3
SEGMENTACIÓN E IMPLANTACIÓN
El acto de la fecundación libera al óvulo de un metabolismo aletargado y evita su desintegración posterior en el tracto reproductor. Inmediatamente después de la fecundación, el cigoto sufre un marcado aumento del metabolismo y comienza la segmentación (varios días de divisiones). Durante este tiempo, el embrión, todavía encerrado en su zona pelúcida, es transportado desde la trompa hacia el útero. Unos seis días más tarde, el embrión se desprende de la zona pelúcida y se ancla al endometrio.
Con el desarrollo intrauterino y la conexión placentaria entre el embrión y la madre, en los mamíferos superiores, incluidos los seres humanos, han evolucionado modos de desarrollo precoz muy diferentes de los de la mayoría de los invertebrados y los vertebrados inferiores. Los huevos de los animales inferiores, que éstos ponen fuera del cuerpo, deben contener todos los materiales que el embrión necesita para alcanzar la etapa de alimentación independiente. Han surgido dos estrategias principales; la primera consiste en completar el desarrollo precoz lo más pronto posible, estrategia que ha sido adoptada por las moscas Drosophila, los erizos de mar y muchos anfibios. Esta estrategia implica el almacenamiento de una cantidad moderada de mucha de la maquinaria molecular necesaria para que el embrión pase rápidamente por la segmentación hasta el comienzo de la gastrulación. Los oocitos de esas especies suelen producir y almacenar abundantes cantidades de ribosomas, ácido ribonucleico mensajero (RNAm) y de transferencia (RNAt). Éstos representan productos de los genes maternos, y ello significa que el desarrollo precoz en estas especies está ante todo bajo el control del genoma materno. La otra estrategia de desarrollo independiente, adoptada por las aves y los reptiles, consiste en la producción de un huevo de gran tamaño que contiene yema suficiente para que el desarrollo precoz se realice a un ritmo más lento. Esto elimina la necesidad de que el oocito sintetice y almacene grandes cantidades de RNA y ribosomas ante la fecundación.
La embriogénesis de los mamíferos utiliza unas estrategias fundamentalmente diferentes de las que usan los vertebrados inferiores. Puesto que la conexión placentaria con la madre evita la necesidad de que el oocito en desarrollo almacene grandes cantidades de yema, los huevos de los mamíferos son muy pequeños. No obstante, debido a que el desarrollo de los mamíferos es interno y el embrión recibe la nutrición de su madre, la segmentación es un proceso prolongado que suele coincidir con el tiempo requerido para transportar al embrión precoz desde el lugar de la fecundación en la trompa de uterina hasta el lugar de la implantación en el útero. Una de las innovaciones importantes en la embriogénesis inicial de los mamíferos es la formación del trofoblasto, el tejido especializado que forma la interfaz trófica entre el embrión y la madre, durante el periodo de segmentación. La placenta representa la manifestación última de los tejidos trofoblásticos.
SEGMENTACIÓN. Morfología
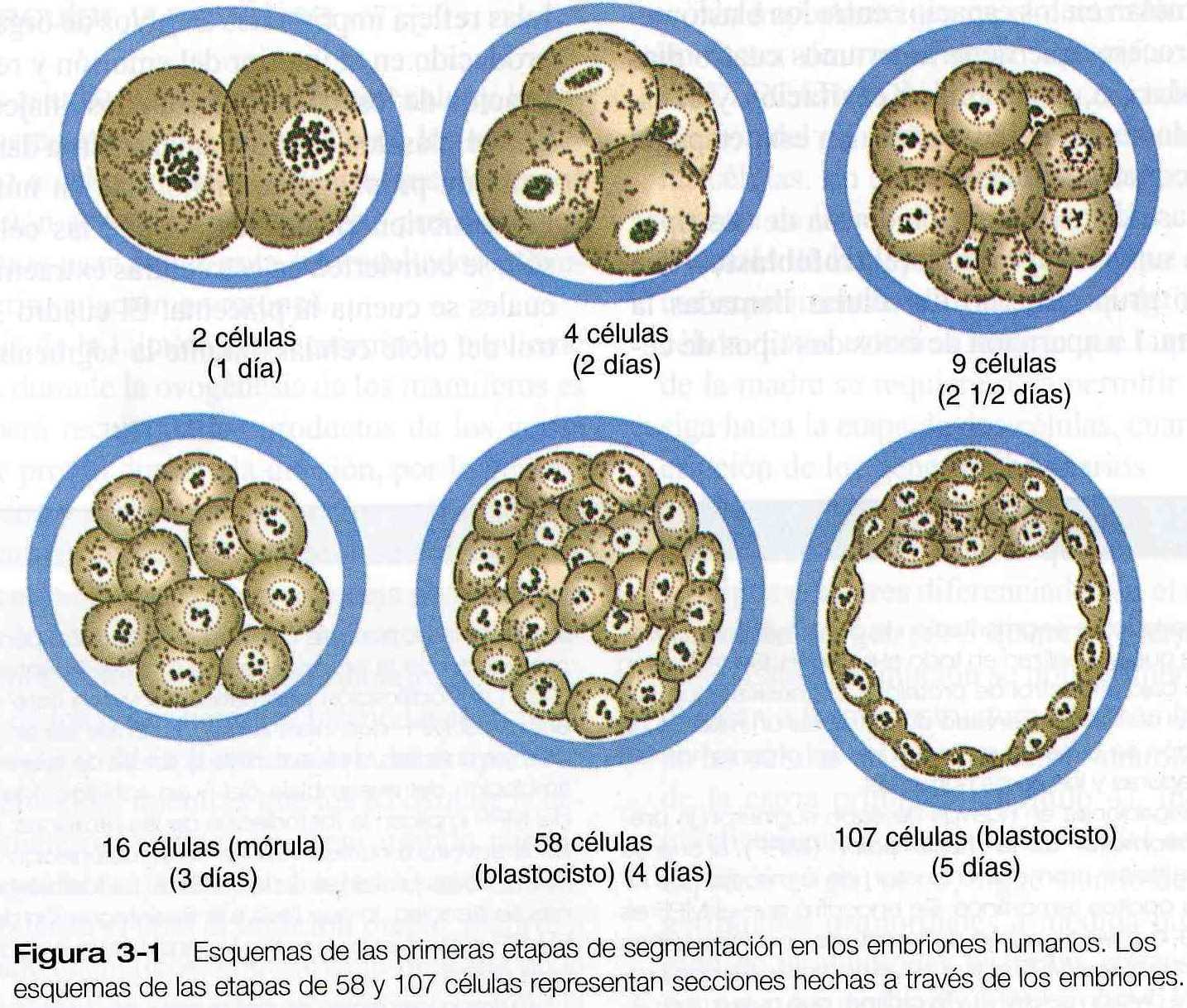
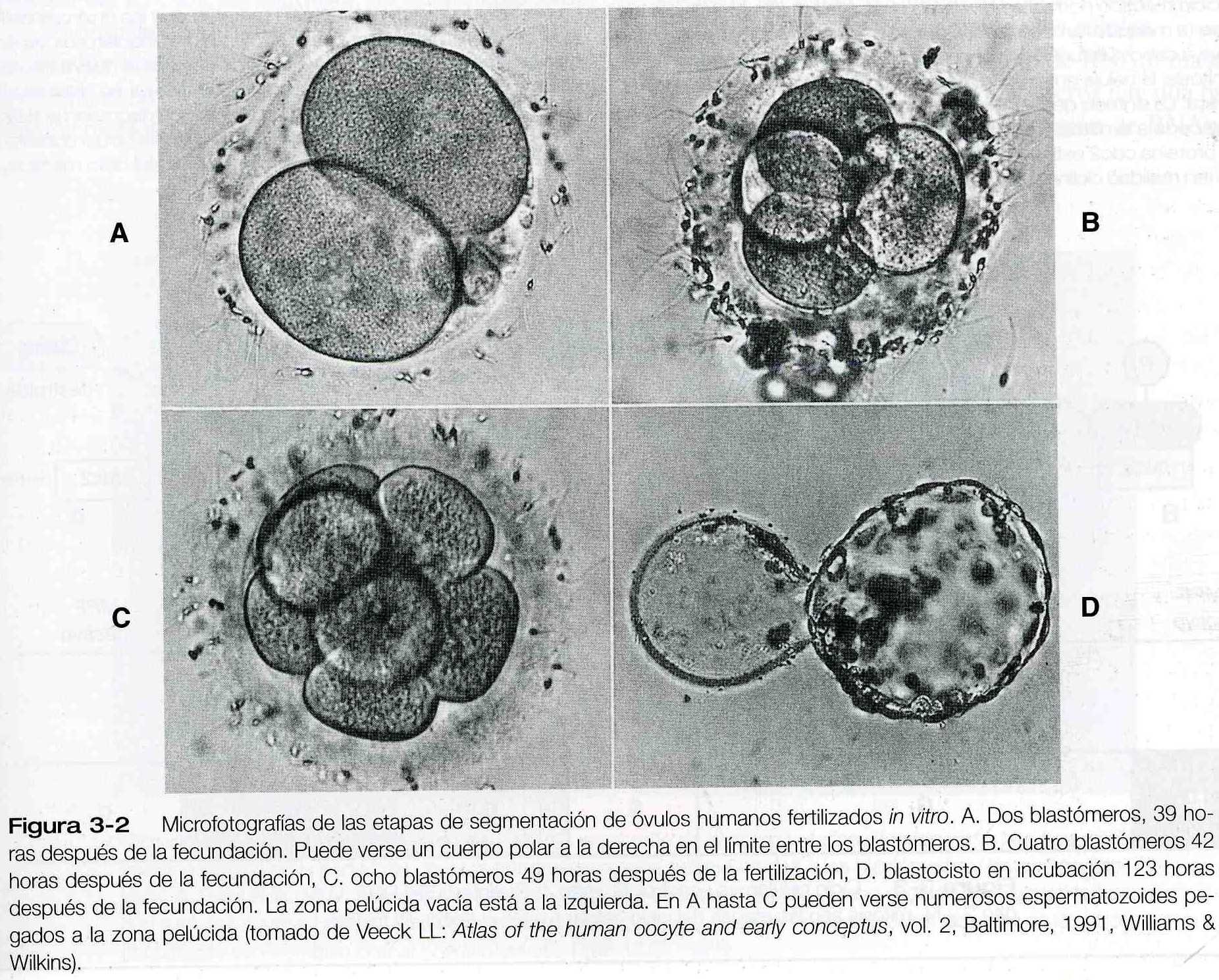
En comparación con la mayoría de las demás especies la segmentación en los mamíferos es un proceso lento que se mide en días y no en horas. El desarrollo procede a una velocidad de cerca de una división por segmentación y por día durante los primeros tres a cuatro días (figuras 3_1 y 3_2).
Después de la etapa de dos células, la segmentación de los mamíferos es asincrónica, es decir, que una de las dos células (blastómeros) se divide para formar un embrión de tres células. Cuando el embrión consta de unas 16 células, recibe el nombre de mórula ( derivado de las palabras griega y latina que significan “mora”).
Después de varias divisiones por segmentación, los embriones de los mamíferos placentados entran en una fase llamada de compactación, durante la cual los blastómeros exteriores se aprietan entre si por medio de uniones densas y comunicantes (tigh and gap junctions) y pierden su identidad individual cuando se ven desde la superficie. A través de la actividad de sistemas de transporte de Na basados en la bomba de Na+ K+ ATPasa, el Na+ y el H2O se desplazan a través de los blastómeros exteriores que ahora se parecen a un epitelio y se acumulan en los espacios entre los blastómeros internos.
Este proceso, que tiene lugar unos cuatro días después de la fecundación, se denomina cavitación y el espacio lleno de liquido se llama blastocele. En esta capa, el embrión se conoce como blastocisto.
En la etapa de blastocisto e embrión consta de dos tipos de células: una capa superficial externa (el trofoblasto), que rodea a un pequeño grupo interno de células llamadas la masa celular interna. La aparición de estos dos tipos de células refleja importantes cambios de organización que se ha producido en el interior del embrión y representa la especialización de los blastómeros en dos linajes celulares distintos.
Las células de la masa celular interna dan lugar al cuerpo del embrión propiamente dicho más un número de estructuras extraembrionarias, mientras que las células del trofoblasto sólo se encuentran en la placenta. El cuadro 3_1 describe el control del ciclo celular durante la segmentación.
Cuadro 3-1. Control del ciclo celular durante la segmentación
El sello distintivo del periodo de segmentación lo constituyen las sucesivas ondas de mitosis que se realizan en todo el embrión. Cada división mitótica se encuentra bajo el control de proteínas tan básicas que sus estructuras y funciones se han preservado durante casi un millardo de años. Esta interpretación de basa en su presencia en organismos tan distintos como las levaduras y los seres humanos.
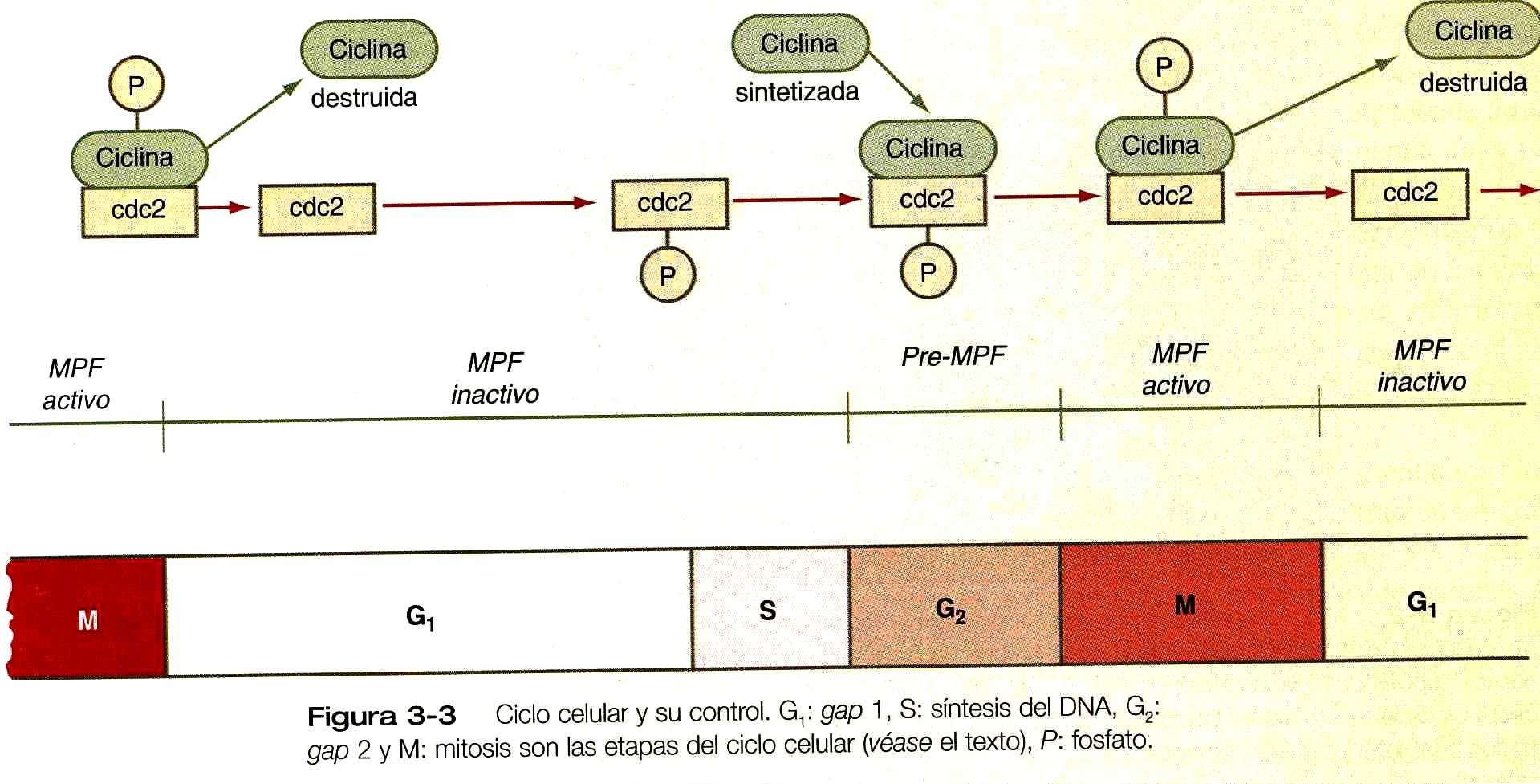
Las primeras investigaciones en huevos de sapo sugirieron la presencia de un factor promotor de la maduración (MPF), al cual se suele denominar ahora factor promotor (inductor) de la mitosis. Investigaciones más profundas revelaron que el MPF activo es un complejo de dos proteínas, la cdc2 (ciclo de división celular) y la cíclica, que guía a una célula a lo largo de su ciclo mitótico.
