Capítulo 11.
CRESTA NEURAL
ORÍGENES DE LA CRESTA NEURAL
MIGRACIONES DE LA CRESTA NEURAL
DIFERENCIACIÓN DE LAS CÉLULAS DE LA CRESTA NEURAL
Tabla 11-1 Principales derivados de la cresta neural.
Tabla 11-2 Factores ambientales que favorecen la diferenciación de las células de la cresta neural
Cuadro 11-1 Principales neurocristopatías
CORRELACION CLINICA 11_1
Neurocristopatias
PREGUNTAS DE REPASO
Aunque se sabe de su existencia desde hace más de un siglo, la cresta neural sólo se convirtió en uno de los componentes más estudiados del embrión de los vertebrados cuando se desarrollaron métodos adecuados para marcar sus células –primero con marcadores isotópicos y después con marcadores biológicos estables, anticuerpos monoclonales, marcadores intracelulares y marcadores genéticos-. La mayoría de los estudios sobre la cresta neural se han realizado en embriones de aves debido a su fácil acceso y a la disponibilidad de marcadores específicos. Las investigaciones en embriones de mamíferos sugieren que, excepto por ciertos detalles estructurales relativamente menores, la información obtenida en las aves puede aplicarse en forma directa a los embriones de mamíferos. Las malformaciones congénitas de la cresta neural se presentan en la correlación clínica 11-1 que aparece al final de este capítulo.
ORÍGENES DE LA CRESTA NEURAL
La cresta neural se origina en las células localizadas a lo largo de los bordes laterales de la placa neural. Las células de la cresta neural se determinan como resultado no neutral (posiblemente mediada por la proteína 4 morfogenética ósea [BMP-R] sobre las células laterales de la placa neural las células inducidas de la cresta neural expresan el gen slug, un factor de trascripción de la familia de dedo de cinc, que caracteriza a las células que se desprenden de la capa epitelial embrionaria y que a continuación migran como células mesenquimatosas.
Las células de la cresta neural se desprenden de la placa neural o del tubo neural después de modificar su forma y sus propiedades originales de células neuroepiteliales y adoptar las de las células mesenquimatosas.
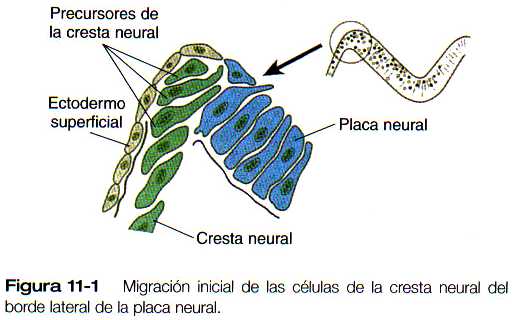
En la región cefálica, la células iniciales de la cresta neural emiten prolongaciones que penetran en la lámina basal que subyace al neuroepitelio mucho antes de que se cierre el tubo neural (figura 11-1). Después de que la lámina basal se disocia un poco más, las células de la cresta neural, que ya han adoptado una morfología mesenquimatosa, atraviesan los restos de la lámina basal y empieza una gran cantidad de migraciones.
Otro cambio importante durante la transformación de las células de la cresta neural de epiteliales a mesenquimatosas es la pérdida de la adhesividad entre las células. Este fenómeno se acompaña de la pérdida de las moléculas de adhesión celular (CAM), características del tubo neural (por ejemplo, ·N-CAM y N-cadherina), por las células de la cresta neural durante su fase migratoria. Después de que estas célula han terminado su migración y se han diferenciado ciertas estructuras (como los ganglios espinales), las CAM por lo general vuelven a expresarse.
En el tronco, las células de la cresta neural sólo abandonan el neuroepitelio después de que se ha formado el tubo neural. Sin embargo, no tienen que atravesar la lámina basal porque la parte dorsal del tubo neural carece de esta estructura hasta después de la migración de las células de la cresta neural.
MIGRACIONES DE LA CRESTA NEURAL
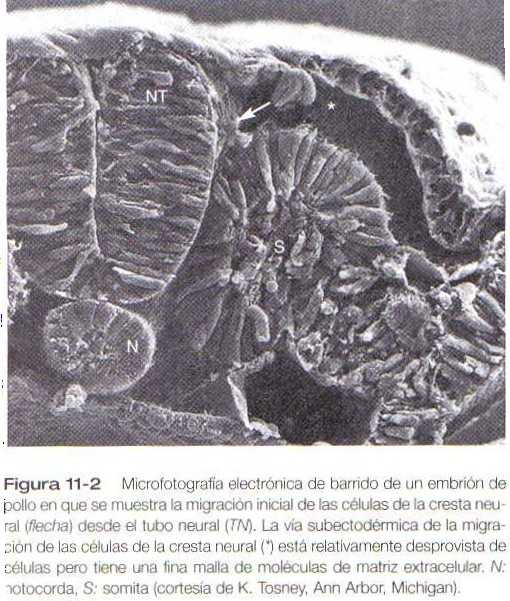
Después de abandonar el neuroepitelio, las células de la cresta neural se encuentran en un ambiente relativamente desprovisto de células y rico en moléculas de la matriz extracelular (figura 11-2). En este ambiente realizan grandes migraciones por varias vías bien definidas; estas migraciones están determinadas tanto por las propiedades intrínsecas de las células de la cresta neural como por las características del ambiente extracelular en el cual están las células en migración.
Debido a la falta de marcadores apropiados, todavía no se han definido bien los mapas de las vías migratorias de las células de la cresta neural en los mamíferos. Sin embargo, la información disponible sugiere que podrían ser similares a las del pollo. La disponibilidad de marcadores celulares estables y anticuerpos monoclonales ha convertido a los embriones quimera pollo/codorniz en la base de los estudios de las investigaciones sobre las migraciones y de la diferenciación de las células de la cresta neural.
