4
FORMACIÓN DE LAS CAPAS GERMINALES Y SUS DERIVADOS INICIALES
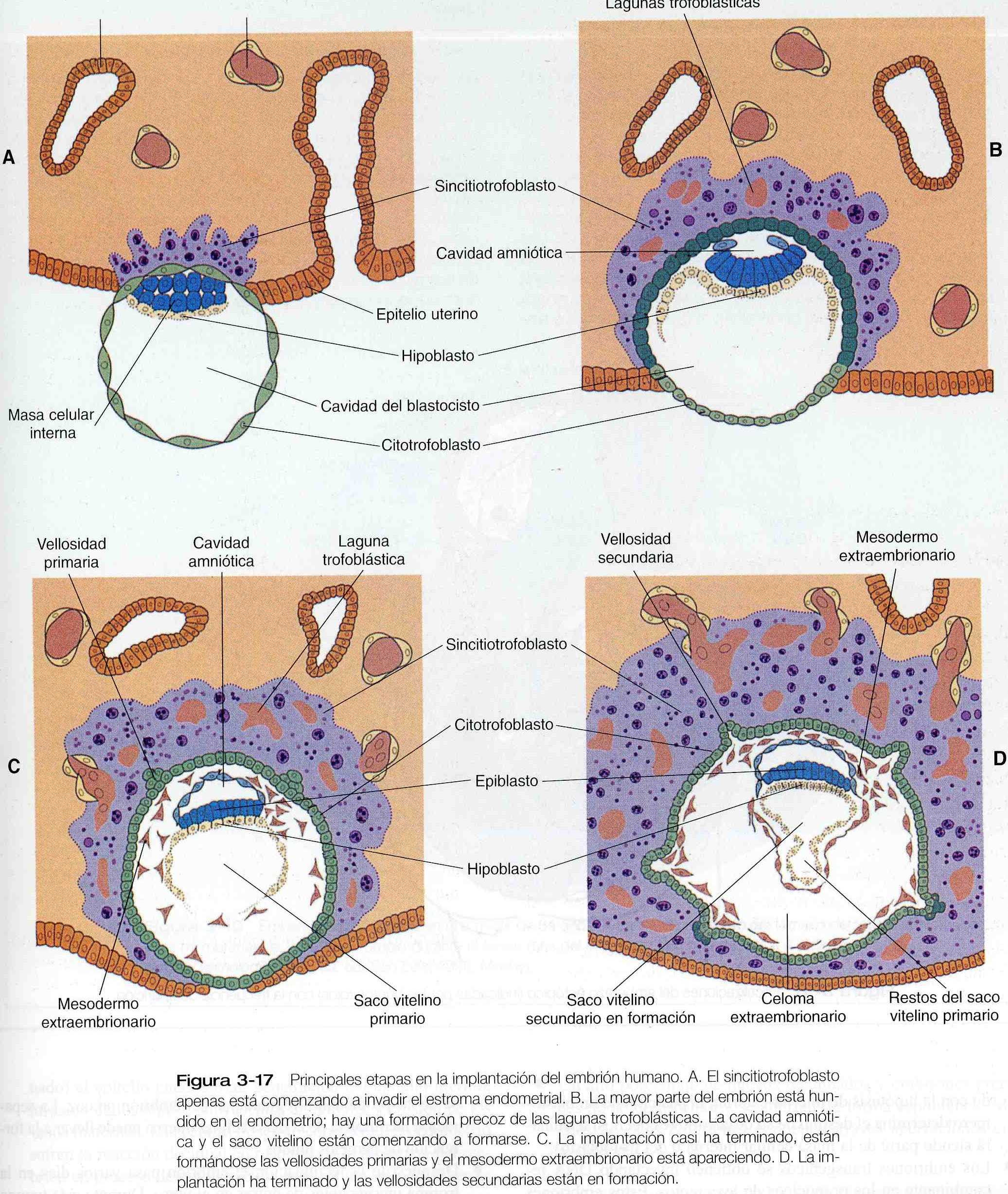
Conforme se va implantando en la pared uterina, el embrión sufre profundos cambios en su organización. Hasta el momento de la implantación, el blastocisto esta compuesto por la masa celular interna, a partir de la cual surge el cuerpo del embrión propiamente dicho, y el trofoblasto externo, que representa el futuro tejido de conexión entre la madre y el embrión. Ambos componentes del blastocisto sirven como precursores de otros tejidos que aparecen en etapas posteriores del desarrollo. En el capitulo 3 se examinó como el citotrofoblasto da origen a una capa sincitial externa, el sincitiotrofoblasto, poco antes de anclarse en el tejido uterino (figura 3-17). No mucho tiempo después, la masa celular interna comienza también a dar origen a otros derivados titulares. La subdivisión de la masa celular interna tiene como resultado un cuerpo embrionario que contiene las tres hojas o capas germinales primarias: el ectodermo (capa externa), el mesodermo (capa intermedia), y el endodermo (capa interna). El proceso por el cual se forman las capas germinales mediante desplazamientos celulares se denomina gastrulación.