El ciclo mitótico normal se divide en cuatro fases (figura 3_3). La interfase (a menudo llamada fase G1) es el periodo durante el cual la célula lleva a cabo sus funciones asignadas. A medida que se prepara para la mitosis , la célula entra en la fase S, durante la cual su DNA nuclear se replica. La síntesis del DNA va seguida de una breve fase G2 (GAP2) que procede a la mitosis real (fase M).
La proteína cdc2 está presente durante todo el ciclo de la mitosis. La ciclina (en realidad ciclina B) se sintetiza y se acumula en la célula durante la interfase, pero se combina con la cdc2 para formar un factor de promoción de la premaduración (pre-MPF) antes de la mitosis (figura 3_3).
La modificación enzimática convierte este complejo en la forma activa del MPF que inicia la mitosis. Entre las acciones específicas de MPF se cuentan la iniciación de la rotura de la envoltura nuclear y la estimulación del ensamblaje del huso mitótico. Muchas de las acciones del MPF implican la fosforilación de las proteínas. Por ejemplo, la rotura de la envoltura nuclear resulta de la fosforilación de láminas nucleares, o de las proteínas de la cubierta. La fosforilación hace que las láminas se disocien, lo que lleva a la desintegración de la envoltura nuclear.
El MPF activo también activa las enzimas que degradan de forma brusca la ciclina.
Cuando los niveles de ciclina caen por debajo de un cierto umbral, la proteína cdc del MPF pierde su actividad, con lo cual termina la mitosis.
La pérdida de la actividad del MPF permite que las enzimas de la fosfatasa celular retiren los grupos fosfato que se añadieron a la proteína bajo la influencia del MPF. Un efecto de esto es la nueva formación de la cubierta nuclear cuando las láminas nucleares se desfosforilan. Las fosfatasas también inactivan a las enzimas que degradan la ciclina, y esto hace que la ciclina se acumule de nuevo en la célula durante la interfase. Esto crea el escenario para la repetición del ciclo mitótico.
BIOLOGÍA MOLECULAR Y GENÉTICA
La mayoría de los estudios de la biología molecular y la genética de las primeras etapas del desarrollo de los mamíferos se han llevado a cabo en ratones. Hasta que contemos con más información sobre la embriogenésis precoz de los primates, deberemos usar como guía los resultados provenientes de ka experimentación en ratones.
La consecuencia de la falta de almacenamiento previo de ribosomas y RNA durante la ovogénesis de los mamíferos es que el cigoto deberá recurrir a los productos de los genes embrionarios muy pronto durante la división, por lo general hacia la etapa de dos o cuatro células ( cuatro a ocho células en los seres humanos) . Sin embargo, no parece haber una aguda transición entre el momento en que deja de recurrirse a los productos de los genes puramente maternos y la iniciación de la transcripción del genoma embrionario. Por ejemplo, los productos de los genes paternos (como la isoformas de la b--glucuronidasa y de la b2-microglobilina) aparecen muy pronto en el embrión, mientras que los RNAm de la actina y la histona maternas todavía se están usando para la producción de las proteínas correspondientes. Como una indicación del grado hasta el cual el embrión precoz recurre a sus propios productos genéticos, el desarrollo más allá de la etapa de dos células no tiene lugar en el ratón si se inhibe la transcripción del RNAm. En contraste, un tratamiento similar en los embriones de anfibios no interrumpe el desarrollo hasta la división tardía, tiempo en el cual los embriones comienzan a sintetizar el RNAm necesarios para controlar los movimientos morfogenéticos y la gastrulación.
Un importante gen en el desarrollo incipiente es el oct-3,un factor de transcripción especifico que fija el octámero ATTTGCAT al DNA. Hay una estrecha relación entre la expresión del gen oct-3 y el estado altamente indiferenciado de las células. En el ratón, el oct-3 derivado de la madre se encuentra en los oocitos en desarrollo y es activo en el cigoto.
Después de la pérdida de la proteína oct-3 inducida de forma experimental, el desarrollo se detiene en la etapa de una célula. Estos estudios indican que la proteína oct-3 derivada de la madre se requiere para permitir que el desarrollo prosiga hasta la etapa de dos células, cuando comienza la transcripción de los genes embrionarios.
El gen oct-3 se expresa en todos los blastómeros hasta la etapa de mórula. A medida que comienzan a aparecer diversos tipos celulares diferenciados en el embrión, el nivel de la expresión del gen oct-3 disminuye hasta que no puede detectarse. Esta disminución se nota primero en las células que se dedican a formar estructuras extraembrionarias y por último en las células de las etapas embrionarias a mediada que surgen de la estría primitiva (capitulo 4). Incluso después de que prácticamente todas las células del embrión han dejado de expresar el gen oct-3, sigue siendo detectable en las células germinales primordiales a medida que migran desde la región de la alantoides hacia las crestas gonadales. Debido a sus patrones de distribución, se cree que la proteína oct-3 desempeña un papel regulador en las primeras decisiones de determinación o diferenciación de las células.
Junto con una pequeña cantidad de RNAm materno preformado, en los embriones de mamíferos hay una baja capacidad correspondiente para la traducción de RNAm.
Diversos experimentos con inyección sugieren que el factor que limita la efectividad de la traducción puede se el pequeño número de cromosomas almacenado en el óvulo.
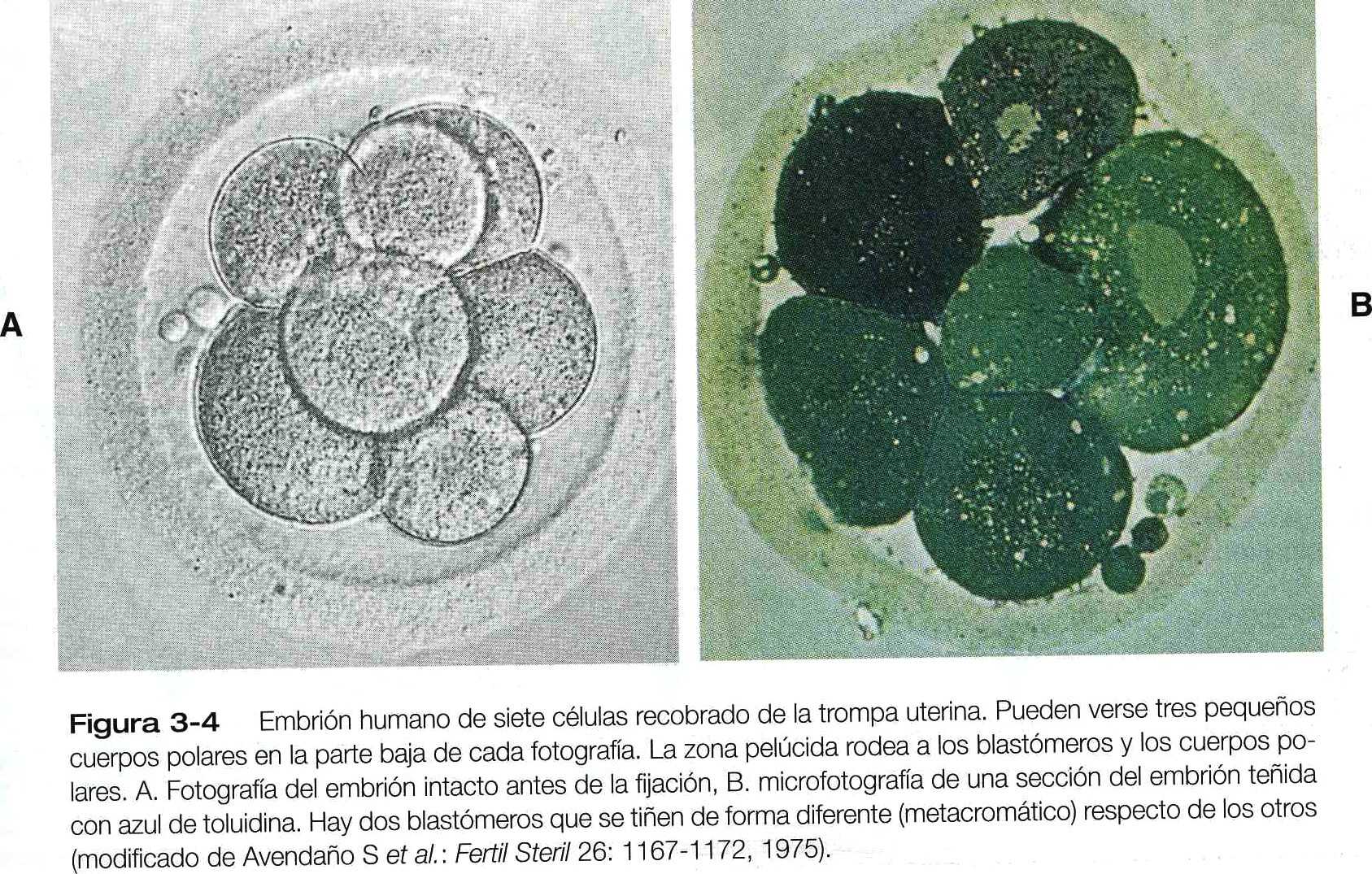
Ni siquiera en una etapa precoz los blastómeros de un embrión en segmentación son homogéneos. Los métodos de tinción simple revelan pronunciadas diferencias entre las células en los embriones humanos incluso desde la etapa de siete células (figura 3_4). Los estudios autorradiográficos han demostrado que todos los blastómeros de los embriones humanos de cuatro células tienen niveles bajos de síntesis de RNA extranucleolar y no nucleolar. En la etapa de ocho células, algunos blastómeros tienen niveles muy altos de síntesis de RNA, pero otros blastómeros todavía presentan el patrón que se ve en los blastómeros de los embriones de cuatro células. Los estudios morfológicos muestran diferencias correspondientes entre los blastómeros activos y los inactivos desde el punto de vista de la transcripción.
Una vez que comienza la segmentación, los productos de la transcripción de los cromosomas provenientes tanto del padre como de la madre son activos en la guía del desarrollo. Por lo general los embriones haploides mueren durante la segmentación o poco después de la implantación. Sin embargo, hay crecientes evidencias de que el control del desarrollo precoz implica algo más que un juego diploide de cromosomas en cada célula.
IMPRONTA DE LOS PADRES
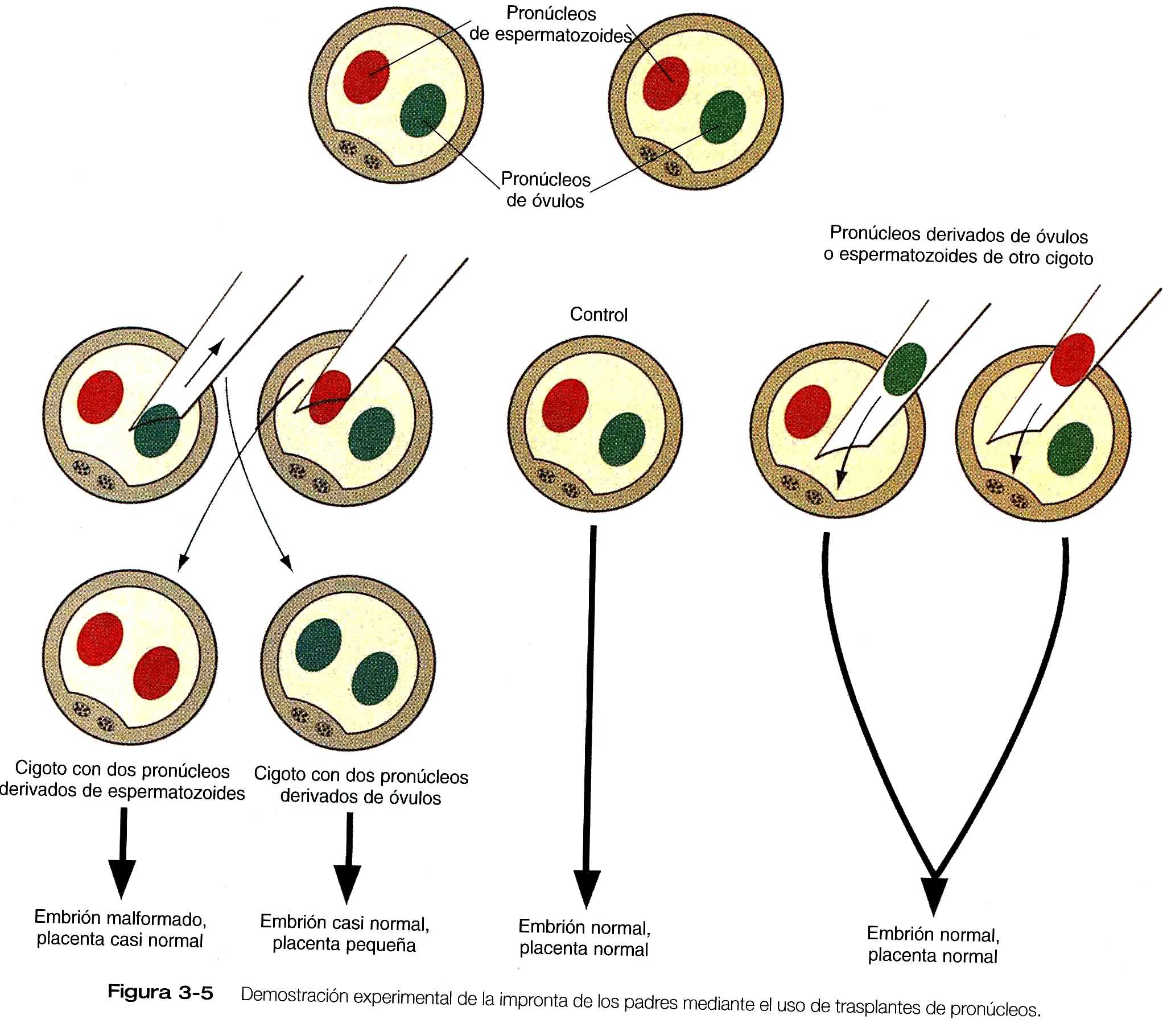
La experimentación, junto con la observación de algunos trastornos del desarrollo en ratones y seres humanos, ha demostrado que la expresión de ciertos genes derivados del óvulo difieren respecto de la expresión de los mismos genes derivados del espermatozoide. Los efectos de este fenómeno, llamado impronta de los padres, se manifiestan de diversas maneras.