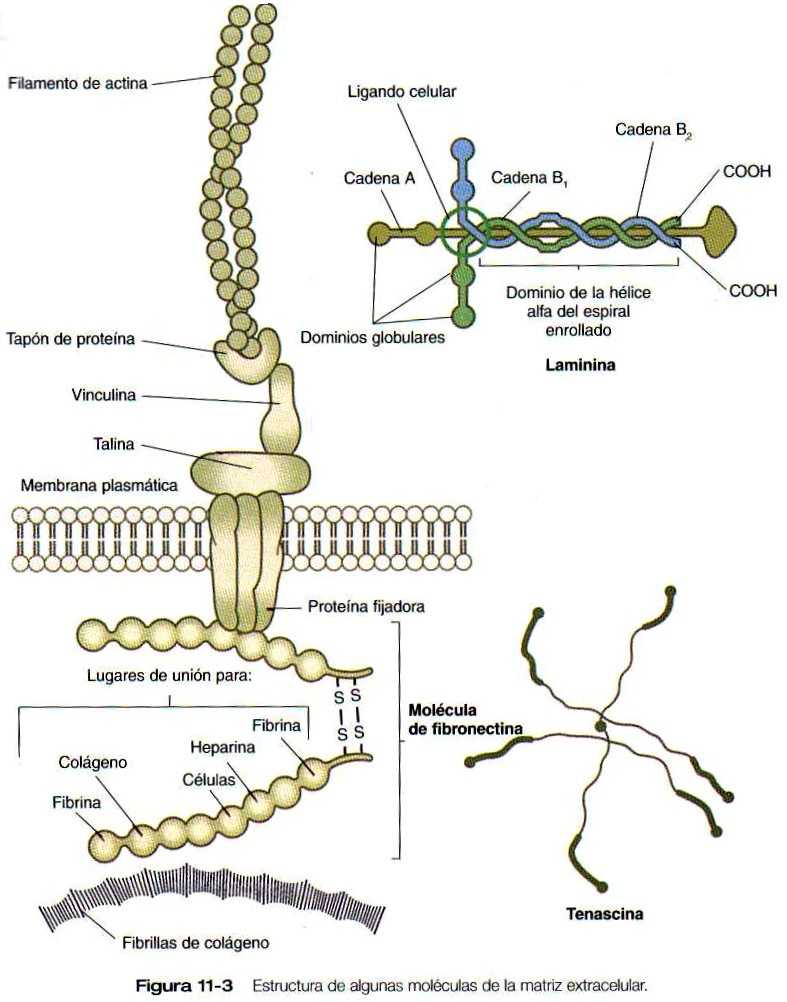
Sólo pueden hacerse consideraciones generales sobre los controles específicos que dominan las migraciones de las células de la cresta neural. Los factores permisivos son los sustratos que contienen fibronectina, laminina y colágeno de tipo IV (figura 11-3). La fijación de estas moléculas de sustrato y la migración a través de ellas está mediada por la familia de las proteínas de fijación llamadas integrinas. Las láminas basales estructuradas (por ejemplo, la del ectodermo superficial y las de tubo neural ventral) actúan como barreras que guían a las células de la cresta neural a lo largo de sus superficies. Otras moléculas de la matriz extracelular, en especial los proteoglicanos ricos en condroitín sulfato, no son buenos sustratos para la migración de las células de la cresta neural; se ven muy pocas de ellas en las áreas cuya concentración de condroitín sulfato es alta, (por ejemplo, en el esclerotoma de los somitas). La distribución de las células migratorias de la cresta neural en los somitas ilustra bien este principio. Las células de la cresta neural sólo entran en la parte anterior de los somitas, donde muchas de ellas se detienen para formar los ganglios sensitivos; no penetran en las regiones posteriores de las somitas, ya que tienen altas concentraciones de condroitín sulfato. Las células de la cresta neural del tronco y la cabeza siguen diferentes vías migratorias,
DIFERENCIACIÓN DE LAS CÉLULAS DE LA CRESTA NEURAL
Las células de la cresta neural acaban por diferenciarse en una gran variedad de estructuras adultas (tabla 11-1); uno de los principales interrogantes en la biología de la cresta neural es: ¿que controla tal diferenciación ?. A este respecto, se han propuesto dos hipótesis; según una de ellas, todas las células de la cresta neural son iguales en cuanto a su potencial de desarrollo, y su diferenciación final depende por completo del ambiente por el cual migran y en el cual terminan estableciéndose. La otra hipótesis sugiere que antes de migrar las células de la cresta neural ya están programadas para alcanzar los distintos destinos de su ulterior desarrollo y que algunas células progenitoras son favorecidas en tanto que a otras se les inhibe de su ulterior desarrollo durante la migración. Las investigaciones más recientes indican que la respuesta real puede encontrarse en algún punto entre las dos posiciones.
Cada vez hay más evidencias que muestran una correlación entre el momento de inicio de la migración de las células de la cresta neural del tubo neural y su desarrollo potencial. Por ejemplo, las células que primero comienzan a migrar tienen el potencial de diferenciarse en muchos tipos diversos de células. En cambio, las que comienzan su migración más tarde sólo pueden dar lugar a derivados característicos de localizaciones más dorsales (por ejemplo, los ganglios espinales) pero no neuronas simpáticas ni células de la médula suprarrenal. Las últimas en salir del tubo neural están restringidas a la vía más dorsal de migración y sólo pueden formar células pigmentarias.
Varios experimentos han mostrado que los destinos de las células de la cresta neural no están vinculados de forma irreversible a una sola vía. Un tipo de experimentos es el trasplante de célula de la cresta neural de una parte del cuerpo a otra. Por ejemplo, muchas células de la cresta neural del tronco se convierten en neuronas simpáticas que utilizan la norepinefrina como neurotransmisor. No obstante, en la región craneal, las células de la cresta dan lugar a neuronas parasimpáticos que producen acetilcolina. Si se trasplantan a la cabeza las células de la cresta neural torácica, algunas de ellas se transforman en neuronas parasimpáticos colinérgicas en vez de en las simpáticas adrenérgicas que se producen en la región torácica. Por su parte, las células de la región craneal de la cresta neural trasplantadas a la región torácica responden a su nuevo ambiente transformándose en neuronas simpáticas adrenérgicas. Un ejemplo mas impresionante es la conversión de las células del mesénquima periocular derivadas de la cresta neural, que en las aves normalmente formarían cartílago, en neuronas si se asocian in vitro con tejido del intestino posterior embrionario. Hoy se sabe que muchas de las influencias regionales que actúan en la diferenciación de las poblaciones locales de células de la cresta neural se deben a interacciones entre las células migratorias de la cresta neural y los tejidos específicos con que se encuentran durante su trayecto. En la tabla 11-2 se presentan algunos ejemplos de interacciones titulares que favorecen la diferenciación de derivados específicos de la cresta neural.
