TRANSPORTE DE LOS GAMETOS
Y FECUNDACION
Se describen tanto los orígenes como la maduración de los gametos masculinos y femeninos y las condiciones hormonales que hacen posible dicha maduración. También se describen los cambios cíclicos controlados por las hormonas, que tienen lugar en el tracto reproductor femenino y lo preparan para la fecundación y el soporte del desarrollo embrionario. Primero la forma en que el óvulo y el espermatozoide se juntan en el tracto reproductor femenino para que pueda tener lugar la fecundación. Se analiza el complejo conjunto de interacciones que intervienen en la fecundación del óvulo por el espermatozoide.
OVULACIÓN Y TRANSPORTE DEL ÓVULO
Y DEL ESPERMATOZOIDE
Ovulación
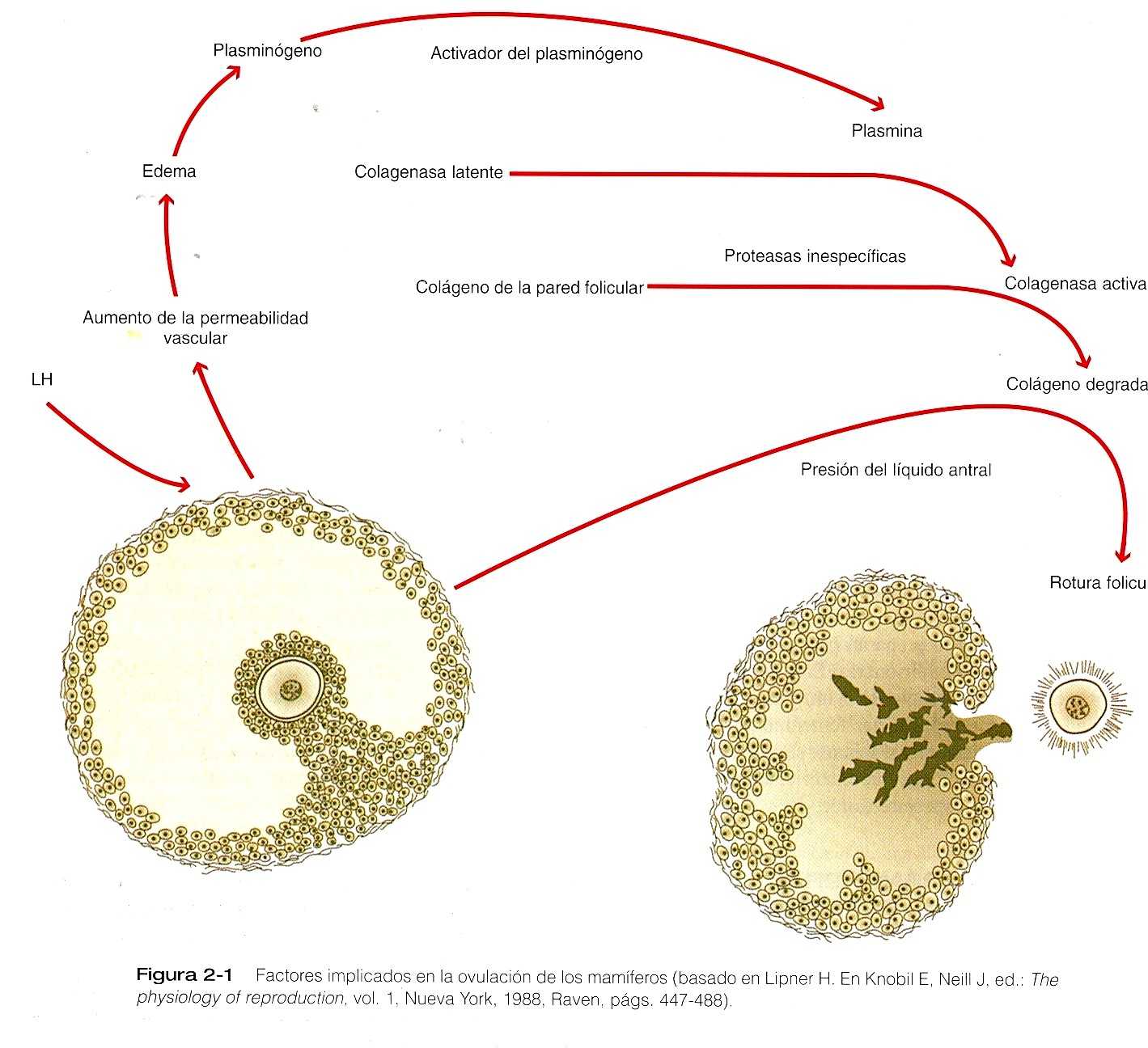
Hacia la mitad del ciclo menstrual, el folículo de Graff maduro que contiene al óvulo que está detenido en la profase de la primera división meiótica se ha desplazado hacia la superficie del ovario. Bajo la influencia de las hormonas folículo-estimulante (FSH) y luteinizante (LH), el folículo se expande en forma considerable. Al terminar la primera división meiótica continúa la segunda, que avanza hasta la etapa de metafase, cuando se produce la segunda detención de la meiosis. A la expulsión del primer cuerpo polar, el folículo sobresale en la superficie del ovario. El vértice de la protrusión será el estigma.
El estímulo para la ovulación es el pico de LH liberado por la adenohipófisis en el punto medio del ciclo menstrual. El edema y la liberación de varios compuestos con actividad farmacológica, como las prostaglandinas, la histamina y la vasopresina, sirven de punto de partida para una serie de reacciones que redundan en la producción local de colagenasa (figura 2-1). La degradación del colágeno, la isquemia y la muerte de algunas de las células suprayacentes causan un debilitamiento de la pared externa del folículo. Este debilitamiento conduce a la rotura de la pared externa del folículo unas 8 a 36 horas después del pico de LH
La ovulación da como resultado la expulsión tanto del líquido astral como del óvulo desde el ovario hacia la cavidad peritoneal. El óvulo se expulsa como un complejo que consta de: 1)el óvulo, 2)la zona pelúcida, 3)la corona radiada de 2 o 3 capas celulares de espesor y 4) la matriz pegajosa que contiene las células que rodean al cumulus oophorus. Las células adheridas al óvulo se denominan como corona radiada después de que se ha producido la ovulación. La expulsión y fecundación de 2 óvulos pueden dar origen a un embarazo de gemelos fraternos.
El leve dolor en el momento de la ovulación que algunas mujeres experimentan se denomina como mittelschmerz (término alemán que significa “intermedio”), puede acompañar a la ligera hemorragia de la rotura folicular. Otro signo de ovulación es una leve elevación de temperatura corporal basal. A menudo esto se ha usado como punto de referencia en los métodos del ritmo para la anticoncepción.