Es posible eliminar un pronúcleo de un óvulo de ratón recién inseminado y remplazarlo por un pronúcelo tomado de otro óvulo que esté en una etapa similar del desarrollo (figura 3_5). Si un pronúcleo masculino o femenino es reemplazado por un pronúcleo correspondiente masculino o femenino, el desarrollo es normal. No obstante, si se extrae un pronúcleo masculino y se remplaza por un femenino ( de lo que resulta un cigoto con dos pronúcleos femeninos), el embrión en si mismo se desarrolla de una forma bastante normal, pro la placenta y el saco vitelino tienen un desarrollo deficiente. Por el contrario, un cigoto con dos pronúcleos masculinos produce un embrión gravemente malformado, mientras que la placenta y el saco vitelino son casi normales.
La impronta paterna inactiva de forma selectiva algunos genes implicados en el desarrollo del cuerpo del embrión, mientras que la impronta materna inactiva algunos genes implicados en la formación de estructuras extraembrionarias como la placenta. La impronta de los padres tiene lugar durante la gametogénesis a través de la metilación de ciertas bases ( en especial, citosinas en secuencias de CpG). Esta metilación redunda en la expresión diferencial de alelos paternos y maternos de los genes troquelados ( que llevan la impronta) durante el desarrollo embrionario. Los genes troquelados operan durante el desarrollo y posiblemente hasta la edad adulta, pero una impronta dada no pasa a la progenie de ese individuo; en vez de ello, las improntas de los padres troquelados en los genes en los gametos se borran y se establecen unas nuevas, que corresponden al sexo de ese individuo, en los óvulos o los espermatozoides durante la gametogénesis.
No todos los genes llevan una impronta de los padres. Hasta ahora, se han identificado menos de dos docenas de genes troquelados, pero los cálculos sugieren que hasta 500 de los casi cien mil genes humanos pueden ser afectados por el troquelado (tabla 3_1). En la Correlación clínica 3-1 se examinan algunas situaciones y síndromes asociados con la impronta de los padres.
Tabla 3_1 efectos diferenciados de la expresión de genes paternos y maternos
|
Función |
Impronta Paterna |
Impronta Materna |
|
Crecimiento global |
Estimulado (IGF-2) |
Reducido (H-19) |
|
Comportamiento de las células madre |
PROLIFERACIÓN |
diferenciación |
|
Refuerzo de la diferenciación |
músculo |
epidermis |
|
Localización de la actividad en el cerebro |
hipotálamo |
Neocórtex |
INACTIVACIÓN DEL CROMOSOMA X
Oro ejemplo de la desigualdad de la expresión genética durante el desarrollo precoz es el patrón de inactivación del cromosoma X en los embriones femeninos.
A partir de los estudios citogenéticos es bien sabido que uno de los dos cromosomas X de las células femeninas se inactiva mediante intensa condensación. Ésta es la base de la cromatina sexual o el corpúsculo de Barr que puede demostrarse en las células femeninas pero no en las masculinas normales.
El propósito de la inactivación del cromosoma X es compensar la dosificación, o preservar las células de un exceso de productos genéticos del cromosoma X.
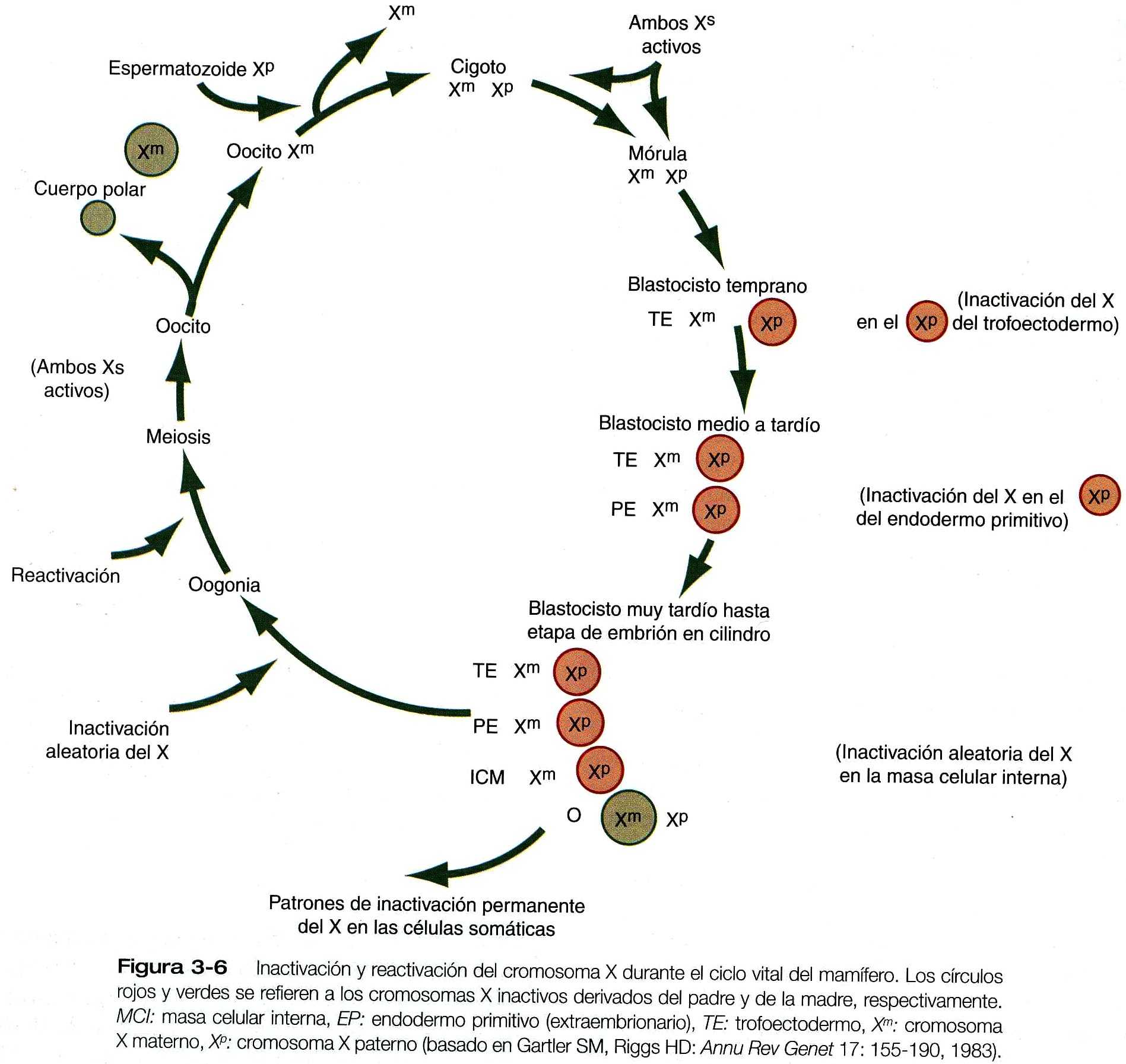
Los estudios de los niveles de enzimas codificados en el DNA del cromosoma X en ratones han demostrado que ambos cromosomas X son activos desde el punto de vista de la transcripción durante la segmentación inicial de los embriones femeninos. Después de la diferenciación de los blastómeros ya sea en trofoblasto o en masa celular interna, el patrón cambia (figura 3_6). Ambos cromosomas X continúan activos en las células de la masa celular interna, mientras que en todas las células del trofoblasto (trofoectodermo), el cromosoma X derivado del padre se inactiva selectivamente. A medida que las células de la masa celular interna se subdividen en otros linajes, también se produce la inactivación diferencial del cromosoma X.
CORRELACION CLINICA 3-1
Situaciones y síndromes asociados con la impronta de los padres
Un ejemplo impresionante de la impronta paterna en el ser humano es la mola hidatidiforme (figura 6_17), que se caracteriza por el desarrollo excesivo de tejido trofoblástico y el subdesarrollo importante del embrión. Esta situación puede resultar de la fecundación de un óvulo por dos espermatozoides y la imposibilidad subsiguiente del genoma materno del óvulo de participar en el subdesarrollo, o de la duplicación de un pronúcleo masculino en el óvulo “huero” o vacío. Ésta forma de desarrollo muy anómalo es coherente con la hipótesis de que la impronta paterna favorece el desarrollo del trofoblasto a expensas del desarrollo del embrión.
Otros síndromes se basan también en la impronta de los padres. El síndrome de Beckwith-Wiedemann, que se caracteriza por macrosomía fetal y un aumento de la incidencia de cáncer en la infancia, ha sido mapeado en la región troquelada del cromosoma 11, que contiene los genes para el factor 2 de crecimiento similar a la insulina (IGF-2, que promueve la proliferación celular) y el H19 (un supresor del crecimiento). Se produce cuando ambos alelos del gen IGF-2 expresan un patrón de impronta paterna. Otro ejemplo implica la deleción del brazo largo del cromosoma 15. los niños de cualquier sexo que heredan la deleción de la madre sufren el síndrome de Angelman, que incluye retraso mental profundo, convulsiones y ataxia. Un niño que hereda del padre la deleción de la misma región sufre el síndrome de Prader-Willi, caracterizado por obesidad, corta estatura, hipogonadismo, labio superior en arco y retraso mental breve.
La inactivación X tiene lugar en último término en todas las células, y solo durante la oogénesis ambos cromosomas X de los oocitos vuelven a activarse. La inactivación selectiva del cromosoma X paterno en el trofoblasto es otro ejemplo de la impronta genética de los padres, aunque el mecanismo de la inactivación puede ser diferente.
Un gen recién descubierto, el transcriptor especifico del X inactivo (XIST), interviene en la inactivación de uno de los cromosomas X. Sus transcritos de RNA permanecen el núcleo y no forman proteínas. A través de un mecanismo de conteo, que todavía no está del todo claro, un cromosoma X rompe sus RNAm del XIST. El otro cromosoma X acumula RNAm del XIST estabilizado y posteriormente se inactiva. La inactivación del cromosoma X se produce cuando las células sufren restricción y comienzan a diferenciarse, en vez de permanecer totipotenciales (capaces de formar todo tipo de progenesis). La impronta paterna en el gen XIST explica la inactivación selectiva del cromosoma X paterno en el trofoectodermo extraembrionario y en el endodermo extraembrionario durante la segmentación (figura 3_6).
Puesto que conservan durante más tiempo su totipotencialidad, las células de la masa celular interna sólo sufren la inactivación del cromosoma X más adelante, pero no se sabe por qué en ellas se produce la inactivación al azar del cromosoma X ya sea derivado del padre o de la madre.
PROPIEDADES DEL DESARROLLO DE LOS EMBRIONES EN SEGMENTACIÓN
Se considera que la embriogénesis inicial de los mamíferos es un proceso muy regulado. La regulación es la capacidad del embrión o del primordio de un órgano de producir una estructura normal aunque se le hayan quitado o añadido partes.*
A nivel celular, esto significa que los destinos de las células de un sistema regulador no están fijos de manera irrecuperable y que las células todavía pueden responder a las influencias ambientales. Puesto que la asignación de blastómeros a diferentes linajes celulares es una de las características principales del desarrollo de los mamíferos, es importante identificar los factores ambientales implicados.
La técnica experimental más sencilla de las que se usan para demostrar las propiedades reguladoras de los embriones precoces, es separar los blastómeros de los embriones al comienzo de la segmentación y determinar si cada uno de ellos puede dar lugar a un embrión completo. Este método se ha usado para demostrar que los blastómeros únicos de embriones de dos, y en ocasiones hasta cuatro, células pueden formar embriones normales, aunque los blastómeros de capas posteriores no pueden hacerlo. En estudios en mamíferos suele tomarse una sola célula de un embrión al comienzo de la etapa de segmentación e inyectarla en el blastocele de un huésped diferente desde el punto de vista genético. Estas células inyectadas se incorporan al embrión huésped, y dan lugar a quimeras o mosaicos celulares.
Cuando se inyectan blastómeros de un donante genéticamente diferente en embriones huésped, las células del donante pueden identificarse mediante análisis histoquímico o citogenético, y se puede determinar se destino (los tejidos a que dan lugar).Los experimentos de mapeo del destino son importantes en embriología porque permiten seguir las vías a lo largo de las cuales pueden diferenciarse una célula en particular. Los experimentos de mapeo del destino celular, que incluyen varias isoenzimas de la enzima glucosa fosfato isomerasa, han demostrado que todos los blastómeros de un embrión de ocho células de ratón siguen siendo totipotenciales, es decir, que mantienen la capacidad de formar cualquier tipo de célula en el organismo. Incluso en la etapa de segmentación de 16 células, algunos blastómeros pueden producir progenies que producen tanto los linajes de la masa celular interna como el trofoblasto.