La plasticidad de la diferenciación de las células de la cresta neural puede demostrarse si se clonan en cultivo sólo células de la cresta neural. En el mismo medio y aparentemente bajo las mismas condiciones ambientales, la progenie de las células clonadas a menudo se diferencian en fenotipos neuronales y no neuronales (por ejemplo, células pigmentarias). Así mismo, si a las células individuales de la cresta neural in vivo se les inyecta un colorante, más del 50% de las que han recibido la inyección dan origen a una progenie de dos a cuatro fenotipos diferentes que contiene dicha sustancia. Si se cultivan in vitro células precursoras de la cresta neural que han sido clonadas en unas condiciones ambientales específicas, es posibles comenzar a comprender los mecanismos que pueden determinar el fenotipo in vivo. Por ejemplo, en un experimento las células de la cresta neural de ratas que se cultivaron in vitro en condiciones estándar se transformaron en neuronas, pero cuando se les añadió factor β de crecimiento glial se diferenciaron en células de Schwann porque ese factor suprimió su tendencia a convertirse en neuronas. De igual manera, los factores de crecimiento BMP-2 y BMP-4 hacen que las células de la cresta neural en cultivo se conviertan en neuronas autonómicas, en tanto que la exposición de estas células al factor β de transformación del crecimiento hace que se diferencien en músculo liso.
No todos los tipos de transformaciones pueden producirse entre los posibles derivados de la cresta neural. Por ejemplo, las células de la cresta del tronco trasplantadas a la cabeza no pueden formar cartílago ni elementos esqueléticos, aunque esto sea lo normal para las células de la cresta neural craneal. La mayoría de los experimentos sugiere que las células primitivas de la cresta neural pueden segregarse en linajes intermedios que conservan la posibilidad de diferenciarse en varios pero no en todos, los tipos de fenotipos individuales. En el embrión de pollo, algunas células de la cresta neural son antigénicamente diferentes de otras, incluso antes de haber dejado el tubo neural.
Algunas células de la cresta neural son bipotenciales y dependiendo de las señales que reciben de su ambiente local adquieren su diferenciación final. Un línea llamada linaje simpatoadrenal forma las células de la médula suprarrenal si se exponen a los glucocorticoides suprarrenales. En contraste, si antes se exponen al factor de crecimiento de los fibroblastos (FGF) y después al factor de crecimiento de los nervios, las mismas células se convierten en neuronas simpáticas. Así mismo, las células cardíacas cultivadas segregan una proteína que convierte las neuronas simpáticas posmitóticas de fenotipo adrenérgico (cuyo neurotransmisor es la norepinefrina) en neuronas de fenotipo colinérgico (cuyo neurotransmisor es acetilcolina), como se ve en la figura 10-20. Durante el desarrollo normal, las neuronas simpáticas que inervan glándulas sudoríparas son catecolaminérgicas hasta que sus axones establecen contacto efectivo con la glándula; en ese momento se vuelven colinérgicas. Con tan amplia diversidad de derivados de la cresta neural, es probable que durante el desarrollo se descubran otros cambios de un tipo funcional de células en otro.
Tabla 11-1 Principales derivados de la cresta neural
Cresta del tronco Cresta craneal
Sistema nervioso Ganglios espinales Ganglios del trigémino (V),
sensitivo el facial (VII), el glosofa-
ríngeo (ganglio superior)
(IX) y el vago (ganglio yu-
gular) (X)
Sistema nervioso Ganglios de la cresta Ganglios parasimpáticos:
autónomo simpática, ganglios ciliar,etmoidal, esfenopalati-
colaterales: celíaco y no, submandibular, visceral
mesentérico. Células satélitales de los
Ganglios parasimpáticos: ganglios sensitivos, células
Plexo pélvico, viscerales. de Schwann de los nervios
Células satélitales de los periféricos, leptomeringes
Ganglios sensitivos, cè- del prosencéfalo y parte del
lulas de Schwann de los mesencéfalo
nervios periféricos, célu-
las gliales entéricas.
Células pignen- Melanocitos Melanocitos
tarias.
Células endocri- Médula suprarrenal, célu- Cuerpo carotídeo (células
nas y paraendo- las neurosecretoras del de tipo I), células parafo-
crinas corazón y de los pulmones liculares (tiroides)
Células mesodér- Ninguna Bóveda craneal (escama y
micas. parte del frontal), nasal y
Esqueleto orbital, cápsula óptica
(parte), paladar y maxilar,
esfenoides (pequeña con-
tribución), trabéculas
(parte), cartílagos visce-
rales, cartílago del oído
externo (parte)
Tejido conectivo Ninguna Dermis y grasa cutánea;
córnea (fibroblastos del
estroma y endotelio corne-
al); papilas dentales (odon-
toblastos); tejido conectivo
del estroma glandular de:
tiroides, paratiroides, timo,
salívales, lacrimales; tracto
de salida del corazón (re-
gión troncoconal); válvulas
semilunares cardíacas; pa-
redes de la aorta y arterias
derivadas del cayado aór-
tico
Músculo Ninguna Músculos filiares; músculo
liso de la dermis, músculo
liso vascular, elementos
accesorios del músculo
esquelético
.
Tabla 11-2 Factores ambientales que favorecen la diferenciación de
las células de la cresta neural
Derivados de la cresta Estructura con la cual
neural interactúan
Huesos de la bóveda craneal Encéfalo
Huesos de la base del cráneo Notocorda, encéfalo
Cartílagos de los arcos bran- Endodermo faríngeo
quiales
Cartílago de Meckel Ectodermo craneal
Maxilar Ectodermo maxilar
Mandíbula Ectodermo mandibular
Paladar Ectodermo palatino
Cápsula ótica Vesícula ótica
Dentina Ectodermo oral
Estroma glandular de: tiroides, Epitelio local
paratiroides, timo, salivales
Células cromafines de la Glucocorticoides segregados
médula suprarrenal por la corteza suprarrenal
Neuronas entéricas Pared del intestino
Neuronas simpáticas Médula espinal, notocorda,
somitas
Neuronas sensitivas Tejido diana periférico
Células pigmentarias Matriz extracelular a lo
largo de la vía migratoria
CRESTA NEURAL DEL TRONCO
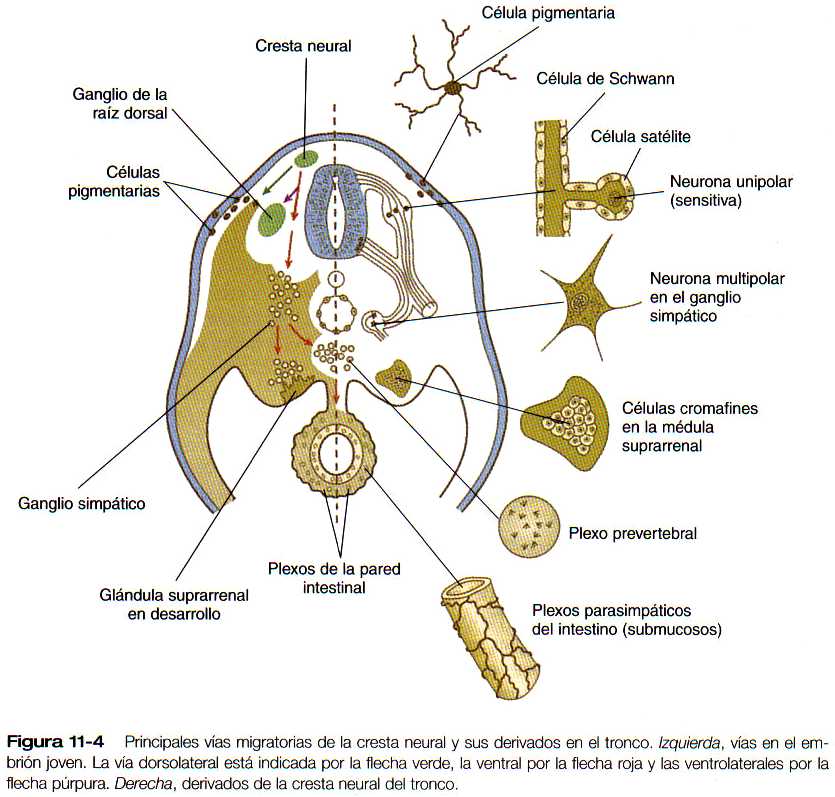
La cresta neural del tronco se extiende desde el nivel del sexto somita hasta los somitas más caudales. Si se hace un corte transversal, en la mayor parte del tronco pueden verse tres vías migratorias de las células de la cresta neural (figura 11-4). La primera es la vía dorsolateral entre el ectodermo y los somitas. Las células que eligen esta vía se distribuyen por debajo del ectodermo y al final entran en el ectodermo como células pigmentarias (melanocitos).