Transporte del óvulo
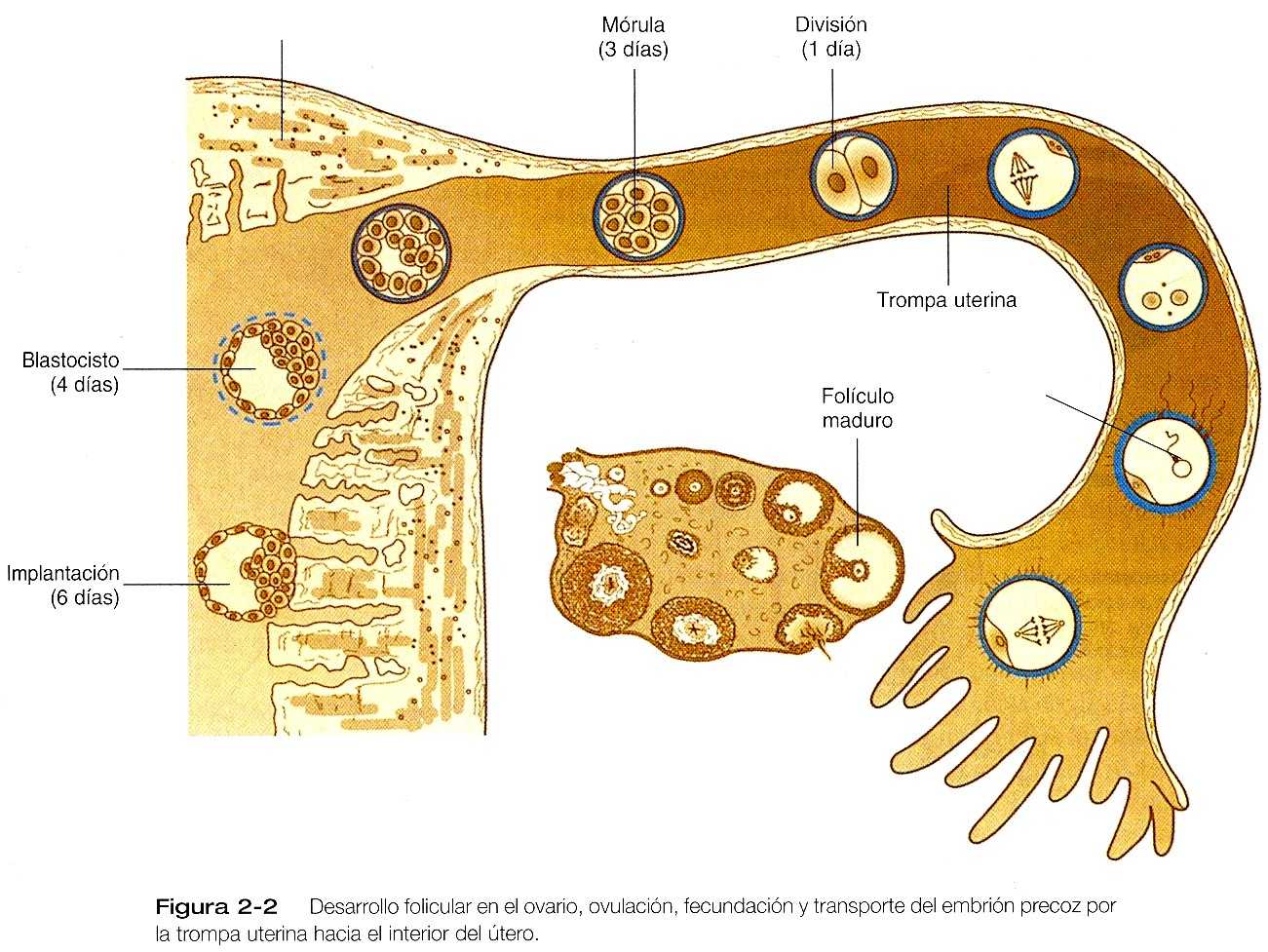
El primer paso en el transporte es la captación del óvulo expulsado por la trompa de Falopio. Poco antes de la ovulación, las células epiteliales de la trompa uterina se vuelven más ciliadas y aumentan la actividad de la musculatura lisa tubárica y del ligamento suspensorio como resultado de influencias hormonales. Al momento de la ovulación, las fimbrias del extremo de la trompa se acercan al ovario y parecen hacer un barrido rítmico sobre su superficie. Esta acción, más las corrientes originadas por los cilios, capta con gran efectividad el complejo ovular expulsado del folículo. Los estudios experimentales en conejos han demostrado que el aumento de volumen que hacen las cubiertas celulares alrededor del óvulo expulsado es importante para facilitar la captación de éste y su transporte por la trompa uterina. Los óvulos desnudos o los objetos inertes del mismo tamaño no se transportan tan fácilmente. Algunas evidencias sugieren que el óvulo humano puede necesitar menos de sus cubiertas celulares para el transporte que el óvulo del conejo.
Aun sin estos tipos de adaptaciones naturales, la capacidad de las trompas uterinas para captar el huevo es notable: Si las fimbrias de la trompa se extirpan, la captura del huevo se produce con gran frecuencia; ha habido embarazos en mujeres que tras una intervención quirúrgica sólo tenían un ovario y la trompa contralateral. En estos casos, el óvulo habría tenido que recorrer libre en la cavidad pélvica una distancia considerable antes de entrar en el ostium de la trompa del otro lado, o una trompa de gran longitud podría haber llegado hasta el ovario contralateral. Una vez dentro de la trompa, el óvulo se transporta hacia el útero, en principio como resultado de las contracciones de la musculatura lisa de la pared tubárica.
Aunque la superficie ciliada de la mucosa de la trompa también desempaña una función en el transporte del huevo, su acción no es obligatoria porque las mujeres que tienen el síndrome de los cilios inmóviles suelen ser fértiles.
Mientras se encuentra en la trompa, el óvulo es bañado por el líquido tubárico que es una combinación de la secreción del epitelio de la trompa y un trasudado procedente de los capilares situados inmediatamente debajo del epitelio. En algunos mamíferos, la exposición a las secreciones del oviducto es importante para la supervivencia del óvulo, pero la función de este líquido en los seres humanos no se ha establecido.
El transporte tubárico del óvulo suele tardar de tres a cuatro días, con independencia de si tiene lugar o no la fecundación figura 2_2). El transporte del óvulo suele efectuarse en dos fases: un transporte lento en la ampolla (aproximadamente 72 horas) y una fase más rápida (ocho horas) durante las cuales el óvulo o el embrión pasa a través del istmo hacia el interior del útero (pág 40). Mediante un mecanismo aún no comprendido del todo, quizás edema o reducción de la actividad muscular, se impide de forma transitoria al óvulo penetrar en la porción ístmica de la trompa, pero bajo la influencia de la progesterona se relaja la unión tubouterina y permite la entrada del óvulo.
Alrededor de 80 horas después de la ovulación, el óvulo o el embrión ha pasado de la trompa al útero. Si no ha habido fecundación, el óvulo degenera y es fagocitado (implantación del embrión se discute en el capítulo 3)
Transporte espermático
El transporte de los espermatozoides tiene lugar tanto en el varón como en la mujer. En el varón, el transporte de los espermatozoides se relacionan en forma estrecha con la madurez estructural y funcional, mientras que en el tracto reproductor femenino es importante que tantos espermatozoides como sea posible pasen con rapidez hasta el tercio externo de la trompa uterina, donde pueden encontrarse con el óvulo.
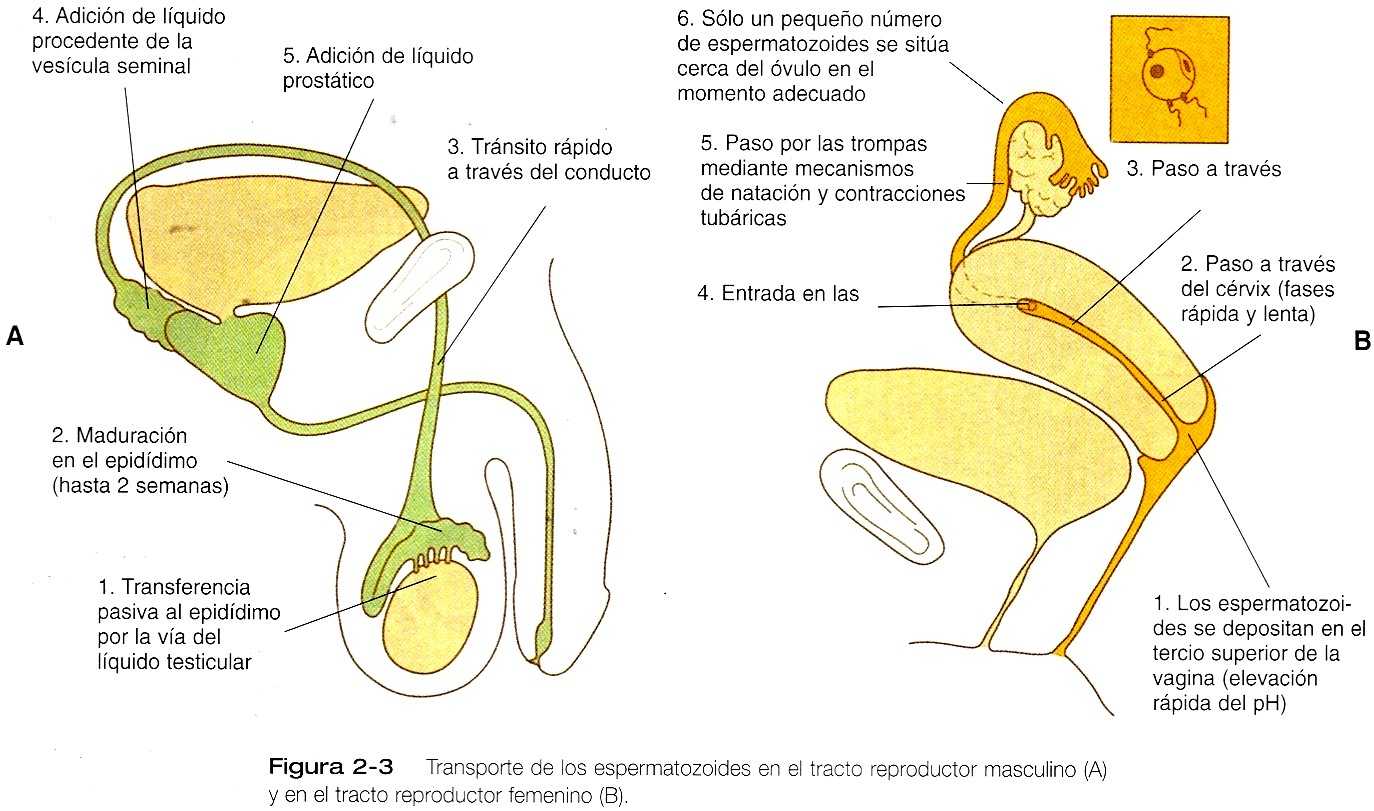
Después de la espermiogénesis en los túmulos seminíferos, los espermatozoides están maduros desde el punto de vista morfológico pero son inmóviles e incapaces de fecundar al óvulo (figura 2_3). Los espermatozoides son transportados pasivamente en el líquido testicular desde los túmulos seminíferos hasta la cabeza del epidídimo a través de la retetestis y los conductillos eferentes.