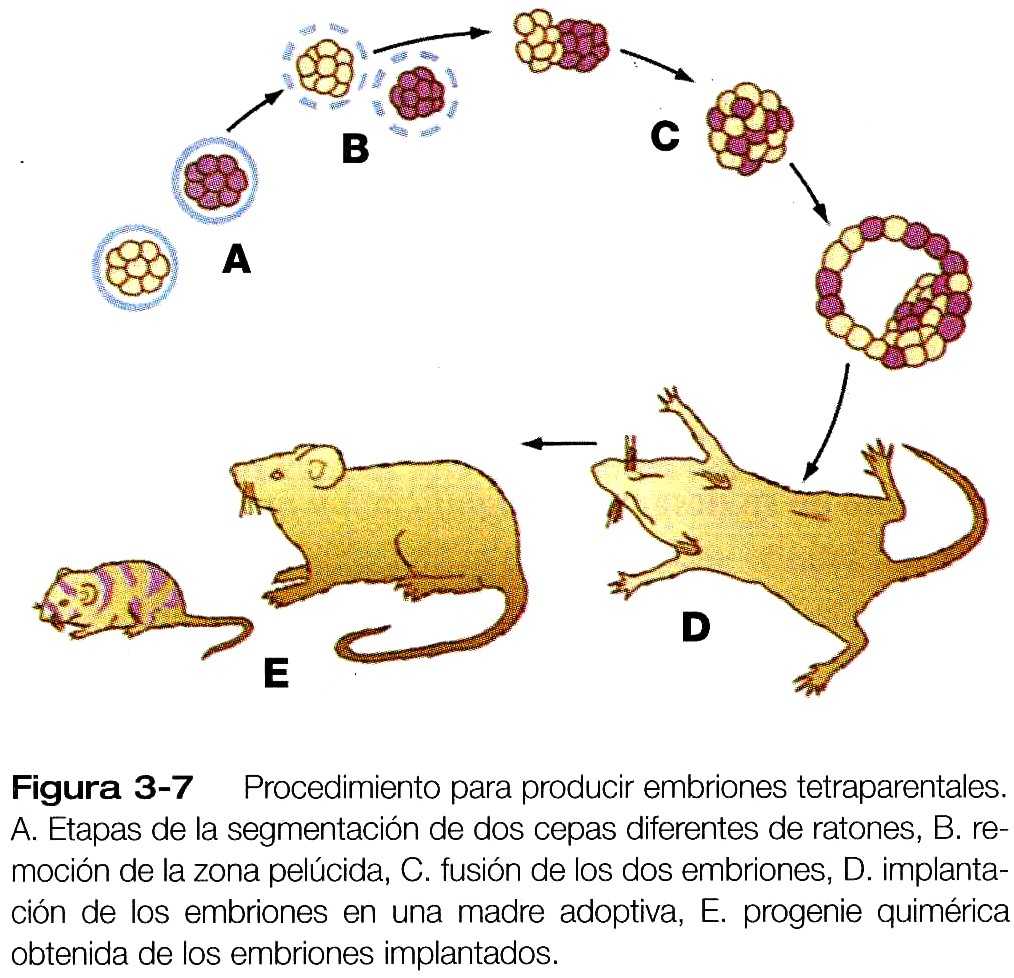
Otro método para demostrar las propiedades reguladoras de los embriones precoces de mamíferos es disociar embriones de ratón en blastómeros separados y entonces combinar los blastómeros de dos o tres embriones( figura 3_7). Los blastómeros combinados pronto se agregan y se reorganizan para transformarse en un gran embrión único, que prosigue hasta convertirse en un ratón tetra o hexaparental de apariencia normal. Mediante diversos métodos para hacer embriones quiméricos, es posible incluso combinar blastómeros para producir quimeras entre especies ( por ejemplo, una cabra-oveja).
*contrario al desarrollo regulador es el desarrollo en mosaico, que se caracteriza por la incapacidad de compensar los defectos o integrar células extra en un todo unificado. En un sistema de mosaico, los destinos de las células están determinados rígidamente, y la eliminación de células tiene como resultado la falta de un embrión o una estructura a la que le faltan los componentes que estaban destinados a formar las células que se eliminaron. La mayoría de los sistemas reguladores tienen una tendencia creciente a mostrar propiedades de mosaico a medida que el desarrollo avanza.
Uno de los pasos mas importantes en el desarrollo inicial de los mamíferos es la decisión que da como resultado la aparición de dos líneas separadas de la célula (el trofoblasto y la masa celular interna) a partir de los blastómeros tempranos. Hasta la etapa de ocho células en el ratón, todos los blastómeros son prácticamente idénticos en su potencial para el desarrollo. Poco después se notan diferencias entre las células que tienen al menos una superficie situada en el borde exterior del embrión y las que están totalmente rodeadas por otros blastómeros.
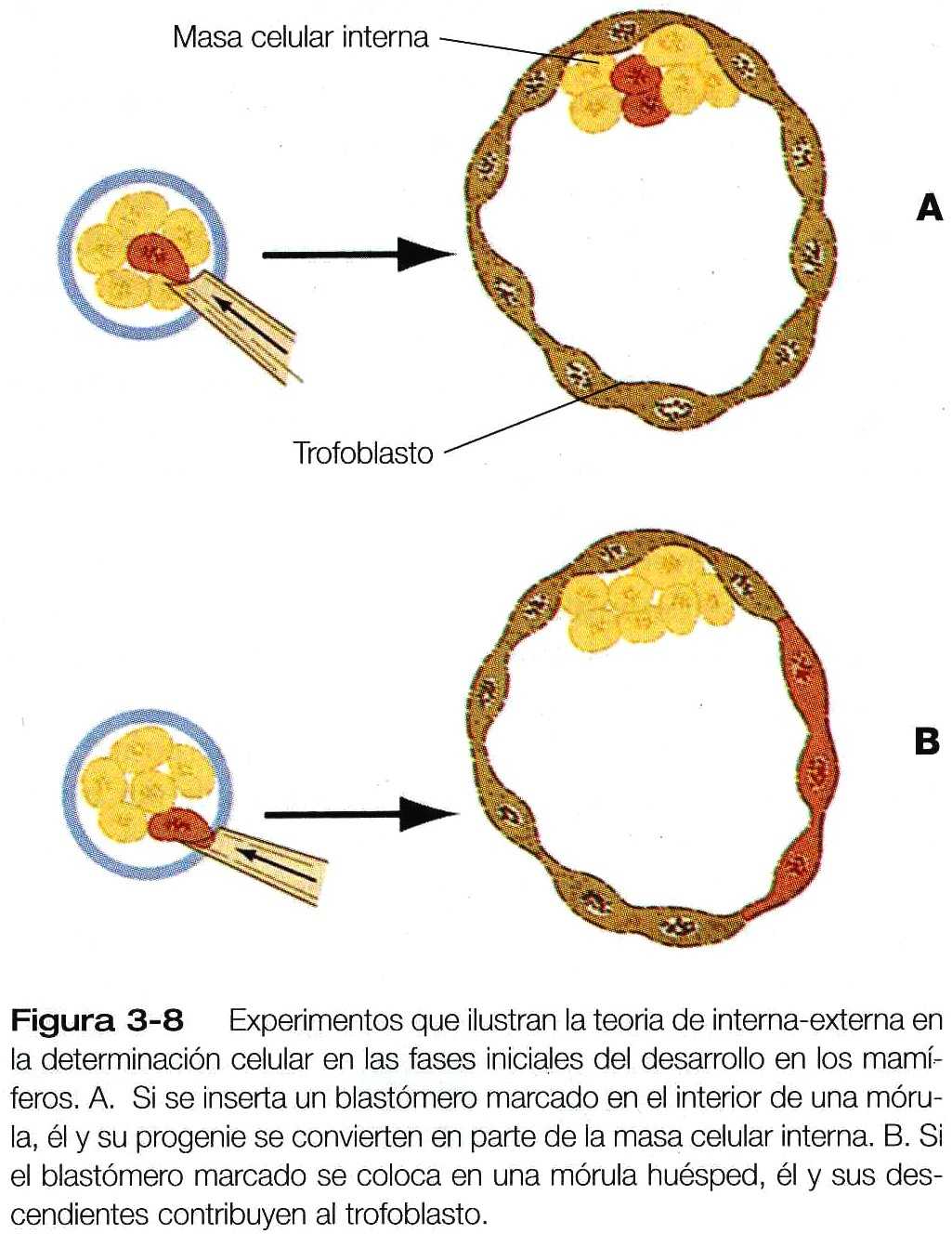
La relación entre la posición de los blastómeros y el destino final de su desarrollo se expresó a través de la hipótesis de interna-externa. Los blastómeros externos se diferencian en el trofoblasto, mientras que los internos forman la masa celular interna, de la cual surge el cuerpo del embrión. Aunque esta hipótesis ha sido confirmada por diversos experimentos, aún es esquivo y poco claro el mecanismo por el cual los blastómeros reconocen sus posiciones y después se diferencian en consecuencia. Si se colocan blastómeros marcados procedentes de embriones desagregados en la zona externa de otro embrión en fase de mórula, es característico que contribuyan a la formación del trofoblasto. Por el contrario, si las mismas células marcadas se colocan en el interior del embrión huésped, participan en la formación de la masa celular interna (figura 3_8). Las células externas del embrión precoz de los mamíferos están ligadas entre sí mediante uniones comunicantes y densas, pero no se sabe si esta característica morfológica es una causa o un efecto de su diferenciación en trofoblasto. Los experimentos de este tipo demuestran que el potencial o la potencia del desarrollo (potencia prospectiva) –los tipos de células que una célula precursora puede formar- de muchas células es mayor que el destino de su desarrollo (significación prospectiva) – los tipos de células que una célula precursora forma normalmente-.
Otra característica de los embriones de los mamíferos en etapa de segmentación es la ausencia de ejes corporales fijados rígidamente. En muchos vertebrados inferiores, el eje dorsoventral esta fijo en el óvulo antes de la fecundación y el eje anteroposterior lo determina el lugar de la penetración del espermatozoide en el óvulo en el momento de la fecundación. Por otra parte, los embriones de las aves y los mamíferos nos muestran evidencias de fijación axial hasta mucho tiempo después, durante la segmentación. En el blastocisto humano de cuatro a cinco días, que tiene una masa celular interna bien definida, la superficie dorsal es la parte de la masa celular interna que hace cuerpo con el trofoblasto externo, y la superficie ventral es la parte que se proyecta hacia la cavidad del blastocisto.
Apenas dos semanas después de la fecundación puede identificarse el eje longitudinal. El reconocimiento de los extremos craneal y caudal en el embrión (eje longitudinal) solo es posible después de la aparición de la estría primitiva al comienzo de la tercera semana.
MANIPULACIÓN EXPERIMENTAL
DE LOS EMBRIONES EN SEGMENTACIÓN
Muchos de los conocimientos sobre las propiedades del desarrollo inicial de los embriones de los mamíferos es el resultado de técnicas recientemente diseñadas para manipularlos de forma experimental. En general, estas técnicas deben combinarse con otras que se han diseñado para la fecundación in vitro, el cultivo de embriones y la transferencia embrionaria (capitulo 2).
Las estrategias clásicas para investigar las propiedades del desarrollo de los embriones son: 1) remover una parte y determinar la manera en que el resto del embrión compense la pérdida (estos experimentos se llaman de deleción o ablación) y 2) añadir una parte y determinar como el embrión integra el material añadido en su plan corporal global ( estos experimentos se llaman de deleción, la estrategia de los experimentos de adición ha probado ser mas fructífera para dilucidar mecanismos que controlan la embriogénesis de los mamíferos.
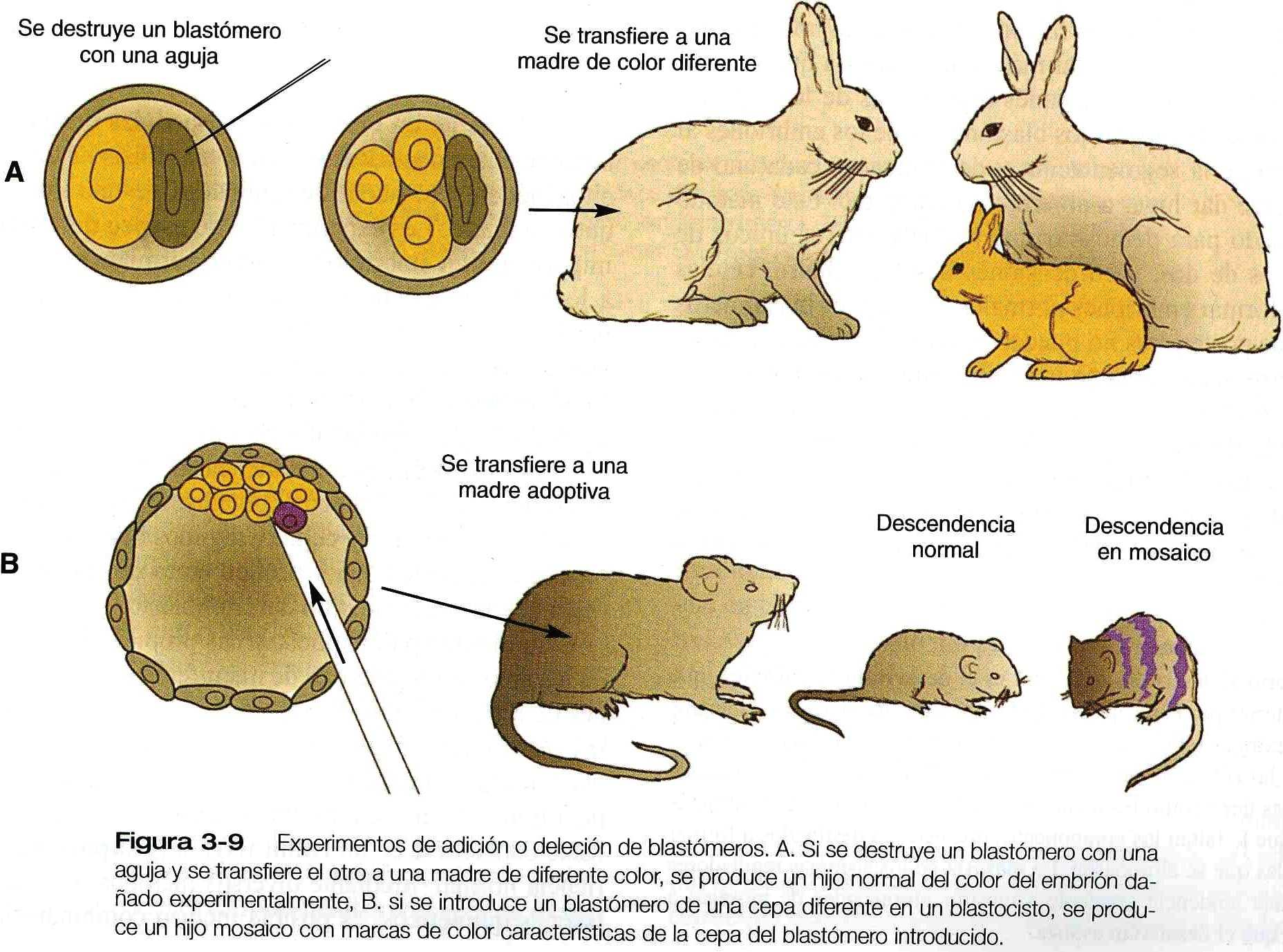
Los experimentos de deleción y adición de blastómeros (figura 3_9) han demostrado en forma convincente la naturaleza reguladora ( es decir, la fuerte tendencia del sistema a restablecer su totalidad) de los embriones precoces de los mamíferos.
Es importante conocer esto para comprender por que la exposición de los embriones humanos precoces a influencias ambientales desfavorables suele dar como resultado la muerte o un embrión normal.
Una de las técnicas experimentales mas poderosas de los últimos 20 años ha sido la inyección de células marcadas genética o artificialmente en la cavidad del blastocisto de un embrión huésped (figura 3_9 B). Esta técnica se ha usado para demostrar que las células añadidas llegan a integrase al cuerpo del embrión huésped; esto ofrece evidencias adicionales sobre la regulación embrionaria. Otra utilización de esta técnica ha sido en el estudio de los linajes celulares del desarrollo inicial del embrión. Al identificar al progenie de las células marcada inyectadas, los investigadores han podido determinar la potencia del desarrollo de las células del donante.