La segunda vía migratoria es ventral; a lo largo de ella las células de la cresta neural inicialmente se desplazan hacia el espacio que queda entre los somitas y el tubo neural. Prosigue justo por debajo de la superficie ventromedial del somita hasta llegar a la aorta dorsal. Las células que siguen este trayecto pertenecen al linaje simpatoadrenal; estas células contribuyen a la formación de la médula suprarrenal y de los elementos del sistema nervioso simpático.
Una tercera vía ventrolateral conduce hacia las mitades anteriores de los somitas. Las células que siguen esta ruta forman los ganglios sensitivos que se disponen de formas segmentarias.
El linaje simpatoadrenal deriva de una célula progenitora simpatoadrenal determinada que ya ha pasado varios puntos de restricción, de manera que ya no puede formar neuronas sensitivas, ni células gliales ni melanocitos. Esta célula da lugar a cuatro tipos de progenies celulares, 1) células cromafines suprarrenales, 2) pequeñas células intensamente fluorescentes que se encuentran en los ganglios simpáticos, 3) neuronas simpáticas adrenérgicas y 4) una pequeña población de neuronas simpáticas colinérgicas.
En un escalón más debajo de este linaje celular se encuentra la célula progenitora bipotencial que da lugar a las células cromafines suprarrenales o a neuronas simpáticas. Esta célula progenitora bipotencial tiene algunas características neuronales, pero la diferenciación final dependerá del ambiente que los rodee. En presencia del FGF y del factor de crecimiento neural en los primordios de los ganglios simpáticos, estos precursores se convierten en neuronas simpáticas definitivas. Por otra parte, las células precursoras situadas en la medula suprarrenal en desarrollo se encuentran con los glucocorticoides segregados por las células de la corteza suprarrenal. Bajo esta influencia hormonal, pierden sus propiedades neuronales y se transforman en células cromafines. Sin embargo, esta determinación de la diferenciación no queda fijada de forma definitiva, ya que después del nacimiento las células cromafines pueden ser inducidas a transdiferenciarse en neuronas si se exponen in Vitro al factor de crecimiento neural.
A lo largo de todo el intestino existe una población de neuronas parasimpáticas derivadas de la cresta neural y sus células asociadas, la glía entérica. Provienen de las células de la cresta neural de los niveles cervical (vagal) y sacro y migran por todo el intestino en desarrollo. En el tubo digestivo las células de la cresta neural forman el sistema nervioso entérico, que en muchos aspectos actúa como un componente independiente del sistema nervioso. El número de neuronas entéricas se aproxima al de neuronas de la médula espinal, y la mayoría de ellas no están conectadas directamente con el cerebro ni con la médula. Esta independencia explica por qué el intestino puede mantener su actividad refleja cuando no recibe estímulos del sistema nervioso central.
Una gran cantidad de datos sugieren que las células de la cresta neural no están comprometidas a formar el tejido nervioso asociado con el intestino antes de migrar de la médula espinal. Si la cresta vagal se remplaza por cresta neural del tronco, que por lo general no da lugar a derivados asociados con el tubo digestivo, el intestino resulta colonizado por las células transplantadas de la cresta neural del tronco. Las evidencias de que las vías de migración afectan la diferenciación se ven en los neurotransmisores que estas células transplantadas producen. Las neuronas parasimpáticas procedentes de las células del tronco producen en el intestino serotonina pero no catecolaminas. Si se hubiesen diferenciado en sus lugares normales en el tronco, estas neuronas habrían producido catecolaminas y no serotonina.
A pesar de la fuerte influencia del ambiente del intestino en la diferenciación de las células de la cresta neural expuestas a él, estas conservan un alto grado de flexibilidad en su desarrollo. Al parecer, si las células de la cresta neural que ya están en el intestino de embriones de aves se trasplantan a la región del tronco de embriones más jóvenes, pierden la memoria de los derivados de su asociación con el intestino. Entran en las vías de diferenciación común a las células de la cresta del tronco (por ejemplo, suprarrenales o nervios periféricos, pero no entran en las vías de los melanocitos) y se diferencian como si fueran derivados del tronco.
La mayor parte de los precursores de la cresta neural de las neuronas parasimpáticas asociadas con el intestino y de las neuronas parasimpáticas asociadas con el intestino y los de las neuronas simpáticas expresan el factor básico de trascripción hélice-asa-hélice Mash-1, pero no los de las neuronas sensitivas. La expresión del Mash-1 en estas células esta bajo el estimulo de los factores de crecimiento BMP-2 y BMP-4 y al parecer el Mash-1 mantiene la competencia de las células posmigratorias en el intestino para que se diferencien en neuronas. Se requieren otros factores ambientales para contemplar la determinación de estas células a formar neuronas autonómicas. Hay datos recientes de la existencia de una segunda población de aparición tardía de precursores de la cresta neural de neuronas entéricas que no expresan Mash-1.
