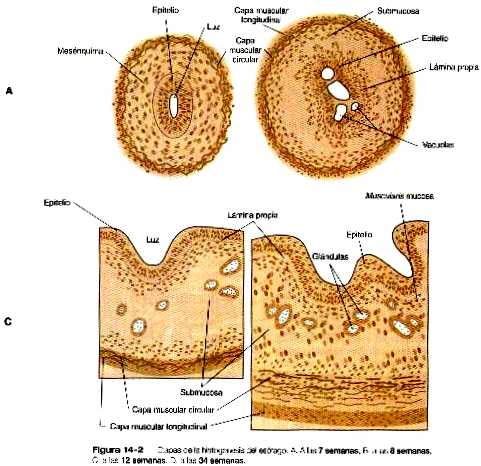
Aunque el esófago a nivel microscópico se asemeja a un simple tubo, experimenta una serie de cambios importantes de diferenciación a nivel tisular. En sus etapas más incipientes el epitelio endodérmico de revestimiento interno es cilíndrico estratificado. Hacia la octava semana, el epitelio ha ocluido parcialmente la luz del esófago y en él aparecen unas grandes vacuolas (figura 14_2).
En las semanas siguientes, las vacuolas se fusionan y la luz del esófago se recanaliza pero esta vez forma un epitelio ciliado de múltiples capas. Durante el cuarto mes, este epitelio es remplazado por fin por el epitelio escamoso estratificado que caracteriza al esófago adulto.
Externamente, en la pared del esófago, las capas musculares se diferencian como respuesta a señales inductivas procedentes del endodermo intestinal. El primordio de la capa muscular circular interna del esófago puede reconocerse muy pronto (a las cinco semanas de gestación) y hacia la octava semana se comienza a formar la capa longitudinal externa. La pared del esófago tiene músculo liso y esquelético. Las células de músculo liso se diferencian a partir del mesodermo esplácnico local asociado al intestino. Los experimentos realizados en embriones de ratón sugieren que las células de músculo esquelético presentes en la pared del esófago surgen debido a la transformación directa de las células de músculo liso preexistentes en músculo esquelético. Toda la musculatura esofágica está inervada por el neumogástrico o vago (X par).
Muy pronto, en el desarrollo del tracto digestivo es posible reconocer el estómago como una región dilatada cuya forma es bastante similar a la del estómago adulto ( figura 14_1)
La transformación directa de un tipo celular diferenciado en otro se llama transdiferenciación. Es un fenómeno infrecuente en los vertebrados, y se presenta en procesos como la regeneración del cristalino a partir de la cara dorsal del iris o la retina neural a partir de la retina pigmentaria en los anfibios.
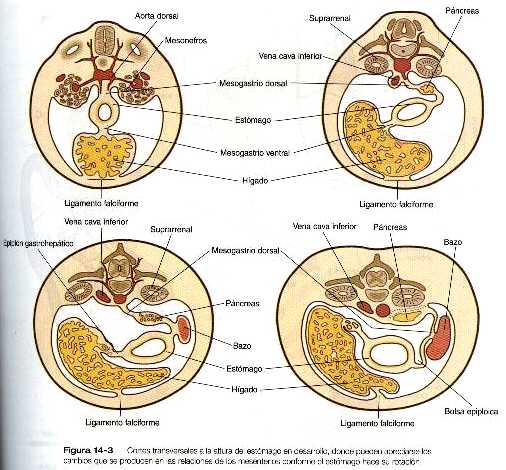
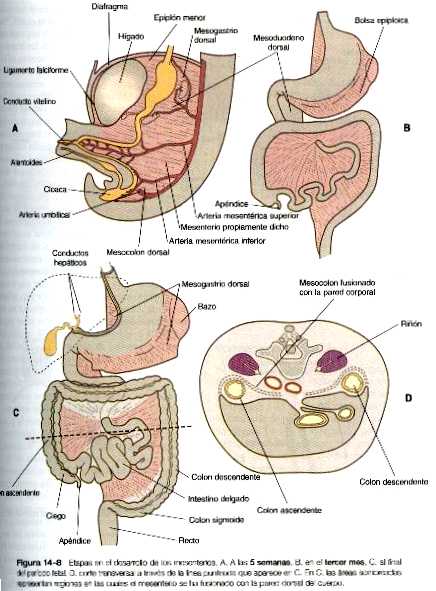
El estómago inicial está suspendido de la pared dorsal del cuerpo por una porción del mesenterio dorsal llamada mesogastrio dorsal. Está unido a la pared ventral del cuerpo mediante el mesenterio ventral, que también engloba al hígado en desarrollo (figura 14_3).
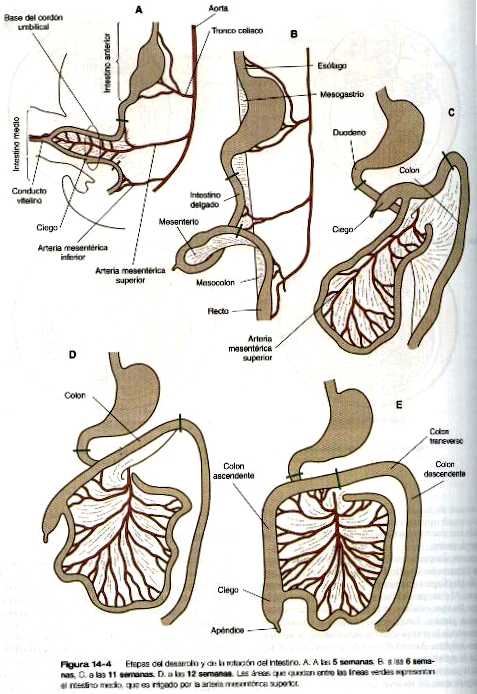
Cuando el estómago hace su aparición, su borde cóncavo mira hacia la región ventral, mientras que el convexo lo hace hacia la región dorsal. Dos desplazamientos concomitantes de la posición sitúan al estómago en su configuración adulta. El primero es una rotación de casi 90 grados sobre su eje craneocaudal, de forma que su borde convexo originalmente dorsal queda mirando hacia la izquierda y el borde cóncavo lo hace hacia la derecha. El otro desplazamiento consiste en una pequeña inclinación del extremo caudal (pilórico) del estómago en dirección craneal, de manera que el eje largo del estómago queda colocado algo diagonal con relación al eje del cuerpo (figura 14_4).
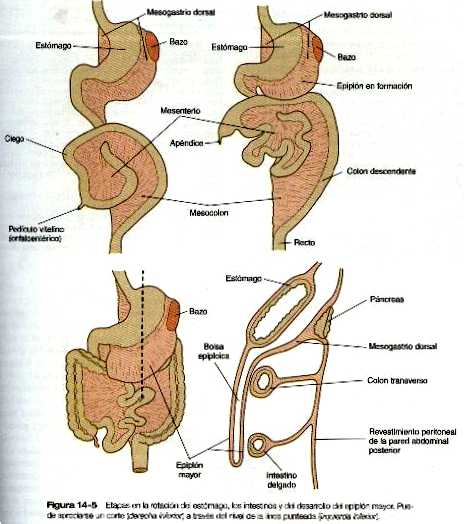
Durante la rotación, el estómago arrastra consigo al páncreas mesogastrio dorsal, dando lugar al desarrollo de una estructura llamada bolsa epiploica. Tanto el bazo como la cola del páncreas están incluidos en el mesogastrio dorsal (figura 14_3). Conforme el estómago realiza su rotación, el mesogastrio dorsal y la bolsa epiploica a la que encierra aumentan considerablemente. Pronto, parte del mesogastrio dorsal que conforman el epiplón mayor, cuelga por encima y por debajo del colon transverso y partes del intestino delgado como el delantal grande y doble de tejido adiposo (figura 14_5). Al final, las dos caras del epiplón mayor se fusionan, con lo que se oblitera la bolsa epiploica situada dentro de él.
El hígado en rápido crecimiento ocupa una porción cada vez mayor del mesenterio ventral.
En cuanto al aspecto histológico, la mucosa gástrica empieza a formarse hacia el final del segundo mes con la aparición de los pliegues (arrugas) y las primeras depresiones gástricas. Al principio del periodo fetal, comienzan a diferenciarse los tipos celulares individuales que caracterizan a la mucosa gástrica. Estudios bioquímicos y citoquímicos han demostrado la gradual diferenciación funcional de algunos tipos celulares durante el final del periodo fetal. En la mayoría de mamíferos, incluidos los seres humanos, las células de la mucosa gástrica comienzan a segregar ácido clorhídrico poco antes del nacimiento.
Los intestinos se forman de la parte caudal del intestino anterior, el medio y el posterior (tabla 14_1 falta). Hay dos puntos de referencia útiles para comprender la transformación macroscópica del intestino primitivo tubular con la forma de un cilindro relativamente recto hasta la compleja organización en plegamientos característica del tracto intestinal adulto. El primero es el pedículo vitelino, que se extiende desde el suelo del intestino medio hasta el saco vitelino (onfaloentérico). En el adulto, el lugar de situación del pedículo vitelino se encuentra en el intestino delgado más o menos a 60 cm craneal a la unión existente entre el intestino delgado y el grueso (válvula ileocecal).
En el lado dorsal del intestino primitivo una rama ventral impar de la aorta, la arteria mesentérica superior y sus ramas irrigan el intestino (figura 14_4). La arteria mesentérica superior misma sirve como eje alrededor del cual se producirá más tarde la rotación del intestino.
