Capítulo 13
CABEZA Y CUELLO
DESARROLLO INICIAL DE LA CABEZA Y EL CUELLO
Componentes tisulares y segmentación del primordio de la región craneofacial
Primeras migraciones celulares y desplazamientos titulares en la región craneofacial
ORGANIZACIÓN FUNDAMENTAL DE LA REGIÓN BRANQUIAL (FARÍNGEA)
DESARROLLO DE LA REGIÓN FACIAL
Formación de la cara y de la región mandibular
Formación de la nariz y del aparato olfatorio
Formación de las glándulas salivales
Etapas del desarrollo de los dientes
Interacciones tisulares en el desarrollo del diente
Formación de la dentina y el esmalte
Erupción y sustitución de los dientes
DESARROLLO DE LA FARINGE Y SUS DERIVADOS
Desarrollo externo de la región faríngea
Estructuras de la línea media que surgen de la faringe
Timo y órganos linfoides
PREGUNTAS DE REPASO
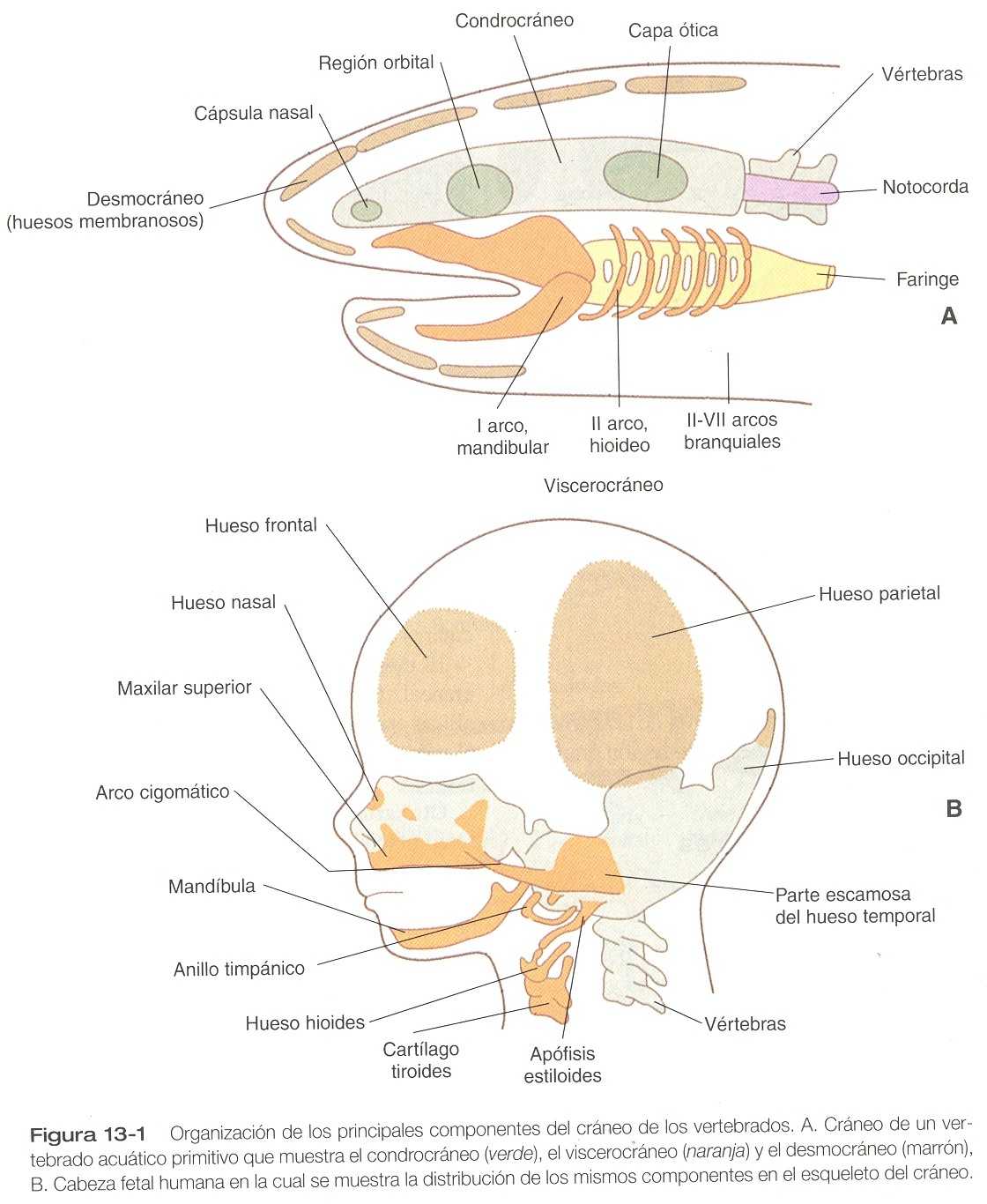
La región craneal de los primeros vertebrados constaba de dos componentes principales: 1) un condrocráneo asociado con el cerebro y los principales órganos de los sentidos (nariz, ojo, oído) y 2) un viscerocráneo, una serie de arcos branquiales (faríngeos) asociados con la región oral y la faringe (figura 13_1 A). Conforme los vertebrados fueron haciéndose más y más complejos, las contribuciones de la cresta neural a la cabeza se volvieron mucho más importantes, y se añadieron a la cara y a muchos huesos dérmicos (de formación intramembranosa) del cráneo (desmocráneo).
Con la evolución de la cara primitiva, los arcos branquiales más anteriores se transformaron para constituir las mandíbulas superior e inferior así como dos de los huesos del oído medio: el martillo y el yunque. A la par con el aumento en la complejidad de la cara (figura 13_1B) se produce un aumento correspondiente en la complejidad del prosencéfalo (compuesto por telencéfalo y diencéfalo). Desde los puntos de vista estructural y molecular, la parte rostral de la cabeza muestra características que la diferencian de la región faríngea, como son:
En capítulos anteriores se ha descrito con detalle el desarrollo de algunos de los componentes de la cabeza (por ejemplo, el sistema nervioso, la cresta neural y los huesos del cráneo) la primera parte de este capitulo ofrece una visión integrada del desarrollo craneofacial primario para mostrar las interrelaciones de los principales componentes. El resto del capitulo se centra en el desarrollo de la cara, la faringe y el sistema de los arcos faringeos. En las correlaciones clínicas 13_1 y 13_2a, b y c) al final del capitulo, se presentan malformaciones asociadas con la cabeza y el cuello.
DESARROLLO INICIAL DE LA CABEZA Y EL CUELLO
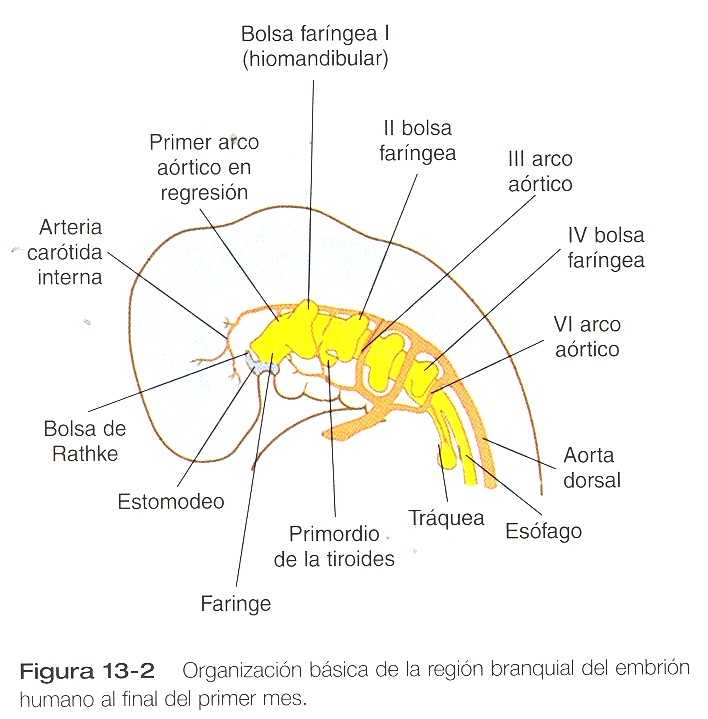
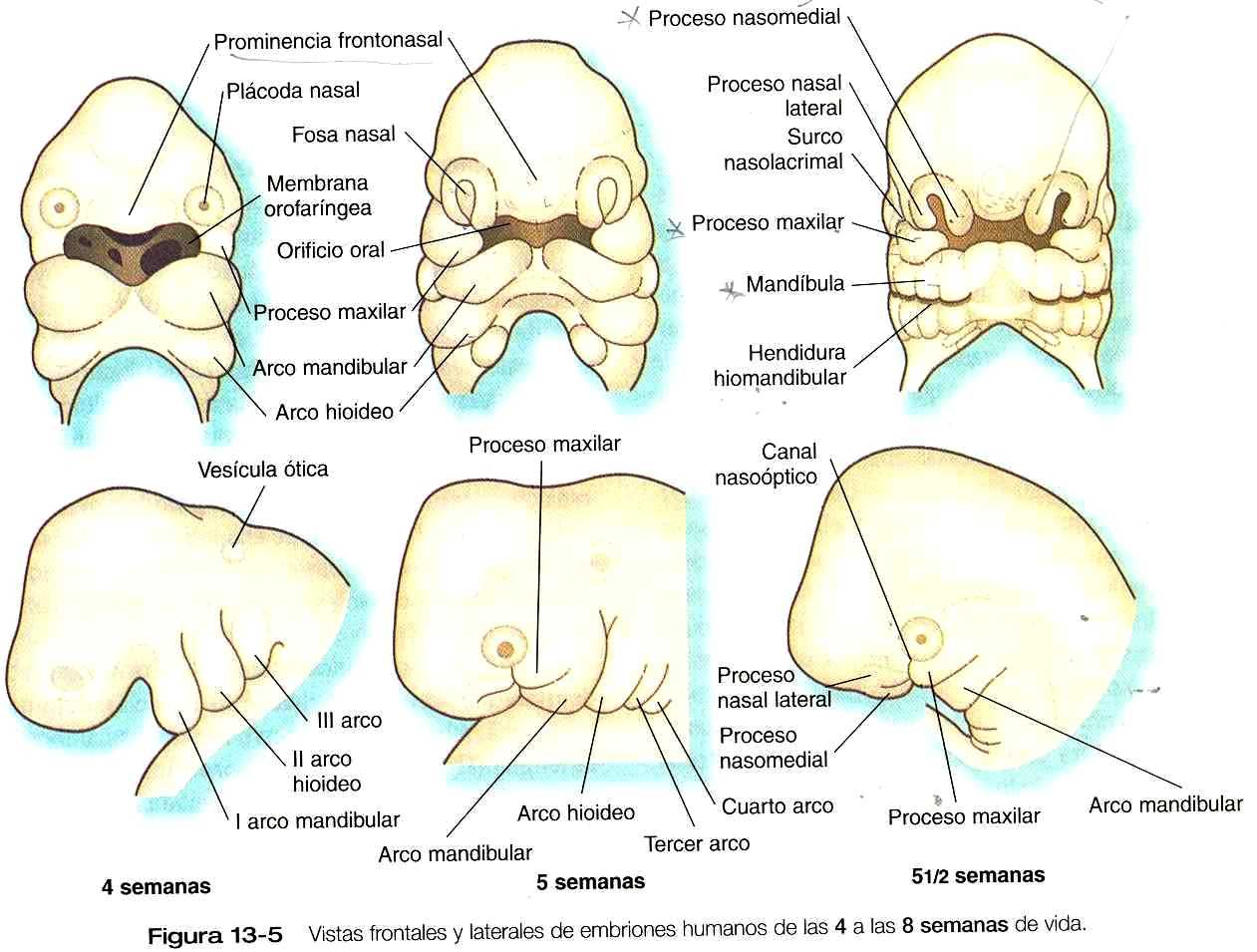
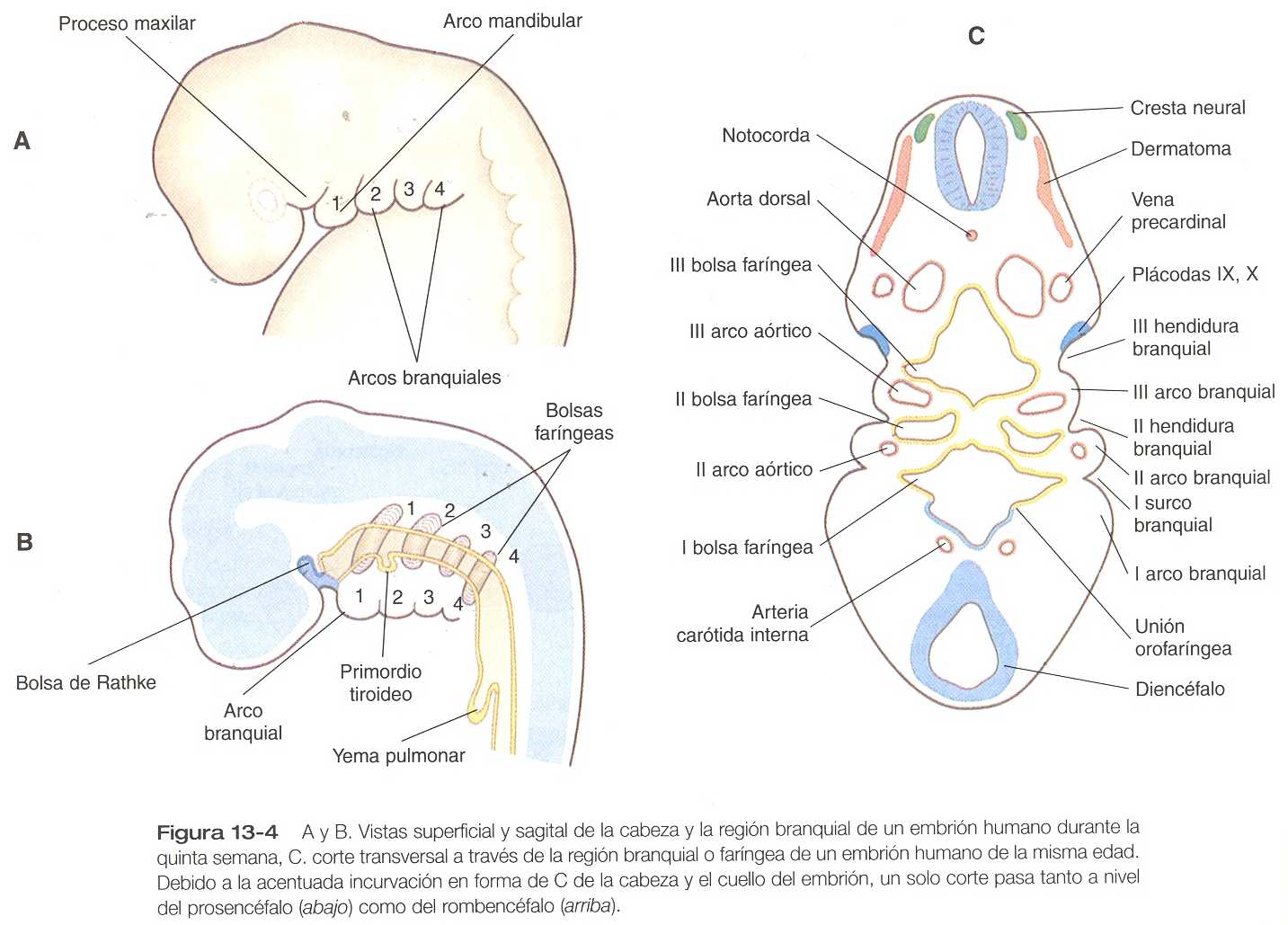
El desarrollo de la cabeza y el cuello comienza pronto en la vida embrionaria y continúa hasta la terminación del crecimiento posnatal al final de la adolescencia. La cefalización comienza con el rápido crecimiento del extremo rostral de la placa neural. Muy pronto, el futuro cerebro se convierte en el componente dominante de la región craneofacial. Por debajo del cerebro, la cara –que solo adquiere su forma tardíamente durante la embriogénesis- esta representada por el estomodeo (figura 13_2). En el embrión joven el estomodeo está separado del intestino primitivo por la membrana orofaríngea, que se rompe hacia el final del primer mes de vida embrionaria (figura 13_5). En la línea mediorrostral se encuentra la prominencia frontonasal, poblada con células mesenquimatosas derivadas del prosencéfalo y algo de la cresta neural del mesencéfalo. A cada lado de esta eminencia las plácodas ectodérmicas nasales aparecen como estructuras con forma de herradura; en cada una de ellas existe un proceso nasal lateral, derivado de la cresta neural del mesencéfalo. En dirección más caudal, el estomodeo está limitado por los procesos maxilar y mandibular, que están llenos de mesénquima derivado de la cresta neural que se origina de los dos primeros rombómeros.