Después de que se ha formado las capas germinales, el mantenimiento progresivo del desarrollo embrionario depende de una serie de señales llamadas inducciones embrionarias que se intercambian entre las capas germinales u otros precursores de tejidos. En una interacción inductiva, un tejido (el inductor) actúa sobre otro (inducido) de manera que el curso del desarrollo de este último es diferente de lo que habría sido en ausencia del inductor.
Las modificaciones en el embrión que pueden observarse al microscopio durante este periodo son reflejos tangibles de cambios profundos en la expresión genética y en las propiedades celulares de los embriones en la fase de implantación.
En los últimos años la abundante información sobre el desarrollo inicial de las moscas Drosophila a nivel molecular se ha utilizado en los estudios relacionados con el desarrollo inicial de anfibios y mamíferos. Algunos de los principales hallazgos se presentan en este capitulo y en el siguiente.
ETAPA DEL EMBRIÓN BILAMINAR
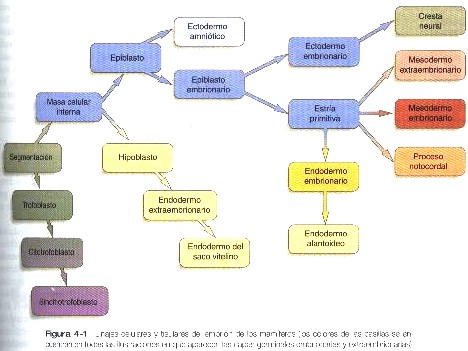
Inmediatamente antes de que el embrión se implante en el endometrio al comienzo de la segunda semana empiezan a producirse cambios importantes en la masa celular interna así como el trofoblasto. Conforme las células de la masa celular interna se reorganizan en una configuración epitelial, aparece una delgada capa de células ventral a la masa celular interna (figura 3-17). La capa superior principal de células se conoce como epiblasto, y la inferior se denomina hipoblasto, o endodermo primitivo (figura 4-1).
Todavía no se sabe cómo se forma el hipoblasto en los embriones humanos, pero la formación proveniente de la embriología comparada sugiere que las células de esta etapa surgen por delaminación (separación o esfacelo) a partir de la masa celular interna. El hipoblasto se considera como un endodermo extraembrionario, y en el último término da lugar al revestimiento endodérmico del saco vitelino (figura 3-17). Después de que el hipoblasto se ha convertido en una capa bien definida y el epiblasto ha adquirido una configuración epitelial, la antigua masa epitelial interna se transforma en un disco bilaminar, con el epiblasto en la superficie dorsal y el hipoblasto en la superficie ventral.