Ya en la quinta semana, el rápido crecimiento del tubo intestinal obliga a formar una asa similar a una horquilla. La principal causa de que el intestino adopte su posición adulta es una rotación en el sentido contrario a las manecillas del reloj, del brazo inferior del asa intestinal (con el anclaje del pedículo vitelino y la arteria mesentérica superior como puntos de referencia) alrededor del brazo superior cruzando la cara ventral de éste.
La principal consecuencia de esta rotación es que se lleva al colon por delante del intestino delgado, de manera que pueda adoptar con facilidad su posición en forma de “C” a lo largo de la pared ventral del abdomen (figura 14_4). Por detrás del colon, el intestino delgado experimenta un gran crecimiento en longitud y queda situado en su posición característica dentro de la cavidad abdominal.
Las malformaciones más frecuentes del esófago se asocian con anomalías del desarrollo del tracto respiratorio. Otras afecciones bastante infrecuentes son la estenosis y la atresia del esófago; la primera suele atribuirse a la recanalización anómala del esófago; después de la oclusión epitelial de su luz, en tanto que la segunda se asocia con más frecuencia con anomalías del desarrollo del tracto respiratorio. En ambas afecciones la alteración de la deglución en el feto puede llevar a un aumento excesivo del líquido amniótico (polihidramnios). Poco después del nacimiento, el neonato enfermo suele tener dificultad para deglutir la leche; la regurgitación y el atragantamiento durante la lactancia son indicaciones para examinar la permeabilidad del esófago.
La estenosis pilórica, que parece ser más fisiológica que anatómica, es la hipertrofia de la capa circular del músculo liso que rodea la esfínter pilórico (tracto de salida) del estómago. La hipertrofia causa un estrechamiento (estenosis) del esfínter pilórico e impide el paso de los alimentos. Varias horas después de la alimentación, el bebé vomita el contenido gástrico con violencia (vómito en proyectil). En ocasiones es posible palpar el píloro hipertrófico en el examen físico. Aunque por lo común la estenosis del píloro se trata mediante una sencilla incisión quirúrgica en la capa circular del músculo liso, a veces, la hipertrofia cede por sí sola sin tratamiento en algunas semanas. Aún no se conoce la patogénesis de este defecto, pero al parecer su etiología es genética. La estenosis congénita del píloro es más común en varones que en mujeres y se ha informado una incidencia de uno en 200 a uno por mil nacidos vivos.
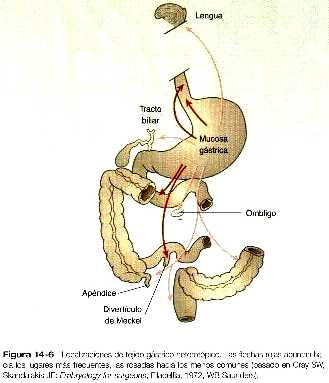
Mucosa gástrica heterotópica
La mucosa gástrica heterotópica se ha encontrado en varios órganos por lo demás normales (figura 14_6). Esta afección puede tener importancia clínica, porque si la mucosa heterotópica segrega ácido clorhídrico pueden formarse úlceras en localizaciones inesperadas.
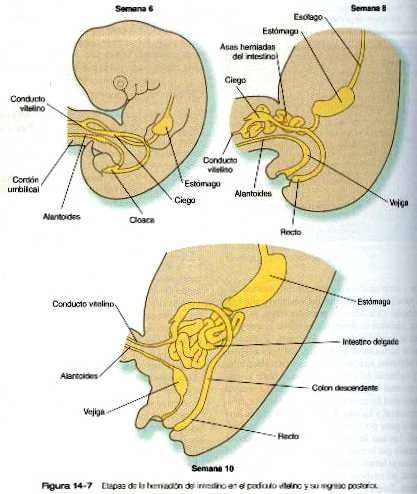
La rotación y otros cambios de posición del intestino tienen lugar en parte por que la longitud del intestino crece más que la del embrión. Casi desde las primeras etapas el volumen del tracto intestinal en desarrollo es mayor al que puede alojarse en la cavidad abdominal. Por tanto, el intestino en desarrollo se hernia en el interior del pedículo vitelino, que formará el cordón umbilical en el curso del desarrollo (figura 14_7). La hernia del intestino comienza hacia la sexta o séptima semanas de gestación. Hacia la décima semana la cavidad abdominal ha crecido lo suficiente para dar cabida al tracto intestinal, y las asas del intestino que se han herniado vuelven a pasar por el anillo umbilical hacia la cavidad abdominal. Las asas del intestino delgado se introducen primero, a medida que lo hacen, empujan la parte distal del colon- que nunca se hernió- hacia el lado izquierdo de la cavidad peritoneal, así establece la posición definitiva del colon descendente. Después de que el intestino delgado ha ocupado su posición intraabdominal, la parte proximal del colon que se había herniado también regresa a la cavidad, esta vez, su extremo cecal se sitúa hacia la derecha y hacia abajo (figura 14_4).
Durante esta rotación, hernias y reingresos, los intestinos están unidos a la pared dorsal del cuerpo por un mesenterio (figura 14_8). Conforme los intestinos adoptan sus posiciones definitivas en el interior de la cavidad abdominal, sus mesenterios los siguen. Partes del mesenterio asociado con el duodeno y el colon ( mesoduodeno y mesocolon) se fusionan con el revestimiento peritoneal de la pared dorsal del cuerpo.
Desde el comienzo de la sexta semana, puede observarse el primordio del ciego como una pequeña evaginación del brazo inferior del intestino medio (figura 14_4). En las semanas siguientes, la evaginación cecal se vuelve tan prominente que la pared distal del intestino delgado se continúa con el colon en ángulo recto.
La punta del ciego se alarga, pero su diámetro no aumenta en proporción con el resto. Este apéndice similar a un gusano recibe el apropiado nombre de apéndice vermiforme.
DIVISIÓN DE LA CLOACA
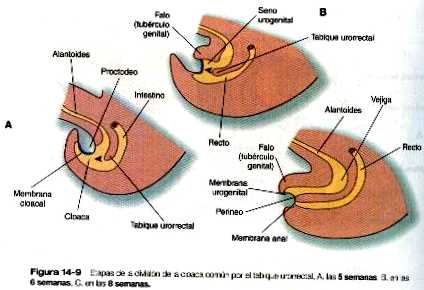
En el embrión joven, el extremo caudal del intestino posterior termina en la cloaca tapizada de endodermo, que en los vertebrados inferiores sirve como zona de terminación común de los sistemas digestivo y urogenital. La cloaca también comprende la base de la alantoides, que más adelante crecerá y formará el seno urogenital (capítulo 15). La membrana cloacal (proctodeo) consiste en varias capas de ectodermo y endodermo en aposición directa y actúa como barrera entre la cloaca y una depresión ectodérmica conocida como el proctodeo (figura 14_9). Un tabique de tejido mesodérmico llamado tabique urorrectal se sitúa entre el intestino posterior y la base de la alantoides. Durante la sexta y la séptima semanas, el tabique urorrectal crece hacia la membrana cloacal. Al mismo tiempo, unas crestas procedentes del mesodermo pericloacal se extienden desde las zonas laterales hacia el interior de la cloaca. La combinación del crecimiento hacia la línea media de las crestas laterales y el crecimiento del tabique urorrectal hacia la membrana cloacal divide a la cloaca en los senos rectal y urogenital (figura 14_9 B). Los mutantes dobles del Hoxa-13 y el Hoxd-13 dan como resultado la ausencia de la división de la cloaca junto con un subdesarrollo del falo (tubérculo genital). Una vez que la cloaca se divide, la membrana cloacal también se subdivide en una membrana anal, que cierra el extremo del intestino posterior, y una membrana urogenital, que cierra el seno urogenital y lo aísla del exterior. Hacia el final de la octava semana la membrana anal se rompe y permite la libre comunicación entre el intestino posterior y el exterior. El área en la cual se fusionan el tabique urorrectal y los pliegues mesodérmicos laterales con la membrana cloacal se convierte en el cuerpo perineal, que representa la zona de división entre los sistemas digestivo y urogenital.
HISTOGÉNESIS DEL TRACTO INTESTINAL
Poco después de su formación inicial el tracto intestinal presenta una capa simple de epitelio endodérmico columnar rodeada por una capa de mesodermo de la esplacnopleura. Hay tres fases principales en la histogénesis del epitelio intestinal: 1) una fase inicial de proliferación epitelial y morfogénesis, 2) un periodo intermedio de diferenciación celular en el cual aparecen los tipos celulares epiteliales característicos y 3) una fase final de maduración bioquímica y funcional de los diferentes tipos de células epiteliales. La pared mesenquimatosa del intestino también se diferencia en varias capas de músculo liso ricamente inervado y tejido conectivo. Existe un gradiente creaneocaudal general en la diferenciación en el intestino en desarrollo.
Al comienzo del segundo mes, el epitelio delgado empieza una fase de proliferación rápida cuyo resultado es que el epitelio ocasiona una oclusión temporal de la luz entre las semanas sexta y séptima. En un par de semanas la luz se ha recanalizado. Alrededor de esta época, aparecen varias pequeñas luces secundarias semejantes a hendiduras por debajo de la superficie del epitelio poliestratificado y unos cordones de mesodermo se invaginan hacia el epitelio. Una combinación de coalescencia de las luces secundarias y un crecimiento continuo y ascendente de mesénquima por debajo del epitelio provoca la formación de numerosas vellosidades intestinales en forma de dedo, que incrementan en gran medida la superficie de absorción del intestino. En esta época el epitelio se ha transformado de estratificado en cilíndrico simple.