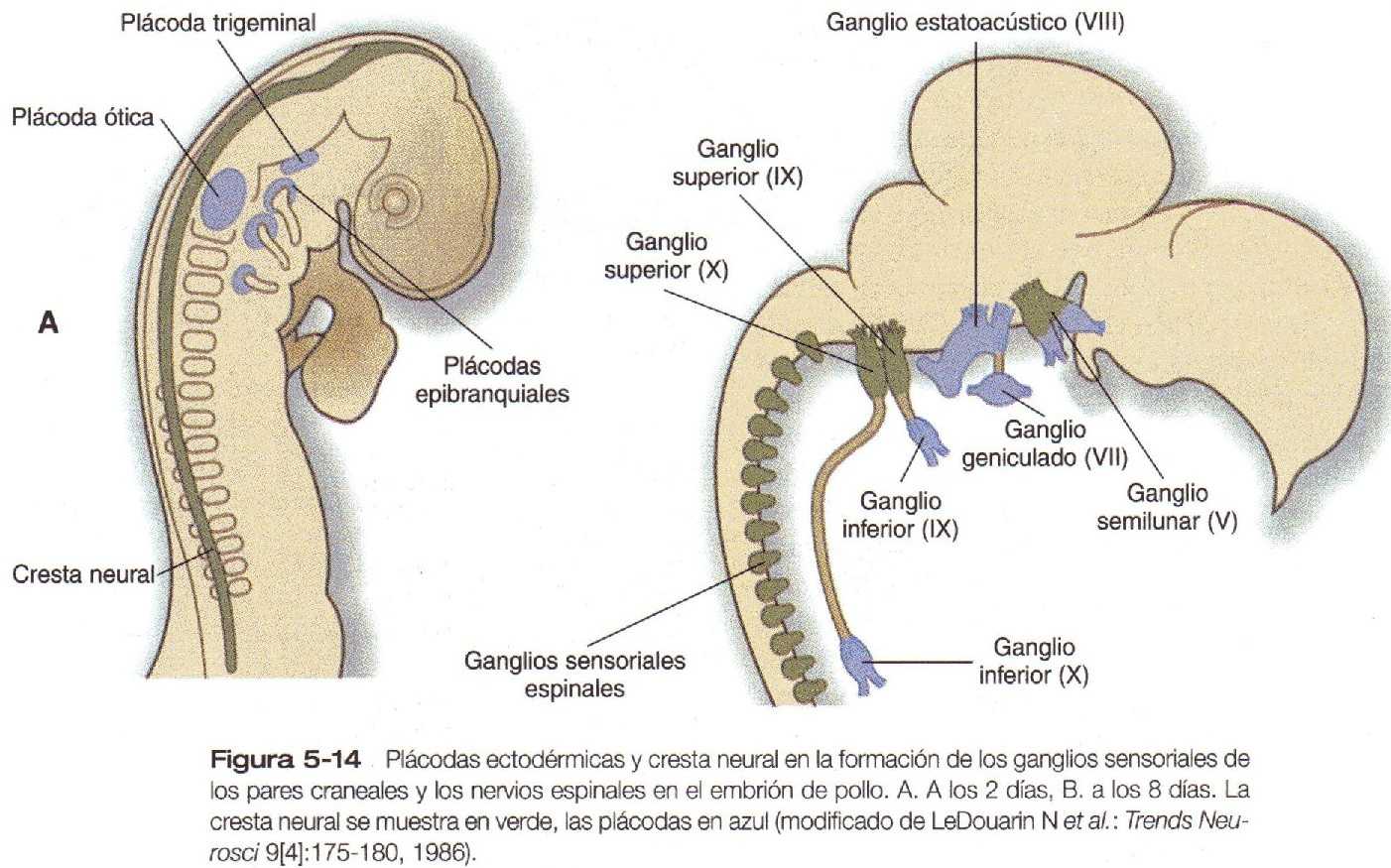
La futura región cervical está formada por las estructuras branquiales, conformadas por una serie de bolsas, arcos y hendiduras faríngeas o branquiales. Muchos componentes de la cara, los oídos y las glándulas de la cabeza y el cuello proceden de la región branquial. Otras estructuras evidentes son los pares de plácodas ectodérmicas (figura 5_14) de los cuales derivará gran parte del tejido sensorial de la región craneal.
Componentes tisulares y segmentación del primordio de la región craneofacial
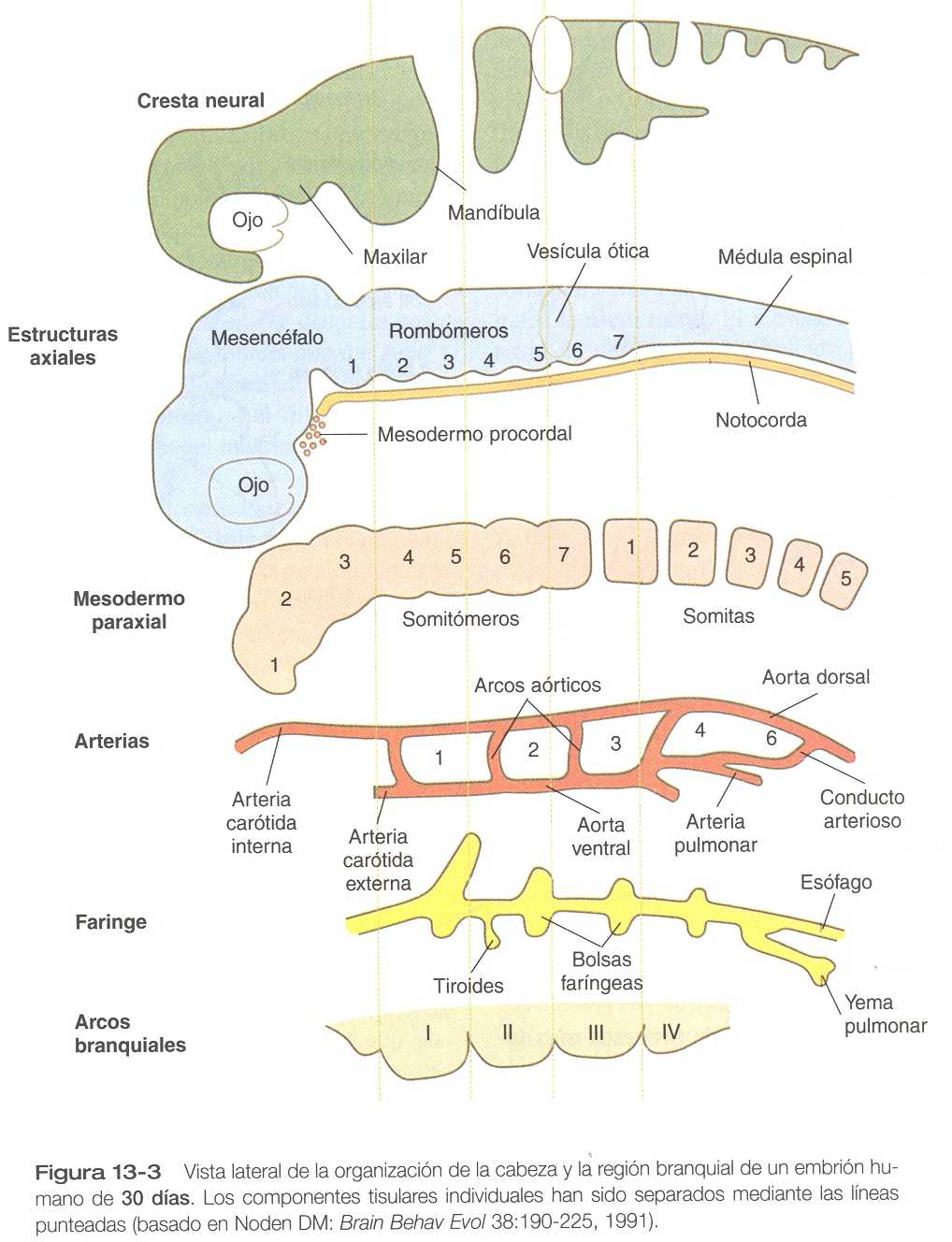
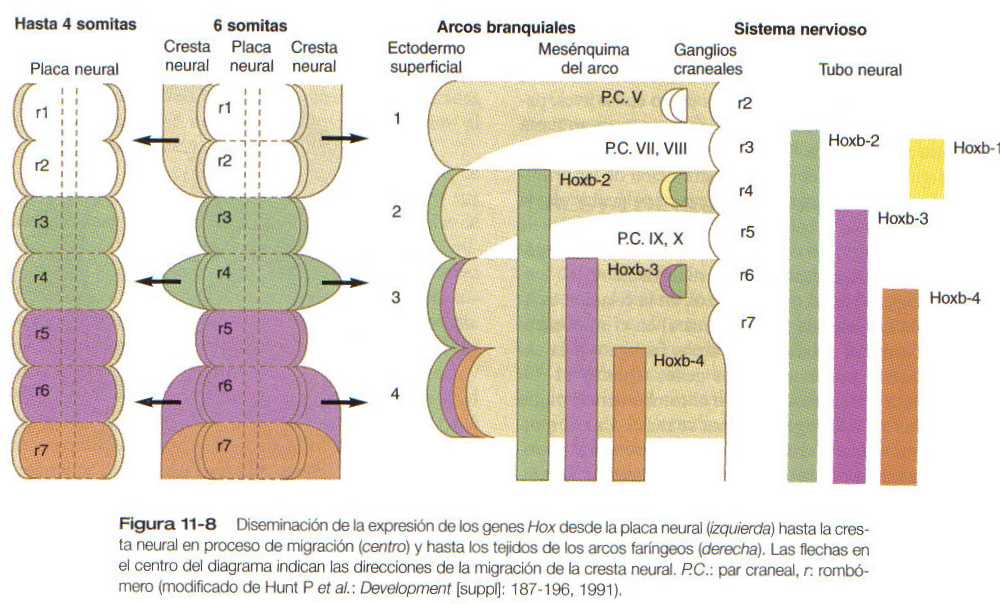
El primordio de la región craneofacial está formado por un tubo neural muy desarrollado, por debajo del cual se encuentran la notocorda y la faringe en situación ventral (figura 13-2). La faringe está rodeada por una serie de arcos branquiales o faríngeos. Muchos de los componentes titulares de la cabeza y el cuello tienen un origen y organización segmentaria. La figura 13-3 ilustra la segmentación de los componentes titulares de la cabeza. Como ya se explicó en capítulos anteriores, la segmentación morfológica de algunos tejidos craneales, en especial del sistema nervioso central (figura 10-10) está asociada con patrones definidos de expresión de ciertos genes portadores de homeobox. La cadena de acontecimientos que se suceden entre los patrones segmentarios de expresión genética y la aparición de la segmentación morfológica en algunas regiones de la región craneal sólo se conoce parcialmente.
Primeras migraciones celulares y desplazamientos titulares en la región craneofacial
El desarrollo craneofacial inicial se caracteriza por varias migraciones y desplazamientos masivos de células y tejidos. La cresta neural es el primer tejido que muestra comportamiento migratorio importante; sus células migran desde el sistema nervioso aun antes del cierre de la región craneal del tubo neural (capitulo 11). Inicialmente, se agregan grupos segmentarios de células de la cresta neural, en especial en la región branquial (figura 13-3). No obstante, estas poblaciones de células confluyen durante su migración. Por ejemplo, cuando se reemplazaron las células presuntivas de la cresta neural del segundo y el tercero arcos branquiales de embriones de aves por células de la cresta neural del primer arco (en esencia todo el aparato de la mandíbula) en un nivel más caudal al de la mandíbula normal del embrión huésped.
El mesodermo craneal primitivo está compuesto principalmente por mesodermo paraxial y procordal (figura 13_3). Las células mesenquimatosas que se originan en el mesodermo paraxial forman el tejido conectivo y los elementos esqueléticos de la región inferior del cráneo y de la región dorsal del cuello. Las células biogénicas del mesodermo paraxial derivadas de los somitómeros realizan migraciones muy amplias para formar la masa de los músculos de la región craneal. Al igual que sus similares en el tronco y las extremidades, estas células biogénicas se integran con el tejido conectivo para formar los músculos. Otro parecido con la musculatura del tronco es que el control morfogenético parece residir en los elementos del tejido conectivo de los músculos no en las células biogénicas mismas. En la cara y la faringe ventral este tejido conectivo procede de la cresta neural.
El mesodermo procordal, que emite importantes señales inductivas prosencéfalicas en el embrión primitivo, es una masa transitoria de células localizada en la línea media, en situación rostral al vértice de la notocorda. Aunque aún no hay consenso acerca del destino de éstas células, algunos investigadores creen que los mioblastos que contribuyen a la formación de los músculos extrínsecos del ojo derivan de ellas. En su trayecto hacia el ojo, las células del mesodermo procordal pueden pasar por los somitas más rostrales. Otros investigadores opinan que el origen de los músculos extraoculares es puramente somitomérico.
El mesodermo lateral no está bien definido en la región craneal. Los experimentos de injerto han demostrado que allí se originan las células endoteliales y musculares lisas y, al menos en las aves, algunas porciones de los cartílagos laríngeos.
Otro grupo importante de desplazamientos tisulares en la región facial es la confluencia de las células derivadas de las plácodas ectodérmicas con la de la cresta neural para formar partes de los órganos de los sentidos y de los ganglios de algunos pares craneales (figura 5_14).
ORGANIZACIÓN FUNDAMENTAL DE LA REGIÓN BRANQUIAL (FARÍNGEA)
Puesto que muchos componentes de la cara se derivan de la región branquial, es muy importante comprender su organización básica. En el embrión de un mes de edad la parte faríngea del intestino anterior contiene cuatro pares de evaginaciones laterales tapizadas de endodermo llamadas bolsas faríngeas y un divertículo impar en la línea media, el primordio tiroideo (figura 13_4). Si se siguen los contornos del ectodermo que cubre la región branquial, se observa cómo existen unas invaginaciones llamadas hendiduras branquiales que casi hacen contacto con la extensión más lateral de las bolsas faríngeas (figura 13_4C).
Con las hendiduras y las bolsas faríngeas alternan unas masas pareadas de mesénquima que se llaman arcos branquiales (faríngeos). En el centro de cada uno de ellos se encuentra una arteria que constituye un arco aórtico, que se extiende desde la aorta ventral hasta la dorsal (véanse el capítulo 16 y la figura 13_2). El mesénquima de los arcos branquiales tiene un doble origen. El mesénquima del esbozo de la musculatura se origina del mesodermo, específicamente de los somitómeros. Gran parte del resto del mesénquima del arco faríngeo, en especial el de la región ventral, se deriva de la cresta neural, en tanto que el mesodermo hace contribuciones variables al mesénquima de la región dorsal del arco branquial. El inicio del desarrollo de los arcos branquiales está muy relacionado con la diseminación de la expresión de los productos de los genes de la familia Hoxb desde los rombómeros del tubo neural al mesénquima del arco branquial y en última instancia al ectodermo que lo cubre (figura 11_8).
DESARROLLO DE LA REGIÓN FACIAL
Formación de la cara y de la región mandibular
Las estructuras de la cara y de la región mandibular se originan de varios primordios que rodean a la depresión del estomodeo en el embrión humano de cuatro a cinco semanas de edad (figura 13_5). Estos primordios son una prominencia frontonasal única, un par de procesos nasomediales, que son estructuras pertenecientes al primordio olfatorio (nasal) en forma de herradura, un par de procesos maxilares y un par de prominencias mandibulares, ambos componentes de los primeros arcos branquiales. Hoy se sabe que los diferentes primordios que forman la cara primitiva tienen propiedades distintivas en cuanto a su desarrollo. Según los datos que poseemos, las células de la cresta neural, que constituyen la masa mesenquimatosa de estos primordios de la cara. La naturaleza de esta información y la forma como se usa todavía no se ha aclarado. El maxilar contiene células mesenquimatosas derivadas de la cresta neural del mesencéfalo y del rombencéfalo (rombómeros 1 y 2).
Como sucede con las yemas de las extremidades, el crecimiento de los primordios faciales depende de interacciones ectodermo-mesénquima, aunque el unto de la interacción no está marcado por un engrosamiento ectodérmico apical como en el caso de la yema de la extremidad. Los experimentos de recombinación en aves han demostrado que el mesénquima de los tres primordios faciales (la eminencia frontonasal y los primordios maxilar y mandibular) puede mantener la diferenciación de la cresta ectodérmica apical engrosada de la yema de la extremidad. El sonic hedgehog y el factor de crecimiento de los fibroblastos (FGF), que hace poco tiempo se identificaron en el ectodermo del vértice de los procesos frontonasal y maxilar actúan, respectivamente, como organizador morfogenético y como estímulo para el crecimiento del mesénquima de los primordios faciales. El gen portador del homebox Msx-1 se expresa en el mesénquima muy proliferativo del vértice del primordio facial. El parecido con la expresión del este gen en la zona de progreso de la extremidad sugiere que unos mecanismos similares actúan en el crecimiento de los primordios tanto de los miembros como de la cara. El ácido retinoico también tiene una gran participación en el desarrollo inicial de la cara; tanto su deficiencia como su exceso son responsables de impresionantes anomalías faciales. Por ejemplo, se sabe que en exceso inhibe el crecimiento de los procesos frontonasal y nasomediales. También se sabe que este ácido influye sobre la expresión de los 3-parálogos de los genes Hox, pero su posición exacta en las secuencias de la expresión genética implicada en el control del desarrollo facial todavía no está del todo clara.