Los impulsa la presión del líquido generada en los túmulos seminíferos y los ayudan las contracciones del músculo liso y las corrientes filiares de los conductillos eferentes. Los espermatozoides están alrededor de doce días en el conducto muy enrollado del epidídimo, tiempo durante el cual experimentan su maduración bioquímica. Este período de maduración se asocia con cambios en las glucoproteínas de la membrana plasmática de la cabeza del espermatozoide. Cuando los espermatozoides han alcanzado la cola del epidídimo, ya pueden fecundar al óvulo.
En el momento de la eyaculación, los espermatozoides pasan rápidamente por el conducto deferente y se mezclan con las secreciones líquidas de las vesículas seminales y la próstata. El líquido prostático contiene abundante ácido cítrico, fosfatasa ácida, iones de cinc y magnesio, mientras que el líquido de las vesículas seminales es rico en fructuosa (la principal fuente de energía de los epermatozoides) y prostaglandinas. Los 2 a 6 mL de una eyaculación (semen o líquido seminal) suelen constar de 40 a 250 millones de espermatozoides mezclados con líquido alcalino procedente de las vesículas seminales (60% del total) y secreción ácida (pH, 6,5) de la próstata (30% del total). El pH del semen normal suele estar entre 7,2 y 7,8. A pesar del gran número de espermatozoides normalmente presentes en el eyaculado, sólo 25 millones de cada eyaculación pueden ser compartibles con una fertilidad normal.
En la mujer, el transporte de los espermatozoides comienza en el tercio superior de la vagina y termina en la ampolla de la trompa uterina, donde los espermatozoides establecen contacto con el óvulo. Durante la cópula, el líquido seminal suele depositarse en el tercio superior de la vagina, donde su composición y su capacidad de amortiguación inmediata protegen a los espermatozoides el líquido ácido que se encuentra en esta zona de la vagina. El líquido vaginal ácido suele ejercer una acción bactericida que protege al canal cervical de los organismos patógenos. En unos diez segundos, el pH de la vagina se eleva desde 4,3 hasta 7,2. El efecto amortiguador dura sólo unos pocos minutos en los seres humanos, pero da tiempo suficiente a los espermatozoides para que se aproximen al cérvix en un ambiente óptimo para su mortalidad (pH de 6,0 a 6,5).
La siguiente barrera que los espermatozoides han de superar es el canal cervical y el moco que lo bloquea. Los cambios en la presión intravaginal pueden succionar a los espermatozoides hacia el orificio cervical, pero los movimientos natatorios también parecen ser importantes para que la mayoría de los espermatozoides puedan penetrar el moco cervical.
La composición y la viscosidad del moco cervical varían mucho durante el ciclo menstrual. Compuesto por mucina cervical (una glucoproteína con un alto contenido de carbohidratos) y componentes solubles, el moco cervical no es fácil de penetrar. No obstante, entre los días 9 y 16 del ciclo, su contenido de agua aumenta, lo cual facilita el paso de los espermatozoides por el cerviz alrededor del tiempo de la ovulación; este tipo de moco suele denominarse moco E.
Después de la ovulación, bajo la influencia de la progesterona, cesa la producción del moco cervical acuoso y se produce un nuevo tipo de moco viscoso con un contenido de agua mucho menor, que se denomina moco G. Este moco progestacional es casi del todo resistente a la penetración de los espermatozoides. Conocer las propiedades del moco cervical ha sido un método bastante efectivo de planificación familiar natural.
Hay dos modos de transporte de los espermatozoides a través del cérvix Uno es una fase de transporte rápido inicial, por el cual algunos espermatozoides pueden alcanzar las trompas uterinas entre cinco y 20 minutos después de la eyaculación. Este transporté rápido depende más de los movimientos musculares del tracto reproductor femenino que de la mortalidad de los espermatozoides en sí misma. El otro, la fase lenta del transporte de los espermatozoides, implica que estos naden a través del moco cervical (viajando a una velocidad de 2 a 3 mm/h), su almacenamiento en las criptas cervicales y su paso final por el canal cervical incluso dos a cuatro días más tarde.
Se sabe relativamente poco sobre el paso de los espermatozoides a través de la cavidad uterina, pero la contracción del músculo liso uterino, más que la mortalidad espermática, parece ser el principal mecanismo de transporte en el órgano. En este punto los espermatozoides entran en una de las trompas uterinas. Aunque se ha sugerido que el líquido folicular o algo asociado con el óvulo pueden ejercer un quimiotactismo sobre los espermatozoides en los mamíferos, esto no se ha comprobado y parece bastante probable que sea el azar lo que determine en cuál trompa uterina entrarán los espermatozoides. Una vez dentro de la trompa, los espermatozoides se acumulan en el istmo hasta que alguna señal asociada con la ovulación estimula su migración adicional por trompa arriba. En el momento de su liberación, los espermatozoides entran en un período de hiperactividad transitorio que concisita en movimientos natatorios exagerados que puedan liberarlos de los sitios de su unión temporal en el istmo.
Un número sorprendentemente reducido de espermatozoides (sólo unos 200) se encuentran en el extremo distal de la trompa en el momento adecuado. Con los movimientos musculares de la trompa y algunos movimientos natatorios, los espermas siguen su camino por la trompa arriba e incluso fuera del infundíbulo y hacia la cavidad peritoneal. El transporte simultáneo de un óvulo hacia el útero y de espermatozoides hacia el extremo distal de la trompa se explica en la actualidad con base en las contracciones peristálticas de la musculatura tubárica, que subdividen la trompa en compartimientos. En uno de ellos en particular, los gametos quedan atrapados en movimientos de rotación que en uno o dos días aproximan al óvulo y al espermatozoide.
Los cálculos más recientes sugieren que los espermatozoides pueden mantener su actividad en el tracto reproductor femenino hasta por unas 80 horas. Durante su paso por el aparato reproductor, los espermatozoides experimentan la reacción de capacitación. Ésta consiste en la alteración de la superficie de las glucoproteínas de los espermatozoides bajo la influencia de las secreciones de los tejidos del tracto reproductor femenino, y es necesaria para que los espermatozoides puedan fecundar al óvulo. Esto último por lo general ocurre en la porción ampular de la trompa uterina.
Formación y función del cuerpo lúteo
de la ovulación y del embarazo
Mientras que el óvulo está pasando por las trompas uterinas, el folículo roto del cual surgió sufre una serie de cambios profundos esenciales para la progresión de los acontecimientos que den lugar a un embarazo normal.
Poco después de la ovulación, la membrana basal que separa las células de la granulosa de las células de la teca interna se rompe y permite que crezcan vasos sanguíneos tecales hacia el interior de la cavidad del folículo roto. Al mismo tiempo, las células de la granulosa pasan por una serie de cambios importantes en su forma y su función (luteínización). En unas 30 a 40 horas desde el pico de LH, estas células –que ahora se llaman células luteínicas de la granulosa- comienzan a producir cantidades crecientes de progesterona con algo de estrógenos. Este patrón de secreción sirve de base hormonal para los cambios de los tejidos reproductores femeninos en la última mitad del ciclo menstrual, durante este período el folículo sigue creciendo. A causa de su color amarillo se le conoce como cuerpo lúteo. Las células luteinizadas de la granulosa se distinguen en la fase final; aunque han dejado de dividirse, continúan segregando progesterona durante diez días.
En ausencia de fecundación y de un estímulo hormonal suministrado por el embrión precoz, el cuerpo lúteo comienza a degenerar (luteólisis) al final del ciclo menstrual. La luteólisis parece implicar tanto la programación previa de las células lúteas para la apoptosis (muerte celular) como la de los factores luteolíticos uterinos, por ejemplo la prostaglandina F2 . La regresión del cuerpo lúteo y la reducción concomitante de la producción de progesterona causan una de privación hormonal que redunda en los cambios degenerativos del tejido endometrial durante los últimos días del período menstrual.
Durante la regresión del cuerpo lúteo, las células luteinizadas de la granulosa degeneran y son remplazadas por tejido de colágeno cicatricial. A causa de color blanquecino, el antiguo cuerpo lúteo se conoce ahora con el nombre de corpus albicns (cuerpo blanco).