Comparada con la cresta neural craneal, la del tronco tiene un espectro relativamente limitado de opciones de diferenciación. Sus derivados se resumen en la tabla 11-1.
CRESTA NEURAL CRANEAL
La cresta neural craneal es uno de los principales componentes del polo cefálico del embrión. Las investigaciones de anatomía y desarrollo comparado sugieren que la cresta neural craneal puede representar el principal sustrato morfológico en la evolución de la cabeza de los vertebrados. Debido a los métodos precisos de marcaje celular, la comprensión de la cresta neural craneal ha avanzado de forma notable. La mayoría de los estudios se han realizado en embriones de aves; sin embargo, los que se han hecho en mamíferos sugieren que las propiedades y el papel de la cresta neural en el desarrollo craneal de los mamíferos son bastante similares a los de las aves.
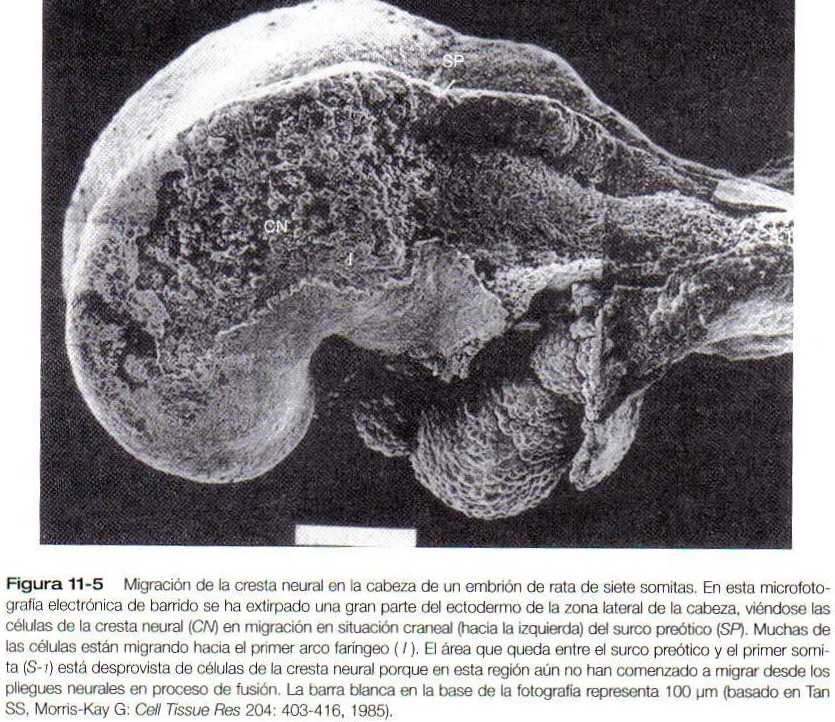
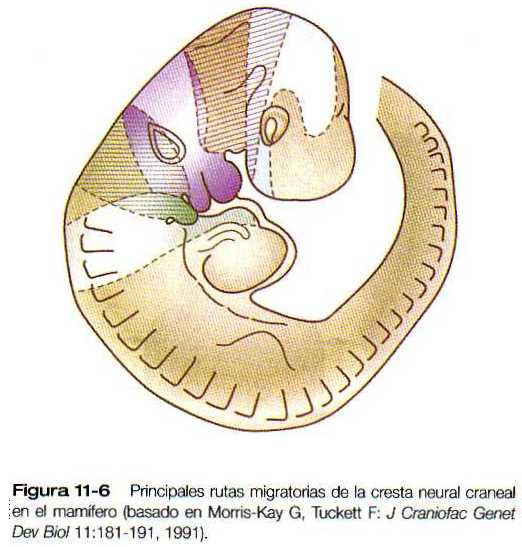
En la cabeza de los mamíferos, las células de la cresta neural abandonan al futuro encéfalo mucho antes de la fusión de los pliegues neurales (figura 11-5). Aunque las vías migratorias de las células de la cresta neural craneal en los mamíferos no están tan bien definidas como en las aves, parece que existen territorios migratorios distintos pero hasta cierto punto solapados en la cabeza del embrión de los mamíferos (figura 11-6). Debido a la distribución de las primitivas células mesenquimatosas y de las moléculas de la matriz extracelular en la cabeza del mamífero, las células de la cresta neural migran en corrientes difusas a través del mesénquima craneal para alcanzar su destino final.
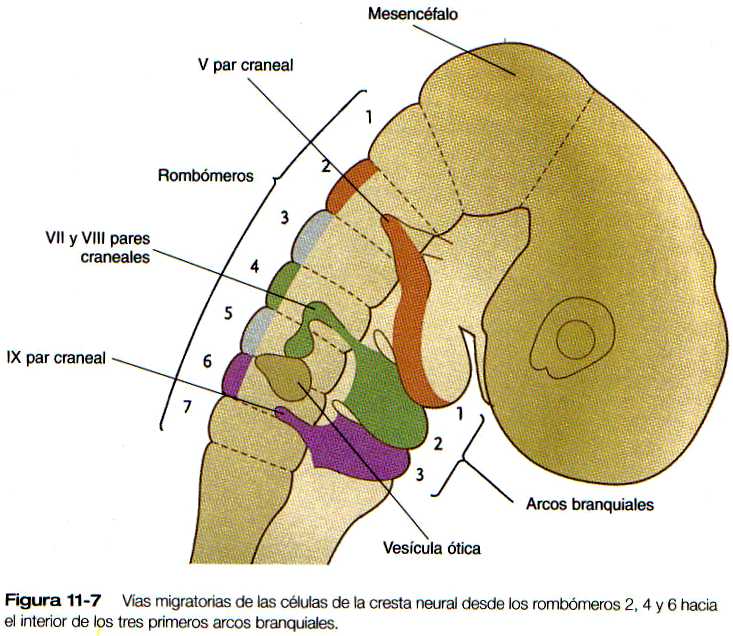
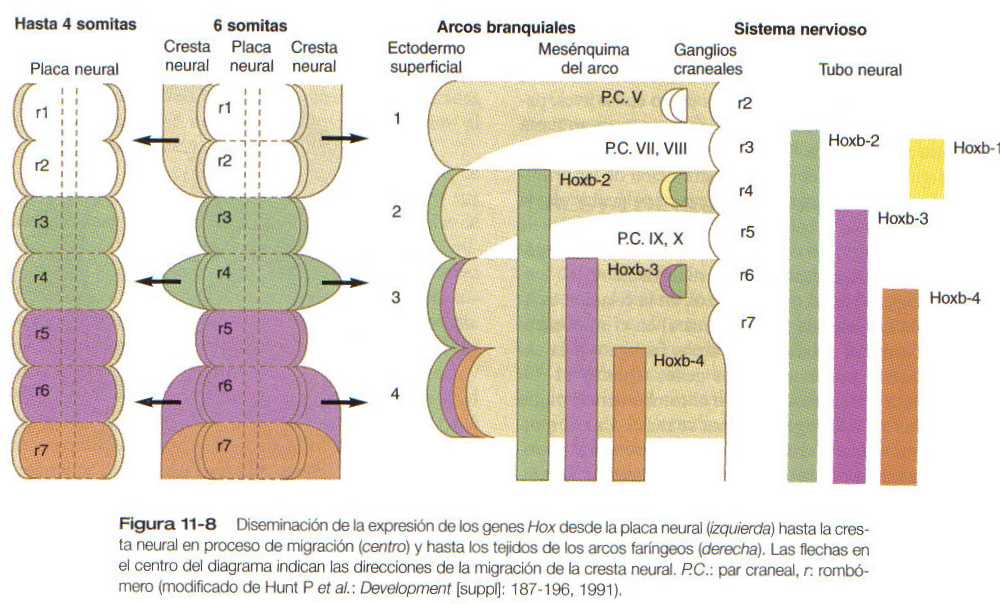
Hay una especificidad muy marcada en la relación existente entre los orígenes de la cresta neural en el rombencéfalo, su destino final dentro de los arcos faríngeos y la expresión de ciertos productos de los genes (figura 11-7 y 11-8). La cresta neural asociada con el rombómero 2 migra hacia el primer arco faríngeo y forma su masa. La del rombómero 4 va al segundo arco y la del rombómero 6 al tercero. Las células de la cresta neural no parecen migrar directamente en dirección lateral de los rombómeros 3 y 5. Según algunos investigadores estas células no se forman en los rombómeros 3 y 5, o si lo hacen sufren luego una apoptosis. Los estudios sugieren que en estos rombómeros se forman células de la cresta neural pero que, conforme salen de ellos, hacen un giro agudo y se unen a las vías migratorias de las células de la cresta neural que migran desde los rombómeros adyacentes.