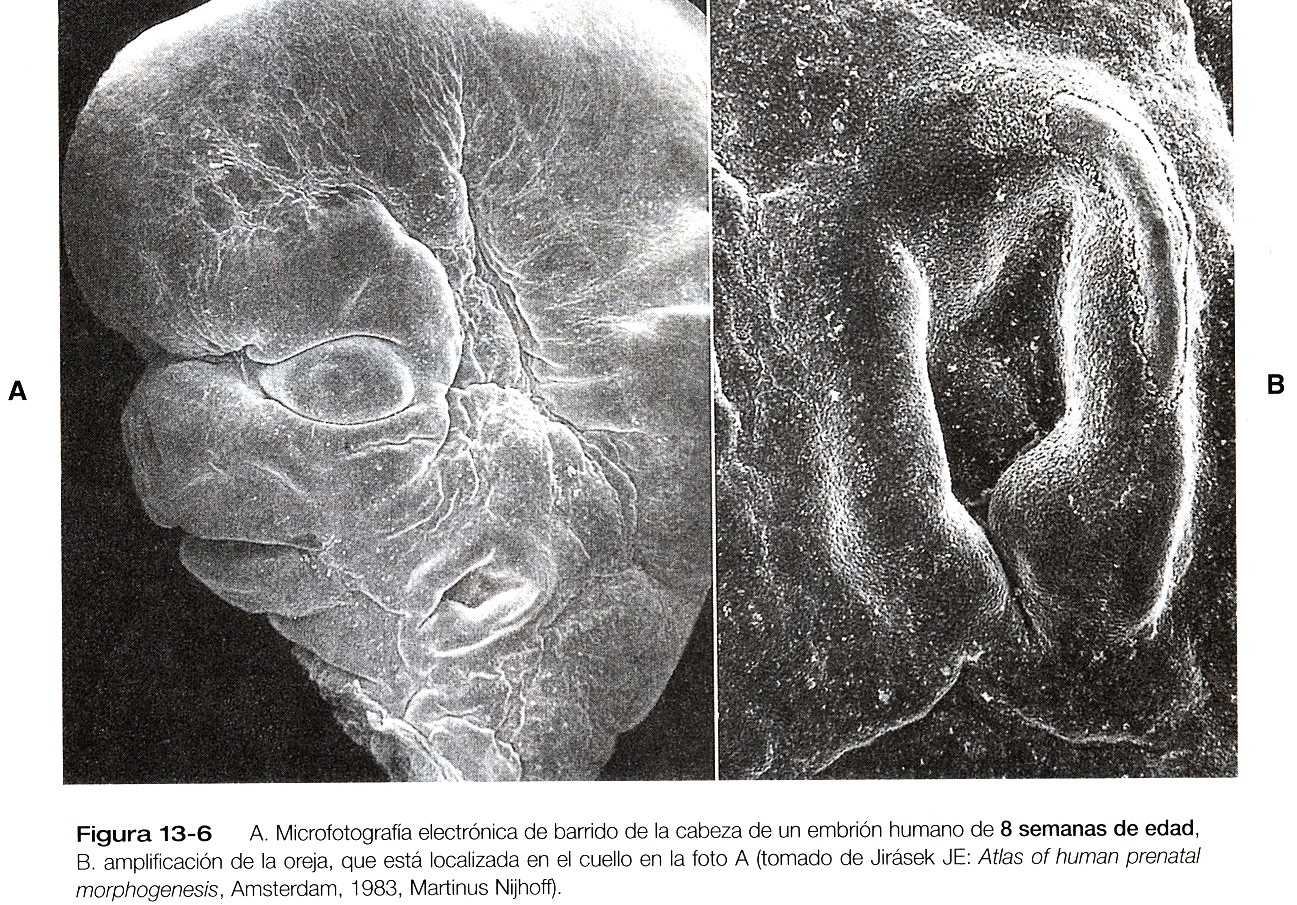
En virtud del crecimiento diferencial que se produce entre la cuarta semana y la octava (figura 13_5), los procesos nasomediales maxilares y mandibulares se hacen proporcionalmente más prominentes y terminan por fusionarse para formar el labio superior y la mandíbula (figura 13_6). A medida que esto va sucediendo y conforme los dos procesos nasomediales se fusionan, la prominencia frontonasal –que era una excrecencia de tejido que rodeaba el área del estomodeo en el embrión de cuatro a cinco semanas— se desplaza cranealmente y no contribuye a la formación del maxilar superior. Estos procesos una vez fusionados forman el segmento intermaxilar, que es un precursor de: 1) el filtro de la nariz, 2) el componente premaxilar del maxilar, y 3) el paladar primario (primitivo).
Entre el proceso maxilar y el proceso nasolateral del primordio nasal se encuentra un surco nasolacrimal (canal nasoóptico) que se extiende hasta el ojo en formación (figura 13_5). El ectodermo del suelo del surco nasolacrimal se engruesa para formar un cordón epitelial sólido, que se desprende del surco. Posteriormente, este cordón epitelial se canaliza y forma el conducto nasolacrimal y, cerca del ojo, el saco lacrimal. El conducto nasolacrimal se extiende desde la comisura interna del ojo hasta la cavidad nasal (meato inferior) y en la vida postnatal será la vía de drenaje de las lágrimas. Esta conexión explica por qué cuando las personas lloran les aumenta la secreción la nariz. Entre tanto, el proceso nasomedial en crecimiento se fusiona con el proceso maxilar, y en la región del surco nasolacrimal, el proceso nasolateral protruye y se fusiona con la zona superficial del proceso maxilar. La región de la fusión de los procesos nasomedial y maxilar está marcada por una sutura epitelial, llamada la aleta nasal. El mesénquima pronto invade la aleta nasal, lo que da como resultado una continuidad entre los procesos nasomedial y maxilar.
La formación de la mandíbula es más sencilla. Las eminencias mandibulares bilaterales se agrandan y sus componentes internos se fusionan en la línea media y así se forma el cuerpo de la mandíbula. El hoyuelo de la línea media que se observa en el mentón de algunos individuos es un reflejo de la variación en el grado de fusión de las prominencias mandibulares. En la mandíbula se diferencia una estructura cartilaginosa alargada llamada cartílago de Meckel (figura 13_22D). Éste, que es un derivado de las células de la cresta neural del primer arco faríngeo, forma el soporte alrededor del cual se disponen los huesos membranosos (que formarán el esqueleto definitivo de la mandíbula). Los estudios experimentales indican que la forma de bastón del cartílago de Meckel está relacionada con la inhibición de la condrogénesis en la zona posterior ejercida por el ectodermo que rodea al cartílago de Meckel, se desarrollan grandes masas de cartílago en vez de la formación de aspecto de bastón. Estas propiedades se parecen a las interacciones inhibitorias observadas entre el ectodermo y la condrogénesis en la yema de la extremidad.
Poco después de que las estructuras faciales básicas se han formado, son invadidas por células mesodérmicas derivadas del primero y el segundo arcos branquiales. Estas células forman los músculos masticadores (derivados del primer arco que son inervados por el V par craneal) y los músculos de la expresión facial no mímicos (derivados del segundo arco e inervados por el VII par).
Aunque la estructura básica de la cara se define entre la cuarta y la octava semanas, los cambios en la proporcionalidad de las regiones continúan hasta mucho después del nacimiento. En particular, la región media de la cara permanece menos desarrollada durante toda la embriogénesis y aun al comienzo de la vida posnatal.
Formación del paladar
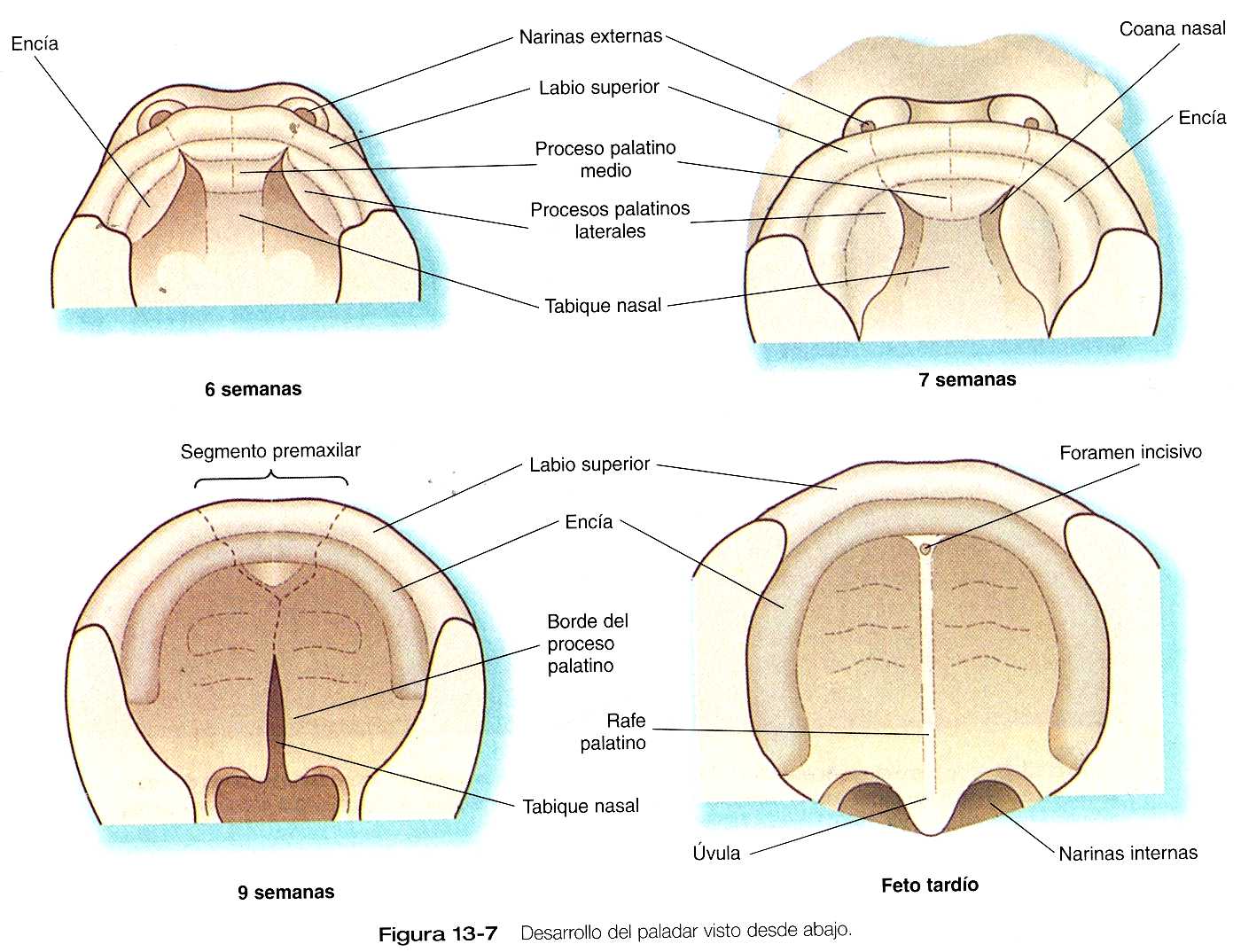
El embrión joven tiene una cavidad oronasal única, pero en los mamíferos el paladar se forma entre la sexta y la décima semanas y separa las cavidades nasal y oral. El paladar se deriva de tres primordios: un proceso palatino medio único y un par de procesos palatinos laterales (figura 13_7).
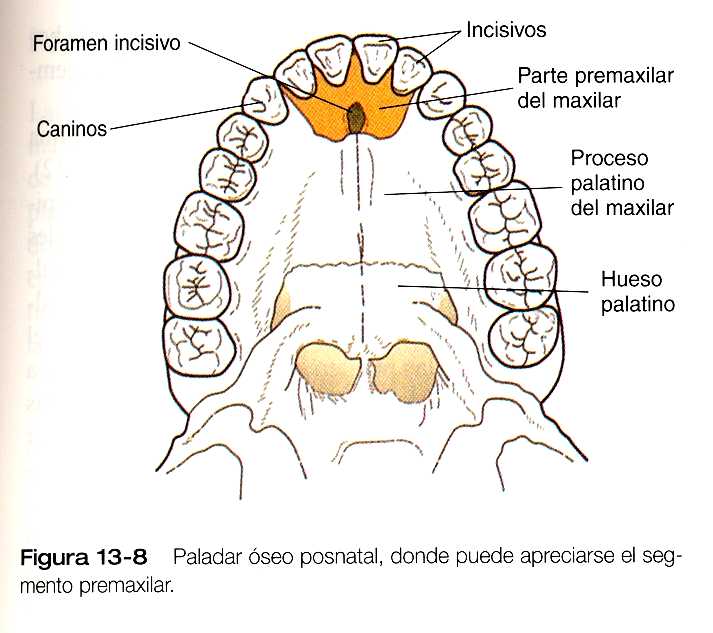
El proceso palatino medio es una invaginación de los procesos nasomediales recién fusionados. Conforme va creciendo, en el proceso palatino medio se va formando una estructura ósea triangular llamada paladar primario. En la vida posnatal el componente esquelético del paladar primario se denomina componente premaxilar del maxilar. Los cuatro dientes incisivos superiores provienen de esta estructura (figura 13_8).
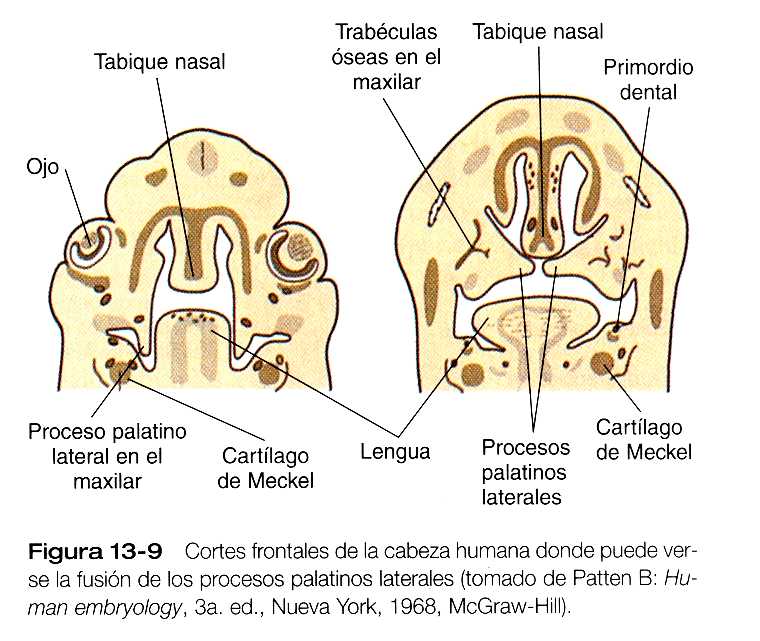
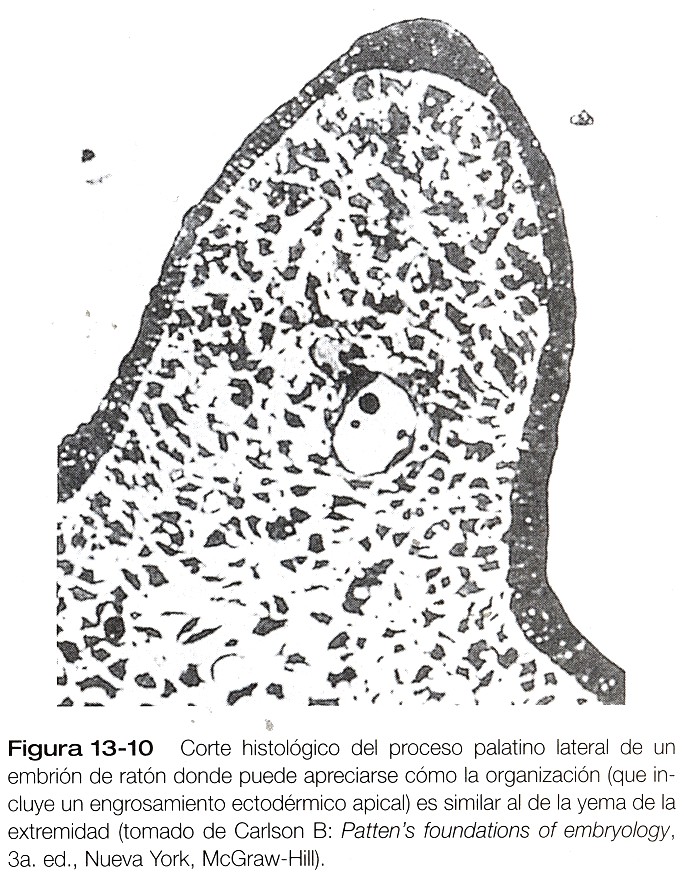
Los procesos palatinos laterales, que son los precursores del paladar secundario, hacen su aparición durante la sexta semana. Al principio crecen hacia abajo a ambos lados de la lengua (figura 13_9). En el borde apical de los procesos palatinos en crecimiento pueden verse en ocasiones engrosamientos ectodérmicos que recuerdan la cresta ectodérmica apical de la yema de la extremidad (figura 13_10). Al parecer, el crecimiento de los procesos palatinos laterales implica tanto interacciones de ectodermo y mesénquima como acciones de factores de crecimiento específicos en las células mesenquimatosas, como el factor de crecimiento epidérmico y el factor alfa de transformación del crecimiento. Durante la séptima semana, los procesos palatinos laterales modifican radicalmente la posición perpendicular a los procesos maxilares. Los vértices de estos procesos se encuentran en la línea media y comienzan a fusionarse.