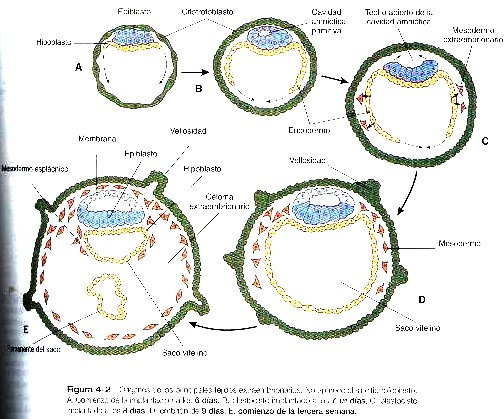
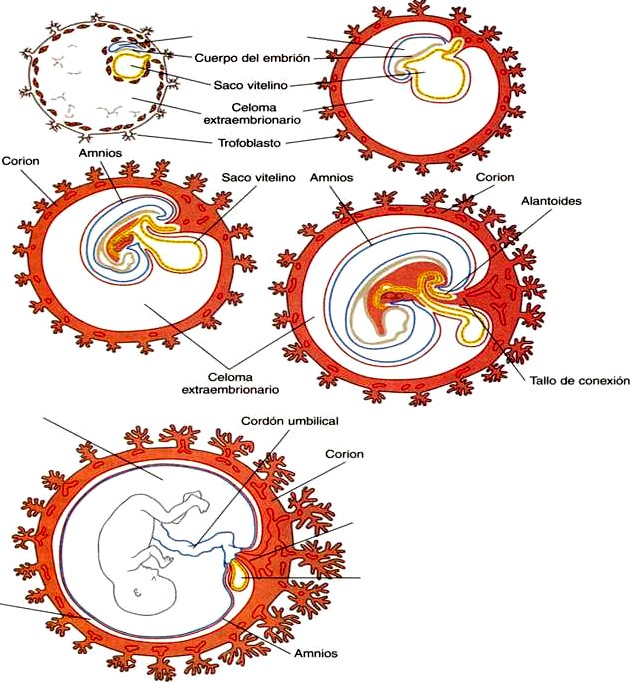
El epiblasto contiene las células que constituirán el embrión propiamente dicho, pero los tejidos extraembrionarios también surgen a partir de esa capa. La siguiente capa en aparecer después del hipoblasto es el amnios, una capa de ectodermo extraembrionario que encerrara a todo el embrión dentro de una cavidad llena de liquido denominada cavidad amniótica (capitulo 6). A causa de la escasez de especimenes, las primeras etapas de formación del amnios y la cavidad amniótica todavía no se comprenden del todo. Los estudios de embriones de primates sugieren que primero surge una cavidad amniótica primordial mediante la cavitación (formación de un espacio interno) en el interior del epiblasto preepitelial; esta cavidad esta cubierta por células derivadas de la masa celular interna (figura 4-2). De acuerdo con algunos investigadores, el techo del amnios se abre entonces exponiendo la cavidad amniótica primordial al citotrofoblasto situado por encima de ella. Poco después (unos ocho días después de la fecundación), el epitelio amniótico original vuelve a formar un techo sólido y continuo sobre la cavidad amniótica.
Mientras que el embrión aún esta hundiéndose en el endometrio (unos nueve días después de la fecundación), algunas células del hipoblasto comienzan a extenderse y a tapizar toda la superficie interna del citotrofoblasto, con una capa continua de endodermo extraembrionario llamado endodermo parietal (figura 4-2). Cuando la extensión endodérmica ha terminado, ha adquirido forma una vesícula llamada saco vitelino primario (figura 3-17C).En Este punto (unos diez días después de la fecundación), el complejo embrionario constituye un disco embrionario bilaminar que se localiza entre el saco vitelino primario en su superficie ventral y cavidad amniótica en su superficie dorsal. Poco después de que se forma, el saco vitelino primario se constriñe formando un saco vitelino secundario dejando atrás un remanente del saco vitelino primario (figuras 3-17D y 4-2E).
Unos doce días después de la fecundación, comienza a aparecer otro tejido extraembrionario, el mesodermo extraembrionario (figura 4-2). Las primeras células mesodérmicas extraembrionarias parecen surgir a partir de una transformación de las células endodérmicas parietales. Estas células más adelante se unen a las células mesodérmicas extraembrionarias que se han originado en la estría primitiva. El mesodermo extraembrionario se convierte en el tejido que sostiene al epitelio del amnios y al saco vitelino así como las vellosidades coriales, que surgen de los tejidos trofoblásticos (capitulo 6). El sostén que ofrece el mesodermo extraembrionario no solo es mecánico sino también trófico, pues el mesodermo sirve como sustrato morfológico a través del cual los vasos sanguíneos abastecen de oxigeno y nutrientes a los diversos epitelios.