Existe una estrecha correlación entre el patrón de migración de las células romboméricas de la cresta neural y la expresión de productos del complejo de los genes Hoxb. Los productos de Hoxb-2, Hoxb-3 y Hoxb-4 se expresan según una secuencia regular tanto en el tubo neural como en el mesénquima de los arcos faríngeos 2, 3 y 4 derivados de la cresta neural. El Hoxb no se expresa en el rombómero 2 ni en el mesénquima del primer arco faríngeo. Sólo después de que los arcos faríngeos se han poblado con células de la cresta neural el ectodermo que los cubre expresa un patrón similar de los productos de los genes Hoxb (figura 11-8). Estos genes Hoxb intervienen en la especificación de la posición de las células de la cresta neural con las cuales están asociados. Además, las interacciones existentes entre las células de la cresta neural y el ectodermo superficial de los arcos faríngeos pueden especificar el ectodermo de los arcos.
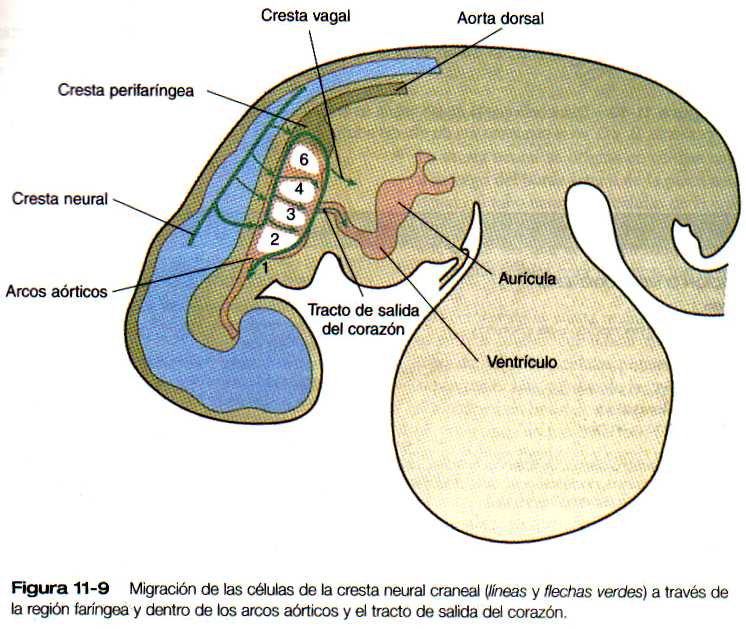
En la parte posterior de la faringe una cresta perifaríngea pasa por detrás de sexto arco faríngeo (figura 11-9); ventral a la faringe se dirige en dirección craneal, creando la vía a través de la cual pasan el nervio hipogloso (XII) y las células precursoras de músculo esquelético. Los músculos inervados por el hipogloso y los músculos hipofaríngeos son los únicos músculos esqueléticos derivados de los somitas en los que las células del tejido conectivo se originan en la cresta neural.
Una de las principales diferencias entre las células de la cresta neural craneal y las del tronco es que las primeras se crean con el patrón de instrucciones específicas de su nivel cefálico, mientras que las del tronco no parecen tener incorporadas instrucciones específicas del nivel. Concretamente, si las células de la cresta neural que en circunstancias normales dan lugar a estructuras características del primer arco faríngeo se trasplantan al nivel del tercero se forman estructuras ectópicas del primer arco (por ejemplo, una mandíbula supermuneraria) en el nivel del tercero. Trasplantes de un nivel similar de cresta neural trasplantada no dan lugar a la formación de estructuras anómalas.
Tanto en los embriones de las aves como en la de los mamíferos de la vía de migración de la cresta perifaríngea que tiene forma de S (figura 11-9) marca aproximadamente una frontera entre las estructuras craneales y las del tronco. En esta región los somitas dorsales (del tronco) y los arcos faríngeos (craneales) se superponen a lo largo del eje craneocaudal. La cresta perifaríngea marca la vía de la migración de las células entéricas de la cresta hacia el intestino y de las células cardíacas de la cresta neural hacia el tronco de salida del corazón (figura 11-9). Un trastorno en esta área puede producir defectos en la reptación cardiaca (tabique aortopulmonar) así como malformaciones glandulares y craneofaciales.