A pesar de los muchos años de investigación que se han dedicado a este tema, el mecanismo que regula y procede la elevación de los procesos palatinos laterales no se ha aclarado. El edema en la matriz extracelular de los procesos palatinos les provoca una elasticidad que les permite aproximarse en el transcurso de los minutos y las horas que siguen a su cambio de posición a ambos lados de la lengua. Se ha especulado que el cambio de posición, que se produce durante un periodo en el cual la cabeza está creciendo en altura pero no en anchura, puede ser precipitado por el hipo del embrión. Cuando los procesos palatinos se elevan, por lo general aún no han tenido lugar ni la apertura de la mandíbula, ni la deglución ni la succión.
Otra estructura implicada en la formación del paladar es el tabique nasal (figuras 13_7 y 13_19). Esta estructura de la línea media, que es una prolongación descendiente de la prominencia frontonasal, alcanza el nivel de los procesos palatinos laterales en el momento en que éstos se unen para formar el paladar secundario definitivo. En dirección rostral, el tabique nasal se continúa con el paladar primario.
A nivel macroscópico, los procesos palatinos se fusionan en la línea media, pero en dirección rostral también se unen al paladar primario. El punto de fusión del paladar primario con los procesos palatinos laterales en la línea media está marcado por el foramen incisivo (figura 13_8).
Por su importancia clínica, la fusión de los procesos palatinos laterales ha sido objeto de intensa investigación. Cuando estos procesos establecen contacto por primera vez en la línea media, los cubre un epitelio homogéneo. Sin embargo, durante la fusión la sutura epitelial de la línea media desaparece. El epitelio de la superficie nasal del paladar se convierte en un epitelio columnar de tipo ciliado, en tanto que el de la cavidad oral se vuelve escamoso y estatificado. Algunas preguntas importantes desde el punto de vista del desarrollo son:
¿Qué causa la desaparición de la sutura de la línea media?
¿Qué señales dan como resultado las diversas vías de diferenciación del epitelio a ambos lados del paladar?
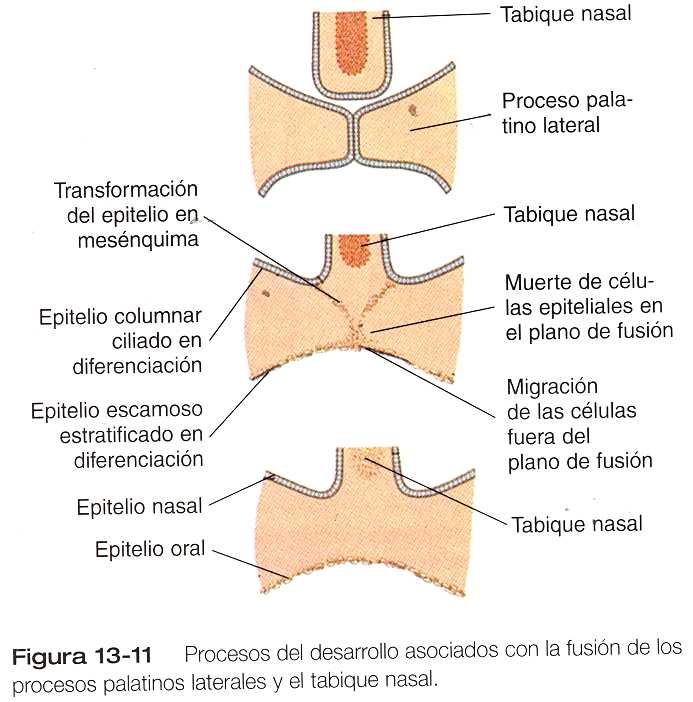
La desaparición de la sutura epitelial de la línea media después de la aproximación de los procesos palatinos laterales implica varios procesos fundamentales del desarrollo (figura 13_11). Algunas de las células epiteliales situadas en la zona de la sutura de fusión sufren apoptosis y desaparecen; otras experimentan una transformación del crecimiento. Otras pueden migrar fuera del plano de fusión y terminar localizándose en el epitelio que tapiza la cavidad oral.
Los experimentos de cultivo in Vitro en numerosas especies de un solo proceso palatino lateral han demostrado con claridad que todos los aspectos de la diferenciación epitelial (muerte celular en la línea media y vías de diferenciación diversas en las superficies oral y nasal) pueden tener lugar en ausencia de contacto con el proceso palatino del lado opuesto. Estas distintas vías de diferenciación no son intrínsecas del epitelio regional sino que son mediadas por el mesénquima subyacente produce factores de crecimiento que influyen sobre la producción y la distribución regional de las moléculas de la matriz extracelular (por ejemplo, el colágeno de tipo IX). Se desconoce la forma en que las células epiteliales reciben e interpretan estas señales.
Formación de la nariz y del aparato olfatorio
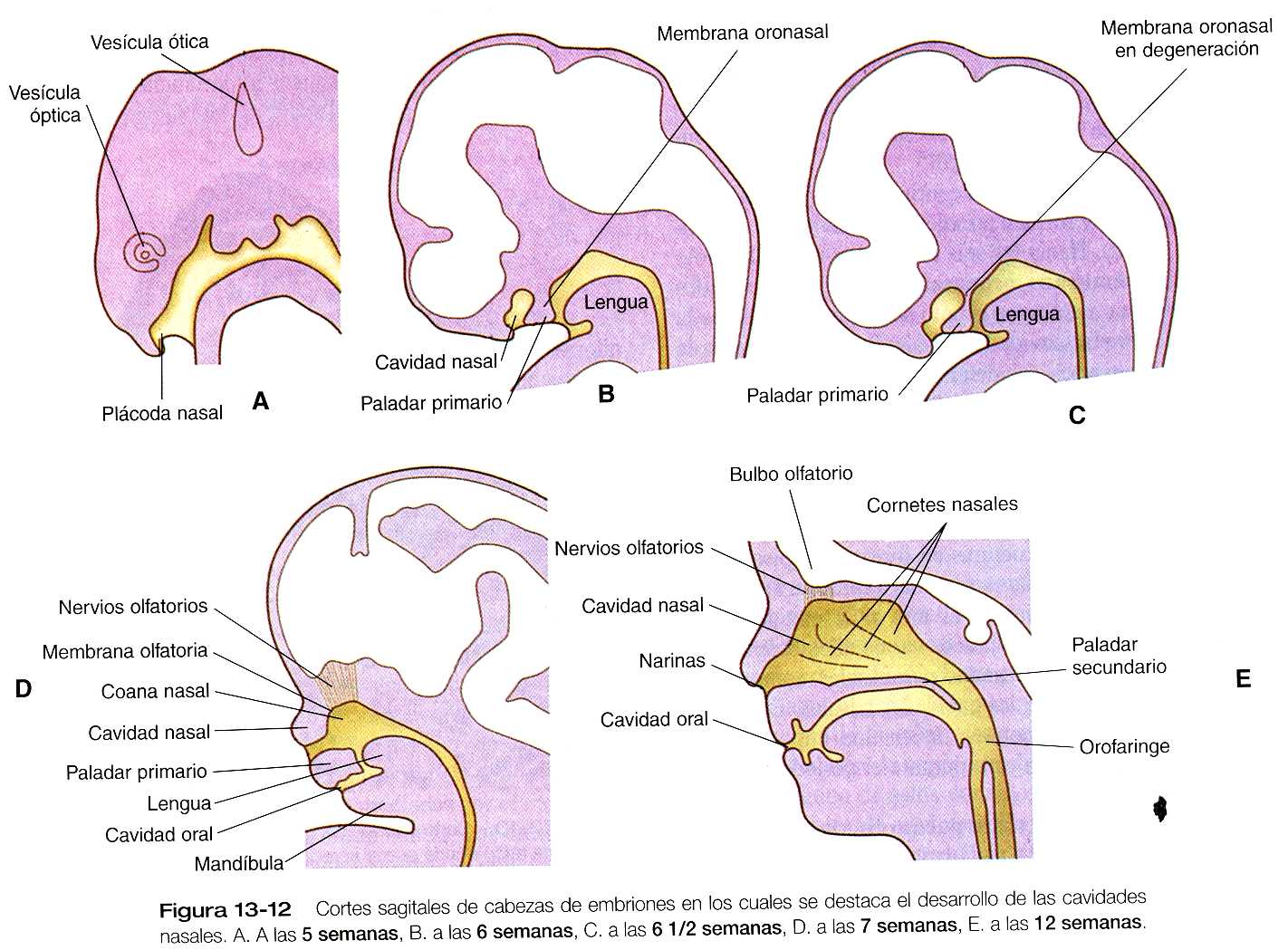
El aparato olfatorio humano comienza a hacerse visible al final del primer mes como un par de plácodas nasales ectodérmicas engrosadas localizadas en la región frontal de la cabeza (figura 13_12A). Estas plácodas se originan a partir del borde anterolateral de la placa neural antes de su cierre. La formación de las plácodas nasales, al igual que las del cristalino, requiere la expresión del Pax-6. si este gen no se expresa ninguno de estos dos tipos de plácodas se forma.
Poco después de su desarrollo, las plácodas nasales dan lugar a una depresión superficial (fóvea o fosa nasal) rodeada por unas elevaciones de tejido mesenquimatoso en forma de herradura cuyos extremos abiertos miran hacia la futura boca (figura 13_5). Los dos brazos de las elevaciones mesenquimatosas son los procesos nasal medial y nasal lateral. A medida que los primordios nasales se dirigen hacia la línea media durante la sexta y la séptima semanas, los procesos nasomediales forman la punta y la cresta nasal junto con parte del tabique nasal, y los procesos nasolaterales dan origen a las alas de la nariz. El proceso frontonasal contribuye en parte al fuente de la nariz.
Entre tanto, las fosas nasales continúan invaginándose hacia la cavidad oral y forman también unas cavidades evidentes (figura 13_12). Hacia las seis semanas y media tan sólo una delgada membrana oronasal separa a las cavidades oral y nasal. Esta membrana pronto se rompe y establece la continuidad entre las cavidades nasales y la oral a través de unos orificios situados por detrás del paladar primario que se llaman coanas nasales primitivas. Con la fusión de los procesos palatinos laterales, la cavidad nasal se alarga de forma considerable y termina comunicándose con la región superior de la faringe.
Durante el tercer mes, se desarrollan en la pared lateral de la cavidad nasal unas estructuras en forma de repisa llamadas cornetes nasales. Esas estructuras aumentan la superficie disponible para aclimatar el aire a la cavidad. Más adelante en la vida fetal y durante varios años después del nacimiento se forman los senos paranasales como crecimientos de las paredes de las cavidades nasales. El tamaño y la forma de estas estructuras tienen una importante influencia en la forma de la cara durante el periodo de crecimiento posnatal.
Hacia la mitad del embarazo puede observarse un par de invaginaciones epiteliales a cada lado del tabique nasal cerca del paladar. Estos divertículos, llamados órganos vomeronasales, alcanzan su tamaño máximo de 6 a 8 mm. Cerca del sexto mes de la gestación y entonces comienzan a regresar por completo o dejan pequeñas estructuras quísticas. En la mayoría de los mamíferos y en muchos otros vertebrados los órganos vomeronasales, que están revestidos por un epitelio olfatorio modificado, tienen importancia y al parecer intervienen en la percepción del olor de la comida en la boca o en la percepción de los estímulos sexuales (por ejemplo, las feromonas).
El epitelio más dorsal de las fosas nasales se transforma en un epitelio olfatorio muy especializado (figura 13_12). Desde el periodo embrionario y durante toda la vida, el epitelio olfatorio puede formar neuronas bipolares sensoriales primitivas que emiten prolongaciones axonales hacia el bulbo olfatorio del cerebro. Antes del crecimiento axonal, algunas células se desprenden del epitelio y migran hacia el cerebro. Algunas de ellas pueden sintetizar un sustrato para guiar el crecimiento de los axones olfatorios. Otras células que migran de la plácoda olfatoria son capaces de sintetizar la hormona liberadora de la hormona luteinizante y alcanzan el hipotálamo. Las células de la plácoda olfatoria también forman células de soporte (sustentaculares) y células glandulares en la región olfatoria de la nariz. Hay evidencias fisiológicas que indican que el epitelio olfatorio puede desempeñar alguna función al final de la vida fetal pero su función olfatoria completa sólo se alcanza después del nacimiento.
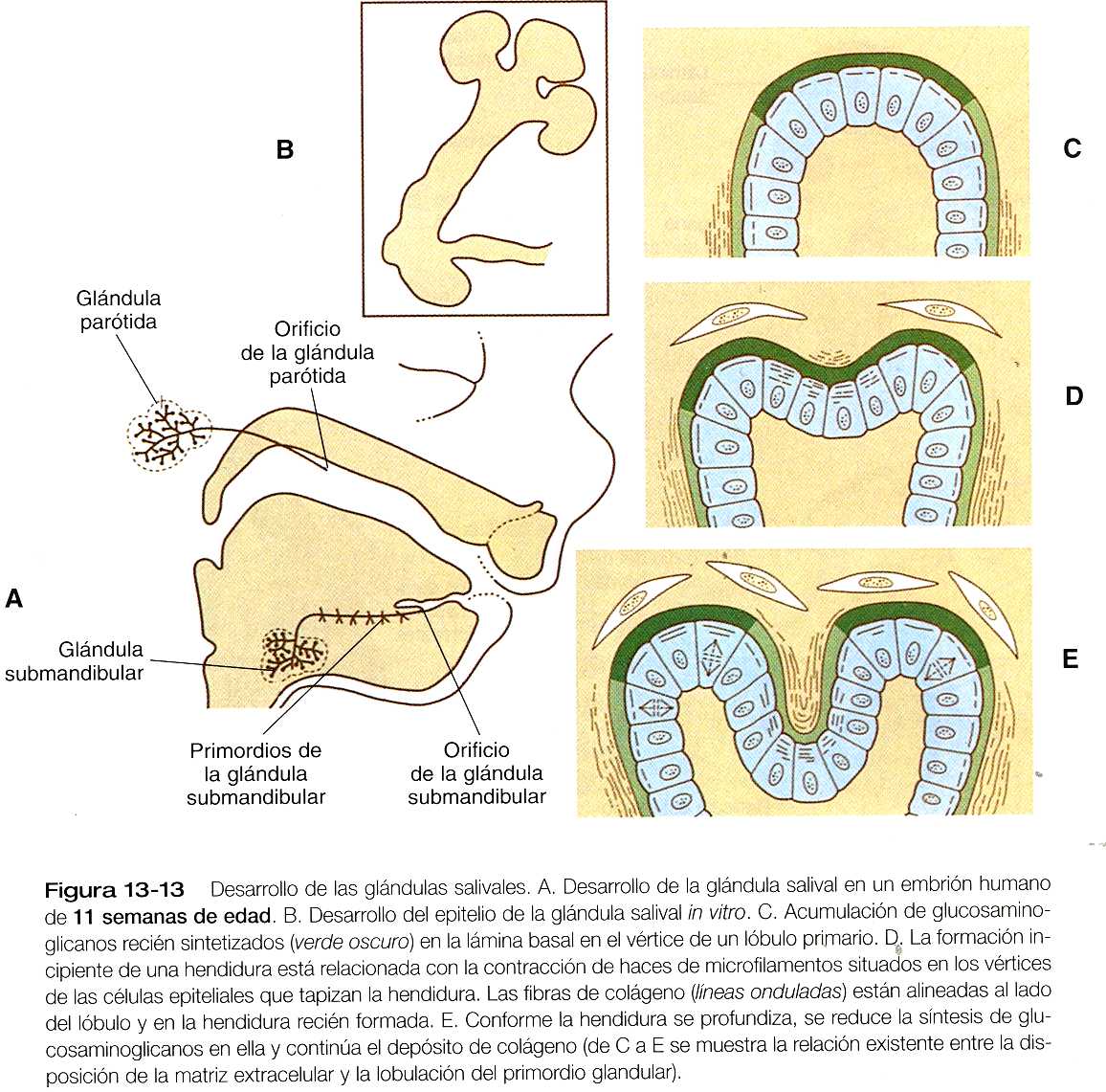
Formación de las glándulas salivales
Las glándulas salivales comienzan a aparecer a la sexta semana y se originan como engrosamientos macizos en forma de crestas del epitelio oral (figura 13_13). Los grandes desplazamientos epiteliales que tienen lugar en la cavidad oral dificultan el poder determinar la capa germinal de origen del epitelio de las glándulas salivales. Es probable que las parótidas se deriven del ectodermo, y que tanto las submandibulares como las sublinguales provengan del endodermo.