Si se produce la fecundación, la producción de la hormona proteica gonadotropina coriónica humana por los futuros tejidos placentarios mantiene al cuerpo lúteo en estado funcional e incluso hace que aumente su tamaño y su producción de hormonas. Dado que las células luteinizadas de la granulosa no pueden dividirse y también dejan de producir progesterona al cabo de diez días, el gran cuerpo lúteo gestacional se compone principalmente de células teca luteínicas.
El cuerpo lúteo del embarazo conserva su función durante los primeros meses de la gestación. Después del segundo mes, la placenta produce cantidades suficientes de4 estrógenos y progesterona para mantener el embarazo por si sola. En este punto la gestación continuaría aún si se extirparan los ovarios.
FECUNDACIÓN
La fecundación es una serie de procesos, más que un acontecimiento único. Vistos en el más amplio, estos procesos comienzan cuando los espermatozoides empiezan a penetrar la corona radiada que rodea al óvulo y terminan con la mezcla de los cromosomas maternos y paternos después de que el espermatozoide ha entrado en el óvulo.
Penetración de la corona radiada
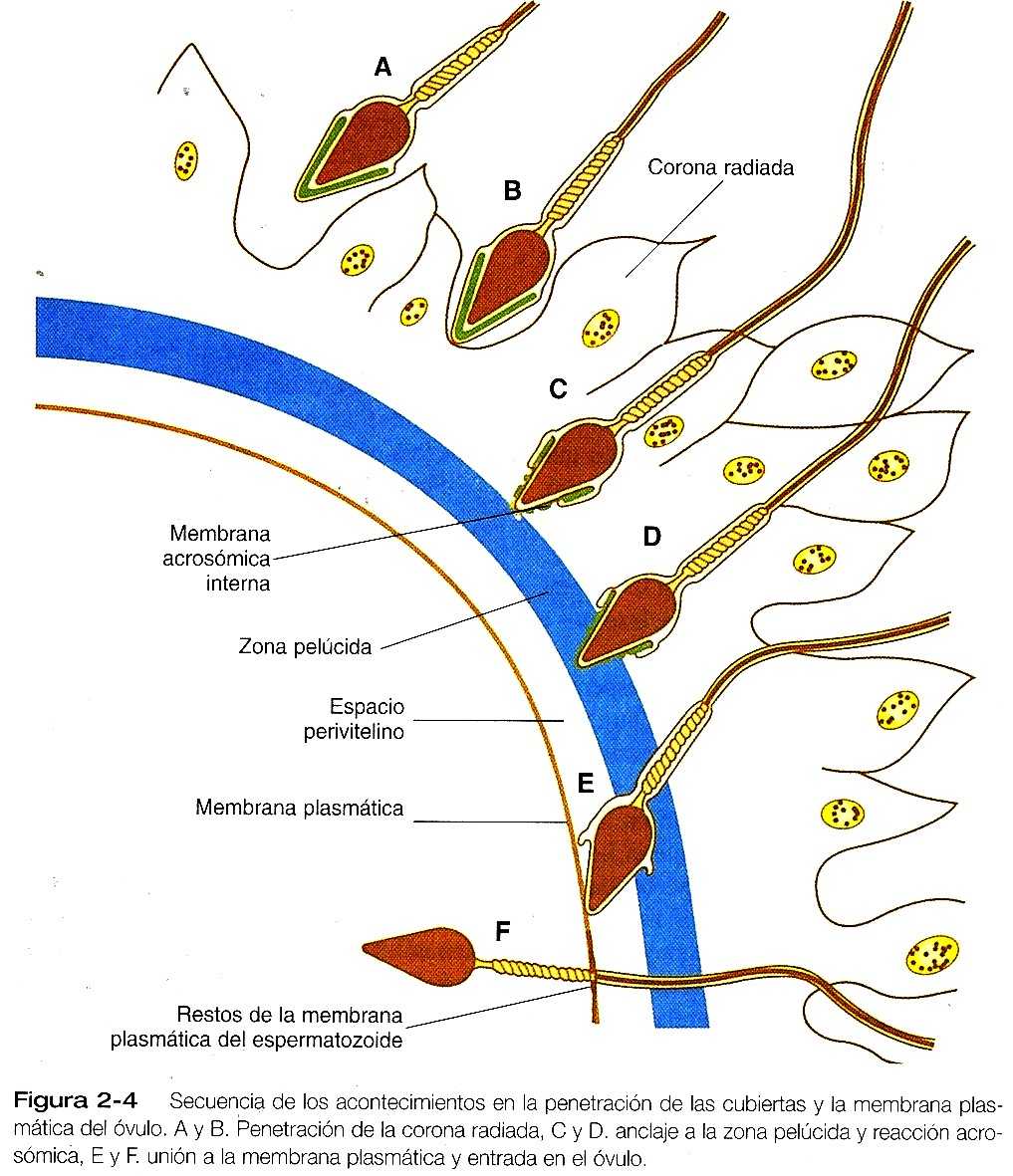
Cuando los espermatozoides se encuentran por primera vez con el óvulo en la ampolla de la trompa uterina, se enfrentan a la corona radiada y posiblemente a algunos restos del montículo ovárico (cúmulus oophorus), que representa la capa más externa del complejo ovular (figura 2_4). La corona radiada es una capa muy celular que tiene una matriz intercelular formada por proteínas y una alta concentración de carbohidratos, en especial ácido hialurónico.
Aunque durante mucho tiempo se ha creído que la hialuronidasa que emana de los espermatozoides desempeña un papel fundamental en la penetración de la corona radiada, la evidencia no es absoluta. Al parecer, los movimientos natatorios activos de los espermatozoides son esenciales en la penetración.
|
Cuadro 2_1 Algunas de las principales enzimas acrosómicas de los mamíferos Proteína ácida Fosfolipasa C Acrosina b-Galactosidasa Arilaminidasa b-Glucuronidasa Arilfufatasa Hialuronidasa Colagenasa Neuraminidasa Esterasa Proacrosina |
Anclaje a la zona pelúcida y penetración de la misma
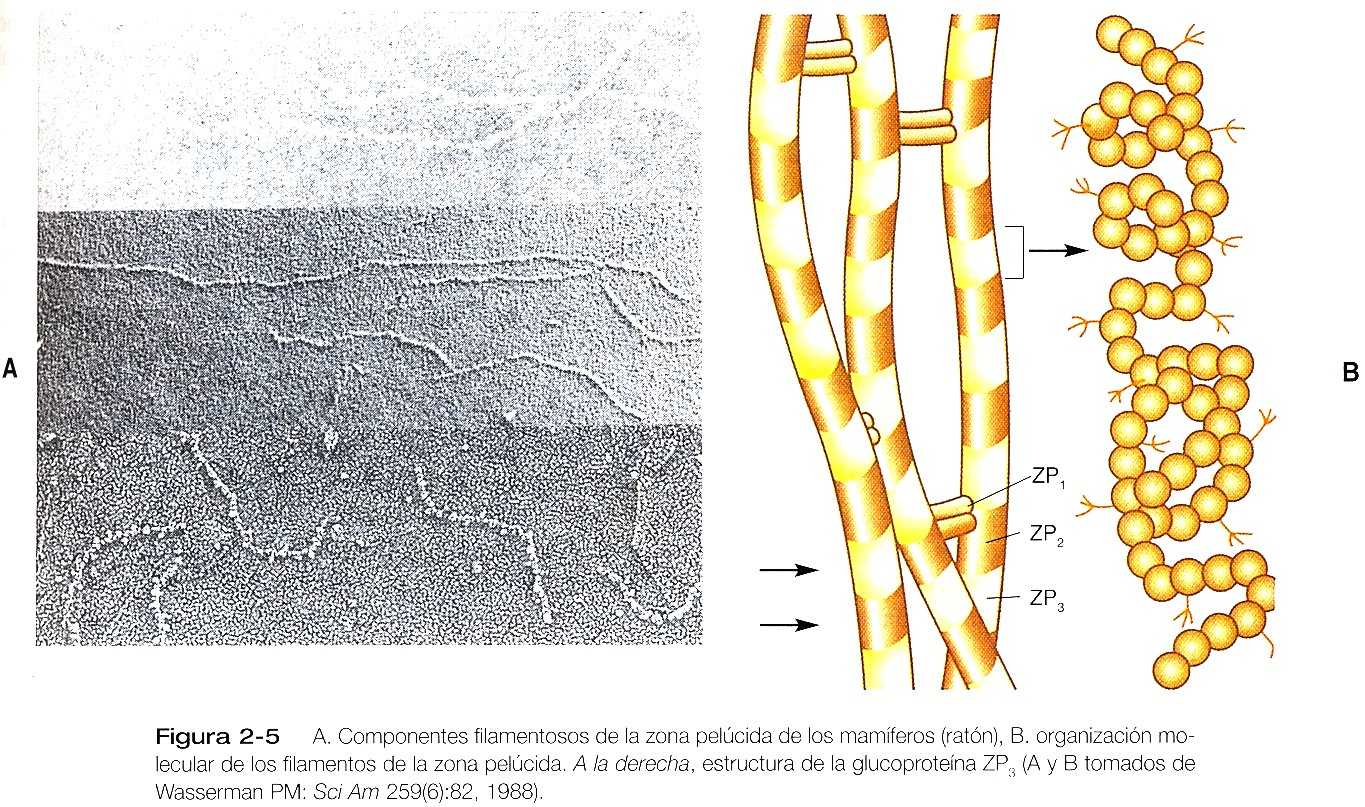
La zona pelúcida, que tiene un espesor de 13 mm, en los seres humanos, consta principalmente de tres glucoproteínas: ZP1’, la ZP2’, y la ZP3’ con pesos moleculares de 200.000, 120.000 y 83.000 daltons. La ZP2’, y la ZP3’, se combinan para formar unidades básicas que se polimerizan en largos filamentos, los cuales se unen a intervalos mediante puentes cruzados de moléculas de ZP1’ (figura 2_5). Se calcula que la zona pelúcida del óvulo no fecundado del ratón contiene más de un millardo de copias de la proteína ZP3’.