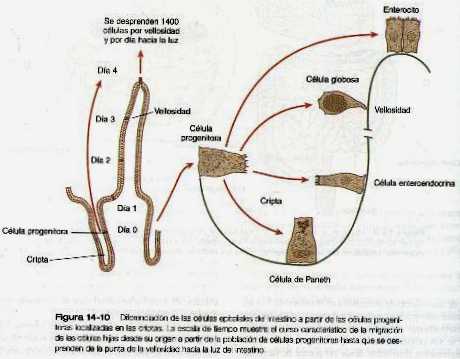
Con la formación de las vellosidades en su base también desarrollan las criptas intestinales. Estas criptas contienen células epiteliales progenitoras que tienen una alta tasa de mitosis y sirven como fuente de células epiteliales para toda la superficie intestinal (figura 14_10). Las pruebas experimentales sugieren que todas las células epiteliales de la misma cripta tienen un origen monoclonal (es decir, todas se derivan de una sola célula precursora en el embrión).
Los estudios autorradiográficos realizados después del nacimiento han demostrado que en tres o cuatro días las células epiteliales que se han originado en las criptas, migran y ascienden para desprenderse finalmente desde las puntas de las vellosidades. El epitelio intestinal está en constante renovación gracias a este mecanismo.
Poco después de la formación de las criptas, las células progenitoras epiteliales individuales que están en su interior, comienzan a formar uno de los cuatro tipos de células epiteliales maduras que se encuentran en el revestimiento interno del intestino *.
Hacia el final del segundo trimestre del embarazo, todos los tipos de células que se encuentran en el revestimiento intestinal del adulto se han diferenciado pero muchas de ellas carecen del patrón funcional del adulto. Numerosos patrones bioquímicos específicos ya están definidos a las 12 semanas de gestación y maduran durante el periodo fetal. Por ejemplo, la lactasa, una enzima que degrada el disacárido lactosa (azúcar de la leche) , es una de las enzimas digestivas que se sintetizan durante el periodo fetal como preparación para el periodo postnatal inmediato durante el cual el recién nacido subsiste en principio gracias a la leche materna. La posterior diferenciación bioquímica del intestino tiene lugar después del nacimiento, a menudo en respuesta a patrones alimentarios específicos.
La histodiferenciación del tracto intestinal no es una propiedad aislada de los componentes tisulares individuales de la pared del intestino. Durante el periodo embrionario inicial y en ocasiones hasta la vida postnatal los componentes epitelial y mesodérmico de la pared intestinal se comunican por medio de interacciones inductivas. Los experimentos de recombinación interespecies han demostrado que el mesodermo intestinal ejerce una influencia regional sobre la diferenciación del epitelio.
La diferenciación enzimática final de las células de absorción del intestino recibe una intensa influencia de los glucocorticoides, y el mesodermo subyacente parece mediar en este efecto hormonal. Una acción inductiva ejercida por el endodermo intestinal provoca la diferenciación del músculo liso a partir de las células mesenquimatosas de la pared intestino. El mecanismo de las interacciones entre el mesénquima y el endodermo aún no está claro. En algunos casos el mesodermo puede ejercer sus efectos directamente sobre el endodermo, en tanto que en otros, las moléculas de la matriz extracelular segregadas por las células mesodérmicas pueden mediar en las interacciones inductivas.
Aunque el intestino desarrolla muchas de sus capacidades funcionales durante el periodo fetal, no se produce ninguna de las principales funciones digestivas hasta que comienza la alimentación después del nacimiento. Los intestinos del feto contienen un material verdoso llamado meconio (figura 17_9), que es una mezcla de vello del lanugo y de vérmix caseosa desprendida de la piel, células descamadas del intestino, secreciones biliares y otros materiales deglutidos con el líquido amniótico.
Durante muchos años se creyó que las células secretoras de las hormonas peptídicas en el tracto intestinal se derivaban de la cresta neural. Los experimentos de transplante y marcaje de células no han aportado datos que permitan comprobar esta afirmación.
GLÁNDULAS DEL SISTEMA DIGESTIVO
Las glándulas del sistema digestivo surgen en virtud de procesos inductivos entre las evaginaciones epiteliales y el mesénquima circundante. Los diversos epitelios glandulares tienen requerimientos que difieren considerablemente en cuanto a los tipos de mesénquima que puede necesitar para su desarrollo. Por ejemplo, en los estudios de recombinación de tejidos, el epitelio pancreático experimenta un desarrollo característico cuando se combina con mesénquima de prácticamente cualquier origen. Por su parte, la formación de las glándulas salivales se mantiene por mesénquima del pulmón o de las glándulas sexuales accesorias, pero no por muchos otros tipos de mesénquima. El soporte inductivo del epitelio normal es mantenido por el mesénquima derivado de la placa lateral o el mesodermo intermedio, el mesénquima axial (ya sea somítico o de la cresta neural) no mantiene la diferenciación hepática. Las propiedades inductivas de ciertos mesénquimas glandulares pueden correlacionarse con los diferentes modelos de vascularización de los mismos.
Formación del hígado
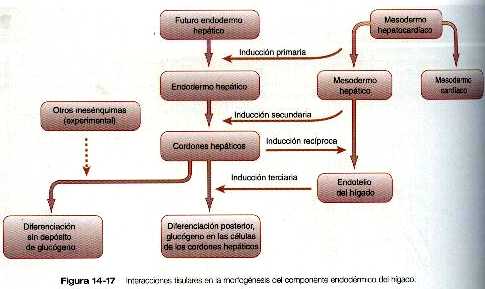
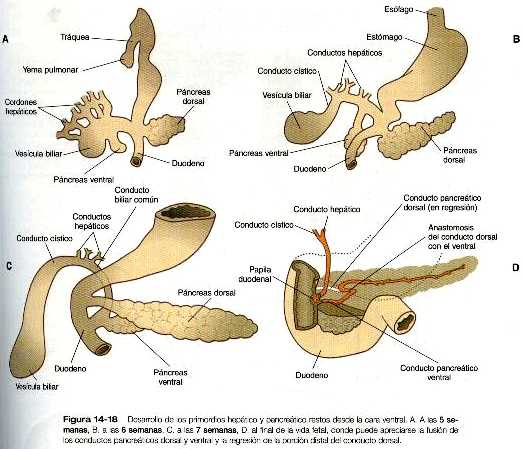
Al comienzo de la tercera semana, surge un divertido endodérmico hepático del suelo del intestino anterior y crece en el mesénquima del tabique transverso (septum transversum) figura 5-25. El divertículo hepático es la manifestación de una serie de procesos inductivos que ya habían empezado (figura 14_17); el divertículo primitivo se ramifica en muchos cordones hepáticos, que guardan una estrecha asociación con el mesodermo esplácnico del tabique transverso. El mesodermo permite el crecimiento continuo y la proliferación del endodermo hepático, en parte, gracias a la acción del factor de crecimiento hepático. Este factor es fijado por la molécula receptora, c-met, localizada en la superficie de los hepatocitos endodérmicos. Diversos estudios experimentales han demostrado que el mesodermo ya sea de los componentes esplacnopleural o somatopleural del mesodermo de la placa lateral pueden sostener el crecimiento y la diferenciación hepática posterior, no así el mesodermo paraxial, cuya capacidad para mantener el desarrollo hepático es limitada.
Además de los cordones endodérmicos hepáticos, en el hígado en desarrollo se forma un sistema de conductos de drenaje biliar. Cerca del área en la cual confluyen los conductos hepáticos, la presencia de una dilatación indica el desarrollo de la vesícula biliar (figura 14_18). Los cordones hepáticos forman una serie de láminas irregulares de disposición laxa que alteran con los sinusoides tapizados de mesodermo, a través de los cuales, la sangre se filtra e intercambia los nutrientes de los hepatocitos. El hígado en desarrollo está muy vascularizado y muchos grandes vasos discurren en él durante el periodo embrionario.
La estenosis y la atresia del duodeno son el resultado de la no recanalización o de la recanalización incompleta de la luz duodenal después de haber sido obliterada por el epitelio. Estas malformaciones son infrecuentes.
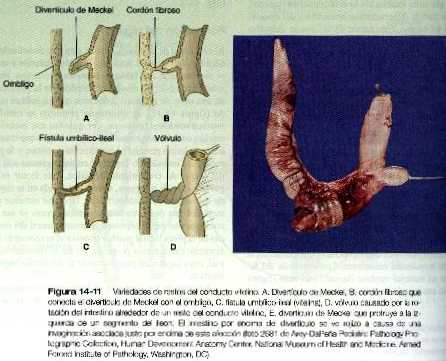
La variedad más común de las anomalías del tracto intestinal es alguna forma de persistencia del conducto vitelino. La más frecuente es el divertículo de Meckel, presente en el 2 al 4% de la población; la forma más típica de esta afección es un corto saco ciego situado en el borde antimesentérico del ileon, aproximadamente a 50 cm de la válvula ileocecal en dirección craneal (figuras 14_11 A y E). Esta estructura representa la persistencia de la porción proximal del conducto vitelino (onfaloentérico). Los divertículos de Meckel simples suelen ser asintomáticos, pero en ocasiones se inflaman o contienen tejido ectópico (por ejemplo gástrico, pancreático e incluso endometrial, que puede ulcerarse).
En algunos casos, un ligamento conecta al divertículo de Meckel con el ombligo (figura 14_11 B) o un ligamento vitelino delgado que puede contener una arteria vitelina persistente conecta el intestino con el ombligo. En ocasiones el intestino rota alrededor de este ligamento, lo cual da lugar a una afección conocida como vólvulo (figura 14_11 D), que puede acompañarse de una estrangulación del intestino.
Un conducto vitelino persistente puede formar una fístula vitelina (figura 14_11 C), que es una comunicación directa entre la luz del intestino y el exterior del cuerpo a través del ombligo. Rara vez se presenta un quiste del conducto vitelino a lo largo del ligamento vitelino.