GASTRULACIÓN Y LAS TRES CAPAS GERMINALES DEL EMBRIÓN
Al final de la segunda semana el embrión consta de dos capas planas de células: el epiblasto y el hipoblasto. Cuando comienza la tercera semana de gestación, el embrión entra en período de gastrulación, durante la cual se establecen claramente las tres capas germinales embrionarias. La morfología de la gastrulación humana sigue el patrón que se ha observado en las aves. A causa de la gran cantidad de vitelo en los huevos de las aves, el embrión forma las capas germinales primarias como tres discos planos superpuestos que descansan sobre el vitelo en forma muy parecida a una pila de monedas. Sólo más tarde las capas germinales se pliegan para formar un cuerpo cilíndrico. Aunque el huevo de los mamíferos en esencia esta desprovisto de vitelo, el conservacionismo morfológico del desarrollo inicial todavía obliga al embrión humano a seguir un patrón de gastrulación similar al que se observa en los reptiles y las aves. Debido a la escasez de material, la morfología de la gastrulación en embriones humanos no se conoce en detalle. No obstante, la extrapolación a partir de la gastrulación de las aves y los mamíferos puede ofrecer un modelo aceptablemente válido de la gastrulación humana.
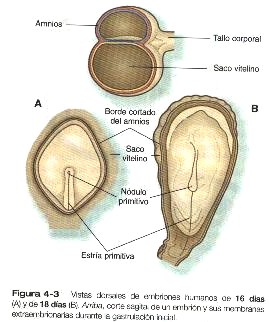
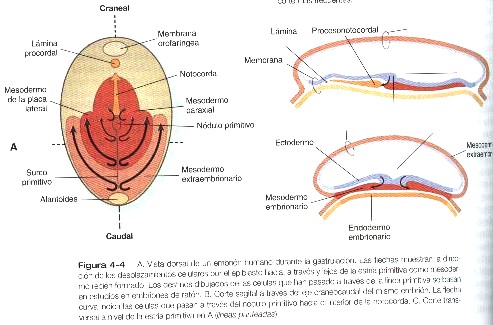
Todas las capas germinales embrionarias derivan del epiblasto (figura 4-1). La primera evidencia de la gastrulación es la formación de la estría primitiva, que aparece primero como un engrosamiento y después como una línea corta en la superficie dorsal del epiblasto (figura 4-3). El esbozo inicial de la estría primitiva es una condensación causada por la convergencia de células epiblásticas hacia esa área. Con la aparición de la estría primitiva, ya pueden identificarse los ejes antero posterior (creaneocaudal) e izquierdo derecho del embrión (figuras 4-4 y 4-5).
El conocimiento
de la dinámica celular asociada a la estría primitiva facilita la comprensión de
la morfología detallada de esta área. A medida que las células del epiblasto
alcanzan la estría primitiva, cambian de forma y pasan a través de ella en su
trayecto hacia la formación de nuevas capas de células por debajo del epiblasto
(ventrales a éste), como se ve en la figura 4-4.
El desplazamiento de células por la estría primitiva tiene como resultado la
formación de un surco (el surco primario o primitivo) a lo largo de la
línea media de ésta. En el extremo de la estría primitiva se encuentra un
pequeño pero bien definido cúmulo de células llamada nódulo primitivo o
nódulo de Hensen*. Esta estructura reviste enorme importancia en el
desarrollo porque es el área a través de la cual las células epiblásticas en
migración se dirigen para formar una estructura redondeada y cordonal de células
mesenquimatosas denominada notocorda (la notocorda y sus funciones en el
embrión precoz se discuten mas adelante.
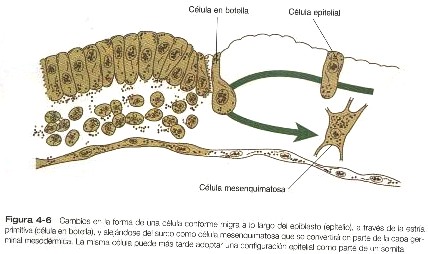
El movimiento de las células para pasar a través de la estría primitiva se
acompaña de cambios profundos en su estructura y su organización (figura
4-6). Mientras que están en el epiblasto, las
células tienen las propiedades de las células epiteliales típicas, con
superficies apicales y basales bien definidas y se asocian como la lámina basal
que subyace al epiblasto. Conforme entran en la estría primitiva, estas células
se alargan, pierden su lámina basal y adoptan una morfología característica que
ha conducido a que se las llame células en botella. Cuando se liberan de
la capa epiblástica del surco primitivo, las células en botella adoptan la
morfología y las características de las células mesenquimatosas, que pueden
migrar como células individuales si se les suministra un sustrato extracelular
apropiado (figura 4-6). En esta transformación
esta incluida la pérdida de moléculas especificas de adhesión celular (Pág. 72
Cuadro 4-1) a medida que las
células pasan de tener una configuración epitelial a una mesenquimatosa.