Como en otras estructuras glandulares asociadas con el tracto digestivo, el desarrollo de las glándulas salivales depende de una serie continua de interacciones entre el epitelio y el mesénquima. La estructura y composición de la lámina basal que rodea a las invaginaciones epiteliales lobulares iniciales difiere en su composición en función del potencial de crecimiento de la región. Alrededor del pedículo y en las hendiduras, la lámina basal contiene colágeno de tipos I y IV y un proteoglicano 1 de la membrana basal (MB-1). Estos componentes no se encuentran en las regiones de los lóbulos que siguen creciendo. Bajo la influencia del mesénquima circundante, la lámina basal de las regiones que crecen pierde los colágenos y los proteoglicanos que se asocian con las estructuras estables (por ejemplo, los pedículos y las hendiduras). Además de las alteraciones en la lámina basal, la ramificación se asocia con la contracción local de microfilamentos situados en los vértices de las células epiteliales ubicados en los puntos de ramificación. El crecimiento continuo en los vértices de las células epiteliales ubicados en los puntos de ramificación. El crecimiento continuo en los vértices de los lóbulos de las glándulas está mantenido por los altos niveles de actividad mitótica del epitelio y por el depósito en el área de glucosaminoglicanos recién sintetizados. La diferenciación estructural y funcional del epitelio de las glándulas salivales continúa durante toda la vida fetal.
Formación de los dientes
Un diente tiene una matriz extracelular muy especializada que consta de dos componentes principales –esmalte y dentina— cada uno de los cuales es producto de un epitelio embrionario diferente. El desarrollo de los dientes es un proceso muy coordinado que comprende íntimas interacciones entre los epitelios que producen el esmalte y la dentina. Haciendo extensión del desarrollo hacia la dimensión macroscópica, los dientes experimentan una transición isomórfica, con la sustitución posnatal de los dientes caducos por sus contrapartes permanentes.
Etapas del desarrollo de los dientes
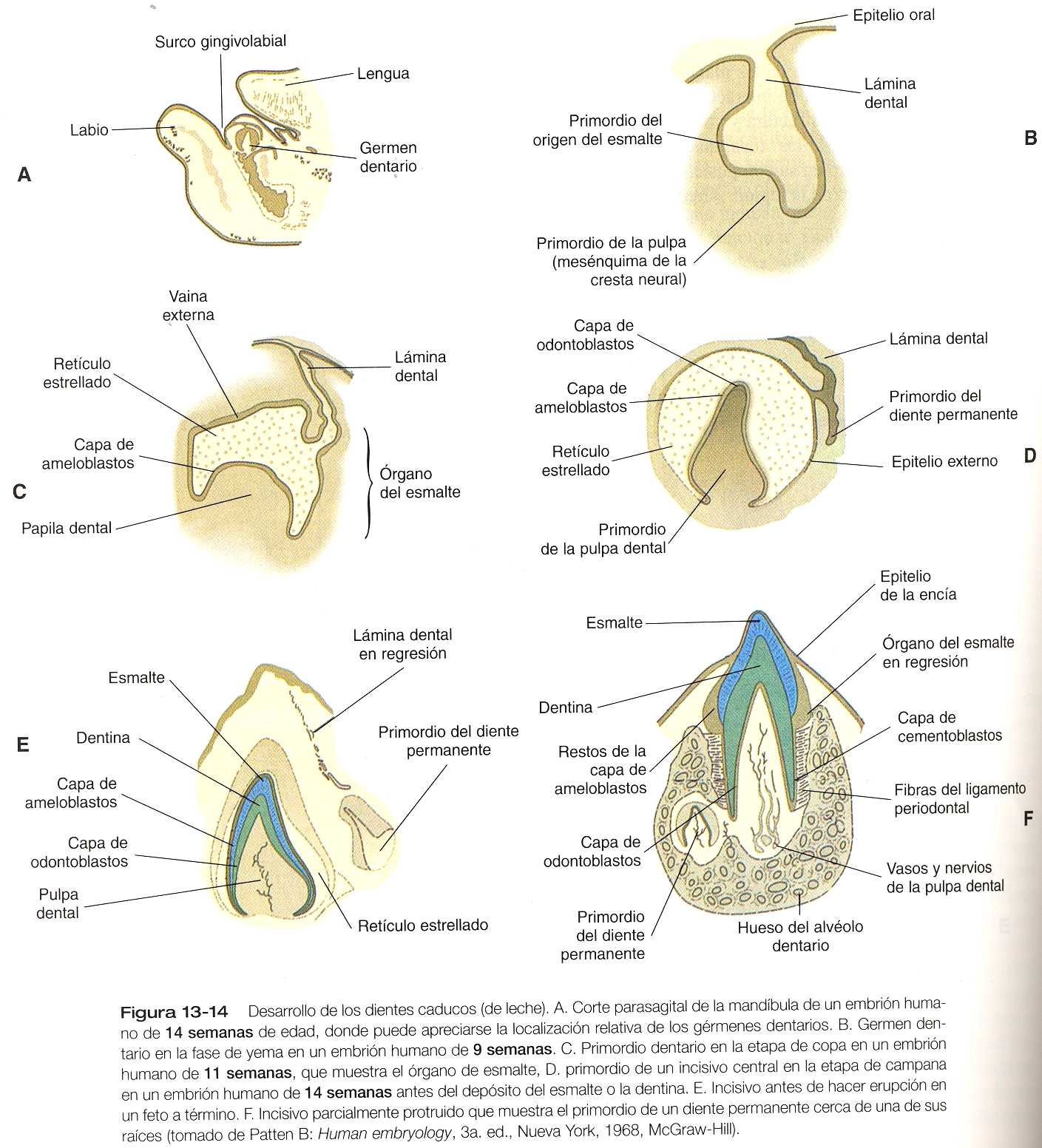
El desarrollo de los dientes comienza con la migración de células de la cresta neural hacia el maxilar y la mandíbula. Algunas de estas células mesenquimatosas derivadas de la cresta neural vienen determinadas para formar estructuras dentarias; Actúan sobre el ectodermo oral que las cubre, el cual se engruesa en bandas con forma de C (láminas dentales) en el maxilar y la mandíbula. La aparición de las láminas dentales durante la sexta semana es la primera manifestación de una serie de interacciones entre el ectodermo y el mesodermo, que continúa hasta que la formación de los dientes está prácticamente terminada.
Aunque cada diente tiene una secuencia de tiempo y un desarrollo morfológico específicos, algunas fases del desarrollo son comunes a todos ellos (figura 13_14). Conforme la lámina dental crece en el mesénquima de la cresta neural, comienzan a formarse los primordios epiteliales de los dientes individuales como brotes o gérmenes dentarios. de acuerdo con el desarrollo interactivo que tienen los gérmenes dentarios se asocian con condensaciones de células mesenquimatosas. El germen pronto se expande y pasa desde una etapa de copa en la que tiene forma de hongo, hasta la etapa de campana.
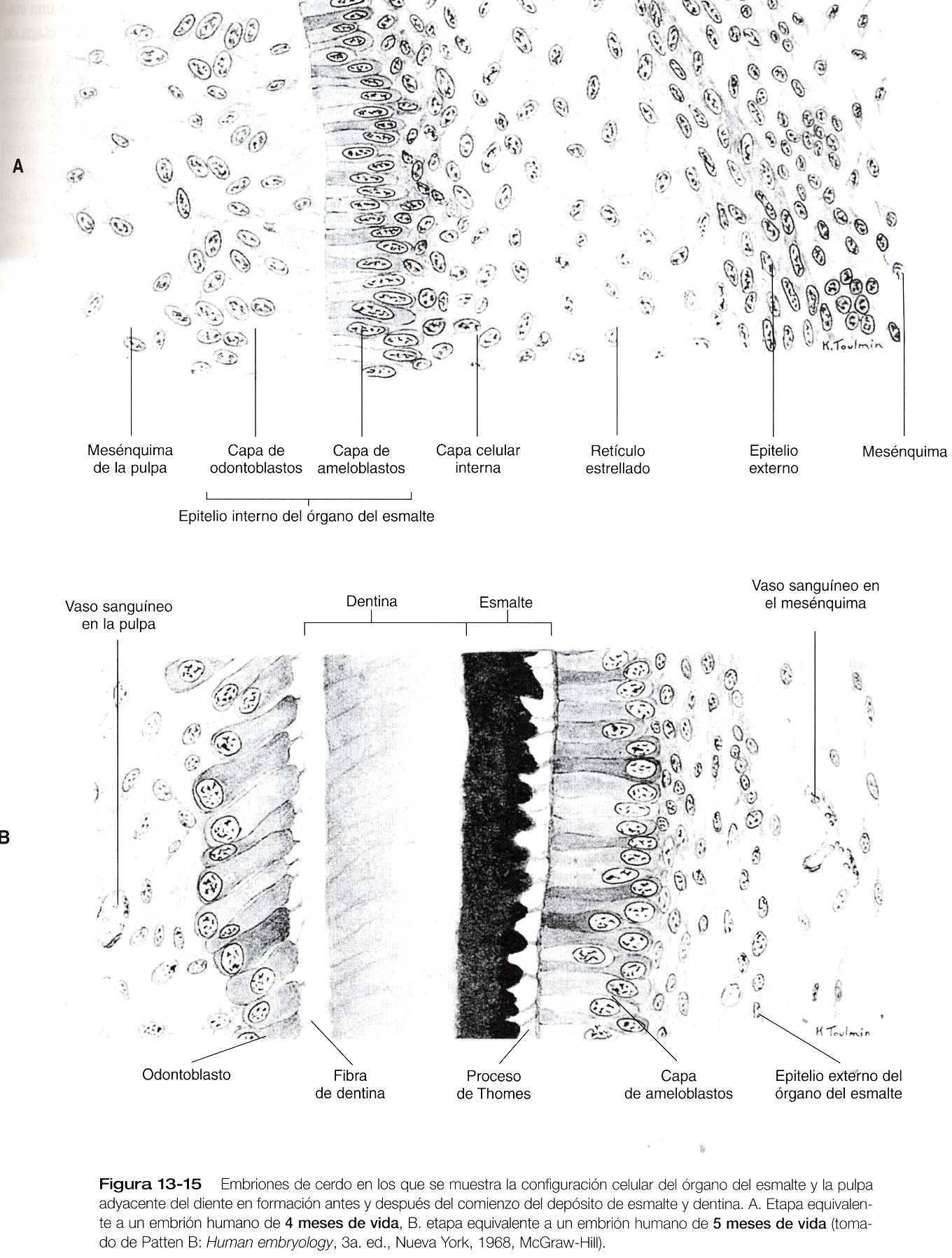
Hacia la etapa de campana el primordio del diente ya tiene una estructura compleja, aunque no ha formado ninguno de los componentes del diente definitivo. El componente epitelial, llamado órgano del esmalte, aún está conectado al epitelio oral por un tallo irregular de lámina dental, que pronto comienza a degenerarse. El órgano del esmalte consta de una vaina externa de epitelio, un retículo estrellado mesenquimatoso y una capa epitelial interna de ameloblastos. Los ameloblastos son las células que comienzan a segregar el esmalte del diente. En la superficie cóncava del órgano del esmalte se encuentra en una condensación de mesénquima de la cresta neural denominada papila dental. Las células de la papila situadas en oposición a la capa de ameloblastos se transforman en células epiteliales cilíndricas llamadas odontoblastos (figura 13_15), que producen la dentina. Conectando a la lámina dental cerca del órgano del esmalte se encuentra una pequeña yema del diente permanente (figuras 13_14 E y F). Aunque más tarde, la yema pasa por las mismas etapas del desarrollo que los dientes caducos.
Ya avanzada la etapa de campana los odontoblastos y los ameloblastos empiezan a segregar los precursores de la dentina y del esmalte, comenzando por el futuro ápice del diente. En el transcurso de varios meses el diente va adquiriendo su forma definitiva (figura 13_14). Entre tanto, alrededor del diente en desarrollo se forma una condensación de células mesenquimatosas. Las células de esta estructura, llamada saco dental, producen componentes especializados de la matriz extracelular (el cemento y el ligamento periodontal) que constituyen un anclaje sólido para el diente en la mandíbula o el maxilar. Mientras estos fenómenos están sucediendo, el diente se alarga y comienza su erupción a través de las encías.
Interacciones tisulares en el desarrollo del diente
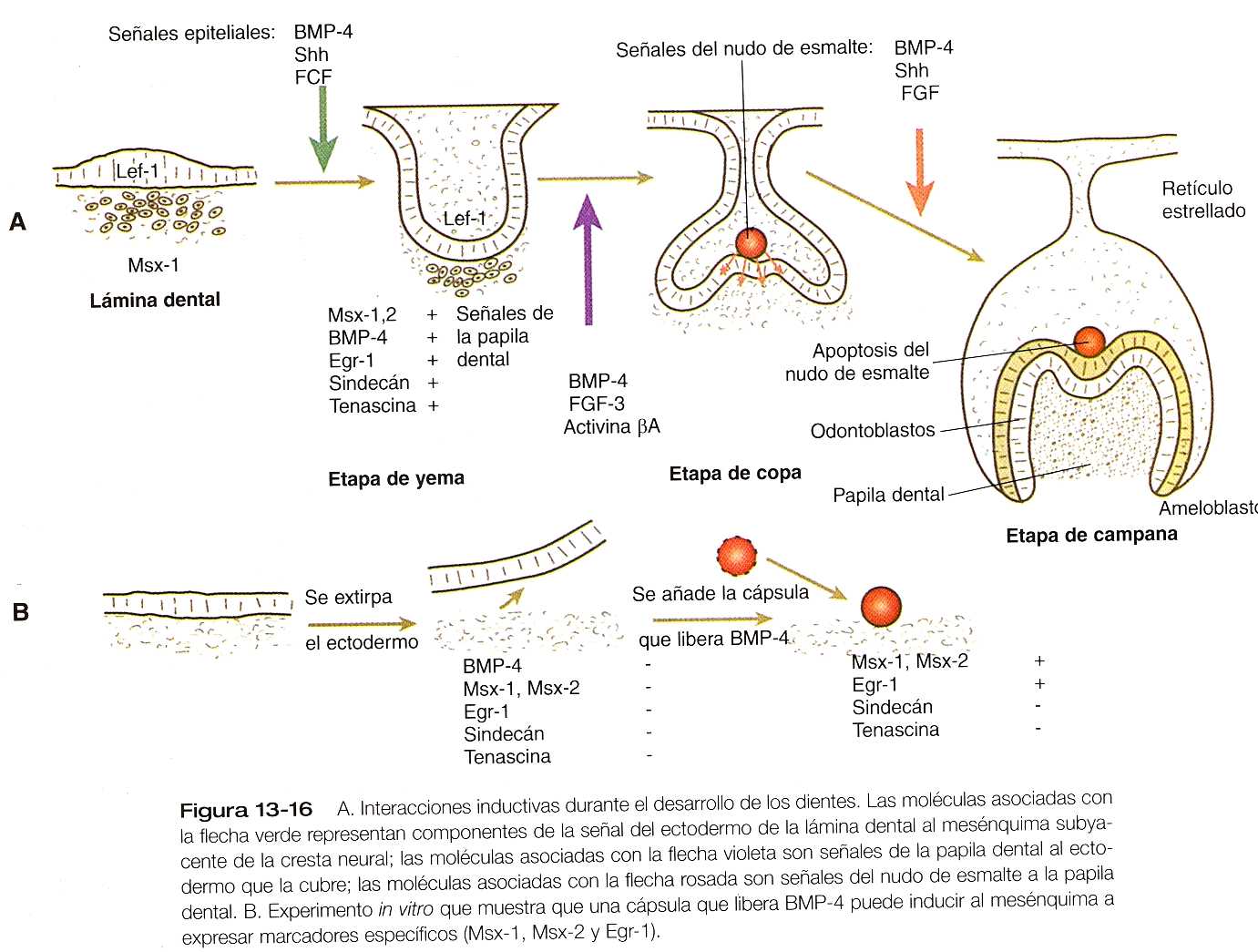
Los experimentos de recombinación tisular han demostrado que el ectodermo engrosado de la lámina dental inicia el desarrollo de los dientes. Las células ectodérmicas expresan el factor de transcripción Lef-1 (factor intensificador de los linfocitos) que, de una forma aún desconocida, lleva a la célula ectodérmica a producir las moléculas de señal BMP-4, FGF-8 y sonic hedgehog (figura 13_16). Estas señales ectodérmicas actúan sobre el mesénquima subyacente derivado de la cresta neural y hacen que participe en la formación del diente. La determinación del mesénquima dental se refleja en la expresión de varias moléculas características, entre las que se cuentan los factores de transcripción Msx-1, Msx-2 y Egr-1 -early growth response- (respuesta temprana de crecimiento-1); las moléculas de la matriz extracelular tenascina y sindecán, las cuales promueven la cohesión de las células de mesénquima; y BMP-4.