El onfalocele representa la ausencia de retorno de las asas intestinales a la cavidad corporal durante la décima semana. Después del nacimiento, las asas intestinales pueden verse en el interior de un saco casi transparente conformado por el amnios en el exterior y la membrana peritoneal en el interior (figura 14_12). La incidencia del onfalocele se acerca a uno de cada 3,500 nacimientos, pero la mitad de los bebés afectados muere antes de nacer.
En la hernia umbilical congénita, bastante común en los bebés prematuros, los intestinos regresan de forma normal hacia la cavidad corporal, pero la musculatura (recto mayor del abdomen) de la pared ventral del abdomen no cierra el anillo umbilical, lo que permite que una cantidad variable de epiplón o del intestino salga del abdomen a través del ombligo. A diferencia del onfalocele, el tejido que se hernia está cubierto de piel y no de membrana amniótica.
Tanto el onfalocele como la hernia umbilical congénita se asocian con defectos de cierre de la pared ventral del abdomen. Si estos defectos son grandes, pueden acompañarse de la protrusión
masiva del contenido abdominal o de otros defectos de cierre como la extrofia vesical (capítulo 15).
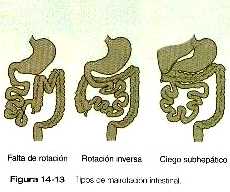
En ocasiones los intestinos no rotan o lo hacen de forma anómala al regresar a la cavidad abdominal. Esto puede dar como resultado un amplio espectro de anomalías anatómicas (figura 14_13), que en la mayoría de los casos son asintomáticas, pero que en ocasiones, pueden dar lugar a un vólvulo u otra forma de estrangulación del intestino.
DUPLICACIONES, DIVERTÍCULOS Y ATRESIA
(ENFERMEDAD DE HIRSCHSPRUNG)
La base del megacolon agangliónico, que se manifiesta por una gran dilatación de ciertos segmentos del colon, es la ausencia de ganglios parasimpáticos en las paredes afectadas del colon. Esto se ha atribuido a un defecto en la migración de las células de la cresta neural hacia esa porción del intestino posterior al comienzo del segundo mes del embarazo. Existen pruebas obtenidas en ratones mutantes en los cuales se desarrollan segmentos agangliónicos del colon, que sugieren que el ambiente local del intestino inhibe la migración de las células de la cresta neural hacia el segmento afectado. Esto se demostró en experimentos en los cuales las células de la cresta de los ratones mutantes podía colonizar al intestino normal, pero las células normales de la cresta no podían migrar hacia segmentos de intestino de ratones como resultado de la sobreproducción de endotelina-3 puede ser la causa que detenga la migración desde la cresta neural. El colon distal es la región más afectada por la ausencia de ganglios, pero en un pequeño porcentaje de los casos, los segmentos agangliónicos se extienden tanto en dirección craneal que alcanzan el colon ascendente. Los datos de la frecuencia del megacolon presentan grandes variaciones, desde uno por cada mil hasta uno por cada 30 mil nacimientos.
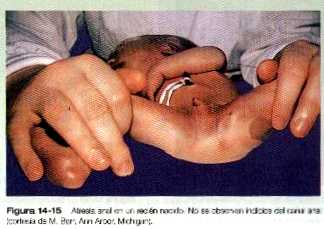
El ano imperforado abarca un espectro de defectos que pueden ir desde la presencia de una simple membrana que cubre el orificio anal (persistencia de la membrana anal), hasta la atresia de longitud variable del canal anal, el recto o de ambos. Desde el punto de vista macroscópico se caracteriza por la ausencia de orificio anal (figura 14_15). En todo examen neonatal debe determinarse la presencia del orificio anal., La longitud del segmento atrésico reviste enorme importancia cuando se considera el tratamiento quirúrgico del ano imperforado. El tratamiento de la membrana anal persistente puede ser sencillo, en tanto que los defectos más extensos, en especial los que implican a la musculatura anal, constituyen problemas quirúrgicos muy complejos
FÍSTULAS DEL INTESTINO POSTERIOR
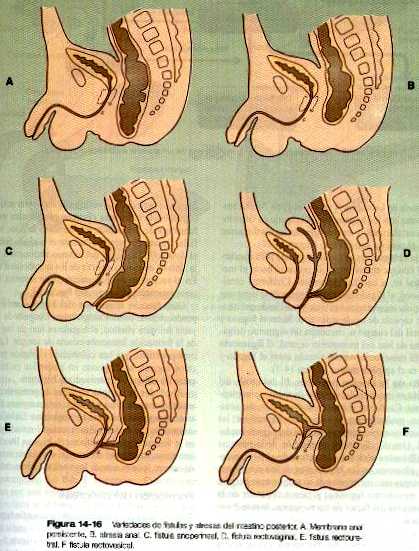
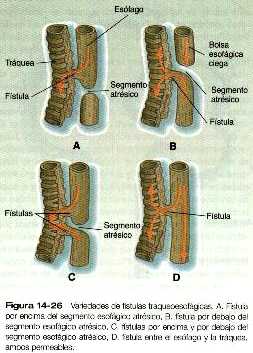
En muchos casos, la atresia se acompaña de una fístula que comunica la porción permeable del intestino posterior con otra estructura perteneciente a la región del seno urogenital original. Los tipos más comunes de fístulas comunican el intestino posterior con la vagina, la uretra o la vejiga y otras pueden comunicarse con la superficie del área perineal (figura 14_16).
Pronto, todo el hígado es demasiado grande para quedarse dentro del tabique transverso, y se proyecta hacia el mesenterio ventral en la cavidad abdominal. Conforme continúa su expansión, el hígado que crece rápidamente tiene una cubierta de tejido mesentérico brillante y traslúcida que ahora le sirve de cápsula de tejido conectivo. Entre el hígado y la pared ventral del cuerpo se encuentra un segmento delgado y en forma de hoz del mesenterio ventral, el ligamento falciforme. El mesenterio ventral situado entre el hígado y el estómago es el epiplón menor (figura 14-3).
Desarrollo de la función hepática. El desarrollo del hígado no sólo es cuestión de aumento de masa y complejidad estructural. Conforme el hígado se desarrolla, sus células adquieren gradualmente la capacidad de realizar las numerosas funciones bioquímicas propias del hígado maduro funcional. Una de ellas es la producción de la proteína plasmática albúmina sérica. Se ha detectado el RNA mensajero de la albúmina en los hepatocitos de los mamíferos durante las etapas más incipientes de su crecimiento hacia el interior del mesodermo hepático, y parece depender de la expresión primaria del factor de trascripción, factor-3 nuclear hepático.
Una de las principales funciones del hígado adulto es la síntesis y el almacenamiento del glucógeno, que se utiliza como una forma de almacenar carbohidratos. Conforme avanza el periodo fetal, el hígado almacena activamente glucógeno. Las hormonas corticosuprarrenales estimulan esta función y de forma indirecta también lo hace la adenohipófisis. De manera similar, durante el periodo fetal se produce el desarrollo funcional del sistema enzimático implicado en la síntesis de la urea a partir de los metabolitos nitrogenados. En el momento del nacimiento, este sistema ha alcanzado completamente si capacidad funcional.
Otra función importante del hígado embrionario es la producción de células sanguíneas. Después de la hematopoyesis del saco vitelino, el hígado es uno de los lugares clave de la formación intraembrionario de sangre. Las células hematopoyéticas, que han colonizado el hígado procedentes de otros lugares, se disponen en pequeños racimos dispersos entre las células del parénquima hepático.
Alrededor de las doce semanas de gestación los hepatocitos comienzan a producir bilis, en gran parte como producto de la degradación de la hemoglobina. La bilis drena por el sistema de conductos biliares recién formado y se almacena en la vesícula biliar. Conforme la bilis va liberándose en los intestinos, va tiñendo el resto del contenido intestinal de verde oscuro, que es una de las características del meconio.
Formación del páncreas
Poco después de la aparición del primordio hepático, dos yemas pancreáticas, comienzan a crecer a partir de las paredes dorsal y ventral del intestino anterior (figura 5_25).
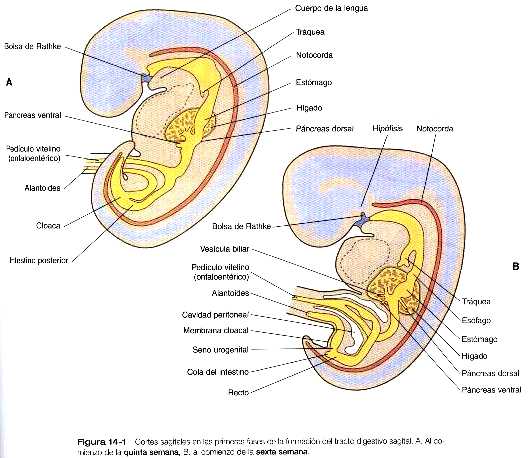
La yema dorsal es una evaginación que surge directamente del endodermo duodenal, mientras que la ventral se origina en el endodermo del divertículo hepático (figura 14_18). La yema pancreática dorsal se induce a partir del endodermo del intestino dorsal por la notocorda, que al comienzo del desarrollo se encuentra en contacto directo con el endodermo (figura 4_7 C) . Al parecer, la yema pancreática ventral es inducida por el mesodermo hepático. Durante las primeras etapas de la formación de la yema pancreática, el homeodominio de la formación de la yema pancreática, el homeodominio de trascripción del Pdx-1 se expresa. Cuando este gen se elimina por una mutación inducida, el desarrollo de la yema pancreática se detiene. El Islet-1, otro gen de homeodominio LIM que se expresa de forma precoz, es necesario para la formación del mesodermo pancreático dorsal y de los islotes. La expresión de estos dos genes (así como la del Pax-6) en la yema pancreática dorsal se elimina en ausencia de la influencia de la notocorda, pero es importante resaltar que su expresión en la yema pancreática ventral no se ve afectada por la extirpación de la notocorda.