El síndrome de DiGeorge, que se asocia con una deleción del cromosoma 22, se caracteriza por hipoplasia y reducción de la función del timo, la tiroides y las paratiroides y defectos cardiovasculares, como la persistencia del tronco arterioso y anomalías de los arcos aórticos. El ratón mutante para Hoxa-3 muestra un espectro similar de defectos faríngeos. El denominador común de estas situaciones patológicas es un defecto de la cresta neural craneal que procede del tercero y el cuarto arcos faríngeos y el tracto de salida del corazón. Se han descrito defectos similares en los embriones humanos expuestos a cantidades excesivas de ácido retinoico al comienzo de la embriogénesis.
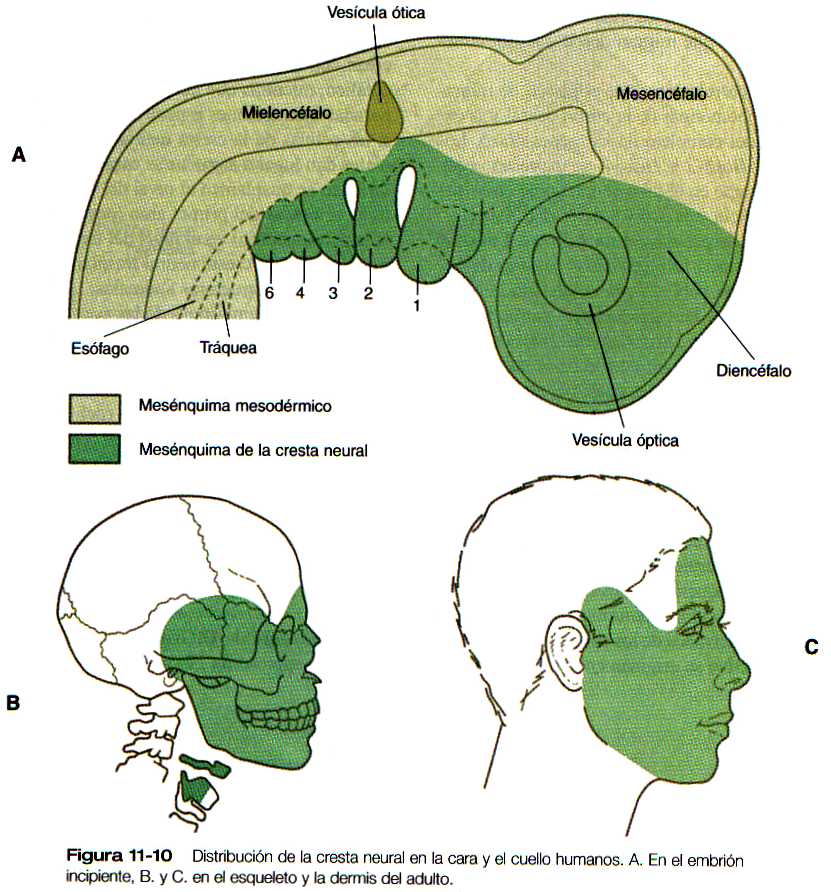
Las células de la cresta neural craneal se diferencian en una amplia diversidad de tipos celulares y tisulares (tabla 11-1), incluidos los tejidos conectivo y esquelético. Estos tejidos constituyen gran parte de los tejidos blandos y duros de la cara (figura 11-10). Los detalles específicos de la morfogénesis de la cabeza se presentan en el capítulo 13.
Cuadro 11-1 Principales neurocristopatías________________________
DEFECTOS DE MIGRACIÓN TUMORES Y DEFECTOS DE
O MORFOGÉNESIS LA PROLIFERACIÓN
Cresta neural del tronco Feocromocitoma: tumor del te-
jido cromafín de la médula su-
prarrenal
Enfermedad de Hirschusprung Neuroblastoma: tumor de la mé-
(megacolon agangliónico) dula suprarrenal, los ganglios
autónomos o ambos
Cresta neural craneal Carcinoma medular del tiroides:
tumor de células parafoliculares
Defectos de septación aorto- (secretoras de calcitonina) de
pulmonar del corazón tiroides
Defectos de la cámara ante- Tumores carcinoides: tumores de
rior del ojo las células enterocromafines del
Labio leporino, paladar hendido tracto digestivo.
o ambos Neurofibromatosis (enfermedad de
Displasia frontonasal von Recklinghausen): tumores
Síndrome de DiGeorge (hipo- de los nervios periféricos
paratiroidismo, deficiencia
tiroidea, displasia del timo DEFECTO GENÉTICO QUE
que conduce a inmunodefi- IMPLICA A LAS CÉLULAS
ciencia, defectos del tracto de DE LA CRESTA NEURAL
salida del corazón y de los
arcos aórticos) Albinismo
Algunas anomalías dentales
Cresta neural y del tronco
Asociación CHARGE
Síndrome de Waardenburg
· La cresta neural se origina en células neuroepiteliales situadas a lo largo del borde lateral de la placa neural. Una vez que han dejado el tubo neural, las células de la cresta neural migran hacia locaciones periféricas en todo el organismo. Algunos sustratos, como los que contienen moléculas de condroitín sulfato, son desfavorables para la migración de las células de la cresta neural
· Las células de la cresta neural se diferencian en muchos tipos de células adultas, como neuronas sensitivas y autonómicas, células de Schwann, células pigmentarias y células de la médula suprarrenal. Las células de la cresta neural craneal también se diferencian en hueso, cartílago, dentina, fibroblastos dérmicos, algunos tipos de músculo liso, tejido conectivo del estroma de las glándulas faríngeas y varias regiones del corazón y los grandes vasos.
· El control de la diferenciación de las células de la cresta neural es variable, ya que algunas de ellas son determinadas antes de comenzar a migrar y otras responden a claves ambientales que hay en el curso de sus trayectos migratorios. Las células de la cresta neural del tronco no pueden convertirse en elementos esqueléticos. La cresta neural se origina en células neuroepitelios situadas a lo largo del bordo lateral de la placa neural. Una vez que han dejado el tubo neural, las células de la cresta neural migran hacia locaciones periféricas en todo el organismo. Algunos sustratos como los que contienen moléculas de condroitin sulfato, son desfavorables para la migración de las células de la cresta neural.
· Las células de la cresta neural se diferencia en muchos tipos de células adultas, como neuronas sensitivas y autonómicas células de Schwann, células pigmentarias y células de la medula suprarrenal. Las células de la cresta neural, craneal también se diferencian en hueso, cartílago, dentina, fibroblastos térmicos algunos de tipos de músculo liso, tejido conectivo del estroma de las glándulas faríngeas y varias regiones del corazón y los grandes vasos.