Si el ectodermo suprayacente se separa del mesénquima de la cresta neural, éste último no se determina para formar un diente. El papel del BMP-4 en la inducción inicial del mesénquima dental se demostró añadiendo una pequeña cápsula empapada en BMP-4 a una pequeña masa de mesénquima de la cresta neural cultivada (figura 13-16B). Bajo la influencia del BMP-4 liberado de la cápsula, el mesénquima comenzó a expresar Msx-1, Msx-2, Egr-1 y BMP-4, pero no produjo tenascina ni sindecán; esto demostró que se requieren otras señales además del BMP-4 para alcanzar la respuesta inductiva completa.
Con la inducción inicial del mesénquima dental, ese tejido se convierte en el motor principal del desarrollo del diente. Las señales inductivas que emanan del mesénquima dental que actúan a continuación sobre el ectodermo del borde dental, que ahora se encuentra en la etapa de yema tardía o copa incipiente. Los experimentos de recombinación han demostrado que el mesénquima dental determina la forma específica del diente. Cuando se combina mesénquima molar in Vitro con epitelio de incisivo, se desarrolla una muela, mientras que al combinar mesénquima de incisivo con epitelio molar el resultado es la formación de un incisivo.
Bajo la influencia inductora del mesénquima dental, ahora llamado papila dental, se forma el nudo de esmalte, un nuevo e importante centro situado en el ectodermo suprayacente (figura 13_16). Las células de este nudo producen varias moléculas de señal, entre ellas el sonic hedgehog, el FGF-4, la BMP-2, la BMP-4, y la BMP-7. estas señales se transmiten hacia los lados a través del epitelio de la copa dental y también hacia atrás, al mesénquima de la papila dental, y parecen estimular la proliferación y la diferenciación de los epitelios ameloblástico y odontoblástico. En último término, las células del nudo del esmalte sufren apoptosis; se ha postulado que debido a este mecanismo terminan las señales inductivas que se producen en esta estructura.
Formación de la dentina y el esmalte
Ya avanzada la diferenciación, los odontoblastos dejan el ciclo celular, se alargan y comienzan a segregar predestina en sus superficies apicales, situadas de cara al órgano del esmalte. La producción de predestina señala un desplazamiento en los patrones de síntesis del colágeno de tipo 1 y otras moléculas (por ejemplo, fosfoproteína de dentina, osteocalcina de dentina) que caracterizan a la matriz de la dentina. La primera dentina se deposita contra la superficie interna del órgano del esmalte en el ápice del diente (figura 13_15B). Con la secreción dentina adicional, el material acumulado empuja el epitelio odontoblástico de la interzona odontoblasto-ameloblasto.
La diferenciación terminal del los ameloblastos se produce después de que los odontoblastos empiezan a segregar predestina. En respuesta a señales inductivas de los odontoblastos, los ameloblastos dejan el ciclo celular y comienzan un nuevo patrón de síntesis, en el cual se producen dos clases de proteínas: las amelogeninas y las esmaltinas aproximadamente el 5% del esmalte es matriz orgánica, y cerca del 90% de ésta corresponde a las amelogeninas; las esmaltinas componen casi todo el resto. Éstas se secretan antes de las amelogeninas y pueden servir de núcleos para la formación de los cristales de hidroxiapatita, el principal componente inorgánico del esmalte.
Los genes de amelogenina están localizados en los cromosomas X y Y en los seres humanos. Los del esmalte se han conservado considerablemente durante toda la filogenia de los vertebrados. Se ha sugerido que en los primeros vertebrados el esmalte sólo servía como parte de un aparato electrorreceptor.
Erupción y sustitución de los dientes
Cada pieza dental tiene un momento específico para su erupción y su sustitución (tabla 13_1). Con el crecimiento de la raíz, la corona cubierta de esmalte empuja a través del epitelio oral y sobresale. La secuencia de la erupción comienza con los incisivos centrales, casi siempre unos pocos meses después del nacimiento, y suele proseguir paso a paso hasta que se forman los últimos molares al final del segundo año. En total se forman 20 dientes caducos o de leche.
Entre tanto, el primordio de los dientes permanentes está embebido en una cavidad que se extiende hacia el hueso del lado lingual de la cavidad alveolar en la cual el diente está anclado (figura 13_14F). Conforme se desarrollan los dientes permanentes, su crecimiento causa la reabsorción de la raíz, éste se cae y deja el espacio para que el diente permanente ocupe su lugar. La secuencia de la erupción de los dientes permanentes es la misma que la de los caducos, pero se forman doce piezas dentales permanentes más (para un total de 32) sin que haya los dientes caducos correspondientes.
La formación y la erupción de los dientes son factores importantes en el crecimiento de la región media de la cara, que tiene lugar en gran parte después del nacimiento. El desarrollo de los dientes y el crecimiento correspondiente de la mandíbula para dar campo a las piezas, junto con el desarrollo de los senos paranasales, explican en gran parte de la masa de la región media de la cara.
DESARROLLO DE LA FARINGE Y SUS DERIVADOS
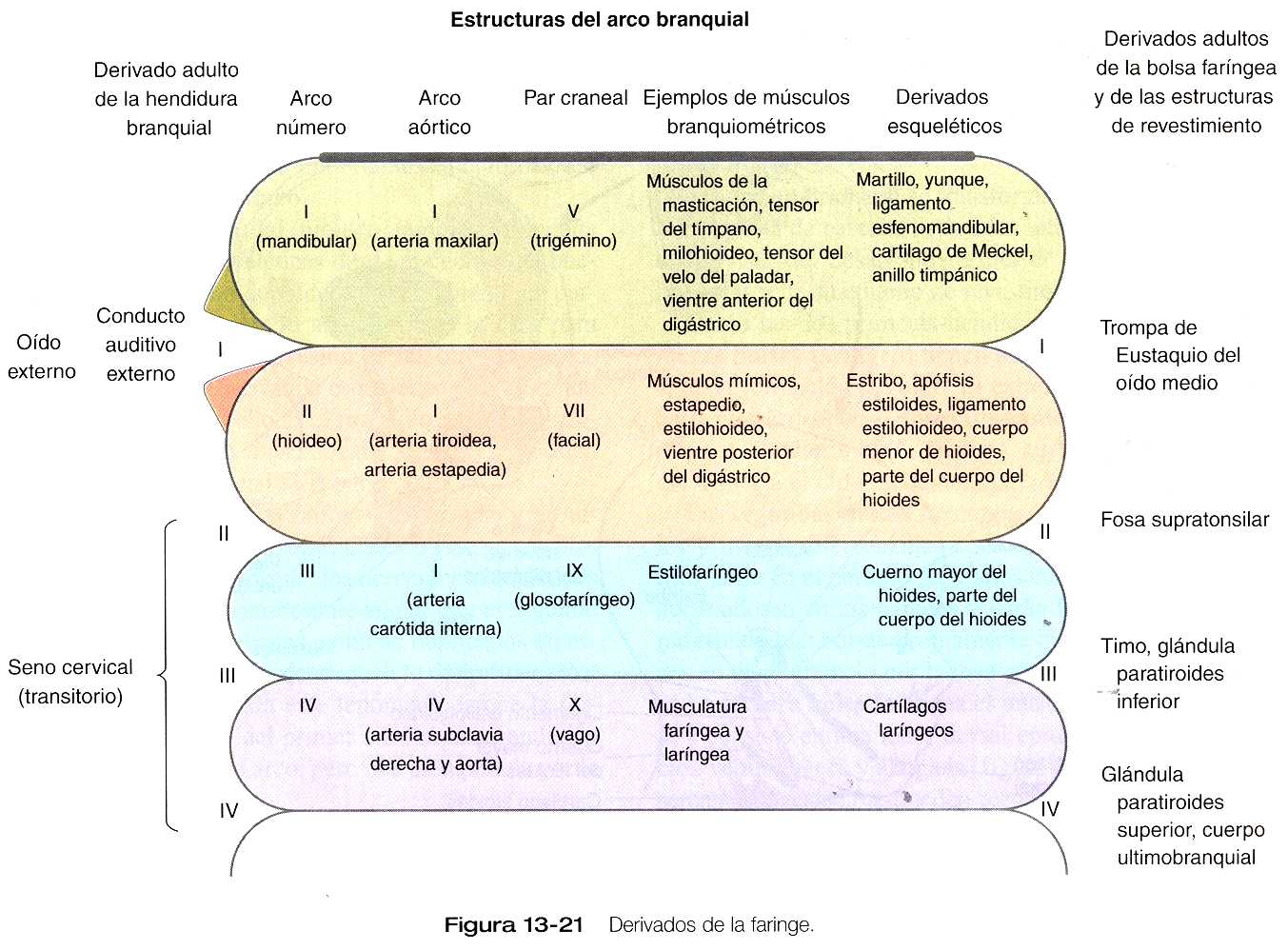
Si se tiene en cuenta la complejidad de la organización estructural de la faringe embrionaria, no es de extrañarse que una gran diversidad de estructuras se originen en la región faríngea; ello permite muchas posibilidades para el desarrollo anómalo, las cuales se discuten en la correlación clínica 13_2a b y c (págs. 317 a 319). En esta sección se explican en detalle aspectos del desarrollo tardío que conducen a la formación de estructuras específicas. Los derivados en el adulto de las regiones de la faringe y los arcos branquiales se resumen en la figura 13_21.
Desarrollo externo de la región faríngea
En su superficie, la región branquial (faríngea) se caracteriza por cuatro arcos branquiales con las hendiduras que los separan (figura 13_4). Estas estructuras dan origen a múltiples derivados.
Arcos branquiales
Además de estar llenos de mesénquima (en su mayoría derivado de la cresta neural, con excepción del mesodermo premuscular, que procede de los somitómeros), cada arco branquial tiene una arteria principal (arco aórtico) y un par craneal (figura 13_21). Cada uno también tiene un cordón central de mesénquima cartilaginoso, que dará lugar a derivados esqueléticos adultos específicos. Es muy importante comprender la relación existente entre los arcos branquiales y su inervación y su irrigación sanguínea, ya que los tejidos suelen mantener su relación con su nervio original conforme emigran o se desplazan desde su lugar de origen en el sistema de los arcos branquiales.
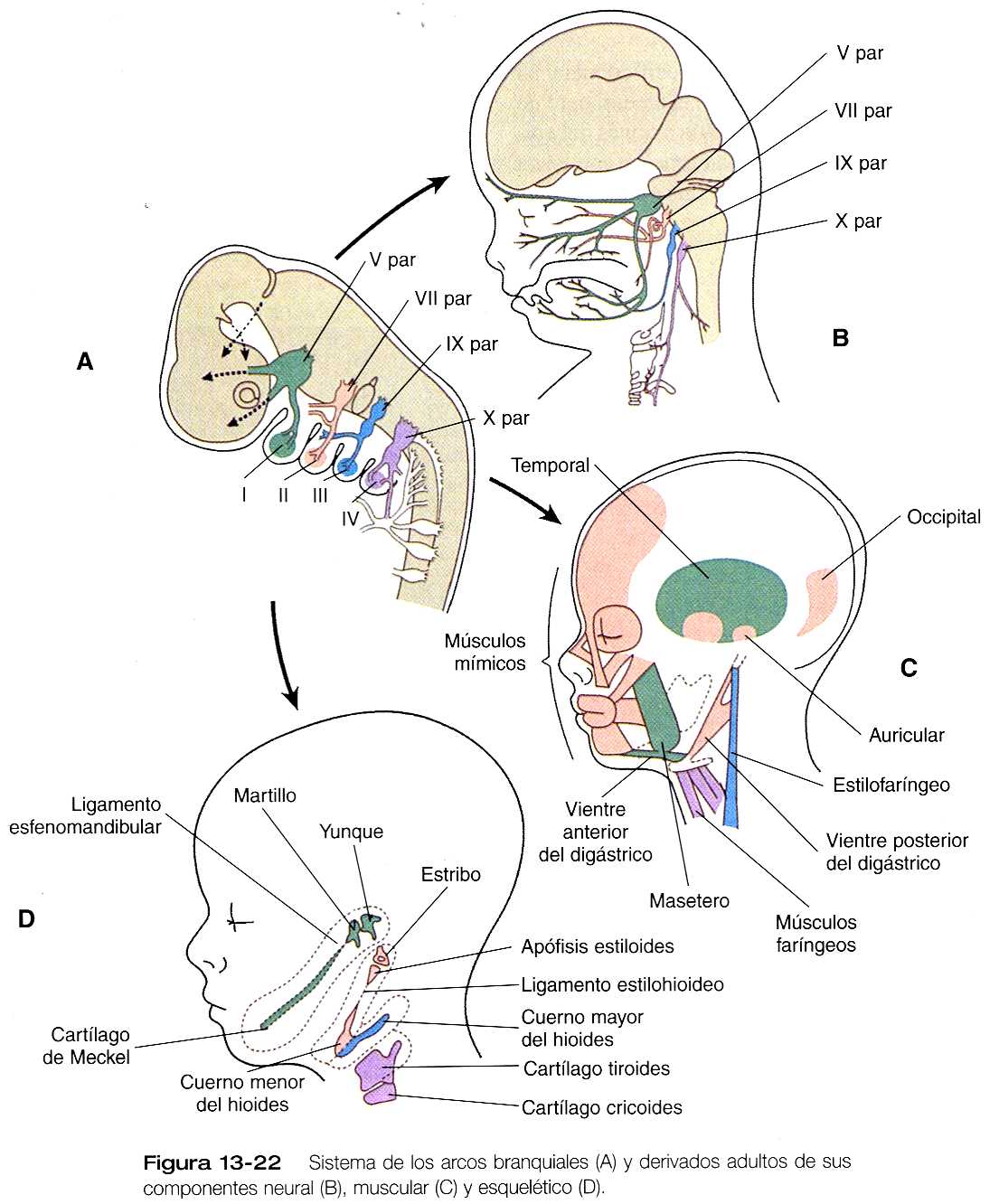
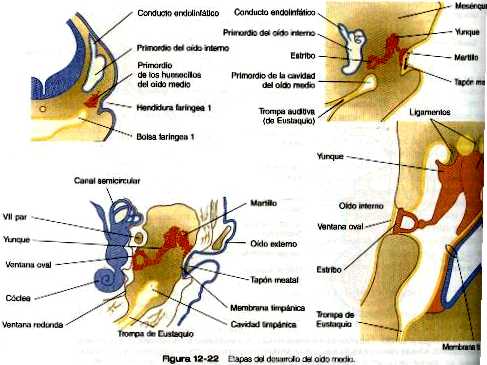
El primer arco branquial (mandibular) contribuye sobre todo a estructuras de la cara (tanto las porciones maxilares como mandibulares) y del oído (figura 13_21). Su cordón cartilaginoso central, el cartílago de Meckel, es un componente importante de la mandíbula embrionaria hasta que es rodeado por hueso intramembranoso formado localmente, y que da lugar a la mandíbula definitiva. Al final del desarrollo la parte distal del cartílago de Meckel experimenta una reabsorción a causa de una gran apoptosis de los condrocitos. En su región más dorsal, el cartílago de Meckel forma el ligamento esfenomandibular, el ligamento anterior del martillo, y el martillo (figura 13_22). Además, el yunque surge de un primordio del cartílago cuadrado. La musculatura del primer arco se relaciona con la masticación, la faringe y el oído medio. Una característica común de estos músculos es su inervación por el trigémino (V par).
Una molécula de activación (señales), la endotelina-1 (ET-1), desempeña un papel importante en el desarrollo del primer arco faríngeo. La ET-1 se expresa principalmente en el epitelio que cubre el arco y, en su ausencia, la mayor parte de las estructuras derivadas de este arco no se forman adecuadamente. La ET-1 está implicada en las señales epiteliales y de la cresta neural que son importantes en la formación de los derivados del primer arco.
El segundo arco branquial (hioideo) también forma una cadena de estructuras esqueléticas desde el cuerpo del hueso hioides hasta el estribo del oído medio. Si bien gran parte del mesodermo del segundo arco migra a la cara para formar los músculos de la expresión facial, otros músculos se relacionan con otros derivados esqueléticos del segundo arco, un buen ejemplo de ello es el músculo estapedio asociado con el estribo. Estos músculos del segundo arco están inervados por el nervio facial (VII par).