Durante la fase inicial del crecimiento, el páncreas dorsal adquiere un tamaño mucho mayor que el ventral. Casi al mismo tiempo el duodeno gira hacia la derecha y da lugar a una asa en forma de ”C”, esto hace que el páncreas ventral y el colédoco queden detrás de él y dentro del mesenterio dorsal. El páncreas ventral pronto establece contacto con el páncreas doral y los dos se fusionan.
Ambos páncreas, el dorsal y el ventral, poseen un conducto principal. Después de la fusión de los dos primordios, el conducto principal del páncreas ventral se anastomosa con el conducto del páncreas dorsal. La porción del conducto pancreático dorsal que queda entre la anastomosis y el duodeno suele regresar, lo que convierte al conducto principal del páncreas ventral (conducto de Wirsung) en la vía de salida definitiva de las secreciones pancreáticas hacia el duodeno (figura 14_18).
El páncreas es un órgano mixto con funciones endocrinas y exocrinas. La porción exocrina consiste en un gran número de acinos, que están conectados con el sistema de conductos secretores. El componente endocrino está formado por cerca de un millón de islotes de Langerhans muy vascularizados y dispersos entre los acinos.
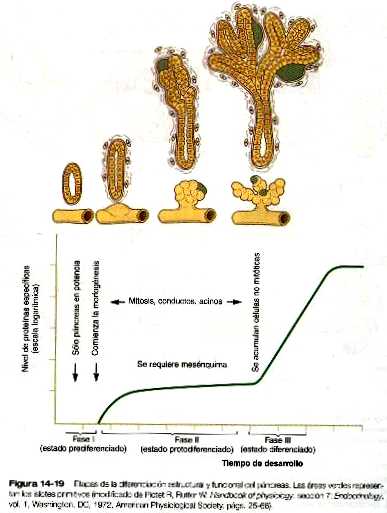
Una vez definidos, los primordios pancreáticos crecen en virtud de varias interacciones epitelio-mesénquima como las que suelen verse en el desarrollo de las glándulas asociadas con el intestino. Durante la fase de evaginación y crecimiento, el epitelio glandular del páncreas se forma gracias a la gemación secuencial de cordones de células derivadas de la población de células pancreáticas primarias (figura 14_19). A partir de los cordones celulares, se diferencian tanto los acinos como sus conductos. Los experimentos de recombinación de tejidos, realizados tanto in vitro como in vivo, han demostrado que el mesénquima es necesario para la formación de los acinos, pero que los conductos pueden formarse sin él si se ponen a las células precursores endodérmicas en contacto con un gel rico en material de membrana basal.
Se presentan muchas alteraciones o variaciones menores en la forma del hígado o de las vías biliares, pero por lo general carecen de importancia clínica. Una de las malformaciones más importantes del hígado es la atresia biliar. Puede afectar a cualquier nivel, desde los minúsculos conductillos hasta los grandes conductos que transportan la bilis. Los neonatos afectados por esta malformación, suelen presentan una pronunciada ictericia muy poco después de nacer. Algunos casos son susceptibles de tratamiento quirúrgico, mientras que otros requieren un transplante hepático.
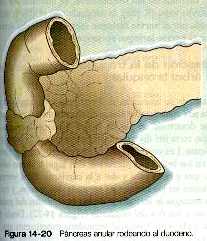
En raras ocasiones un anillo de tejido pancreático rodea por completo al duodeno, lo que se conoce como páncreas anular (figura 14_20). A veces puede causar la obstrucción del duodeno después del nacimiento. No se ha establecido la etiología de esta malformación, pero la explicación más aceptada es que el crecimiento de un páncreas ventral bífido puede rodear al duodeno por ambos lados.
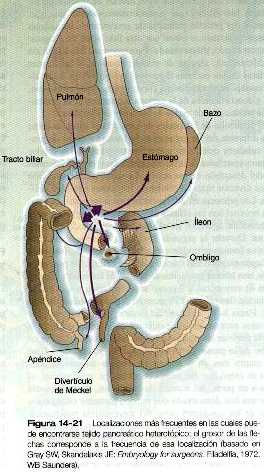
En ocasiones puede encontrarse tejido pancreático heterotópico a lo largo del tracto digestivo y se presenta con mayor frecuencia en el duodeno o en la mucosa gástrica (figura 14_21). Cerca del 6% de los divertículos de Meckel contienen tejido pancreático heterotópico.
Aunque se requiere la presencia de mesénquima para la diferenciación de los acinos, éste no tiene que ser de origen pancreático. In vitro, el endodermo pancreático combinado mesodermo de las glándulas salivales se diferencia aún mejor que cuando se expone al propio mesénquima pancreático. Esto demuestra que en el caso del páncreas la influencia inductiva del mesénquima es permisiva antes que instructiva.
La diferenciación de los acinos se divide en tres fases (figura 14_19). La primera, llamada estado prediferenciado, se produce mientras que los primordios pancreáticos están empezando a formarse. Se establece una población de células pancreáticas primarias que muestran niveles prácticamente indetectables de actividad de las enzimas digestivas. Conforme las yemas pancreáticas comienzan a crecer hacia el exterior, el epitelio experimenta una transición hacia un segundo estado protodiferenciado. Durante esta fase, las células exocrinas presentan bajos niveles de muchas de las enzimas hidrolíticas que posteriormente van a producir. Después del periodo principal de crecimiento, las células acinares pancreáticas pasan por otra fase antes de alcanzar un tercer estado diferenciado. Para entonces han adquirido un diferenciado aparato de síntesis proteica y las formas inactivas de las enzimas digestivas polipeptídicas están almacenadas en el citoplasma como gránulos de zimógeno. Los glucocorticoides de la corteza suprarrenal fetal estimulan la producción de varias de las enzimas digestivas.
El desarrollo de los islotes de Langerhans sigue un curso algo diferente del de los acinos. Se forman a partir de grupos celulares epiteliales durante la segunda fase (protodiferenciada) del desarrollo de los acinos. La secuencia de aparición de los diversos tipos de células de los islotes está bien definida. Las primeras en diferenciarse (hacia las semanas octava y novena) son las células alfa (que segregan glucagón). A continuación aparecen las células beta, que producen la insulina. Durante la segunda fase de la diferenciación pancreática (estado protodiferenciado) los niveles de síntesis de glucagón exceden con mucho a los de insulina. Un poco más tarde, una tercera población de células de los islotes (las células δ) comienzan a secretar somatostatina y una cuarta población (las células PP) secretan el polipéptido pancreático. Hacia la tercera fase del desarrollo del páncreas, los gránulos secretorios son evidentes en el citoplasma de la mayor parte de las células de los islotes. La insulina y el glucagón están presentes en la circulación fetal hacia el final del quinto mes de la gestación. Aunque durante muchos años se creyó que los islotes pancreáticos se derivaban, al menos en parte, del sistema de conductos, investigaciones recientes han demostrado que pueden formarse en ausencia tanto de los conductos como del mesénquima pancreático. Esto sugiere que el desarrollo de los islotes en realidad puede deberse a un proceso por defecto del endodermo pancreático que no se ha expuesto al mesodermo. La expresión del glucagón y la insulina en el endodermo de la yema pancreática muy incipiente apoya este concepto.
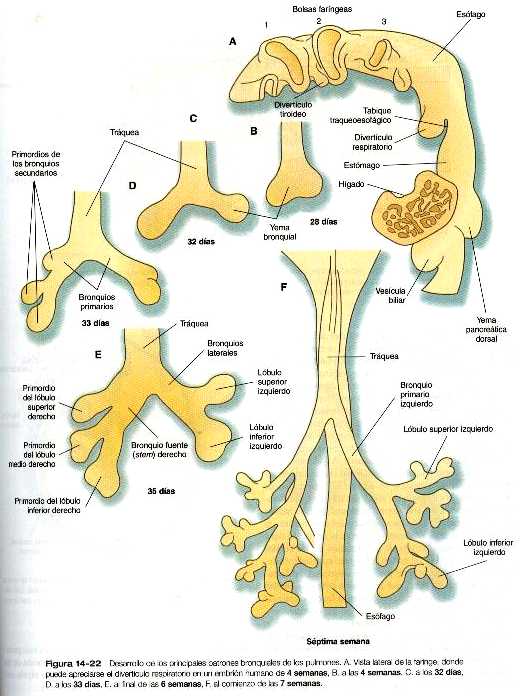
El sistema respiratorio se ve por vez primera durante la cuarta semana de gestación como un surco laringotraqueal, casi imperceptible en la línea media ventral en el límite caudal de la región faríngea. Hacia la quinta semana, el crecimiento ha transformado el surco laringotraqueal en un divertículo respiratorio bien definido (figura 5_25) que crece casi paralelo al esófago hacia el mesodermo esplácnico. Por medio de una serie de interacciones con el mesodermo circundante, el divertículo respiratorio se alarga y forma una porción traqueal y comienza a dar origen a los primeros 23 grupos de bifurcaciones (yemas pulmonares) que se forman incluso hasta la vida postnatal.