· El control de la diferenciación de las células de la cresta neural es variable, ya que algunas de ellas son determinadas antes de comenzar a migrar y otras responden a claves ambientales que hay en el curso y sus trayectos migratorios. Las células de la cresta neural del tronco no pueden convertirse en elementos esqueléticos.
· Las células de la cresta neural del tronco siguen tres vías principales de migración: 1) La vía dorsolateral para las células pigmentarias; 2) una vía ventral para las células de linaje simpatoadrenérgico y 3) una vía ventrolateral que pasa a través de las mitades anteriores de los somitas para las células formadoras de los ganglios sensitivo. Las células de la cresta neural craneal forman muchos tejidos de la región facial. En la región faringe los trayectos migratorios de las células de la cresta guardan una estrecha correlación con regiones de expresión de productos del complejo de los genes Hoxb. Las células de la cresta craneal son moldeadas de acuerdo con instrucciones especificas, en tanto que las del la cresta al tronco no.
· Varias enfermedades y síndromes genéticos se asocian con trastornos de la cresta neural. La neurofibromatosis se caracteriza por la presencia de múltiples tumores y alteraciones de la pigmentación. Los problemas de la cresta neural cardiaca pueden dar como resultado defectos de septación y del tracto de salida del corazón.
CORRELACION CLINICA 11_1
Neurocristopatias
Debido al complejo de la cresta neural, una gran cantidad de malformaciones congénitas se asocia con defectos en su desarrollo. Suelen dividirse en dos categorías principales: los defectos de la migración o la morfogénesis y los tumores de los tejidos de la cresta neural (cuadro 11_1). Algunos de estos defectos solo implican un componente de la cresta otros afectan a varios y se reconocen como síndromes. Solo es posible entender varios síndromes de asociaciones de defectos y se reconoce la amplia distribución de los derivados de la cresta neural. Por ejemplo, una asociación que se llama Charge consiste en coloboma, cardiopatía, atresia de coanas, retraso del desarrollo, hipoplasia genital en los varones y anomalías del oído (capitulo 2). Los tipos 1 y 3 del síndrome de Waardenburg, que es causado por mutaciones del pax-3 comprende varias combinaciones de defectos de pigmentación (por lo común una franja blanca en el cabello y otras anomalías pigmentarias de la piel), sordera, paladar hendido e hipertelorismo ocular (aumento del espacio entre los ojos), el de tipo 1 se caracteriza además por la hipoplasia de los músculos de las extremidades, lo que no es sorprendente dada la importante sujeción que existe entre el pax -3 y las células miogénicas que emigran hacia las yemas de las extremidades desde los somitas. El pax -3 se expresa de forma similar en las células cardiacas procedentes de la cresta neural, pero se dirigen hacia abajo en las paredes del tracto de salida o los arcos aórticos. Los defectos cardiovasculares (xxxx) también se ven en los mutantes pax_3.
La neurofibromatosis (enfermedad de von Recklinghausen ) es una enfermedad genética común que se manifiesta por la presencia de múltiples tumores originados de la cresta neural. Las características comunes son las manchas café con leche (lesiones pigmentarias de color marrón claro) en la piel, múltiples (a menudo cientos) neurofibromas (tumores de los nervios periféricos) gigantismo ocasional de un dedo o una extremidad, y otros defectos. La neurofibromatosis afecta aproximadamente a uno de cada tres mil nacidos vivos y el gen afectado es de gran tamaño y sujeto a una elevada taza de mutaciones. Debido a la contribución se la cresta neural a la cara y a otras partes de la cabeza del cuello, varias malformaciones de la región craneofacial presentan alteraciones de los derivados de la cresta neural. Un amplio espectro y anomalías faciales agrupadas bajo el termino de displasia frontonasal (capitulo 13) implican en alto grado a los tejidos derivados de la cresta neural.
CASO CLINICO
A una recién nacida se le diagnostico una separación incompleta entre las arterias aorta y pulmonar (una forma leve del tronco arterioso persistente). Mas tarde, después de la cirugía cardiaca correctora, comenzó a sufrir mas resfriados y dolores de garganta que sus hermanos. Después de las pruebas, el medico dijo a sus padres que había evidencias de inmunodeficiencia. También les dijo que seria buena idea revisarle los niveles de hormona paratifoidea. ¿Cuál es la base de esta sugerencia?
PREGUNTAS DE REPASO
1.- ¿Cual de estos tipos celulares y titulares proceden de las células de la cresta neural del tronco pero no la del cráneo?
A. Ganglios Sensitivos
B. Medula Suprarrenal
C. Melanocitos
D. Células de Schwann
E. Ninguna de las anteriores
2.-¿Qué molécula es un mal sustrato para la migración de las células de la cresta neural?
A. Laminina
B. Condroitin Sulfato
C. Fibronectina
D. Colágeno de tipo IV
E. Ácido Hialurónico
3.- Las Células de la Cresta Neural proceden de:
A. El Somita
B. El Ectodermo Dorsal No Neural
C. El Tubo neural
D. El Mesodermo Esplácnico
E. El Endodermo del saco Vitelino
4.- Un bebe de seis meses muestra múltiples defectos congénitos, incluidos paladar hendido, sordera, hipertelorismo ocular y un mechón blanco en la frente, pero el resto del cabello es oscuro. El diagnostico más probable es:
A. Asociación CHARGE
B. Enfermedad de von Recklinghausen
C. Enfermedad de Hirschsprung
D. Síndrome de Waardenburg
E. Ninguna de las anteriores
5.- ¿Cuales de las siguientes moléculas participa en la migración de las células de la cresta neural desde el tubo neural?
A. Slug
B. BMP_2
C. Mash_1
D. Norepinefirna
E. Factor de Crecimiento Glial
6.-¿ Cual de las siguientes estructuras no son derivadas de la cresta neural?
A. Neuronas Sensitivas
B. Motoneuronas
C. Células de Schwann
D. Medula Suprarrenal
E. Papilas Dentales
7.-¿ Que mantiene la competencia de las células de la cresta neural para diferenciarse en neuronas autonómicas?
A. Sonic Hedgehog
B. Acetilcolina
C. Mash_1
D. Factor de crecimiento glial
E. Factor beta de transformación de crecimiento
8.- Si las células de la cresta neural del tronco se transplanta a la región craneal, pueden formar todos los siguientes tipos de células, excepto:
A. Células Pigmentarias
B. Células de Schwann
C. Neuronas Sensitivas
D. Cartílago
E. Neuronas autonómicas
9.-¿Cómo se produce la distribución segmentaría de los ganglios espinales?
10.-¿ Cuales son las tres principales diferencia entre la cresta neural del tronco y la del cráneo?
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}