En las aves, las primeras células en abandonar la línea primitiva entran en la capa del hipoblasto, de donde desplazan algunas de las células endodérmicas extraembrionarias para formar el endodermo embrionario definitivo. Este proceso no se ha confirmado en el embrión humano, pero la morfología de la gastrulación es comparable con la que se observa en las aves.
La característica más sobresaliente de la gastrulación humana es la formación del mesodermo. Algunas células que migran a través de la estría primitiva forman el mesodermo extraembrionario. Una gran parte del mesodermo extraembrionario forma el tallo corporal, o de conexión que comunica la parte caudal del embrión con los tejidos extraembrionarios que la rodean (figura 4-7 y 6-1). El tallo corporal mas adelante se convierte en el cordón umbilical.
Después de que la estría primitiva esta bien establecida, la mayoría de las células que pasan por ella se extienden entre el epiblasto y el hipoblasto para formar el mesodermo embrionario (figura 4-4). Los estudios con trazadores han mostrado que las células que abandonan el nódulo y la estría primitiva en diferentes regiones craneocaudales están destinadas a formar tipos específicos y distantes de mesodermo.
Por ejemplo. Las células que pasan por el nódulo primitivo se convierten ya sea en notocorda o en mesodermo paraxial, mientras que aquellas que pasan por la estría primitiva forman mesodermo paraxial (Pág. 92), el mesodermo de la placa lateral (Pág. 96) o el mesodermo extraembrionario, de acuerdo con el nivel craneocaudal en el cual abandonan la estría primitiva (figura 4-4). Las transformaciones de la morfología y el comportamiento de las células que pasan por la estría primitiva se asocian con profundos cambios no sólo en las propiedades de adhesividad y en su organización interna sino también en la forma en que se relacionan con el ambiente extracelular.
Desde el comienzo mismo de la gastrulación, las células del epiblasto producen ácido hialurónico, que entra en el espacio entre el epiblasto y el hipoblasto. El ácido hialurónico, un polímero que consta de subunidades repetidas de ácido D-glucurónico y N-acetilglucosamina, suelen asociarse con la migración celular en los sistemas en desarrollo. La molécula tiene una enorme capacidad de fijar agua (hasta mil veces su propio volumen), y su función es evitar que las células del mesénquima se agreguen y adhieran durante las migraciones celulares. Aunque después de que dejan la estría primitiva las células mesenquimatosas del mesodermo embrionario se encuentran en un sustrato rico en ácido hialurónico, el ácido por si solo no basta para mantener su migración desde la estría primitiva. En todos los embriones de vertebrados que se han estudiado hasta la fecha, se han encontrado que la migración de las células mesodérmicas más allá de la estría primitiva o la estructura equivalente depende de la presencia de fibronectina asociada con la lámina basal por debajo del epiblasto. El mesodermo embrionario en último término se extiende hacia los lados en la forma de una delgada hoja de células mesenquimatosas entre el epiblasto y el hipoblasto (figura 4-4). En el momento en que el mesodermo ha formado una discreta hoja en el embrión humano, la capa germinal superior (restos del antiguo epiblasto) se denomina ectodermo, y la capa germinal inferior, que ha desplazado al hipoblasto original, se llama endodermo. Esta terminología se usará en el resto del texto.