Durante la cuarta y la quinta semanas de la gestación, una rápida proliferación del mesénquima del cuarto y el sexto arcos bronquiales alrededor del lugar de origen de la yema respiratoria en el esófago convierte la hendidura de apertura en una glotis en forma de T limitada por dos engrosamientos aritenoideos laterales y una epiglotis craneal. El mesénquima que rodea al orificio laríngeo termina por diferenciarse en los cartílagos tiroides, cricoides y aritenoides, que forman el soporte esquelético de la laringe. A semejanza del esófago, la luz de la laringe sufre una obliteración epitelial transitoria. En el proceso de recanalización durante las semanas nueve y diez, un par de pliegues y recesos laterales forman la base estructural de las cuerdas vocales y de los ventrículos laríngeos adyacentes. La musculatura de la laringe, derivada de los somitómeros, es inervada por ramas del nervio vago (x par); la asociada con el cuarto arco es inervada por el nervio laríngeo superior, y la del sexto arco por el nervio laríngeo recurrente.
Después de su aparición, el divertículo respiratorio experimenta una considerable elongación antes de que en su extremo se desarrolle un par de yemas broncopulmonares. La porción recta del divertículo respiratorio es el primordio de la tráquea. Las yemas bronquiales, que terminan por convertirse en los bronquios primarios, dan lugar a yemas adicionales: tres a la derecha y dos a la izquierda. Estas yemas se convierten en los bronquios secundarios, o stem, y su número presagia la formación de los tres lóbulos del pulmón derecho y los dos del izquierdo (figura 14_22). Desde este punto, cada yema bronquial secundaria experimenta una larga serie de ramificaciones hasta que se completan 23 generaciones sucesivas de ramas. La morfogénesis del pulmón continúa después del nacimiento y la estabilización del patrón morfológico de este órgano sólo se logra cerca de los ocho años de edad. Al comienzo del desarrollo del tracto respiratorio, se expresa una gran cantidad de genes Hox (Hoxa-3 a Hoxa-5 y Hoxb-3 a Hoxb – 6). Los patrones combinados de expresión de estos genes pueden estar implicados en la especificación regional del tracto respiratorio.
El mesodermo que rodea al endodermo controla en grado de ramificación en el tracto respiratorio. Diversos experimentos de recombinación de tejidos han demostrado que el mesodermo que rodea a la tráquea inhibe la ramificación, en tanto que el que rodea a las yemas bronquiales la favorece. Si se combina endodermo traqueal con mesodermo bronquial, se produce una gemación anómala. A la inversa, si se coloca mesodermo traqueal alrededor del endodermo bronquial, se inhibe la gemación bronquial. El mesodermo de otros órganos, por ejemplo, las glándulas salivales, puede inducir la gemación del endodermo bronquial, pero se induce un patrón de ramificación característico del mesodermo: un mesodermo capaz de promover o sostener la gemación tiene que mantener una alta tasa de proliferación de las células epiteliales. En general, el patrón del órgano epitelial está determinado en su mayor parte por el mesodermo. La diferenciación estructural y funcional del epitelio es una propiedad específica de las células epiteliales, pero en fenotipo epitelial coincide con la región de procedencia del mesodermo.
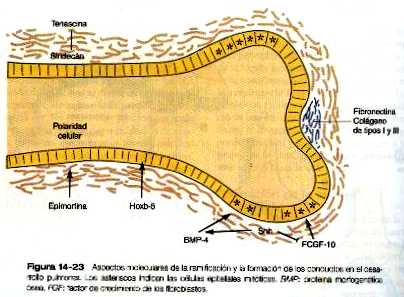
Los principios básicos que subyacen a la ramificación pulmonar son similares a los que operan en el desarrollo de las glándulas salivales y el páncreas. En algunos puntos de la ramificación se reduce la proliferación de las células epiteliales, y el depósito del colágeno de tipos I y III, fibronectina y proteoglicanos que estabilizan la morfología del punto de ramificación y de las regiones ductales más proximales. Por otra parte, el aumento de la proliferación de las células epiteliales caracteriza a las porciones de rápido crecimiento de las yemas epiteliales (figura 14_23).
El grupo más frecuente de malformaciones del tracto respiratorio se relaciona con la división anormal de la yema traqueal y el esófago al comienzo del desarrollo del sistema respiratorio. Existen muchas variedades anatómicas de las fístulas traqueoesofágicas (figura 14_26), pero prácticamente todas comprenden la estenosis o atresia de un segmento de la tráquea o del esófago y una conexión anómala entre ellos. Se manifiestan al poco tiempo del nacimiento por que el bebé se atraganta o regurgita la leche cuando lo alimentan.
Estas raras malformaciones son incompatibles con la vida. Es probable que su causa fundamental sea un defecto en la interacción epitelio-mesénquima, de las cuales depende la formación del sistema respiratorio.
MALFORMACIONES MACROSCÓPICAS DE LOS PULMONES
Debido a su complejidad estructural, los pulmones están sujetos a numerosas variaciones o malformaciones estructurales (por ejemplo, formación anómala de los lóbulos). Por lo general son asintomáticas pero pueden ser foco de infecciones crónicas. Es importante que el cirujano pulmonar reconozca la posibilidad de que haya una anomalía de este tipo.
SÍNDROME DE DIFICULTAD RESPIRATORIA
(ENFERMEDAD DE MEMBRANA HIALINA PULMONAR)
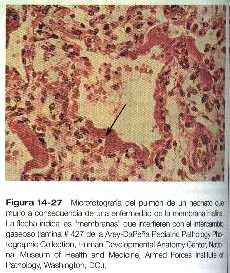
El síndrome de dificultad respiratoria suele manifestarse en los bebés prematuros y se caracteriza por una respiración dificultosa. En los bebés que mueren por esta afección los pulmones están poco inflados, y los alvéolos están parcialmente llenos de un fluido proteico que forma una membrana sobre las superficies respiratorias (figura 14_27), lo que es debido a la insuficiencia en la formación del surfactante por las células alveolares llamadas neumocitos de tipo II.
QUISTES CONGÉNITOS DEL PULMÓN
En el pulmón o en otras partes del tracto respiratorio, pueden formarse estructuras quísticas anómalas, que van desde quistes únicos de gran tamaño hasta numerosos quistes pequeños diseminados por todo el parénquima del pulmón. Pueden asociarse con riñones poliquísticos. Si los quistes son numerosos, pueden causar alteraciones respiratorias.
La actividad de muchas moléculas contribuye a la morfogénesis del pulmón, pero todavía no se ha establecido una jerarquía verdaderamente coherente en el control molecular. Varios factores de crecimiento, entre los cuales se cuenta el factor 10 de crecimiento de los fibroblastos (FCF-10) producido por el mesénquima local (mesodermo esplácnico) estarían implicados en la estimulación del crecimiento inicial de las yemas endodérmicas del pulmón en desarrollo. Las puntas de las yemas Epiteliales producen sonic hedgehog que parece inducir la expresión de la proteína 4 morfogenética ósea (BMP-4) Y reprimir al FGF-10 en la mesénquima circundante. En su área mas distal, la lámina basal que rodea al epitelio pulmonar es porosa, lo que permite que el contacto directo entre las prolongaciones de las células epiteliales y mesenquimatosas.
El protoncogén N-myc también está implicado en la ramificación porque si se inhibe su expresión, aquella no se produce. La actividad de este protoncogén se inhibe con altas concentraciones de factor beta 1 de transformación de crecimiento, y la elevación de este factor a lo largo de las paredes de los bronquios ya formados puede explicar, en parte, la ausencia de ramificación en estas áreas. La presencia concomitante del proteoglicano asociado con las células epiteliales – el sindecán - es importante para mantener la estabilidad de las láminas basales epiteliales a lo largo de los conductos; se encuentra a lo largo de los conductos ya formados interactuando con la proteína de la matriz extracelular, tenascina, pero no en áreas donde está teniendo lugar la ramificación como en las regiones saculares terminales de la vía aérea en desarrollo.
Al igual que con la morfogénesis de la ramificación, la formación y el mantenimiento de los conductos tapizados de epitelio comprende un conjunto especial de componentes moleculares. El hoxb-5 se expresa al comienzo del desarrollo de los bronquíolos más pequeños (por ejemplo, los terminales) pero no en los componentes del pulmón que participarán en el intercambio gaseoso efectivo (es decir, bronquíolos, conductos alveolares, alvéolos). La proteína, epimorfina es importante en la formación posterior de los tubos epiteliales; se localiza en el mesénquima y al parecer emite una señal que permite que las células epiteliales suprayacentes establezcan una polaridad apropiada u ordenamientos celulares. En el pulmón embrionario se si bloquea la epimorfina con anticuerpos específicos los conductos epiteliales en desarrollo se desorganizan y no forman luces.
ETAPAS DEL DESARROLLO PULMONAR
Etapa embrionaria (semanas 4 a 7)
La etapa embrionaria abarca desde la formación inicial del divertículo respiratorio hasta la de los principales segmentos broncopulmonares. Durante este periodo, los pulmones en desarrollo crecen hacia el interior de las cavidades pleurales bilaterales y comienzan a llenarlas; éstas representan los principales componentes de la cavidad torácica del cuerpo por encima del pericardio.