Después de que han penetrado la corona radiada, los espermatozoides establecen una fuerte unión con la zona pelúcida por medio de la membrana plasmática de sus cabezas (figura 2_4). La molécula ZP3’ específicamente los O-ligandos oligosacáridos anclados al núcleo polipeptídico, actúa con receptor de espermatozoides en la zona pelúcida del ratón. Las moléculas de la superficie de la cabeza del espermatozoide actúa como lugares específicos de unión para los receptores espermáticos ZP3’ de la zona pelúcida. Se han propuesto varias moléculas, de las cuales una candidata muy probable es la galactosil transferasa.
Al unirse a la zona pelúcida, los espermatozoides de los mamíferos experimentan la reacción acrosómica. La esencia de esta reacción es la fusión de partes de la membrana acrosómica externa con la membrana plasmática suprayacente y la liberación de las partes fusionadas en forma de pequeñas vesículas. El resultado es la expulsión de una gran cantidad de enzimas que están almacenadas en el acrosoma (cuadro 2_1).
La reacción acrosómica de los mamíferos parece ser estimulada por la molécula ZP3’ (figura 2_5). En contraste con la función de receptor espermático de la ZP3’, un gran segmento de la cadena polipeptídica de la molécula de ZP3’ debe estar presente para inducir la reacción acrosómica. Uno de los cuales sucesos iniciadores de la reacción acrosómica es el paso masivo de Ca a través de membrana plasmática de la cabeza del espermatozoide. Este proceso, que se acompaña de entrada de Na y de salida de H eleva el pH intracelular. Pronto se produce la fusión de la membrana plasmática suprayacente. A medida que las vesículas se fusionan, las membranas se desprenden, el contenido enzimático del acrosoma se libera y puede ayudar a los espermatozoides a encontrar su camino a través de la zona pelúcida.
Después de la reacción acrosómatica, la membrana acrosómatica interna forma la superficie externa que cubre la mayor parte de la cabeza del espermatozoide (figura 2_4D). Hacia la base de las cabeza del espermatozoide (en la región ecuatorial), la membrana acrosómica interna se fusiona con la membrana plasmática postacrosómica remanente para mantener la continuidad de la membrana alrededor de la cabeza del espermatozoide.
Sólo después de terminada la reacción acrosómica el espermatozoide puede comenzar a penetrar con éxito la zona pelúcida. Tal penetración se logra mediante una mezcla de propulsión mecánica en virtud de movimientos de la cola del espermatozoide y de digestión de un trayecto de la zona pelúcida mediante la acción del acrosoma. Aunque es probable que la acción de varias de estas enzimas acrosómicas intervenga en el inicio de la penetración a través de la zona, la enzima más importante es la acrosina, una proteinasa serica que se fija a la membrana acrosómica interna. Muchos investigadores opinan que el túnel bien definido que marca el trayecto por la zona y hacia el interior del espacio perivitelino (el espacio que queda entre la membrana plasmática del óvulo y la zona pelúcida), puede establecer contacto directo con la membrana plasmática del óvulo.
Unión y fusión del espermatozoide
y el óvulo
Después de un breve período de tránsito a través del espacio perivitelino, hace contacto con el óvulo. En dos fases distintas, el espermatozoide primero se fija y después se fusiona con la membrana plasmática del óvulo. La unión entre el espermatozoide y el huevo tiene lugar en la región ecuatorial de la cabeza del espermatozoide, donde la membrana acrosómica interna ya se ha fusionado con la membrana plasmática remanente del espermatozoide y las microvellosidades que rodean a la mayor parte del óvulo. Las moléculas de la membrana plasmática del espermatozoide se ligan a las moléculas de integrina a6b1 de la superficie del óvulo. La reacción acrosómica parece causar algunos cambios en las propiedades de la membrana del espermatozoide porque si la reacción acrosómica no ha tenido lugar, el espermatozoide no puede fusionarse con el óvulo. La fusión real entre el espermatozoide y el óvulo hace que sus membranas plasmáticas establezcan una continuidad. Después de la fusión inicial, el contenido del espermatozoide (la cabeza, la pieza media y usualmente la cola) se sumerge dentro del óvulo, mientras que su membrana plasmática, que es diferente de la del óvulo desde punto de vista antigénico, se incorpora la membrana plasmática del óvulo y sigue siendo reconocible al menos hasta el comienzo de la división del óvulo fecundado o cigoto. Aunque las mitocondrias localizadas en el cuello del espermatozoide entran en el óvulo, no parecen contribuir al complemento mitocondrial funcional del cigoto.
Prevención de la polispermia
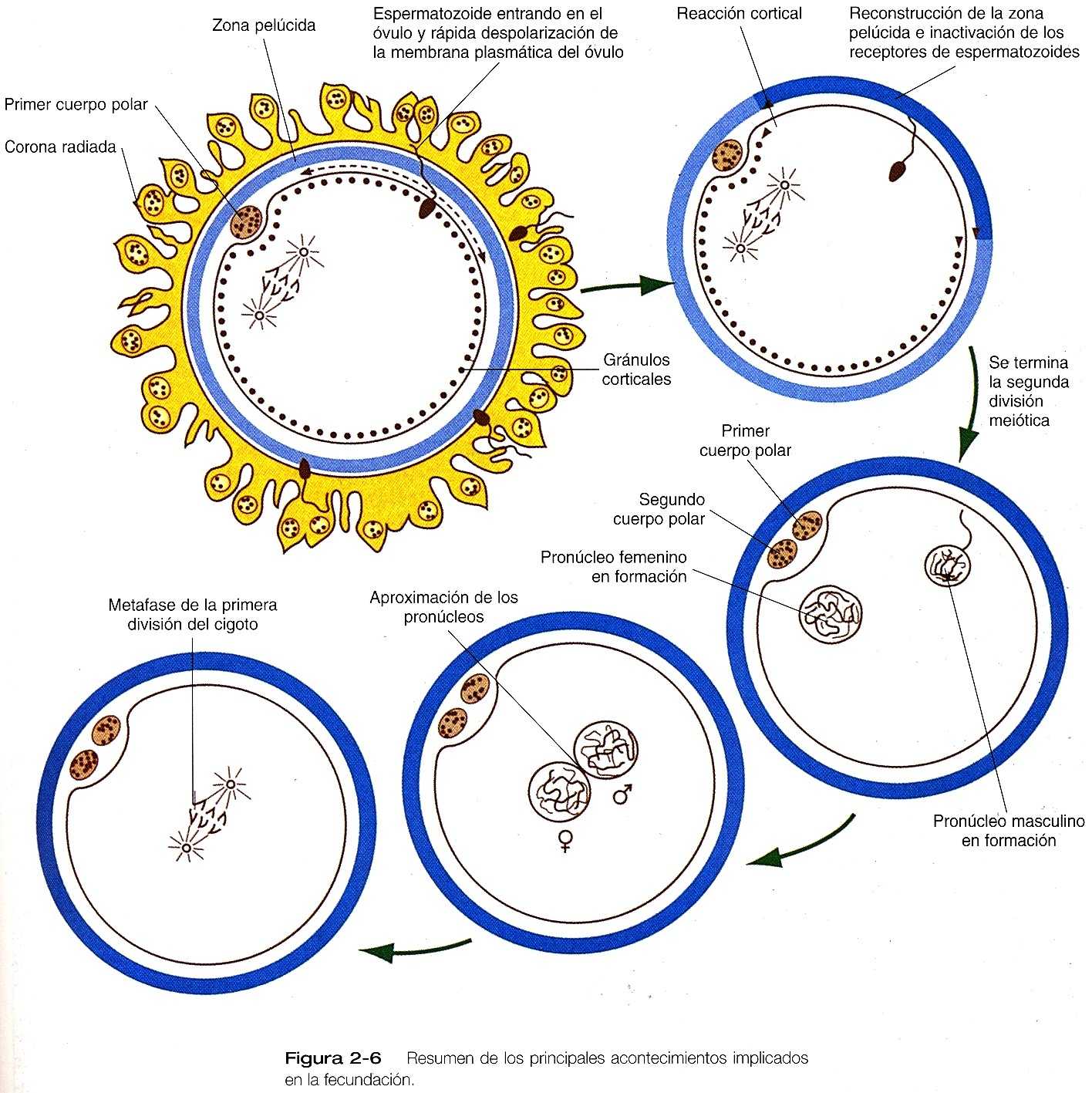
Una vez que un espermatozoide se ha fusionado con un óvulo, debe evitarse la entrada de otros espermatozoides en el óvulo (polispermia) o es probable que se produzca un desarrollo anómalo. En la fecundación de los vertebrados, suele haber dos bloqueos contra la polispermia, uno lento y uno rápido.