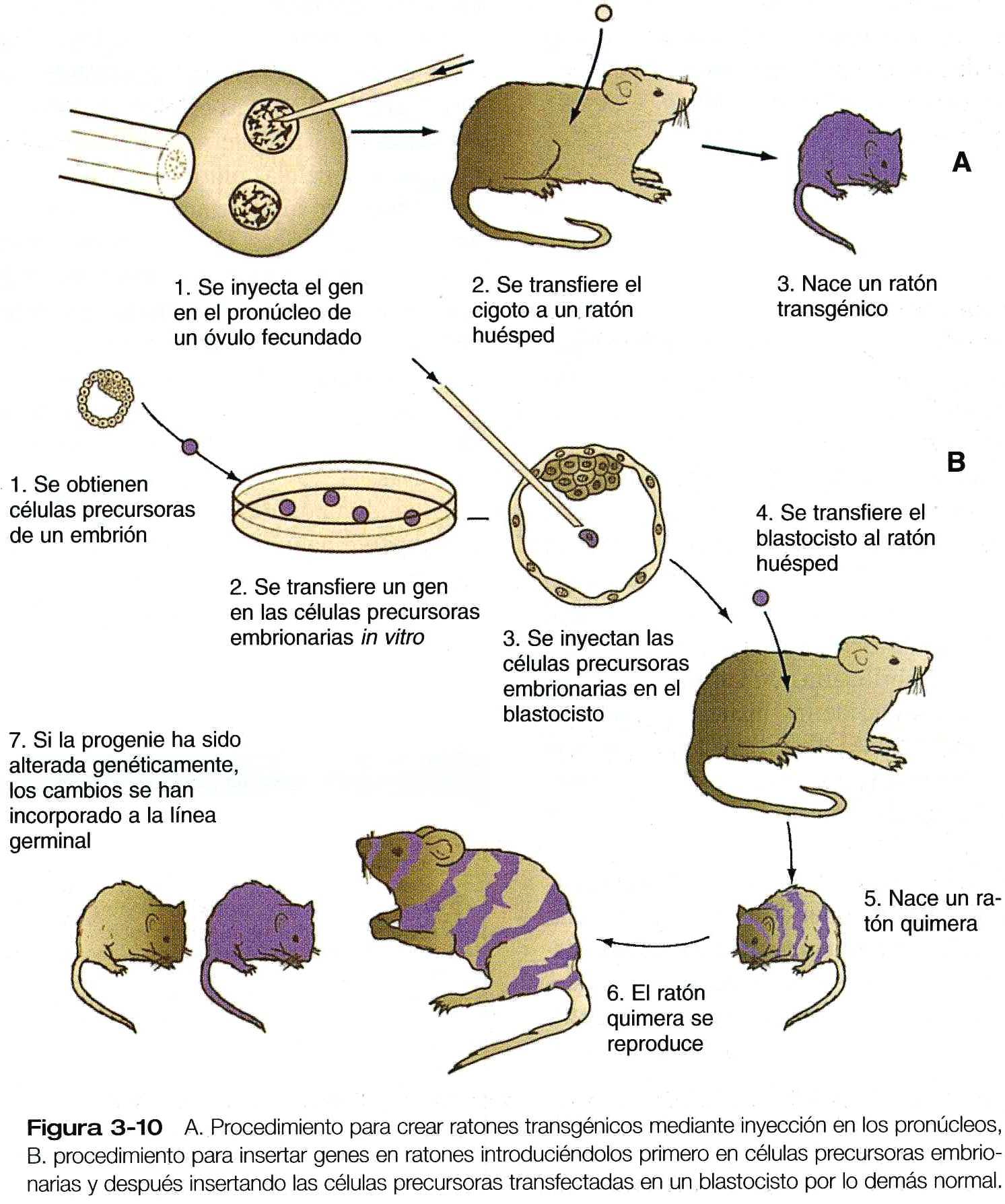
Una técnica que esta ofreciendo nuevos datos sobre los mecanismos de control genético del desarrollo de los mamíferos es la producción de embriones transgénicos. Éstos, por lo común ratones, se producen mediante la inyección directa de DNA ajeno en los pronúcleos de los cigotos(figura 3_10 A). El DNA, específicamente DNA recombinante para un gen especifico, puede fusionarse con un elemento regulador diferente que el investigador puede controlar. Por ejemplo, se han creado ratones transgenicos inyectando el gen de la hormona de crecimiento de la rata acoplado con una región promotora de metalotioneina (MT-1) en los pronúcleos de cigotos de ratón. Los cigotos que han recibido la inyección se transplantan a los úteros de madres adoptivas, que dan a luz ratones transgénicos de apariencia normal. Mas adelante, cuando estos ratones congénitos se alimentan con una dieta alta en zinc que estimula a la región promotora MT-1, se activa el gen de la hormona de crecimiento de la rata, lo que hace que el hígado produzca grandes cantidades del polipéptido de la hormona de crecimiento. La función del gen transplantado es evidente; bajo la influencia de la hormona de crecimiento de rata que estaban produciendo, los ratones transgénicos crecieron hasta un tamaño mucho mayor que el de sus compañeros de camada (figura 3_11). La técnica de producción de ratones transgénicos se usa cada vez mas tanto para examinar los factores que regulan la expresión de genes específicos en los embriones como para alterar genes en el embrión huésped. Además, la efectividad de esta técnica para corregir defectos genéticos conocidos se esta estudiando cada vez mas en los ratones.
Un importante avance tecnológico es la creación de líneas de células precursores derivadas (madre, stem cells) del embrión (células ES). Las células ES derivan originalmente de las masas celulares internas y pueden propagarse in vitro como líneas de células pluripotenciales que pueden mantenerse en un estado indiferencial o estimularlas a diferenciarse en líneas específicas. Hoy es posible introducir genes específicos en las células ES mediante ingeniería genética. Cuando estas células manipuladas genéticamente se introducen en el blastocisto, pueden incorporarse en el embrión huésped (figura 3_10B). Si la progenie de una célula ES cambiada por ingeniería genética se incorpora en la línea germinal, el rasgo genético puede pasar a las generaciones sucesivas.
Algunos tipos de embarazos gemelares representan un experimento natural que demuestra la naturaleza altamente reguladora de los embriones humanos precoces, como se describe en la correlación clínica 3-2.
Cuadro 3-2 resumen de la zona pelúcida
CORRELACIÓN CLÍNICA 3-2 Embarazo gemelar
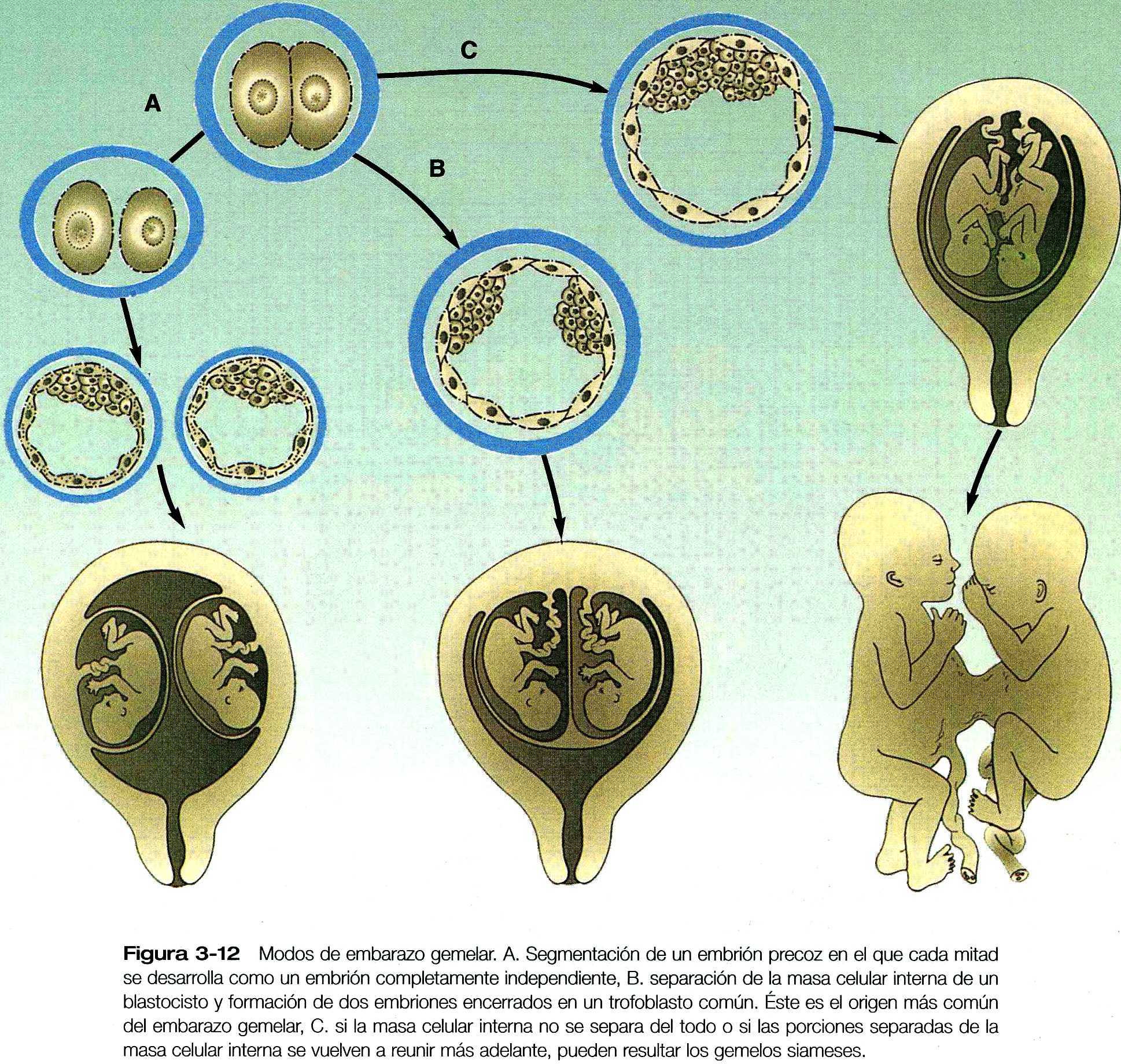
Algunos tipos de embarazos gemelares representan un experimento natural que demuestra la enorme capacidad reguladora del embrión humano en las fases tempranas del desarrollo. En los estados unidos aproximadamente un embarazo de cada 90 es gemelar, y uno de cada 8.000 es de trillizos. Del numero total de gemelos que nace, alrededor de dos tercios son fraternos, o dicigóticos, y un tercio son gemelos idénticos, o monocigoticos. Los dicigóticos son el producto de la fecundación de dos óvulos, y en la etiología de su formación esta implicado el control endocrino de ovulación. Por otra parte, los gemelos monocigóticos y algunos trillizos don el producto de un único óvulo fecundado. Surgen por la subdivisión y separación de un embrión único. Aunque en teoría los gemelos monocigóticos podrían surgir de la división de un embrión de dos células, suelen aceptarse que la mayoría proviene de la subdivisión celular de la masa interna de un blstocisto (figura 3_12). Como la mayoría de los gemelos monocigóticos son perfectamente normales es obvio que el embrión humano precoz puede subdividirse y que cada componente puede regularse para formar un embrión normal. Las inferencias sobre el origen y las relaciones de los nacimientos múltiples puede hacerse a partir de la organización de las membranas extraembrionarias en el momento del parte (capitulo 6).
Los cuádruples o los embarazos múltiples mas numerosos son muy poco frecuentes. Antes, podrían ser producto de ovulaciones múltiples y de la separación de blastocistos únicos. En la era actual de la tecnología reproductora, la mayoría de los nacimientos múltiples, en ocasiones incluso hasta séptuples, pueden atribuirse a los efectos secundarios de los medicamentos que la madre toma durante los tratamientos de la infertilidad.