Etapa canalicular (semanas 17 a 26)
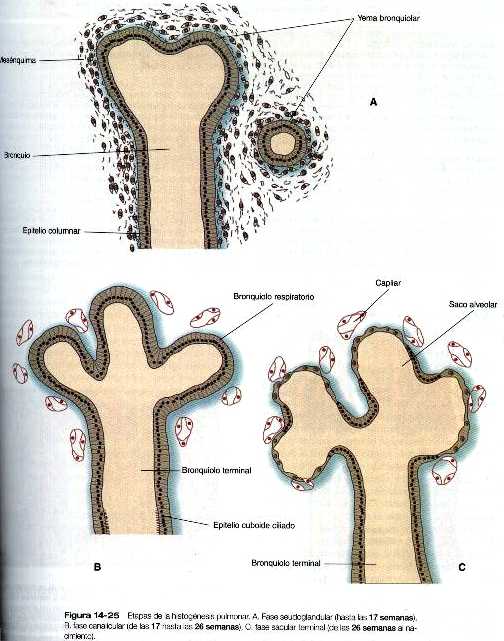
La etapa canalicular se caracteriza por la formación de los bronquíolos respiratorios como resultado de la gemación de los componentes terminales del sistema de bronquíolos que se formaron durante la etapa pseudo glandular. El otro hecho importante que se presenta durante esta etapa es el notable crecimiento de los vasos sanguíneos hacia el interior de los pulmones en desarrollo y la estrecha asociación de los capilares con las paredes de los bronquíolos respiratorios (figura 14_25). En ocasiones, un feto que nace hacia el final de este periodo puede sobrevivir con cuidados intensivos, pero la madurez pulmonar es la principal razón de la falta de viabilidad.
Etapa terminal de saculación (semana 26 hasta el nacimiento)
Durante la etapa final de saculación, los sacos aéreos terminales (alvéolos) procedentes de los bronquíolos respiratorios que se formaron mayoritariamente durante la etapa canalicular. El revestimiento epitelial de los alvéolos se diferencia en dos tipos de células: las células alveolares del tipo I (neumocitos), a través de las cuales tiene lugar el intercambio gaseoso después del nacimiento, y las células epiteliales secretorias de tipo II. Estas últimas forman el surfactante pulmonar, material que se extiende sobre la superficie de los alvéolos para reducir la tensión superficial y facilitar la expansión de los alvéolos durante la respiración. La investigación con marcadores específicos de las células epiteliales ha demostrado que las células de tipo II son las primeras que se forman en el revestimiento epitelial. Después de la proliferación, algunos de estos neumocitos de tipo II se aplanan, pierden sus funciones secretorias características y experimentan diferenciación terminal en neumocitos tipo I. Otras células de este último tipo pueden diferenciarse directamente a partir de una reserva de células epiteliales precursoras que existen en el revestimiento alveolar primitivo. Con el aumento en la producción de surfactante pulmonar, el feto tiene una posibilidad mayor de supervivencia si nace prematuro. En el feto, las vías aéreas intrapulmonares están llenas de líquido (capítulo 17). Durante las últimas 4 semanas del embarazo el gran incremento en la formación de alvéolos da como resultado un aumento exponencial en la superficie respiratoria del pulmón. Estas semanas suelen denominarse periodo alveolar del desarrollo pulmonar.
Etapa postnatal
Al nacimiento el pulmón de los mamíferos todavía no está maduro. Después del nacimiento, se forma alrededor del 90% de los cerca de 300 millones de alvéolos que se encuentran en el pulmón humano maduro. El principal mecanismo de este aumento es la formación de tabiques secundarios de tejido conectivo que dividen los sacos alveolares existentes. Cuando hacen su aparición, los tabiques secundarios son relativamente gruesos. A su debido tiempo, se transforman en los tabiques maduros más delgados capaces de ejercer completamente la función del intercambio respiratorio.
CAVIDADES CORPORALES
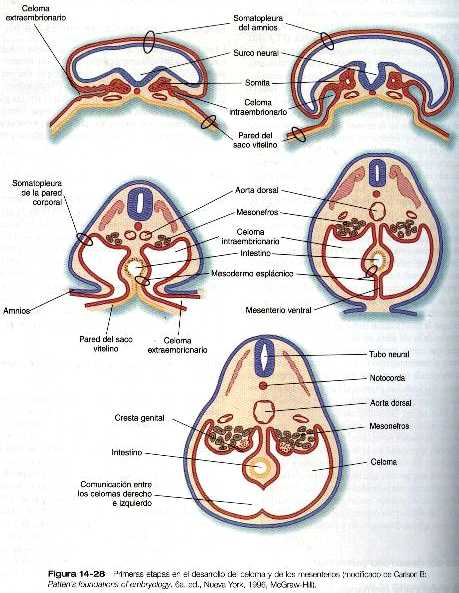
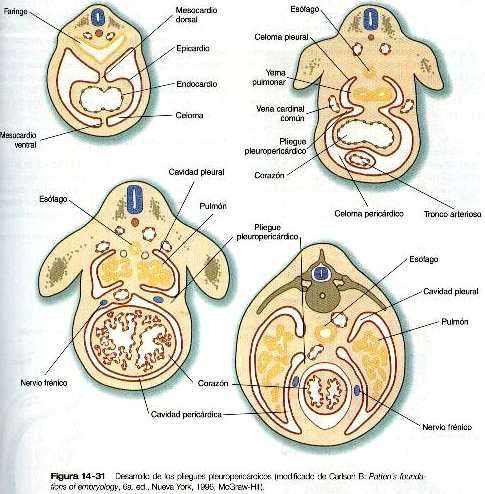
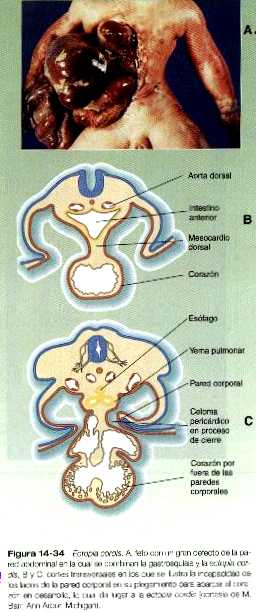
Conforme el mesodermo lateral del embrión primitivo se divide y luego se pliega hacia los lados, el espacio que queda entre las capas somática y esplácnica del mesodermo se convierte en el celoma intraembrionario común (figura 14_28), véase el capítulo 5 para más información. El mismo proceso de plegamiento que da como resultado la formación de la pared ventral del cuerpo y la separación del celoma intraembrionario del extraembrionario también lleva a las dos capas del mesodermo esplácnico a situarse alrededor del intestino recién formado por mesenterio primario o común. El mesenterio primario fija al intestino a la pared dorsal del cuerpo formando el mesenterio dorsal y lo ancla a la pared ventral del cuerpo a través del mesenterio ventral. Esta organización divide la cavidad celómica en dos compartimientos, izquierdo y derecho. No obstante, muy pronto la mayor parte del mesenterio ventral se rompe, y al hacerlo, causa una comunicación entre las mitades derecha e izquierda del celoma. En la región del estómago y el hígado en desarrollo el mesenterio ventral persiste, como mesogastrio ventral y ligamento falciforme del hígado (figura 14_3). En dirección más craneal, el primordio tubular del corazón es fijado de forma similar por un mesocardio dorsal y durante un breve tiempo por un mesocardio ventral que pronto se rompe.
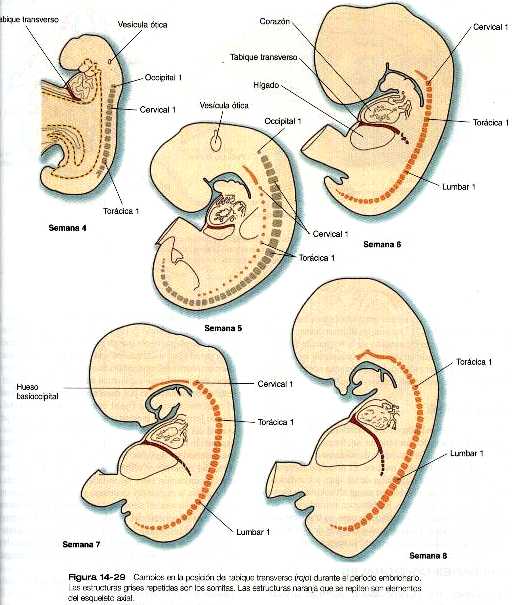
Uno de los elementos más importantes en la división del celoma común en los componentes torácico y abdominal es el tabique transverso (septum transversum). Crece a partir de la pared ventral del cuerpo como un reborde semicircular que separa el corazón del hígado en desarrollo (figura 14_29). Durante el comienzo de su desarrollo, una gran porción del hígado está incluida en el tabique transverso. Al final de su desarrollo, este tabique será un componente importante del diafragma.
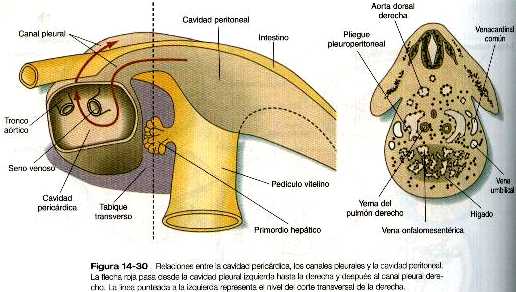
El tabique transverso en crecimiento sirve como separación parcial entre las porciones pericárdica y peritoneal del celoma. En el momento en que el borde libre en crecimiento del tabique alcanza el suelo del intestino anterior, prácticamente ha dividido el celoma común en dos, sin embargo, ambas partes están conectadas por los canales pleurales pericardioperitoneales. Se trata de dos cortos canales localizados a cada lado del intestino anterior (figura 14_30), que representan los espacios dentro de los cuales se desarrollarán los pulmones. Conforme los pulmones aumentan de tamaño, estas estructuras se agrandan y acaban por convertirse en las cavidades pleurales.