El bloqueo rápido de la polispermia, que se ha estudiada en los erizos de mar, consta de una rápida despolarización eléctrica de la membrana del óvulo. El potencial de reposo de la membrana plasmática del óvulo. El potencial de reposo de la membrana del óvulo cambia desde unos -70 mV hasta +10mV en los primeros dos segundos después de la fusión del espermatozoide con el óvulo. Este cambio en el potencial de membrana impide que otros espermatozoides se adhieren a la membrana plasmática del óvulo. Se sabe poco sobre el bloque rápido de la polispermia en el óvulo humano, pero como no hay más que unos pocos espermatozoides en torno al óvulo de los mamíferos en el momento de la fecundación, la necesidad de un bloqueo rápido es corto, suele durar sólo un minuto en los erizos y cinco en los mamíferos. Este tiempo es suficiente para que el óvulo prepare un bloqueo lento permanente.
El bloqueo lento de la polispermia comienza con la propagación de una oleada de Ca-- a partir del lugar de la unión entre el espermatozoide y el óvulo. En un par de minutos, la oleada de Ca++ ha recorrido todo el óvulo, actuando de forma secuencial sobre los gránulos corticales a medida que pasa por ellos. La exposición al Ca--hace que los gránulos corticales se fusionen con la membrana plasmáticas y que liberen su contenido (enzimas hidrolíticas y polisacáridos) hacia el espacio perivitelino. Los polisacáridos liberados en el espacio perivitelino se hidratan y se hinchan, y esto hace que la zona pelúcida se reorganice en la superficie del óvulo.
Los productos secretorios de los gránulos corticales se difunden hacia la zona pelúcida porosa e hidrolizan las moléculas receptoras de espermatozoides (ZP3 en el ratón) de la zona. Esta reacción en la zona ha sido observada visualmente en los óvulos humanos que se han sometido a fecundación in vintro.
Las diferencias moleculares entre especies en las regiones de unión de espermatozoides de la molécula de ZP3 pueden servir como base para impedir que los espermatozoides de una especie puedan fecundar el óvulo de otra especie en la composición de ZP3; esto puede explicar porqué la penetración de la zona pelúcida por los espermatozoides de especies de mamíferos estrechamente relacionadas es posible en ocasiones, mientras que es frecuente entre los animales inferiores.
La especificidad entre el sitio receptor ZP3 y los lugares de unión en el espermatozoide podría usarse como base para un método anticonceptivo de tipo inmunológico.
Además de los cambios en la zona pelúcida, las alteraciones de las moléculas receptoras de espermatozoides en la membrana plasmática del óvulo humano pueden hacer que este mismo se vuelva refractario a la penetración por otros espermatozoides.
Activación metabólica del óvulo
Uno de los cambios más importantes suscitados por la penetración del espermatozoide es una rápida intensificación de la respiración y el metabolismo del óvulo. Los mecanismos subyacentes es estos cambios no se conocen a cabalidad, ni siquiera en los sistemas mejor estudiados, pero la liberación precoz de Ca++desde almacenamiento internos, se cree que es el suceso iniciador. En algunas especies, la liberación de Ca ++ va enseguida en corto tiempo por un intercambio de Na + extracelular a través de la membrana plasmática. El resultado es una elevación en el pH intracelular, que precede a un incremento del metabolismo oxidativo.
Descondensación del núcleo del espermatozoide
En el espermatozoide maduro, la cromatina nuclear está empacada en forma muy apretada, debido en gran parte a las uniones cruzadas –SS— (disulfuro) que se producen entre las moléculas de protamina que constituyen complejos con el ácido desoxirribonucleico (DNA) durante la espermatogénesis. Poco después de que la cabeza del espermatozoide entra en el citoplasma del interior del óvulo afecten el contenido nuclear comienza a aumentar, lo cual permite que los factores citoplasmáticos del interior del óvulo afecten el contenido nuclear del espermatozoide. Después de la reducción de las uniones disulfuro de las protaminas a grupos sulfhidrilo (--SH--) por el glutación reducido en el ooplasna, las protaminas se pierden con rapidez de cromatina del espermatozoide, la cromatina comienza a extenderse en el núcleo (ahora denominado pronúcleo) a medida que se desplaza para acercarse al material nuclear del óvulo. después de un corto período durante el cual los cromosomas masculinos están desnudos, las historias empiezan a asociarse con los cromosomas.
Terminación de la meiosis y desarrollos del pronúcleo en el óvulo
Después de la penetración del óvulo por el espermatozoide, el núcleo del óvulo, que ha estado detenido en la metafase de le segunda división meiótica, termina la última división y libera un segundo cuerpo polar dentro del espacio perivitelino (figura 2_6). La membrana del pronúcleo, derivada en su mayor parte del retículo endoplasmático del óvulo, se forma alrededor del material cromosómico femenino. Algunos factores citoplasmáticos parecen controlar el crecimiento tanto del pronúcleo femenino como del masculino. Parece haber replicación del DNA en los pronúcleos se aproximan entre sí. Cuando los pronúcleos masculino y femenino entran en contacto, las membranas se rompen y los cromosomas se entremezclan. Los cromosomas maternos y paternos se organizan con rapidez alrededor de un huso mitótico en preparación para una división mitótica corriente. En este punto, puede decirse que el proceso se denomina cigoto.
CASO CLÍNICO (2-0)
Una mujer de 33 años que fue a una histerectomía desea fervientemente tener un hijo propio. Puede producir óvulos porque sus ovarios son funcionales. Ella y su esposo, que es un hombre pudiente, desean intentar la fertilización in vintro y la transferencia embrionaria. Encuentran a una mujer, que por $10,000 está dispuesta a permitir que le transfieran a su útero el embrión de la pareja y a servir como madre sustituta durante el embarazo. La inducción de la superovulación se realiza con mucho éxito y los médicos logran fertilizar ocho óvulos in vintro. Se implantan tres embriones en la madre sustituta; el resto se congela para posible uso en el futuro. La transferencia embrionaria tiene éxito, y la madre sustituta queda embarazada de gemelos. Después de que éstos nacen, la madre sustituta siente que ha establecido un vínculo tan intenso con ellos que cree que tiene derecho a criarlos. Los padres genéticos llevan el caso ante los tribunales, pero antes de que la demanda llegue ante los jueces, la pareja muere en un accidente aéreo. La madre sustituta ahora sostiene que ella debe recibir la enorme herencia en nombre de sus gemelos, pero la hermana del padre, igualmente al tanto de las implicaciones monetarias, reclama que debe ser ella quien se encargue de los gemelos. También surge el problema de qué hacer con los cinco embriones congelados restantes.
Este caso es ficticio, pero todos sus elementos has acaecido de forma aislada. ¿Cómo resolvería usted los siguientes aspectos éticos y legales?:
¿A quién se deben adjudicar los gemelos?
¿Qué debe acres con los embriones congelados restantes?
¿Qué se logra con la fecundación?
El proceso de la fecundación ata varios casos biológicos sueltos:
1.-Estimula al huevo a terminar la segunda división meiótica.
2.-Restablece en el cigoto el número diploide normal de cromosomas (46 en los seres humanos).
3.-Se determina el sexo del futuro embrión mediante el complemento cromosómico del espermatozoide (si el espermatozoide contiene 22 autosomas y un cromosoma X, el embrión será femenino desde el punto de vista genético, y si contiene 22 autosomas y un cromosoma Y, el embrión será masculino.)
4.- A través de la mezcla de los cromosomas paternos y maternos, el cigoto es un producto genéticamente único de la redistribución cromosómica, lo cual es importante para la vialidad de cualquier especie.
5.- El proceso de la fecundación causa la activación metabólica del óvulo, que es necesario para que se produzcan la división y el desarrollo embrionario posterior.