REGRESIÓN DE LA ESTRÍA PRIMITIVA
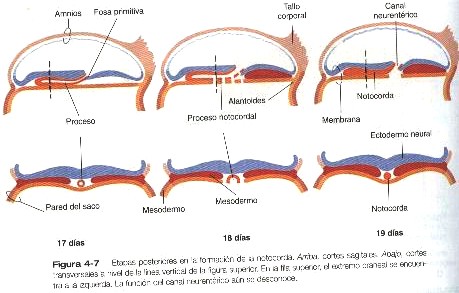
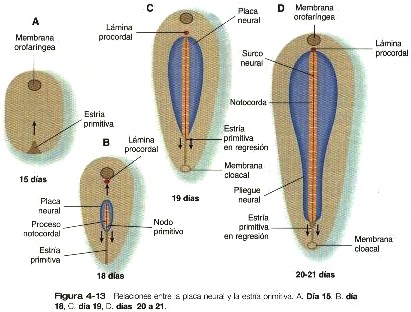
Desde su aparición inicial en el extremo caudal del embrión, la estría primitiva se extiende en dirección craneal hasta unos 18 días después de la fecundación (figura 4-13). Posteriormente regresa en dirección caudal (4-13), extendiendo la notocorda a su paso. Sus vestigios permanecen hasta la cuarta semana; tiempo durante el cual la formación del mesodermo continúa por medio de las células que migran desde el epiblasto a través del surco primitivo.
La estría primitiva normalmente desaparece sin dejar huella, pero en casos raros aparecen grandes tumores llamados teratomas en la región sacrococcigea (figura 1-2A). Los teratomas suelen contener mezclas extrañas de muchos tipos diferentes de tejidos, como cartílago, músculo, grasa, pelo, y tejido glandular. Por ello, se piensa que los teratomas sacrococcigeos tienen su origen en restos de la estría primitiva (que pueden formar todas las capas germinales). También se encuentran teratomas en las gónadas y el mediastino. Se piensa que estos tumores se originan a partir de las células germinales.
NOTOCORDA Y LÁMINA PROCORDAL
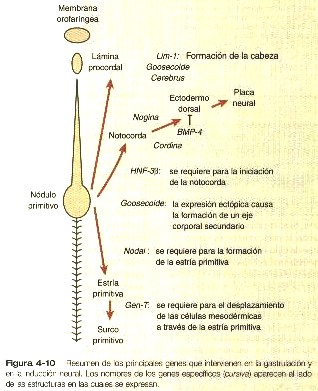
La notocorda –estructura que es la base que da el nombre chordata al phylum al cual pertenecen todos los vertebrados- es un cordón celular que corre a lo largo del eje longitudinal del embrión inmediatamente ventral al sistema nervioso central. Aunque desde los puntos de vista filogenético y ontogenético la notocorda sirve como soporte longitudinal primario del cuerpo, también desempeña un papel crucial como centro primario de una serie de procesos (inducciones) que transforman células embrionarias no especializadas en tejidos y órganos definitivos. En especial, las señales inductivas procedentes de la notocorda 1) estimulan la conversión del ectodermo superficial suprayacente en tejido neural, 2) especifican la identidad de ciertas células (placa del suelo), dentro del sistema nervioso inicial, 3) transforman ciertas células mesodérmicas de los somitas en cuerpos vertebrales y 4) favorecen las primeras fases del desarrollo del páncreas dorsal.
La notocorda surge de la proyección de una población de células epliblasticas a través del nódulo primitivo formando un agregado celular cilíndrico denominado proceso notocordal (figura 4-4). En los mamíferos, poco después de la proyección, las células del proceso notocordal se extienden y se fusionan con el endodermo embrionario (figura 4-7B), de lo cual resulta la formación de un canal neurentérico transitorio que conecta la cavidad amniótica con el saco vitelino. Posteriormente, las células de la notocorda se separan del techo endotérmico del saco vitelino y forman la notocorda definitiva, un cordón sólido de células localizando en la estría media entre el ectodermo y el endodermo embrionario (figura 4-7C).
Hacia el extremo craneal de la notocorda se encuentra una pequeña región en la cual el ectodermo y el endodermo embrionario se adosan sin que haya mesodermo entre ellos. Esta estructura, que se denomina membrana orofaríngea (figura 4-4), marca el lugar de la futura cavidad oral. Entre la punta craneal del proceso notocordal y la membrana orofaríngea hay una pequeña aglomeración de células mesodérmicas en estrecha aposición con el endodermo, que se llama la lámina procordal (figura 4-4). Esta lámina procordal emite señales moleculares que son fundamentales para estimular la formación del cerebro anterior (Pág.69).