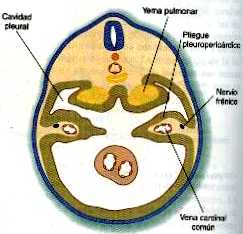
Los canales pleurales están delimitados en parte por dos pares de pliegues de tejido. Los pliegues pleuropericárdicos y los pleuroperitoneales. Los pliegues pleuropericárdicos (figura 14_24) son crestas de tejido asociadas con las venas cardinales comunes, que abultan hacia la pared dorsolateral del celoma a medida que se incurvan hacia la línea media de la porción torácica del celoma para entrar en el seno venoso del corazón (figura 14_31). En un comienzo, los pliegues pleuropericárdicos no son grandes, y sólo ocasionan un estrechamiento en la unión de la cavidad pericárdica y los canales pleurales forman rebordes prominentes que se fusionan en la línea media, dando lugar a la capa fibrosa (parietal) del pericardio.
En el espesor de los pliegues pleuropericárdicos se encuentra el par de nervios frénicos. Surgen de la unión de las ramas de las raíces cervicales 3, 4 y 5 e inervan las fibras musculares del diafragma. Con los cambios de posición de varios componentes del cuerpo durante el crecimiento, el diafragma terminal por descender hasta el nivel de las vértebras torácicas inferiores. A medida que lo hace, se lleva a los nervios frénicos con él. Incluso, en los adultos el trayecto de los nervios frénicos a través del pericardio fibroso recuerda su asociación inicial con los pliegues pleuropericárdicos.
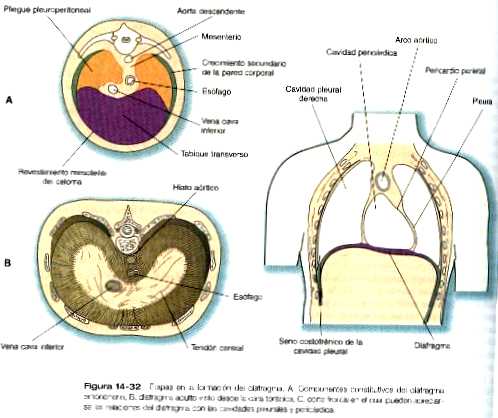
En los extremos caudales de los canales pleurales, otro par de pliegues, los pleuroperitoneales, se vuelven prominentes conforme los pulmones en su expansión empujan hacia el mesodermo de la pared corporal. Los pliegues pleuroperitoneales ocupan progresivamente porciones cada vez mayores del canal pleural hasta que se fusionan con el tabique transverso y el mesenterio del esófago, con lo que se produce la obliteración efectiva del canal pleural (figura 14_32). Así se eliminan todas las conexiones existentes entre la cavidad abdominal y la torácica.
Formación del diafragma
El diafragma, que separa la cavidad torácica de la abdominal en el adulto, es una estructura compleja y derivada de varia estructuras embrionarias (figura 14_32). El gran componente ventral del diafragma procede del tabique transverso, que se fusiona con la parte ventral del mesenterio esofágico.
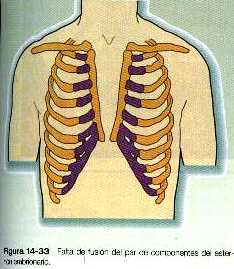
En ocasiones, los lados opuestos de la pared del cuerpo no se fusionan conforme el embrión adopta la forma cilíndrica al final del primer mes. Varios mecanismos anómalos como la hipoplasia tisular pueden explicar estos defectos. Un defecto menor desde el punto de vista cuantitativo en el cierre de la pared del tórax se manifiesta como la ausencia de fusión esternal (figura 14_33). Si el crecimiento de los dos lados de la pared del tórax es muy defectuoso, es probable que el corazón se forme fuera de la cavidad torácica, lo que da lugar a la ectopia cordis (exocardia) (figura 14_34).
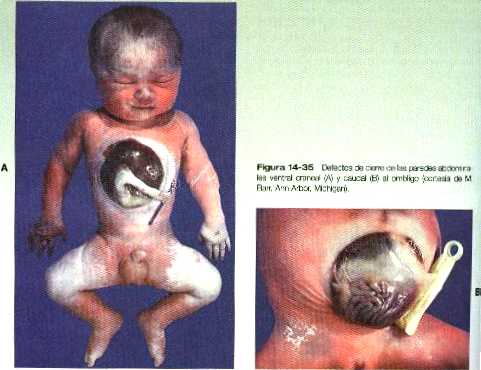
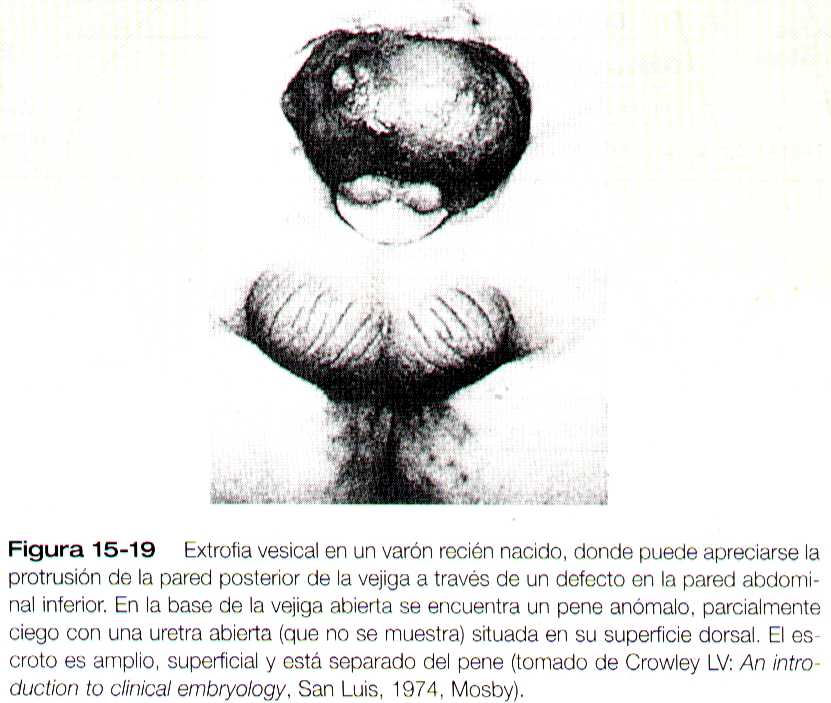
Los defectos de cierre de la pared abdominal pueden conducir a malformaciones macroscópicas similares. En muchos casos de onfalocele (figura 14_12) se hacen evidentes la hipoplasia de la pared abdominal. Los casos mas graves comprenden la hernia del contenido abdominal a través de una fisura situada entre el ombligo y el esternón (gastrosquisis) como se ve en la figura 14_35. En situación caudal al ombligo puede encontrarse la extrofia vesical, un defecto de cierre asociado con la vejiga (figura 15_19).
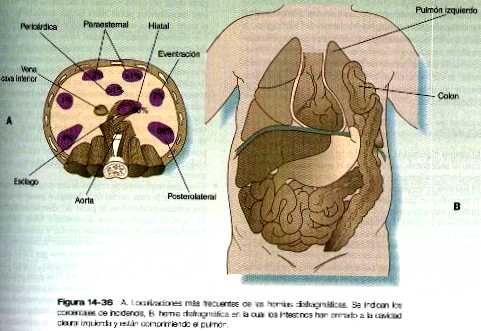
HERNIAS DIAFRAGMÁTICAS
La fusión incompleta o la hipoplasia de uno de los componentes del diafragma puede llevar a la existencia de una comunicación entre las cavidades abdominal y torácica. Si el defecto es de tamaño suficiente, varias estructuras de la cavidad abdominal (por lo general parte del estómago o los intestinos) pueden herniarse hacia la cavidad torácica, o con menor frecuencia las estructuras torácicas pueden penetrar en la cavidad abdominal.
Los casos de hernia menor pueden causar síntomas digestivos. En el caso de los defectos grandes, la hernia masiva de los intestinos puede presionar contra el corazón o los pulmones y entorpecer su función. En la figura 14_36 se muestran algunos de los lugares frecuentes de defectos diafragmáticos.
CASO CLÍNICO
Una niña de 14 años había sufrido molestias caracterizadas por dolor abdominal alto de varios años de evolución que se repetían a intervalos más o menos regulares de cerca de un mes. Después de pasar por varios profesionales e instituciones sin encontrar alivio, la derivaron a un psiquiatra, que tampoco pudo resolver el caso. Por fin, un médico experto sospechó que sus síntomas podrían estar causados por una anomalía congénita. Otras pruebas diagnósticas y una intervención quirúrgica probaron que su sospecha era correcta.
¿Cuál fue el diagnóstico?
Los pliegues pleuroperitoneales convergen sobre el mesenterio esofágico procedentes de las paredes dorsolaterales. Estos componentes constituyen el cuerpo del diafragma. A medida que los pulmones siguen creciendo, sus vértices causales abren espacios adicionales en la pared corporal. El mesénquima de ésta, separado de la pared corporal propiamente dicha, se convierte en un tercer componente del diafragma al formar un delgado anillo de tejido a lo largo de sus bordes dorsolaterales.
a) Atresia anal
b) Divertículo de Meckel
c) Onfalocele
d) Vólvulo
e) Megacolon agangliónico
a) Ileon
b) Colon ascendente
c) Yeyuno
d) Colon transverso
e) Duodeno
a) Páncreas dorsal
b) Pulmón
c) Hígado
d) Timo
e) Bazo
a) Celíaca
b) Umbilical
c) Mesentérica superior
d) Aorta
e) Mesentérica inferior
a) El hígado
b) La notocorda
c) La yema pulmonar
d) El saco vitelino
e) Ninguno de los anteriores
a) Dientes
b) Tráquea
c) Hígado
d) Pulmones
e) Tráquea
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}