RESUMEN
1. Penetración de la corona radiada
2. Anclaje a la zona pelúcida.
3. Reacción acrosómica y penetración de la zona pelúcida.
4. Unción y fusión del óvulo y espermatozoide
5. Evitación de la polispermia.
6. Activación metabólica del huevo.
7. Descondensación del núcleo del espermatozoide.
8. Terminación de la meiosis en el óvulo
9. Desarrollo y fusión de los pronúcleos masculino y femenino
§ El anclaje del espermatozoide a la zona pelúcida es mediado por la proteína ZP3 que también estimula la reacción acrosómica.
§ La reacción acrosómica comprende la fusión de la membrana acrosómica externa con la membrana plasmática del espermatozoide y la fragmentación de las membranas fusionadas, que conduce a la liberación de las enzimas acrosómicas. Una de éstas la acrosina, es una proteinasa de serina, que digiere los componentes de la zona pelúcida y ayuda a la penetración de los espermatozoides a través de la zona.
§ Después de la fusión de los espermatozoides con la membrana del óvulo, una despolarización eléctrica rápida produce el primer bloqueo de la polispermia en el óvulo. Esto va seguido una oleada de Ca++ que hace que los gránulos corticales liberen su contenido hacia el espacio perivitelino y en último termino inactiven los receptores de espermatozoides y el metabolismo del óvulo.
§ La penetración del espermatozoide estimula una rápida intensificación de la respiración y el metabolismo del óvulo.
§ En el interior del óvulo, el material nuclear del espermatozoide se descondensa y forma el pronúcleo masculino. Al mismo tiempo, el óvulo completa la segunda división meiótica, y el material nuclear remanente es rodeado por una membrana, con lo cual se forma el pronúcleo femenino.
§ Después de la replicación del DNA, los pronúcleos masculino y femenino se juntan y sus cromosomas se organizan para una división mitótica. Entonces se completa la fecundación, y el óvulo fecundado recibe en este momento el nombre apropiado de cigoto.
§ El tratamiento de la infertilidad mediante la fecundación in vintro y la transferencia embrionaria es un proceso de varias etapas que comprende la estimulación de la producción de gametos con fármacos como el citrato del clomifeno, la obtención de los óvulos mediante técnicas laparoscópicas o ecográficas, el almacenamiento de los gametos mediante congelación, la realización de la fecundación in vitro y el cultivo de los embriones, la preservación de los mismos y su posterior transferencia a la madre.
§ Otras técnicas que se usan para el tratamiento de la infertilidad son GIFT, la transferencia directa de los gametos en las trompas, y ZIFT, la transferencia de los cigotos en las trompas. Estas técnicas pueden usarse tanto en madres biológicas como sustitutas.
|
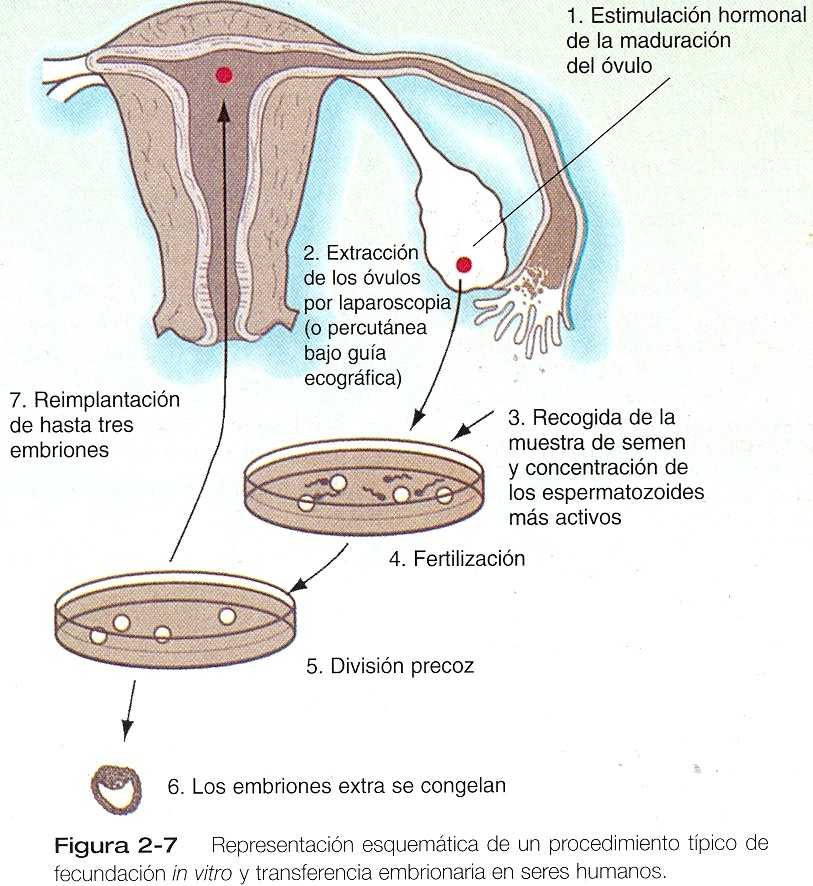
CORRELACIÓN CLÍNICA 2_1 Tratamiento de la infertilidad mediante fecundación in vitro y transferencia embrionaria Ciertos tipos de infertilidad causada por un número o una motilidad inadecuados de los espermatozoides o por la obstrucción de las trompas uterinas se tratan en la actualidad mediante ka fertilización in vitro de un óvulo y la posterior transferencia del cigoto en división al tracto reproductor de la mujer. Para lograr esto se requiere la aplicación secuencial de diversas técnicas que inicialmente se desarrollaron para la reproducción asistida de animales domésticos como vacas y ovejas. Las técnicas pertinentes con: 1)estimulación de la producción de gametos, 2) obtención de gametos masculinos y femeninos, 3) almacenamiento de los gametos, 4) fertilización de los óvulos, 5) cultivo in vitro de los cigotos en división, 6) conservación de los embriones y 7) introducción de los embriones dentro del útero (figura 2_7). ESTIMULACIÓN DE LA PRODUCCIÓN DE GAMETOS La ovulación se estimula mediante la alteración de las relaciones hormonales existentes. Para las mujeres que están en anovulación (que no ovulan), estas técnicas solas pueden bastar para permitir la concepción. En general, se han usado tres de terapias para estimular la producción de gametos. La primera consiste en la administración de citrato de clomifeno, un antiestrógeno no esteroide que compite con los estrógenos por los lugares de unión en la hipófisis y posiblemente en el hipotálamo y los ovarios. Este fármaco suprime la retroalimentación negativa normadle los estrógenos en la hipófisis, de lo que resulta un aumento en las concentraciones séricas de LH y FSH. El resultado característico es una ovulación múltiple; esto es un resultado deseado para la fertilización artificial porque fertilizar más de un óvulo a la vez es más eficiente. En ocasiones, sin embargo, una mujer que ha usado citrato de clomifeno para la inducción de la ovulación produce embarazos múltiples, pues se han registrado varios casos de nacimientos de quíntuples a séptuples. Otros métodos de inducción de la ovulación son la aplicación de la gonadotropina menopáusica humana (GMh) o la administración pulsátil de hormona liberadora de gonadotropinas (GnRH). Estas técnicas son más costosas que la administración del clomifeno. OBTENCIÓN DE LOS GAMETOS Para la inseminación artificial in vivo o la fecundación artificial in vitro, es corriente obtener espermatozoides por masturbación. No obstante, la recogida de los óvulos requiere de asistencia tecnológica. La monitorización continua del curso de la ovulación inducida se logra con la aplicación de técnicas de imagen, en especial la ecografía diagnóstica. La recogida real de los oocitos implica su aspiración de los folículos maduros. Aunque originalmente se hacía por laparoscopia (observación directa mediante la inserción de un laparoscopio a través de una pequeña incisión en la pared abdominal de la mujer), la obtención se hace en la actualidad bajo guía ecográfica. Se inserta una aguja de aspiración en cada folículo maduro y se ejerce una ligera succión con una jeringa; después los oocitos se ponen en un medio de cultivo en preparación para la fertilización in vitro.