El patrón del segundo arco faríngeo recibe una gran influencia del gen homeobox Hoxa-2. Cuando este gen se bloquea en los ratones, no se forman los derivados esqueléticos del segundo arco. Resulta interesante anotar que el segundo arco en estos animales mutantes contiene duplicados en espejo de muchos de los huesos del esqueleto del primer arco. Una posible explicación para este fenómeno es que la formación de las estructuras del primer arco es una condición previa para las del segundo arco, pero que es superada por la influencia del Hoxa-2.
Los arcos branquiales III y IV no tienen otro nombre. El tercero da lugar a las estructuras relacionadas con el hueso hioides y la porción superior de la faringe. El esqueleto del tercer arco se convierte en el cuerno mayor del hueso hioides. El derivado muscular (estilofaríngeo) del tercer arco está inervado por el nervio glosofaríngeo (IX par). El cuarto arco da origen a ciertos músculos y cartílagos de la laringe y la parte inferior de la faringe. Los músculos están inervados por el vago (X par), que también se prolonga hacia las cavidades torácica y abdominal.
Hendiduras branquiales
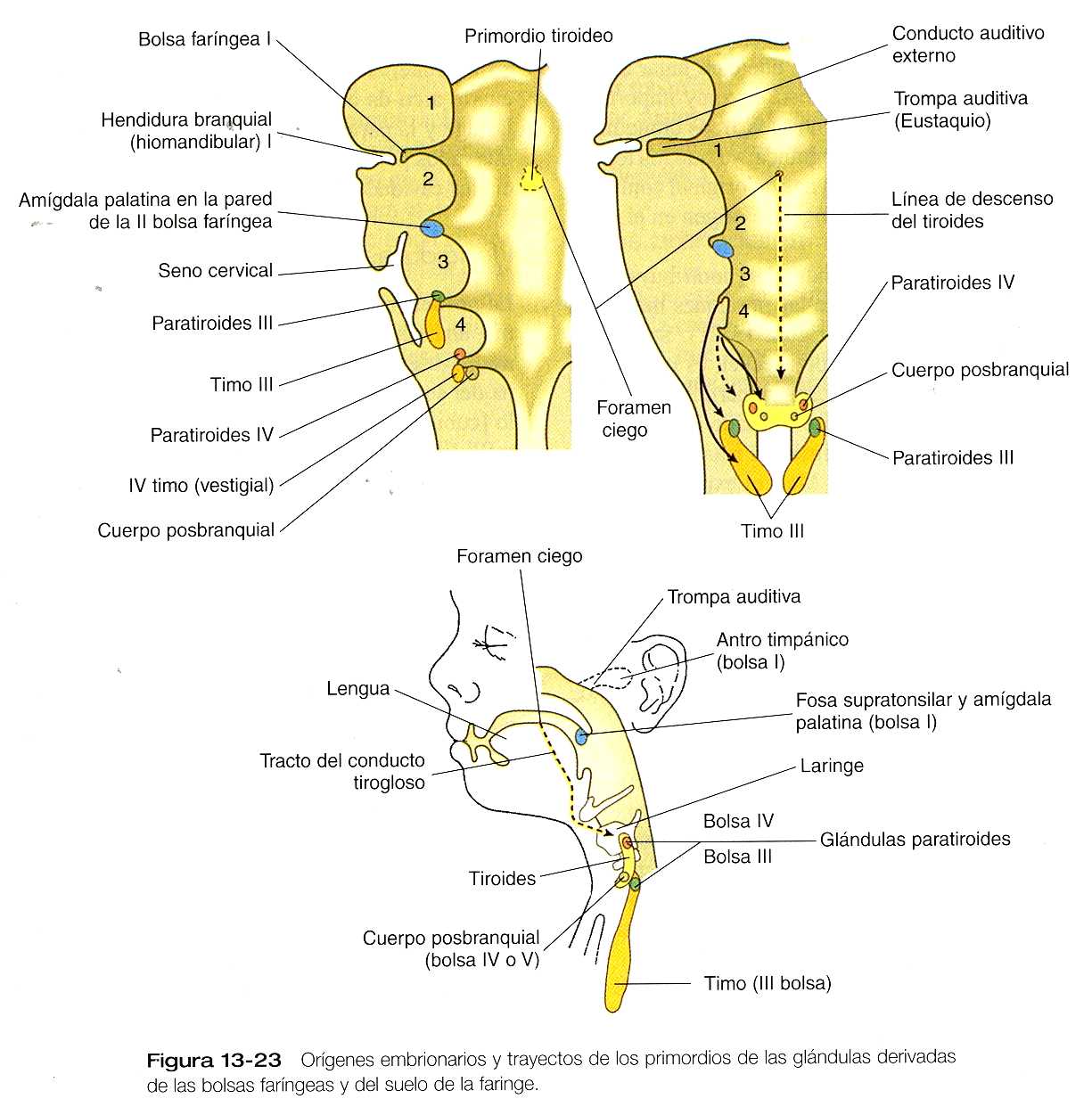
La primera hendidura branquial es la única que persiste como una estructura reconocible en el adulto: el conducto auditivo externo. Al aumentar de tamaño la porción externa del segundo arco (un homólogo filogenético del opérculo [cubierta de agallas] de los peces) cubre los surcos II y III. El crecimiento del segundo arco se debe a la presencia de un centro de emisión de señales en el vértice de su ectodermo, los arcos III y IV carecen de este centro de señales. Al igual que en los primordios faciales, este centro de señales produce sonic hedgehog, FGF-8 y BMP-7, los cuales estimulan el crecimiento del mesénquima subyacente. Durante el periodo en el que el arco hioideo (segundo) los cubre, los surcos II y III reciben el nombre colectivo de seno cervical (figura 13_23). Conforme avanza el desarrollo, el ectodermo de una eminencia (eminencia cardiaca) localizada inmediatamente caudal al cuarto arco, y esto hace que el seno cervical desaparezca y que se modifiquen los contornos externos del cuello.
Faringe y bolsas faríngeas
La faringe embrionaria se transforma directamente en la faringe adulta de paredes lisas. Más interesante desde el punto de vista del desarrollo resulta el destino de las bolsas faríngeas y de su epitelio de revestimiento.
Como las dos primeras hendiduras branquiales, las primeras bolsas faríngeas tienen una participación importante en la formación del oído. El extremo de cada bolsa crece hasta convertirse en el antro timpánico del oído medio, y el resto se convierte en la trompa auditiva (de Eustaquio) que conecta el oído medio con la faringe (figura 12_22).
Las segundas bolsas faríngeas se hacen más superficiales y menos diferenciadas conforme el desarrollo avanza. Más tarde se adosan en las paredes y darán lugar a las amígdalas palatinas. Las bolsas propiamente dichas está representadas en el adulto sólo por la fosa supratonsilar.
La tercera bolsa faríngea es una estructura más compleja y consiste en una masa dorsal epitelial sólida y una porción ventral hueca y alargada (figura 13_23). Hacia la quinta semana de gestación, pueden reconocerse en el endodermo de la masa sólida dorsal unas células identificables como tejido paratifoideo. La elongación ventral de la tercera bolsa se diferencia en la porción epitelial del timo. Los primordios del timo y de las glándulas paratiroides pierden su conexión con la tercera bolsa faríngea y migran en su dirección caudal a partir de su lugar de origen. Aunque el III primordio paratiroideo inicialmente migra junto con el primordio tímico, continúan su migración hacia la línea media. Allí se adosan a la glándula tiroides, y sobrepasan los primordios paratiroideos de la cuarta bolsa para formar las glándulas paratiroides inferiores. La tercera bolsa faríngea desaparece.
La cuarte bolsa faríngea se estructura de forma algo parecida a como lo hace la tercera, con un IV primordio paratifoideo sólido bulboso y dorsal. Además, contiene una pequeña evaginación epitelial ventral, que en algunas especies aporta algún componente al timo. En los seres humanos el componente tímico de la cuarta bolsa es vestigial. En la parte más ventral de cada cuarta bolsa se encuentra otra estructura llamada cuerpo posbranquial (ultimobranquial) como se ilustra en la figura 13_23.
No se sabe con certeza el origen celular y la composición de los cuerpos posbranquiales. Según una teoría los cuerpos posbranquiales derivan exclusivamente del endodermo faríngeo, algunos datos recientes sugieren que las células de la cresta neural migran hacia los cuerpos posbranquiales y terminan por convertirse en el componente secretorio de estas estructuras.
Como sucede con sus homólogas de la tercera bolsa, el IV primordio paratifoideo pierde su conexión con la cuarta bolsa y migra hacia la glándula tiroides y formará las glándulas paratiroides superiores. Los cuerpos posbranquiales también migran hacia la tiroides, y se incorporan a ella como células parafoliculares o C. estas células, cuyo origen está en la cresta neural, producen la hormona polipeptídica calcitonina, cuya acción es reducir la concentración de calcio en la sangre. Las glándulas paratiroides producen hormona paratifoidea, que aumenta los niveles de calcio en la sangre.
Estructuras de la línea media que surgen de la faringe
El primordio impar de la glándula tiroides aparece en la línea media ventral de la faringe entre la primera y la segunda bolsas (figura 13_23). El primordio tiroideo comienza en la cuarta semana como engrosamiento endotelial en situación inmediatamente caudal al tubérculo medio de la lengua (tubérculo impar) y pronto se alarga para formar un conducto descendiente llamada divertículo tiroideo. La extensión caudal de este divertículo crece y se bifurca para formar la glándula tiroides propiamente dicha, que consta de dos lóbulos principales conectados por un istmo. Durante algún tiempo la glándula sigue unida a su lugar de origen mediante un estrecho conducto tirogloso. Hacia la séptima semana, cuando el tiroides ha alcanzado su localización final en el nivel del segundo y el tercero cartílagos traqueales, el conducto tirogloso se mantiene y constituye el lóbulo piramidal del tiroides. El lugar de origen del primordio tiroideo persiste como el foramen ciego, una pequeña depresión ciega localizada en la base de la lengua.
La glándula tiroides experimenta su diferenciación histológica y se vuelve funcional relativamente pronto en el desarrollo embrionario. Hacia la décima semana de gestación, se hacen evidentes los folículos que ya contienen algo de material coloide, y unas pocas semanas después de la glándula comienza a sintetizar tiroglobulina no yodada. La secreción de triyodotironina, una de las formas de la hormona tiroidea, puede detectarse hacia el final del cuarto mes.
Timo y órganos linfoides
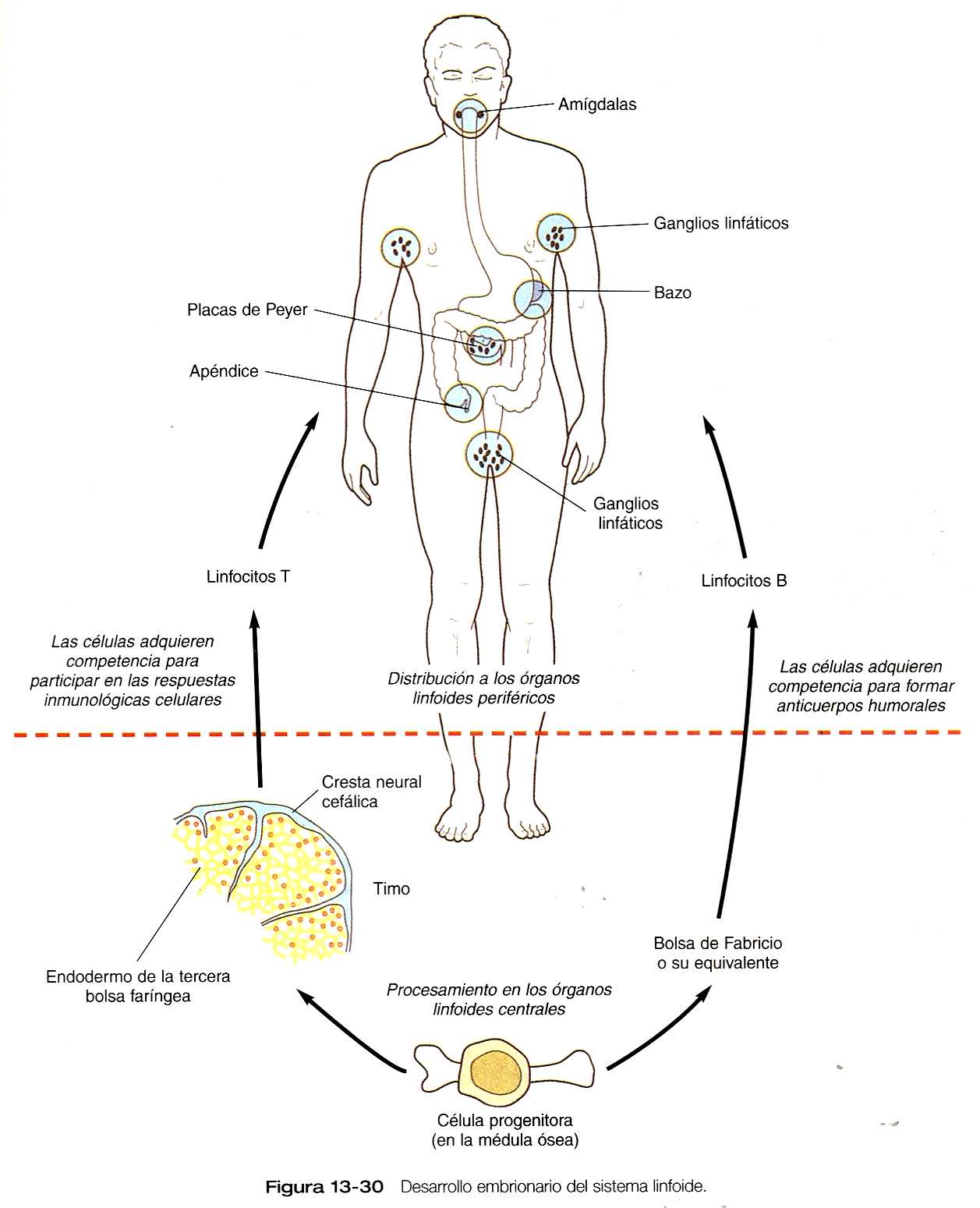
El par de primordios tìmicos endodérmicos comienza a migrar durante la sexta semana desde su origen en la bolsa faríngea. Sus vías migratorias las llevan a través de un sustrato de células mesenquimatosas hasta que alcanzan el área del futuro mediastino dorsal al esternón. Hacia el final de su migración, los dos lóbulos tìmicos que se encuentran en estrecha aposición son todavía estructuras epiteliales; no obstante, muy pronto las cubre una cápsula de tejido conectivo derivado de la cresta neural, que también forma tabiques entre los cordones epiteliales endodérmicos. Si falta la cresta neural, el timo no se desarrolla. Así pues, una interacción entre los componentes de la cresta neural y los endodérmicos del primordio tímico condiciona a este último para la posterior diferenciación de su estructura y su función.
Aproximadamente en la novena y décima semanas de gestación, los precursores de los timocitos (protimocitos) que se transportan por la sangre y que se originan en el tejido hematopoyético, comienzan a invadir el timo epitelial, se supone que como respuesta a la secreción de pépticos por el. En la glándula los protimocitos desplazan a un lado a las células epiteliales, que forman un (retículo epitelial) esponjoso. Bajo la influencia del epitelio tímico los protimocitos proliferan y se redistribuyen, formando las zonas cortical y medular del timo. Hacia la semana 14 o 15, penetran en los vasos sanguíneos y una semana mas tarde se producen cúmulos de células epiteliales a los corpúsculos de Hassal, pequeños y esféricos. En este momento la organización general del timo es la misma que la del adulto. Desde el punto de vista funcional, la acción de varias hormonas timicas hacen que el timo condicione o instruya a los protimocitos que emigran hacia su interior para que se conviertan en miembros competentes de la familia de los linfocitos T, los cuales a su vez dejan el timo y van a poblar otros órganos linfoides (por ejemplo, ganglios linfáticos, bazo) ya como células inmunológicas totalmente funcionales.
Los linfocitos T están implicados ante todo en las respuestas de la inmunidad mediada por células. Otra población de linfocitos que también se origina en la medula ósea recibe las instrucciones necesarias para convertirse en linfocitos B, son los medidores de las respuestas inmunológicas humorales. Los precursores de los linfocitos B (células preB) también deben acondicionarse para adquirir su función total, pero este fenómeno no se produce en el timo. En las aves, las células preB pasan por un órgano linfoide cloacal llamado bolsa de Fabricio donde se condicionan. Los seres humanos no poseemos la bolsa, pero se presume que su equivalente funcional existe, aunque aún no se ha identificado. Se piensa que el acondicionamiento de los linfocitos B tiene lugar en la medula ósea; en los embriones jóvenes es posible que se produzca en el hígado.
El timo y la bolsa o su equivalente en los mamíferos suelen denominarse órganos linfoides centrales. Las estructuras linfoides que son invadidas por linfocitos T y B se denominan órganos linfoides periféricos (la figura13_30 muestra el desarrollo y la función del sistema linfoide.