INTRODUCCIÓN AL SISTEMA NERVIOSO
Inducción Neural.
La relación inductiva entre la notocorda (cordamesodermo) y el ectodermo suprayacente en la génesis del sistema nervioso se reconoció a comienzos del siglo XX. Aunque los primeros experimentos se hicieron en anfibios, otros experimentos similares realizados en vertebrados superiores han demostrado que los elementos esenciales de la inducción neural (o primaria) son los mismos en todos los vertebrados.
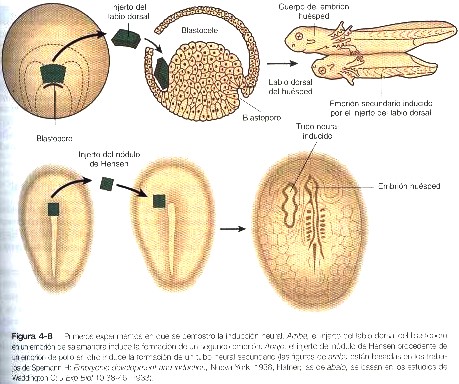
Los experimentos de deleciones y transplantes en anfibios dieron la pauta para la comprensión actual de la inducción neural (véanse los capítulos 5 y 10 para mas detalles sobre la formación del sistema nervioso). En ausencia del cordamesodermo que se desplaza desde el labio dorsal del blastoporo (el equivalente del nódulo primitivo en los anfibios), el sistema nervioso, que esta representado al comienzo por una lámina engrosada de células ectodérmicas transformadas situadas a lo largo de la estría media dorsal del embrión, no se forma a partir del ectodermo dorsal. Por otra parte, si el labio dorsal del blastoporo se injerta por debajo del ectodermo de la región ventral de otro huésped, se forma un segundo sistema nervioso y un segundo eje corporal en el área del injerto (figura 4-8). El labio dorsal se ha denominado organizador por su capacidad para estimular la formación de un eje corporal secundario. Las investigaciones posteriores han demostrado que las interacciones que tienen lugar en la región del labio dorsal de los anfibios son mucho más complejas que una simple inducción entre el cordomesodermo y el ectodermo.
Los experimentos como el descrito también se han realizado en embriones de aves y de mamíferos (figura 4-8); resulta claro que el nódulo primitivo y el proceso notocordal de las aves y los mamíferos son homólogos en su función al labio dorsal y al cordomesodermo de los anfibios. Esto significa que en los vertebrados superiores el nódulo primitivo y el proceso notocordal actúan como inductor neural, y el ectodermo suprayacente es el tejido que responde (inducido). Esta relación fundamental se estableció hace más de cincuenta años. Desde ese momento, los embriólogos han dedicado enormes esfuerzos de investigación a tratar de identificar la naturaleza de la señal inductiva que pasa del cordomesodermo al ectodermo.
Los primeros intentos dirigidos a descubrir la naturaleza del estimulo inductor se caracterizaron por un enorme optimismo. Desde el decenio de 1950, diversos laboratorios habían propuesto que moléculas tan diferentes como proteínas y esteroides eran el estimulo inductor. Poco después se descubrió una gran variedad aún mayor de estímulos, como iones inorgánicos o tejidos muertos, podían suscitar la inducción neural. Con tal plétora de posibles inductores, la atención se volvió hacia las propiedades del tejido que responde (el ectodermo dorsal) y las formas en que podría reaccionar, a través de una vía final común, al estímulo inductor.
La búsqueda de las moléculas inductoras neurales y de su modo de acción ha sido ardua y frustrante, pues en el camino se han encontrado muchos callejones sin salida y giros equivocados.
Varios laboratorios encontraron que el ectodermo aislado podía responder in Vitro implico separar el tejido diana del tejido inductor mediante un filtro con poros que permitían el paso de moléculas pero no de células. Esta técnica se ha usado en el análisis de varios de los sistemas inductores de los mamíferos.