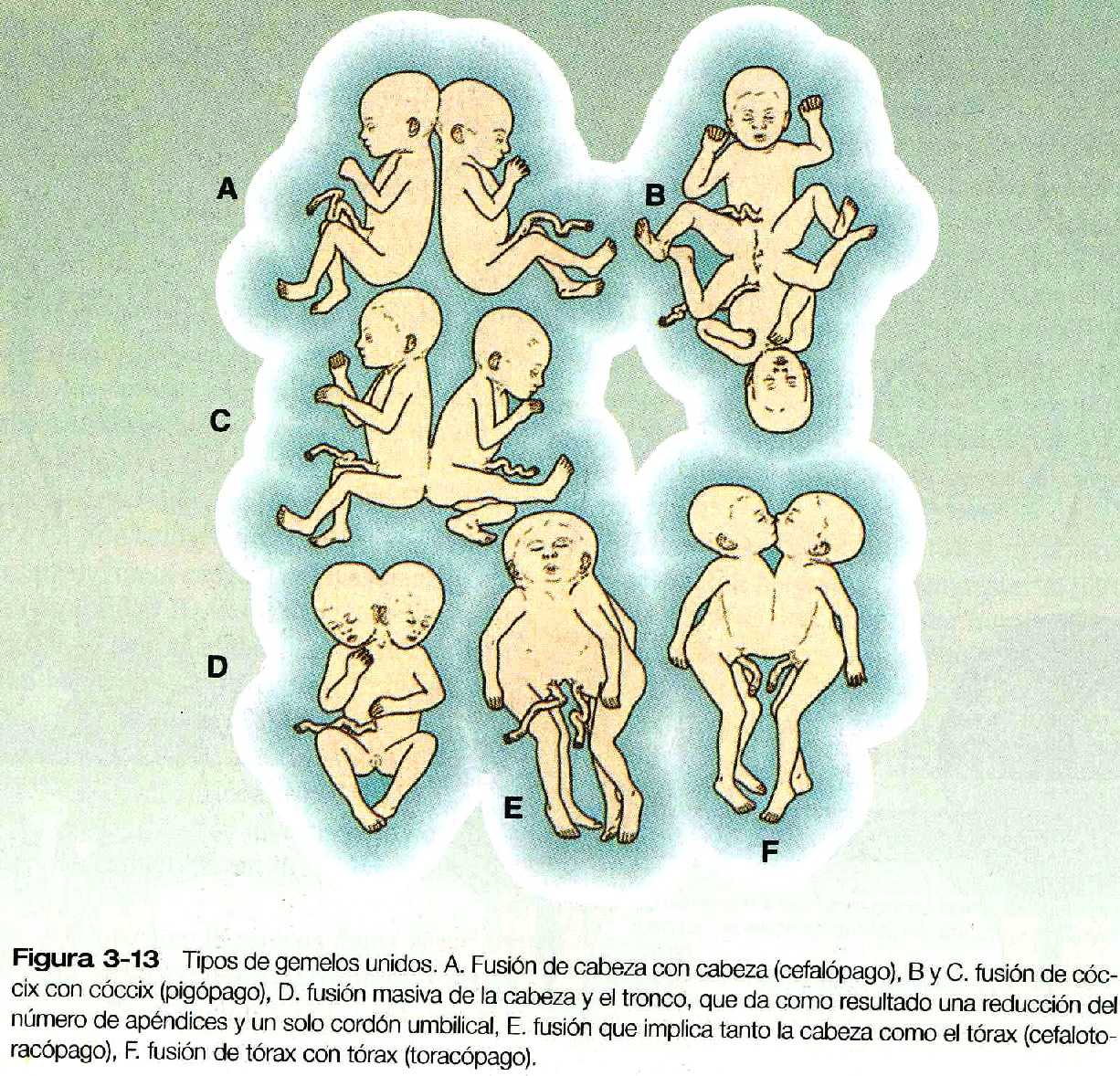
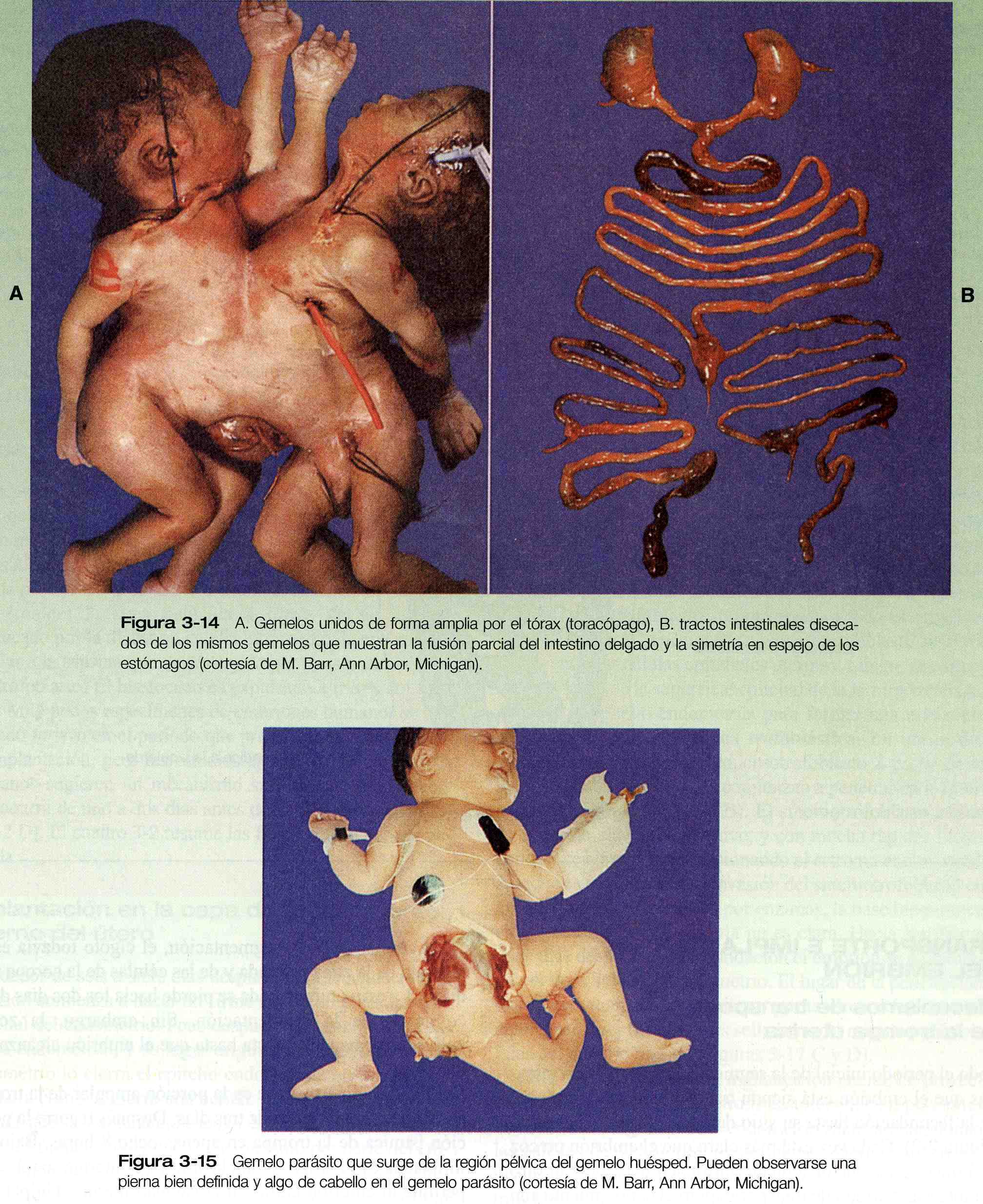
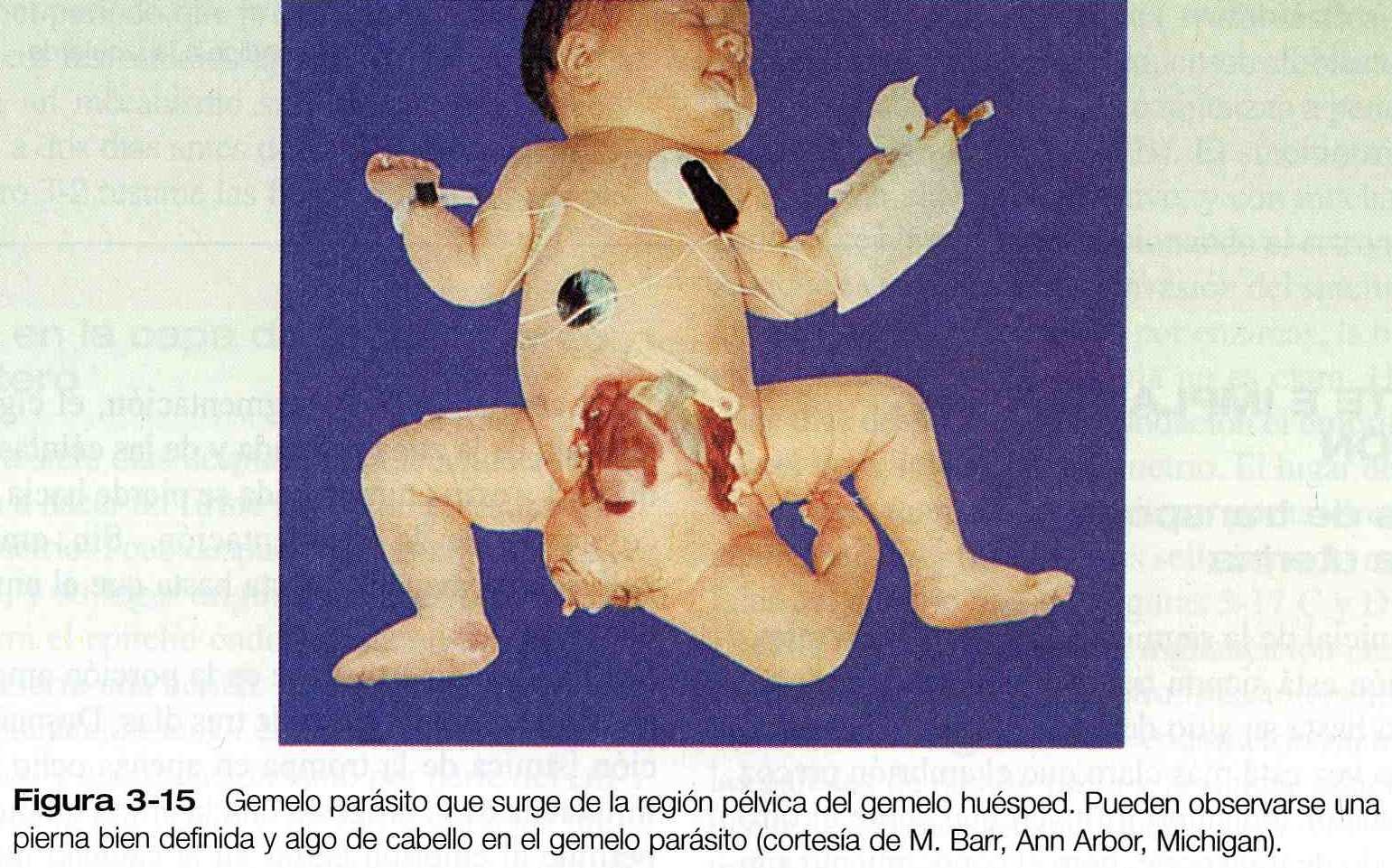
En ocasiones, la separación de porciones de la masa celular interna en un embrión es incompleta y, aunque los dos embriones se forman, están unidos por un puente de tejido de proporciones variables. Cuando esto sucede los gemelos se llaman gemelos unidos (son mas conocidos por el nombre coloquial de gemelos siameses). El grado de unión entre los gemelos varia desde una conexión relativamente delgada en el tórax o la espalda hasta fusiones masivas a lo largo de la mayor parte del eje corporal. En las figuras 3_13 y 3_14 se presentan ejemplos de la amplia variedad de gemelos unidos. Con la creciente mejoría de las técnicas quirúrgicas pueden separase los gemelos con grados de fusión más complejos. Una variedad mucho menos frecuente de gemelos unidos es el gemelo parásito, en el cual una porción mucho mas pequeña, pero notablemente completa, de un cuerpo protruye del cuerpo de un gemelo huésped por lo demás normal (figura 3_15). Los lugares de unión comunes de los gemelos parásitos son la región oral, el mediastino y la pelvis. El mecanismo de los gemelos unidos no se ha demostrado directamente de forma experimental, pero las posibles explicaciones teóricas son la fusión parcial secundaria de porciones originalmente separadas de la masa celular interna o la formación de dos estrías primitivas en un embrión único (capitulo 4).
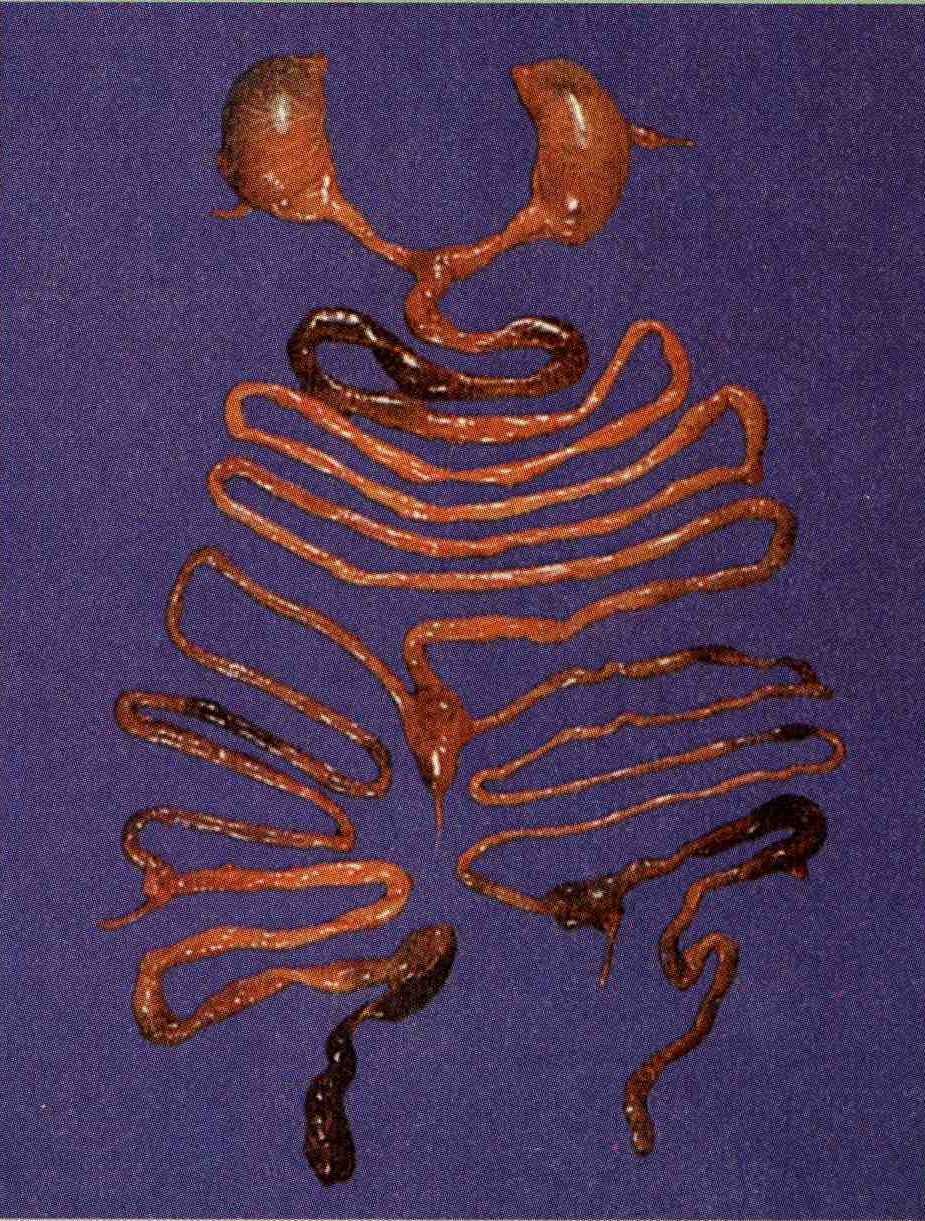
Un fenómeno que se encuentra a menudo en los gemelos unidos es una inversión en la simetría de los órganos de uno de los miembros de la pareja (figura 3_14B). Estas inversiones de la simetría son comunes en los órganos duplicados o en todo el embrión. Hace mas de un siglo, este fenómeno se registro en diversas situaciones biológicas y se incorporó en lo que ahora se denomina la regla de Bateson, según la cual cuando las estructuras duplicadas se unen durante las etapas criticas de desarrollo, una estructura es la imagen en espejo de la otra. A pesar de conocerse desde hace tanto tiempo este fenómeno, solo en época reciente se ha obtenido datos para comprender el mecanismo productor de la inversión de la simetría.