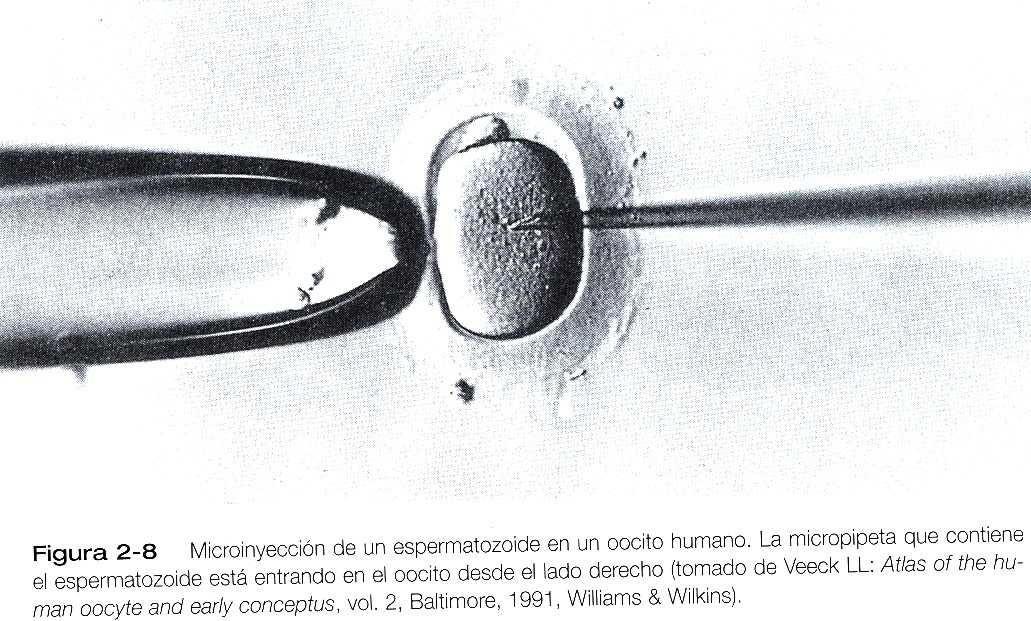
Tratamiento de la infertilidad mediante in vitro y transferencia embrionaria (continuación) ALMACENAMIENTO DE LOS GAMETOS Aunque los óvulos y los espermatozoides suelen ponerse juntos poco después de obtenerlos, en algunas circunstancias los gametos (en especial los espermatozoides), se almacenan durante períodos variables antes de usarlos. Si se llevan los espermatozoides suspendidos en preparación de glicerina a la temperatura del nitrógeno líquido, pueden conservarse durante años sin que pierdan su poder fertilizante normal. La congelación de óvulos es posible, pero mucho más problemática. FECUNDACIÓN Y CULTIVO DE EMBRIONES IN VITRO Los tres ingredientes para una fertilización in vitro con éxito son: 1) óvulos maduros, 2) espermatozoides activos normales y 3) un ambiente de cultivo apropiado. Uno de los factores más importantes para obtener una fecundación in vitro con éxito es conseguir oocitos maduros apropiados. Los óvulos aspirados de una mujer suelen encontrarse en diferentes etapas de madurez. Los óvulos inmaduros se cultivan durante un tiempo corto para que sean más fértiles. Los óvulos aspirados están rodeados por la zona pelúcida, la corona radiada y una cantidad variable de tejido del cúmulo. Los espermatozoides, ya sean frescos o congelados, se preparan separándolos los más posible del líquido seminal. Éste reduce su capacidad de fecundación, en parte porque contiene factores discapacitados. Después de la capacitación, que en los seres humanos puede lograrse exponiendo los espermatozoides a ciertas soluciones iónicas, se añade un número definido de espermatozoides al cultivo en concentraciones de 10.000 a 500.000 /mL. Las tasa de fecundación in vintro varían de un centro a otro, pero el 75% representa un promedio realista. En casos de infertilidad causada por oligospermia (muy pocos espermatozoides) o por porcentajes demasiados altos de formas anormales de espermatozoides, es posible obtener múltiples eyaculados durante un período prolongado. Éstos se congelan y se juntan para obtener un número adecuado de espermatozoides viables. En algunos casos se microinyectan pocos espermatozoides en el espacio perivitelino en el interior de la zona pelúcida. Aunque este procedimiento puede compensar el número demasiado pequeño de espermatozoides viables, introduce el riesgo de polispermia pues se elude la función normal de control de la zona pelúcida. una variación reciente de la fertilización in vitro es la inyección directa de un espermatozoide dentro de un oocito (figura 2_8). Esta técnica se ha usado en casos de grave deterioro de la función o de la cantidad de espermatozoides. Por fortuna, la división in vitro de los embriones humanos tiene mayos porcentaje de éxitos que la mayoría de las especies mamíferos. Suele permitirse que los embriones se desarrollen hasta la etapa de dos a ocho células antes de considerarlos preparados para implantarlos en el útero. En general, todos los óvulos que se obtienen de las ovulaciones múltiples de una mujer se fecundan in vitro durante el mismo período. Existen razones muy prácticas para hacerlo. Una es que a causa de la baja tasa de éxito de transferencia embrionaria, es aconsejable implantar más de un embrión (por lo común hasta tres) en el útero en una sola vez. Otra razón es económica y también tiene que ver con la baja tasa de de éxito de la transferencia embrionaria. Los embriones diferentes de los quien se usan en el procedimiento inicial se almacenan para uso futuro si la primera transferencia embrionaria resulta fallida. Esta reserva ahorra gran cantidad de tiempo y de miles de dólares a la paciente.
Tratamiento de la infertilidad mediante in vitro y transferencia embrionaria ( continuación)
PRESERVACIÓN DE EMBRIONES Los embriones preservados para su posible uso se tratan con crioprotectores (por lo general glicerol o dimetilsulfóxido) para reducir el daño por la formación de cristales de hielo. Se llevan lentamente a temperaturas muy bajas (casi siempre inferiores a -100ºC) para detener toda actividad metabólica. El tiempo durante el cual deben mantenerse los embriones congelados y el procedimiento para tratarlos si la primera implantación tiene éxito son aspectos con implicaciones tanto técnicas como éticas.
TRANSFERENCIA EMBRIONARIA A LA MADRE La transferencia del embrión a la madre es un paso sencillo desde el punto de vista técnico; sin embargo, en todo el proceso, éste es el paso que esta sujeto a un mayor tasa de fracaso. Por los común sólo del 10 al 25% de los intentos de transferencia embrionaria dan como resultado un embarazo viable. La transferencia embrionaria suele realizarse mediante la introducción de un catéter del cérvix hasta la cavidad uterina y después se expulsa el embrión o los embriones del catéter. La paciente permanece inmóvil, preferiblemente acostada durante varias horas después de la transferencia. Las razones para la baja tasa de éxito de las transferencia embrionarios no son del todo claras, pero también es posible que el número de embarazos término después de la fecundación normal in vivo sea sólo de aproximadamente un tercio. Si se produce una implantación normal y terminar en un nacimiento igualmente normal.
TRANSFERENCIA INTRATUBÁRICA Ciertos tipos de infertilidad se deben a factores como un moco cervical hostil y anomalías patológicas o anatómicas de los extremos dístales de las trompas uterinas. Un método no poco más sencillo para superar estas afecciones es introducir gametos femeninos y masculinos directamente en el extremo distal de la trompa uterina (a menudo en la unión entre las regiones ístmica y ampular). La fecundación tiene lugar en el interior de la trompa, y los acontecimientos iniciales de la embriogénesis siguen un curso natural. El método de transferencia intratubárica de gametos (GIFT) ha dado como resultado porcentajes un poco más elevados de embarazos que la fertilización in vitro y la transferencia embrionaria estándar.
MADRES SUSTITUTAS En algunas circunstancias una mujer puede producir óvulos fértiles pero no puede quedar embarazada. En ejemplo sería el de una mujer a quien se ha extirpado el útero pero que conserva sus ovarios funcionales. Una opción en este caso es la fertilización in vitro con transferencia embrionaria, pero el embrión se transfiere al útero de otra mujer (madre sustituta). Desde la perspectiva biológica, este procedimiento difiere poco de la transferencia embrionaria en el útero de la madre biológica, pero introduce muchos aspectos sociales, éticos y legales.
|
PREGUNTAS DE REPASO
1) De las barreras a la supervivencia de los espermatozoides y a su transporte dentro del tracto reproducir femenino, el pH bajo es muy importante en:
A. La parte distal de trompa uterina
B. La parte proximal de la trompa uterina
C. La cavidad uterina
D. El cérvix
E. Huérfana
2) La principal fuente de energía para los espermatozoides eyaculados es:
A. La fosfata ácida prostática
B. La glucosa interna
C. El ácido cítrico prostático
D. La fructuosa del líquido de las vesículas seminales
E. El glucógeno liberado del epitelio vaginal
3) ¿Cuál es el principal estímulo hormonal para la ovulación?
4) ¿Qué es la capitación?
5) ¿Dónde tiene lugar la fecundación?
6) Nombre dos funciones de la proteína ZP3 de la zona pelúcida
7) ¿Qué es la polispermia, y cómo se evita después de que el espermatozoide entra en el óvulo?
8) Una mujer da a la luz unos séptuples. ¿Cuál es la razón probable para el embarazo múltiple?
9) Cuando múltiples oocitos obtenidos por laparoscopia se fecundan in vitro, ¿Por qué suelen congelarse los restantes?
10) ¿Por qué algunos centros de tecnología de la reproducción insertan espermatozoides bajo la zona pelúcida o incluso directamente dentro del oocito?
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}