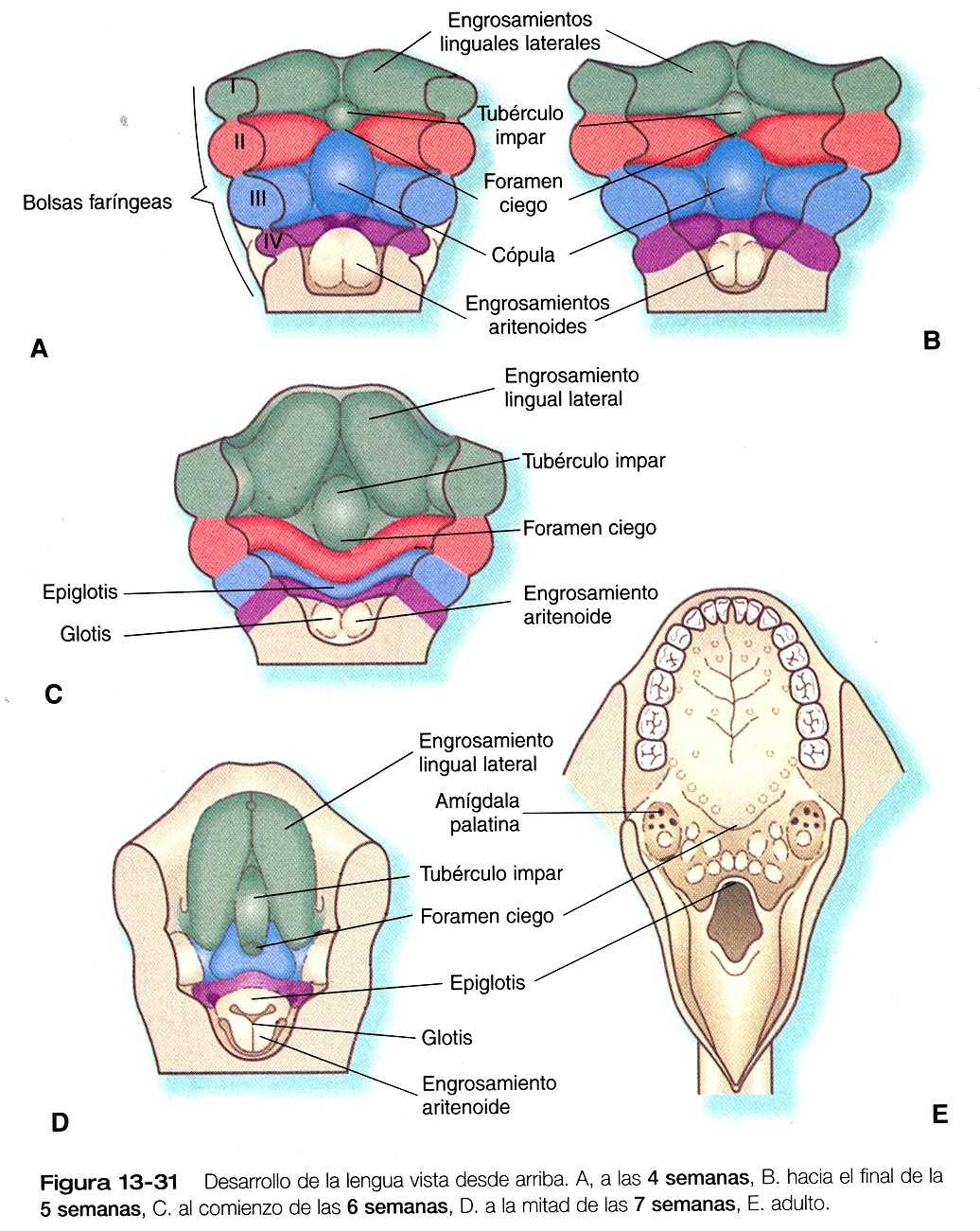
Formación de la lengua
La lengua comienza a desarrollarse a partir de una serie de engrosamientos ventrales situadas en el suelo de la faringe, mas o menos al mismo tiempo que se forma el paladar en la boca. Los tejidos de la lengua se desplazan mucho, lo que dificulta la comprensión de las características morfológicas del adulto si no se conocen los elementos básicos de su desarrollo embrionario.
En los embriones de cinco semanas de vida, la lengua esta representada por un par de engrosamientos linguales laterales situadas en las regiones ventrales de los primeros arcos branquial y dos
engrosamientos ventrales impares. El tubérculo impar esta localizado entre el primero y el segundo arcos, y la cópula (yugo) une el segundo arco con el tercero (figura 13_31A y B). El foramen ciego, que marca la localización original del primordio tiroideo sirve como puente que define el limite entre el tubérculo impar original y la copula.
En situación caudal a la copula se encuentra otro engrosamiento que representa la epiglotis.
El crecimiento del cuerpo de la lengua se logra mediante una gran expansión de las tumefacciones linguales laterales, con una pequeña contribución del tubérculo impar (figuras 13_31 C y D). La raíz de la lengua se deriva de la cópula junto con tejido ventromedial adicional situado entre el tercero y el cuarto arcos branquiales.
Debido a su inervación por el nervio hipogloso (XII par), se supone que la musculatura de la lengua proviene de los miotomas occipitales (prostóticos) y que ha migrado y recorrido una considerable distancia hacia los engrosamientos linguales, sin perder su inervación original. La inervación sensitiva general de la lengua refleja con precisión los orígenes del arco faríngeo del epitelio. Así, el epitelio lingual que cubre al cuerpo de la lengua está inervado por el trigémino (V par) en concordancia con el origen de las tumefacciones laterales en el primer arco. La raíz de la lengua también está inervada por el nervio glosofaríngeo (IX par- tercer arco) y por el vago (X par- cuarto arco). El epitelio del tercer arco sobrepasa el del segundo; por tanto, el VII par no da inervación sensitiva a la lengua.
Los pares VII (facial) y IX inervan las papilas gustativas. La contribución del VII par se ve facilitada por su rama, la cuera del tímpano, que se une a la rama lingual del trigémino y así llega al cuerpo de la lengua. Las papilas gustativas, que aparecen durante la séptima semana de la gestación, provienen de una interacción entre el epitelio lingual y las fibras aferentes viscerales especiales de los pares VII y IX. Hay evidencias que indican que el feto tiene sentido del gusto, y se ha sugerido que lo usa para controlar ambiente intramniótico.
CASO CLÍNICO
Una mujer de 22 años que se veía muy nerviosa acudió a su médico quejándose de que sentía mucho calor, había perdido mucho peso y estaba sudando más de lo acostumbrado. Al examen físico la piel estaba caliente y de textura delgada, se percibía un temblor fino en los dedos y los ojos parecían ligeramente abultados. Como parte del seguimiento diagnóstico se le dio un dosis de yodo radioactivo; luego se le hizo una gammagrafía para localizarlo. Las imágenes mostraron que la mayor parte del yodo radioactivo se concentró en una pequeña masa de tejido localizada en la base de la lengua. Se le diagnosticó una neoplasia en la masa de tejido, pero la localización de la misma se atribuyó a una malformación congénita.
Explique la base embriológica de la localización de la neoplasia.
· La región craneofacial primitiva surge de las porciones rostrales del tubo neural, la notocorda y la faringe, que está rodeada por una serie de pares de arcos aórticos. Entre los arcos aórticos y el ectodermo que los cubre se encuentran grandes masas de mesénquima derivado de la cresta neural y del mesodermo. Varios de estos componentes muestran indicios de segmentación anatómica o patrones segmentarios de expresión genética.
· Las migraciones masivas de los grupos segmentarios de células de la cresta neural suministran el mesénquima de gran parte de la región facial. La musculatura de la región craneofacial se deriva del mesodermo somitomérico o de los somitas occipitales. El componente de tejido conectivo de la musculatura facial se origina en la cresta neural.
· La región faríngea (branquial) se organiza alrededor de pares de arcos branquiales mesenquimatosos que alternan con bolsas faríngeas tapizadas de endodermo y hendiduras branquiales tapizadas de ectodermo.
· La cara y la mandíbula inferior se forman a partir de una prominencia frontonasal impar y de los procesos pares nasomediales, maxilares y mandibulares. Por medio de crecimiento y de la fusión diferencial, los procesos nasomediales forman el maxilar y el labio superiores, y la prominencia frontonasal forma la parte superior de la cara. Los procesos mandibulares al crecer se fusionan para formar la mandíbula y el labio inferiores. Un surco nasolacrimal situado entre los procesos nasolaterales y maxilares termina convirtiéndose en un canal que da lugar al conducto nasolacrimal que conecta la órbita con la cavidad nasal.
· El paladar surge de la fusión de un proceso palatino medial impar y los procesos palatinos laterales pareados. El primero forma el paladar primario y los segundos el secundario.
· El aparato olfatorio comienza como un par de engrosamientos ectodérmicos que forman la plácodas nasales. A medida que estas se hunden para formar las fóveas nasales, son rodeadas por los procesos nasomedial y nasolaterales que tienen forma de herradura. El primero da lugar al puente y al tabique de la nariz, y los segundos, a las alas de la nariz. Las fóveas nasales al profundizarse se introducen en la cavidad oral primitiva y solo más adelante las cavidades nasal y oral son separadas por el paladar.
· Las glándulas salivales se originan como engrosamientos epiteliales del epitelio oral. A través de una serie de interacciones continuas con el mesénquima circundante, el epitelio glandular se invagina, crece, se ramifica y se diferencia.
· Los dientes se forman gracias a la interacción entre el ectodermo oral (lámina dental) y el mesénquima de la cresta neural. Un diente en desarrollo primero es una yema dental, que pasa por las etapas de copa y campana. Al final de la etapa de campana, las células ectodérmicas (ameloblastos) del órgano epitelial del esmalte comienzan a producir esmalte y el epitelio derivado de la cresta neural (odontoblastos) empieza a producir dentina. Los precursores de los dientes permanentes se forman a través de primordios situados a lo largo de los dientes primarios.
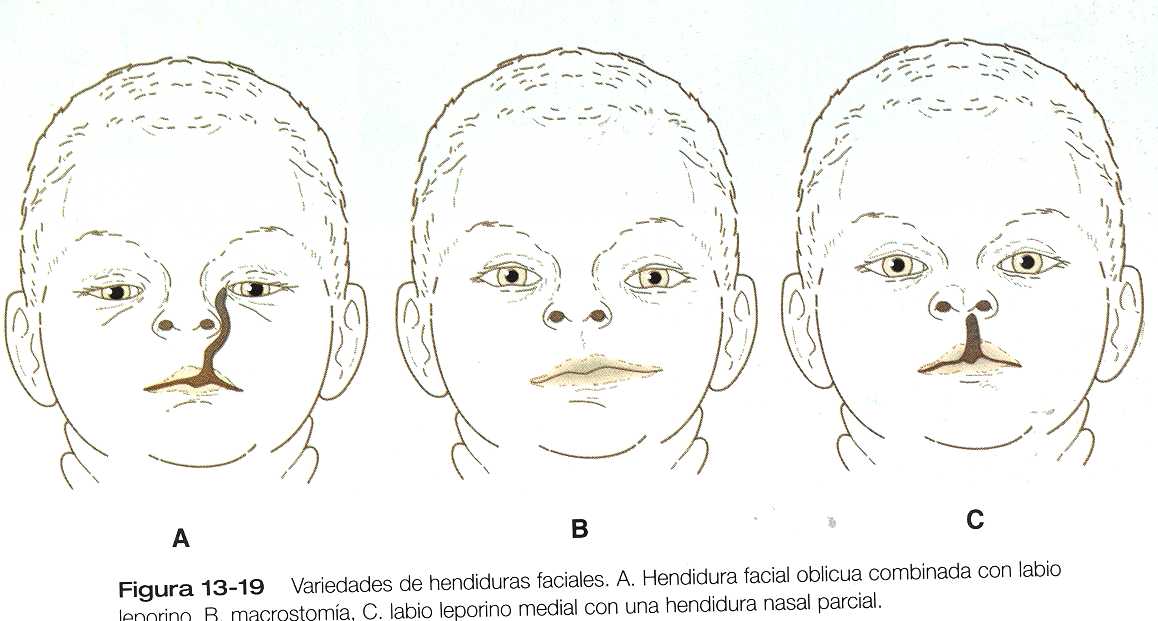
· Las malformaciones de la cara son relativamente frecuentes. Muchas. Como el labio leporino y el paladar hendido, representan la persistencia o la detención de la morfología estructural que es normal para las etapas embrionarias más incipientes. Otras, como la holoprosencefalia y el hipertelorismo, resultan de los problemas de crecimiento del proceso frontonasal. La mayoría de las malformaciones faciales parecen ser de origen multifactorial, e implican tanto susceptibilidad genética como causas ambientales.
· Los componentes de la faringe (surcos, arcos y bolsas) dan lugar a una gran diversidad de estructuras. Del primer arco se derivan el maxilar y la mandíbula y sus estructuras asociadas. El primer surco y la primera bolsa, junto con el mesénquima asociado del primero y del segundo arcos, forman las numerosas estructuras del oído medio y el externo. La segunda, tercera y cuarta hendidura branquial se obliteran y dan origen a la superficie externa del cuello, y los componentes de los arcos segundo a cuarto, forman el esqueleto faríngeo y gran parte de la musculatura y del tejido conectivo de la región faríngea del cuello. El endodermo de las bolsas tercera y cuarta forma el timo y las glándulas paratiroides. La glándula tiroides se origina en un engrosamiento ventral impar de la faringe superior.
· La lengua se deriva de múltiples engrosamientos ventrales ubicadas en el suelo de la faringe. La masa de la lengua procede de las tumefacciones linguales pares laterales situadas en la región de los primeros arcos branquiales. El tubérculo impar y la copula también contribuyen a la formación de la lengua. La musculatura de este órgano, el nervio hipogloso (XII par craneal), inerva los músculos, procede de los somitas occipitales. La inervación sensitiva general de la lengua (de los pares craneales V, IX y X) coincide con el origen embriológico de la parte inervada por cada nervio. Los pares craneales VII y IX inervan las papilas gustativas de la lengua.
· Muchas malformaciones de la región inferior de la cara y de la mandíbula están relacionadas con hipoplasia de los primeros arcos faringeos. Por lo general, los quistes, los senos y las fístulas del cuello se debe a la persistencia anormal de los surcos o las bolsas faríngeas. El tejido glandular ectópico (tiroideo, tímico o paratifoideo) se explica por la persistencia de restos de tejido a lo largo del trayecto migratorio de las glándulas. Algunos síndromes (por ejemplo, el de DiGeorge) que al parecer afectan a órganos disímiles, pueden atribuirse a defectos de la cresta neural.
PREGUNTAS DE REPASO
1. ¿El nervio facial (VII par) inerva a los músculos derivados del arco branquial?
A. Primero
B. Segundo
C. Tercero
D. Cuarto
E. Sexto
2. El labio leporino es el resultado de la falta de fusión de:
A. Los procesos nasomediales y los nasolaterales.
B. Los procesos nasomediales y los maxilares.
C. Los procesos nasolaterales y los maxilares.
D. Los procesos nasolaterales y los mandibulares.
E. Los procesos nasomediales y los mandibulares.
3. En casos de holoprosencefalia, los defectos de las estructuras faciales suelen ser secundarios a defectos de:
A. La faringe.
B. La cavidad oral.
C. El prosencéfalo.
D. Los ojos.
E. El rombencéfalo
4. El cartílago de Meckel es una estructura importante en la formación inicial de:
A. El maxilar.
B. El paladar duro.
C. El tabique nasal.
D. El paladar blando.
E. La mandíbula
5. Una inducción inicial en el desarrollo de los dientes, consiste en que el ectodermo del epitelio dental actúa sobre el mesénquima subyacente de la cresta neural. ¿Cual de las siguientes moléculas es un mediador importante del estimulo inductivo?
A. BMP-4.
B. Tenascina.
C. Hoxb-13
D. Msx-1
E. Sindecán
6. Un muchacho de 15 años con acné leve presentó una masa dolorosa a lo largo del borde anterior de esternocleidomastoideo. ¿Qué afección embriológica estaría incluida en el diagnóstico diferencial?
7. El médico del paciente anterior determino que este tenía un quiste congénito que debía extirparse quirúrgicamente ¿Qué debería tener el cirujano en cuenta durante la cirugía?
8. ¿Por qué a veces cuando una persona llora tiene aumento de la secreción nasal?
9. Una mujer que tomó un medicamento anticonvulsionante durante la décima semana del embarazo dio a luz a un bebe con labio leporino bilateral y paladar hendido. Demandó al médico, a quien culpo de las malformaciones faciales causadas por el medicamento. A usted lo llaman como testigo perito para le defensa. ¿Cuál seria la base para su caso?
10. Una mujer que en promedio tomaba tres bebidas alcohólicas al día durante el embarazo dio a luz a un bebé con retraso mental leve y una pequeña muesca en un labio superior fino y vuelto hacia arriba, amén de una reducción se su capacidad olfativa. ¿Cuál es la base de este grupo de defectos?
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}