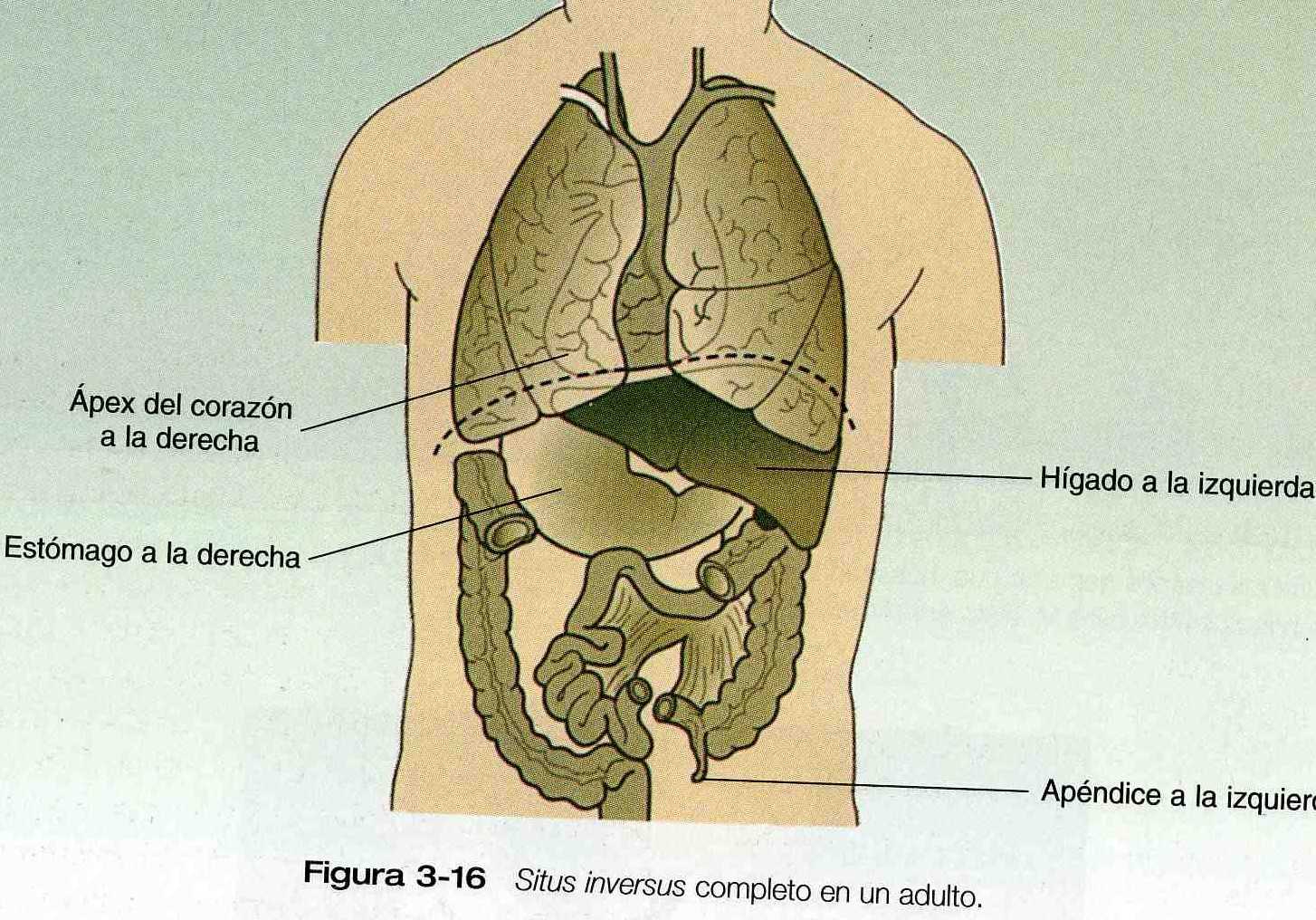
La inversión de la simetría llamada situs inversus, también se observa en alrededor de 1 de cada 10 mil individuos normales nacidos de embarazos únicos (figura 3_16). A esos individuos a menudo solo se les reconoce cuando lo diagnostica un clínico experto en una edad relativamente tardía. La base para la simetría normal del cuerpo (por ejemplo del estómago a la izquierda y el hígado a la derecha) ha comenzado a comprenderse tan solo hace poco tiempo, con el descubrimiento de una mutación recesiva que produce situs inversus de manera constante en los ratones, se ha establecido una base genética para la simetría normal del cuerpo. También esta armando el rompecabezas de la base molecular de la simetría normal con el descubrimiento de que ciertas moléculas de activación y claves para el desarrollo se expresen ya en el lado izquierdo y en el lado derecho de la región que se esta organizando durante la gastrulación (pagina 73).
TRANSPORTE E IMPLANTACIÓN DEL EMBRIÓN
Mecanismos de transporte de la trompa uterina
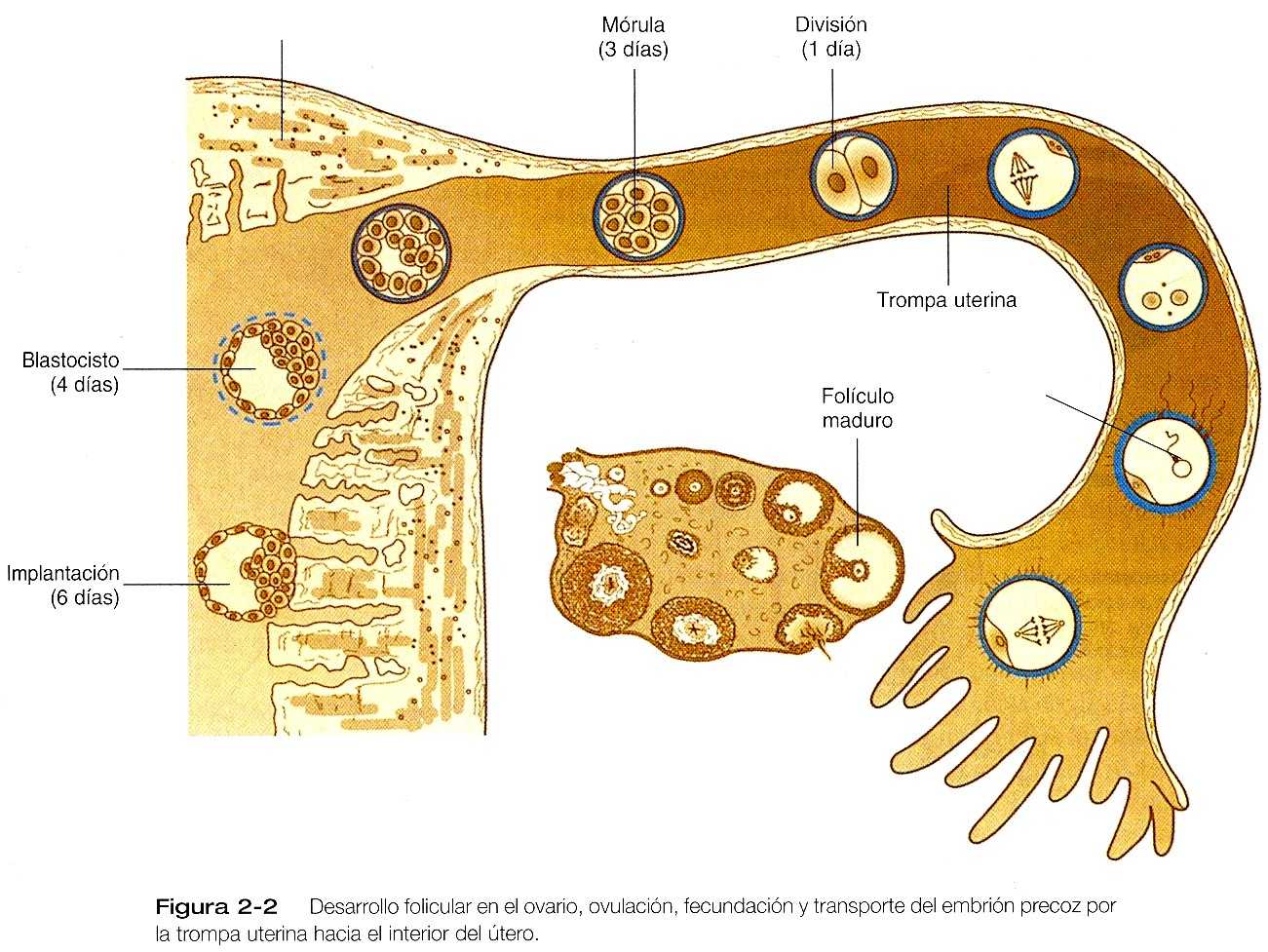
Todo el periodo inicial de la segmentación tiene lugar mientras que el embrión esta siendo transportado desde el lugar de la fecundación hasta su sitio de implantación en el útero (figura 2_2). Cada vez esta mas claro que el embrión precoz y el tracto reproductor femenino influye uno sobre el otro durante este periodo de transporte, pero el conocimiento aún es parcial.
Al comienzo de la segmentación, el cigoto todavía esta rodeado de la zona pelúcida y de las células de la corona radiada. La estructura radiada se pierde hacia los dos días del comienzo de la segmentación sin embrago, la zona pelúcida permanece intacta hasta que el embrión alcanza el útero.
El embrión permanece en la porción ampular de la trompa uterina durante cerca de tres días. Después recorre la porción Ístmica de la trompa en apenas ocho horas. Bajo la influencia de la progesterona la unión tubouterina se relaja y permite al embrión entrar en la cavidad uterina. Un par de días mas tarde (6 a 8 días después de la fecundación), el embrión se implanta en la porción media de la pared posterior del útero.
TABLA 3-2 etapas de la implantación humana
|
Edad (días) |
Maduración del blastocisto |
|
5 |
Perdida de la zona pelúcida del blastocisto |
|
5 |
Anclaje del blastocisto al epitelio uterino |
|
6? |
Penetración del epitelio |
|
7 ½ -9 |
Formación de la placa trofoblástica e invasión del estroma uterino por el blastocisto |
|
9-11 |
Formación de las lagunas junto con erosión de las arterias espirales del endometrio |
|
12-13 |
Formación de las vellosidades primarias |
|
13-15 |
Vellosidades placentarias secundarias, formación del saco vitelino secundario |
|
16-18 |
Ramificación y formación de las vellosidades de anclaje |
|
18-22 |
Formación de las vellosidades terciarias |
|
|
|
Zona pelúcida
Durante todo el periodo que va desde la ovulación hasta su entrada en la cavidad uterina, el óvulo y después el embrión están rodeados por la zona pelúcida. Después de que el embrión alcanza la cavidad uterina, se desprende de la zona pelúcida para preparar la implantación. El blastocisto “eclosiona” (rompe) la zona pelúcida a través de un agujero provocado por la digestión de ella, por medio de un encima similar a la tripsina que es segregada por unas pocas células del trofoblasto. El blastocisto es expulsado a través del agujero. Muy pocos especimenes de embriones humanos se han tomado in vivo en el periodo que precede inmediatamente a la implantación, pero los estudios in vitro de los embriones humanos sugieren un mecanismo similar, que es probable que ocurra de uno a dos días antes de la implantación (figura 3_2 D). El cuadro 3-2 resume las funciones de la zona pelúcida.
Implantación en la capa de revestimiento interno de útero
Alrededor de seis a siete días después de la fecundación, el embrión comienza a ser un firme anclaje al revestimiento epitelial del endometrio. Poco después, se sumerge en el estroma endometrial, y si lugar original de penetración en el endometrio lo cierra el epitelio endometrial, en forma muy similar a como se cierra una herida.
Para que la implantación tenga éxito se requiere un alto grado de preparación y coordinación tanto por parte del embrión como del endometrio (tabla 3-2). Las complejas preparación hormonales del endometrio que comenzaron al final del periodo menstrual anterior están todas dirigidas a proveer un ambiente celular y nutricional propicio para el embrión. La disolución de la zona pelúcida señala la disposición del embrión a comenzar la implantación.
La primera etapa de la implantación consiste en el anclaje del blastocisto expandido al epitelio endometrial. La superficies apicales de las células epiteliales del endometrio condicionadas por la actividad hormonal expresan diversas células de adhesión celular (subunidades de las integrinas) que permiten que la implantación tenga lugar en el estrecho lapso entre los 20 a 24 días del ciclo menstrual ideal.
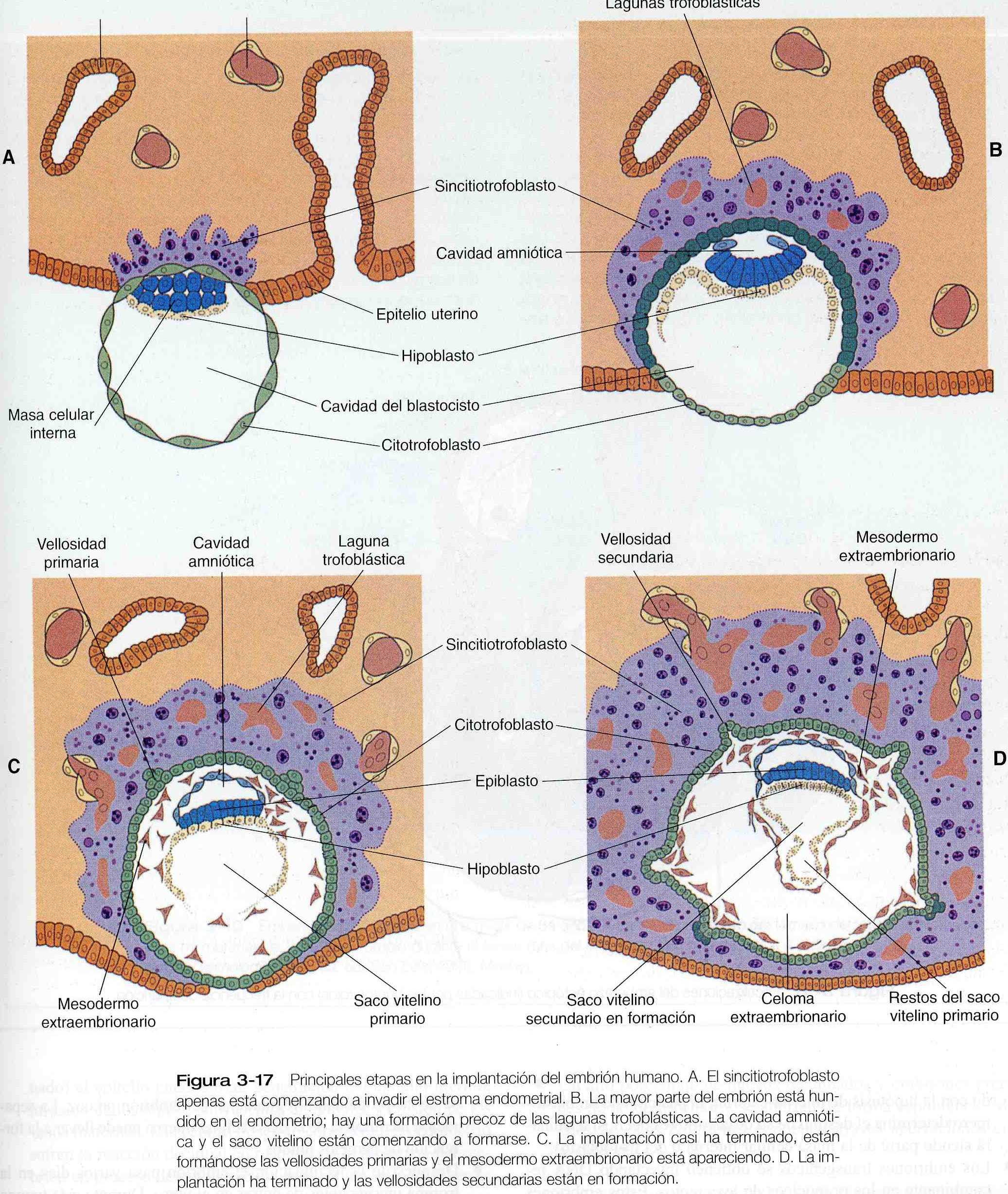
De la misma forma, las células trofoblasticas del blastocisto antes de la implantación también expresan integrinas en sus superficies. De acuerdo con una teoría reciente, el blastocisto se ancla al epitelio endometrial a través de la mediación del ligando que establecen puentes que conectan las integrinas de sus superficies. Estudios tanto in vitro como in vivo han demostrado que el anclaje del blastocisto se produce en el área situada encima de la masa celular interna (polo embrionario), lo cual sugiere que la superficie del trofoblasto no es homogéneo.
La siguiente etapa de la implantación es la penetración del epitelio uterino. En los primates, el trofoblasto celular pasa por otra etapa adicional de diferenciación inmediatamente antes de entra en contacto con el endometrio en el área que roda la masa celular interna las células derivadas del trofoblasto celular (citotrofoblasto) se funcionan para formar un sincitiotrofoblasto multinucleado. Aunque solo una pequeña área del sincitiotrofoblasto se introducen entre las células epiteliales uterinas. Luego, se extienden a lo largo de la superficie epitelial de la lamina basal que subyace al epitelio endometrial para formar una estructura algo aplanada llamada placa trofoblástica. En uno o dos días, las proyecciones del sincitiotrofoblasto a partir de la pequeña placa trofoblástica comienzan a penetra en la lámina basal (figuras 3_17 A y B). El sincitiotrofoblasto inicial es un tejido altamente invasivo y con mucha rapidez se expande y se abre camino erosionando el estroma endometrial.
Aunque es evidente que la invasión del sincitiotrofoblasto en el endometrio esta medida por encimas la base bioquímica en los seres humanos todavía no es clara. Hacia los diez a doce días después de la fecundación el embrión se ha hundido por completo en el endometrio. El lugar de la penetración inicial esta marcado al comienzo por un área desnuda o un tapón acelular mas tardes sellada por la migración de células de epitelio uterino (figuras 3_17 C y D).
A medida que prosigue la implantación inicial, las proyecciones invasivas del sincitiotrofoblasto envuelven porciones de los vasos del endometrio materno, erosionan las paredes de dichos vasos y la sangre materna comienza a llenar las lagunas aisladas que se han estado formando en el trofoblasto (figuras 3_17 C y D). Los procesos trofoblásticos entran en los vasos sanguíneos e incluso comparten complejos de unión con las células endoteliales.
En el momento en que se forman las lagunas llenas de sangre, el trofoblasto cambia de carácter y ya no es tan invasor como durante los primeros días de la implantación.
Mientras que el embrión se enquista en el endometrio y algunas células del citotrofoblasto se funcionan en el sincitiotrofoblasto, las células estromales parecidas a fibroblastos del endometrio que esta algo edematizado, se hinchan, por la acumulación de glicógeno y gotitas de lípidos (figura 6_6).
Estas células, llamadas células deciduales, son muy adherentes y forman una matriz celular masiva que primero rodea al embrión en implantación y mas adelante ocupa la mayor parte del endometrio. Al mismo tiempo de la reacción desigual se denomina esta transformación los leucocitos que han infiltrado el estroma endometrial durante la fase progestacional tardía del ciclo endometrial segregan interleucina 2, que impide que la madre reconozca al embrión como un cuerpo extraño durante las primeras etapas de la implantación. Un embrión es antihigiénicamente diferente de la madre y en consecuencia debería ser rechazado por una reacción inmunológica medida por células similar al tipo de rechazo de un trasplante incompatible de riñón o de corazón. Parece que una función primaria de la reacción decidual es ofrecer un lugar privilegiado desde el punto de vista inmunológico para proteger al embrión en desarrollo para que no sea rechazado, pero aun no se comprende el mecanismo por el cual ello ocurre a pesar de años de intensa investigación.
No es raro que un blastocisto fracase en su anclaje al endometrio y por tanto no se realice la implantación. El fracaso de la implantación es un problema especialmente agudo de los procedimientos de fecundación in vitro y transferencia embrionaria, por lo cual la taza del éxito de la implantación de los embriones transferidos sigue siendo de alrededor del 20% (correlación clínica 3_3).
Muerte embrionaria y aborto espontáneo
Un alto porcentaje de óvulos fecundados (mas del 50%) no se desarrolla hasta la madurez y se aborta de forma espontánea.
La mayor parte de los abortos espontáneos se presenta durante las primeras tres semanas del embarazo.
A causa del pequeño tamaño del embrión en ese momento, a menudo la madre no reconoce los abortos espontáneos como tales y puede comparar el aborto y la hemorragia que lo acompaña con un periodo menstrual un poco retrasado y desusadamente intenso.
El examen de los embriones precoces obtenido después de un aborto espontáneo o del útero extirpado en cirugía durante las primeras semanas del embarazo a mostrado que muchos de los embriones abortados tienen severas anomalías las aberraciones cromosómicas representan la categoría más común de las anomalías en los abortos (cerca del 50 % de lo casos). Cuando se analizan conjuntamente con los estados patológicos mal formativos pueden considerarse los abortos espontáneos como un mecanismo natural para reducir la incidencia de bebes con malformaciones muy graves.
CORRELACIÓN CLÍNICA 3-3 Embarazo ectópico
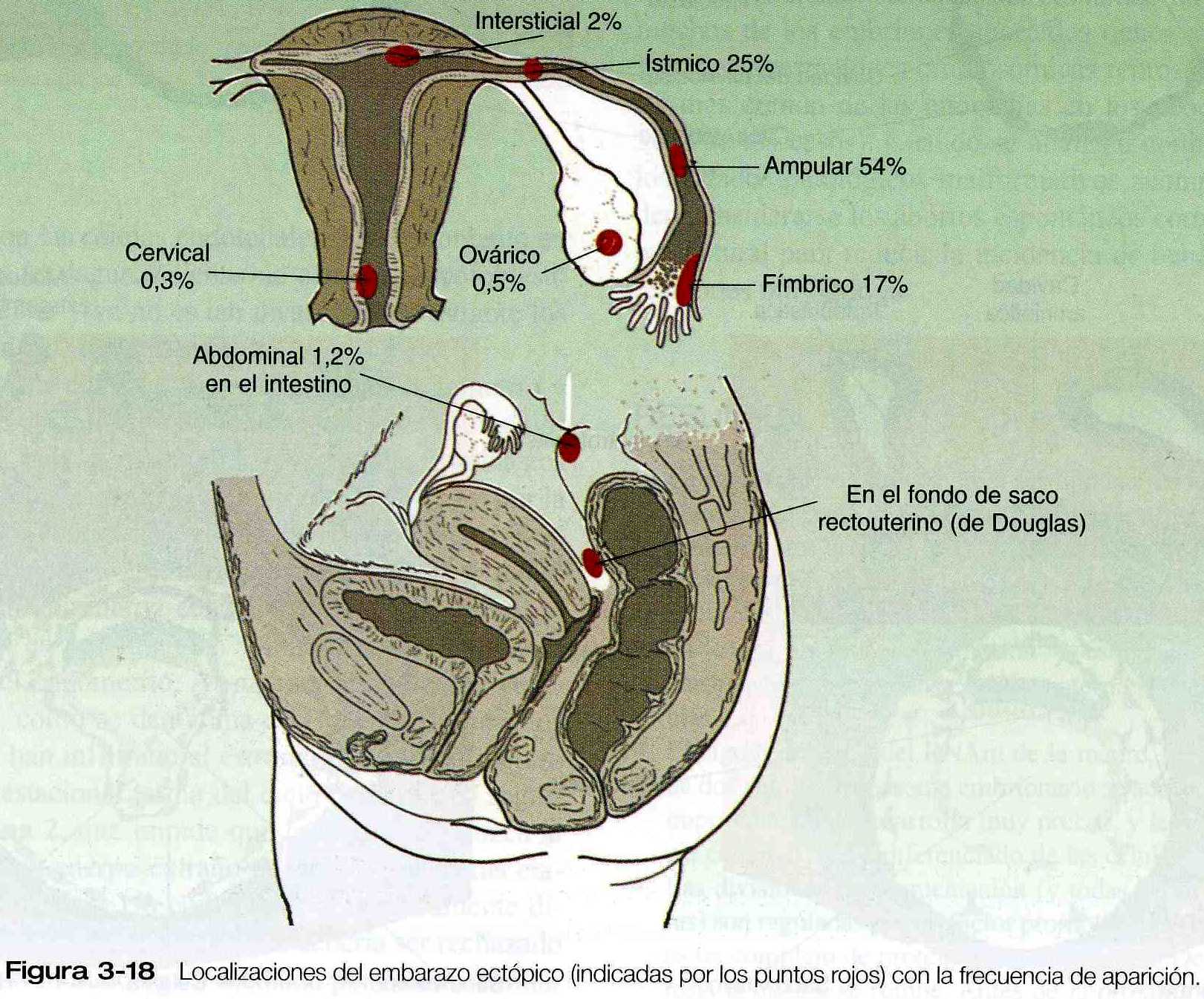
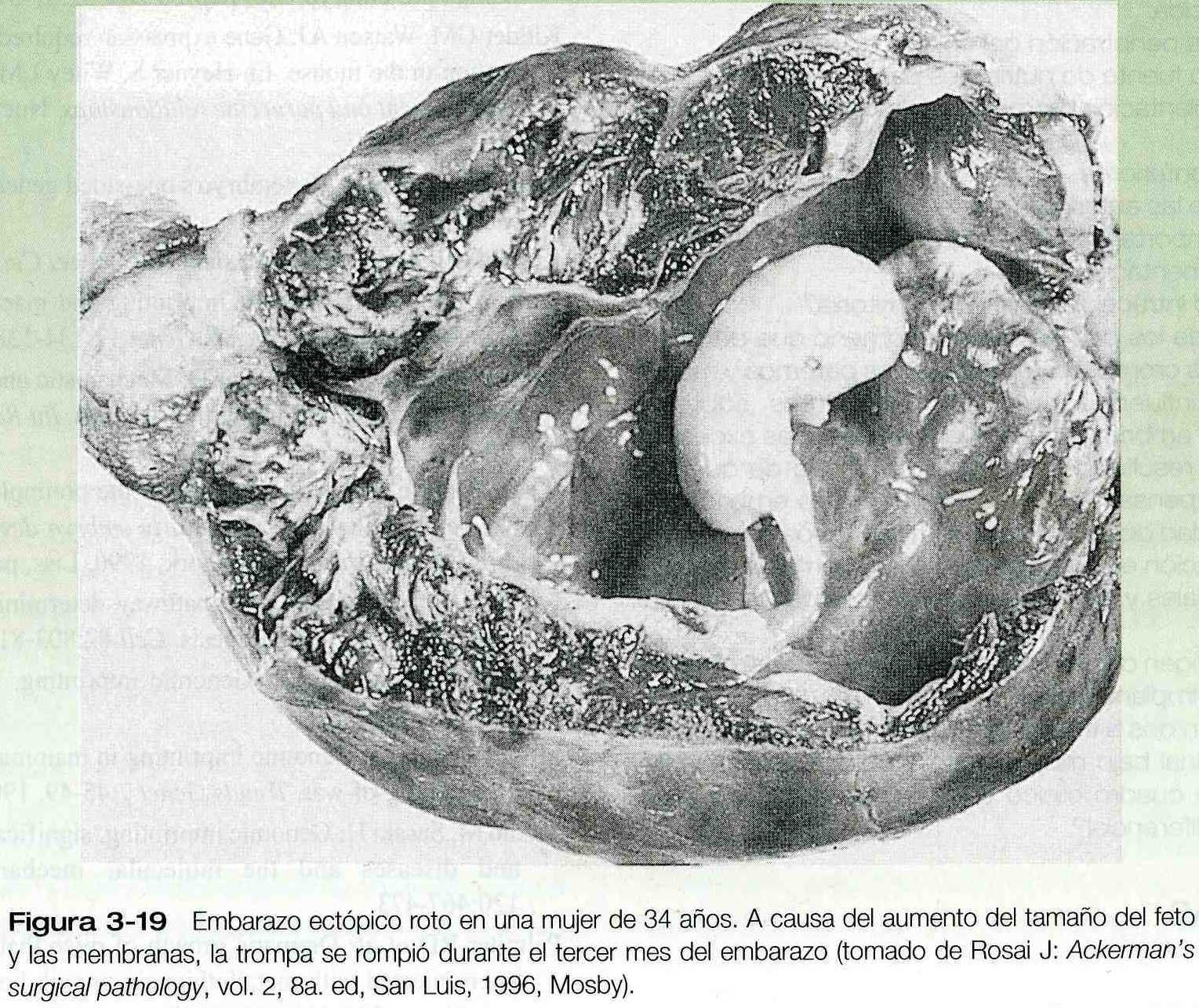
El blastocisto suele implantarse en la pared posterior de la cavidad uterina pero en u8n bajo porcentaje de los casos (0.25% a 1.0%) la implantación se produce en un lugar anómalo. Este fenómeno se conoce como embarazo ectópico. Los embarazos tubáricos son con mucho el tipo mas común de embarazo ectópico. Aunque en casi todos los embarazos tubáricos se encuentran en la porción ampular de la trompa pueden localizarse en cualquier lugar desde el extremo distal (fimbria) hasta la unión tubouterina (figura 3_18). Los embarazos tubáricos (figura 3_19) se encuentran con más frecuencia en mujeres que han tenido endometriosis (una enfermedad caracterizada por la presencia de tejido endometrial en localizaciones anómalas), cirugías anteriores o enfermedad pélvica inflamatoria. La cicatrización posterior a la inflamación o en ocasiones las anomalías anatómicas tienen como resultado bolsas ciegas entre los pliegues de la mucosa de la trompa; aquí puede quedar atrapado un blastocisto. Por lo común la mujer presenta los signos normales del embarazo incipiente, pero cerca de las nueve a once semanas el embrión implantado y sus derivados trofoblásticos asociados han crecido hasta el punto en que el adelgazamiento de la pared de la trompa ocasiona dolor abdominal agudo. Si no se trata, el embarazo tubárico suele terminar con la rotura de la trompa y hemorragia, a menudo de gravedad suficiente para poner en peligro la vida de la madre.
Muy rara vez el embrión se implanta en el ovario (embarazo ovárico) o en la cavidad abdominal (embarazo abdominal).
Estos casos pueden ser el resultado de la fecundación de un óvulo antes de que entre en la trompa o del reflujo de un óvulo fecundado de la trompa o muy raras veces, de la penetración de un embarazo tubárico a través de la pared de la trompa. El lugar de implantación mas común para el embarazo abdominal es en fondo del saco recto uterino (fondo del saco de Douglas) esta localizado detrás del útero. La implantación en la pared intestinal o en el mesenterio es muy peligrosa debido a la posibilidad de hemorragia profusa a medida que crece el embrión. En algunos casos, un embrión se ha desarrollado hasta el término de la localización abdominal.
Si no se extrae el embrión puede calcificarse formando un lithopedion (litopedio).
En el interior del útero el embrión puede implantarse cerca del cerviz. Aunque el desarrollo embrional normal es probable, la placenta suele formar una cubierta parcial sobre el canal cervical. Esta situación llamada placenta previa puede producir hemorragia hacia el final del embarazo y si no se trata, puede llegar a causar la muerte del feto, la madre o de ambos a causa del desprendimiento prematuro de la placenta con la hemorragia que lo acompaña. La implantación directa en el interior del canal cervical es muy rara.
PREGUNTAS DE REPASO. Q03.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}