Diversas manipulaciones experimentales han mostrado con claridad que la inducción neural no es un proceso simple de todo o nada. En vez de ello, existe una considerable especificidad regional (por ejemplo, ciertos inductores artificiales estimulan la formación de estructuras neurales más anteriores, en tanto que otros estimulan la formación de estructuras mas posteriores). En los embriones de anfibios el cordomesodermo anterior tiene posibilidades inductoras diferentes del cordomesodermo posterior.
Investigaciones recientes han identificado moléculas específicas que provocan la inducción neural. En las aves y en los mamíferos, dos moléculas de señales, la nogina y la cordina, producidas por la notocorda, son los agentes inductores. Primero se pensó que estas dos moléculas estimulaban directamente a las células no diferenciadas del ectodermo dorsal para que formaran tejido neural, pero investigaciones posteriores en anfibios han demostrado que estos inductores actúan bloqueando la acción de un inhibidor, la proteína morfogenética ósea-4 (BMP-4), en el ectodermo dorsal (figura 4-10). En ausencia de la actividad de la BMP-4, el ectodermo dorsal forma por defecto tejido neural. Este grupo de interacciones moleculares obliga a las células ectodérmicas que están sobre la notocorda a convertirse en tejido neural, pero éste es sólo el primer paso en la formación del sistema nervioso. Un segundo paso importante es la regionalización del sistema nervioso central.
La regionalización se refiere a la subdivisión del sistema nervioso central en amplias regiones craneocaudales.
Como ya se indicó, los primeros experimentos mostraron que ciertos inductores causaban la formación de estructuras neurales más craneales y otros, más caudales. Hoy se sabe que a través de las acciones de la nogina y de la cordina solas, se forman más estructuras neurales craneales.
No obstante, si se añade a la mezcla el factor de crecimiento de fibroblastos, se forman principalmente estructuras neurales caudales (cerebro posterior) In vivo, un factor aun sin identificar segregado por el mesodermo recién formado ejerce un efecto caudalizante sobre el ectodermo neural. Una de las funciones recientemente descubiertas de la lámina procordal es especificar la formación de la región del cerebro anterior. La lámina procordal por sí sola no puede inducir al ectodermo dorsal a que se convierta en un tejido neural, pero una vez que la inducción neural realizada por la nogina y la cordina se ha producido, la lámina precordal desempeña un papel importante en la regionalización del cerebro anterior del sistema nervioso central.
Inducción Mesodérmica
Cuando se describió por primera vez, se consideró que la inducción neural era el primer proceso inductivo que tiene lugar en el embrión y por ellos se la llamó inducción primaria.
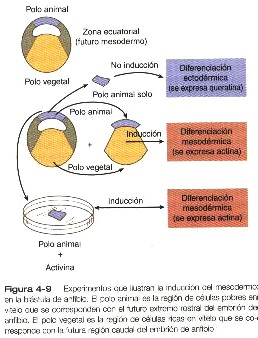
La experimentación posterior llevada a cabo sobre todo en anfibios ha demostrado que otras inducciones importantes suceden antes que la inducción neural. La mejor conocida de ellas es la inducción del mesodermo en la blástula del anfibio. El mesodermo normalmente surge a partir de un anillo de células localizando alrededor de la región ecuatorial de la blástula (figura 4-9). Si se aísla el ectodermo localizado en el techo del blastocele, sigue siendo ectodermo general y produce niveles normales de proteína de queratina, que son moléculas específicas de ectodermo. Si el mismo trozo del ectodermo se pone en aposición con el endodermo, se diferencia en mesodermo, según lo indica su producción de a actina, una molécula característica del músculo. Recientemente, se ha logrado comprender mucho mejor la naturaleza de la inducción del mesodermo por ciertas proteínas especificas –activina, nogina y Vgl (tabla 5-2)- que son efectores de la inducción mesodérmica.
Aunque casi toda la investigación actual sobre la inducción en embriones jóvenes se esta realizando en anfibios, muchos de los principios allí adquiridos pueden transferirse a los embriones de los vertebrados superiores. Si en un ave se aísla el epiblasto inicial y se cultiva, no se forman la notocorda ni el mesodermo axial. Si un hipoblasto intacto o incluso células del hipoblasto disociadas y luego reagregadas se yuxtaponen al epiblasto cultivado, se forman estructuras axiales. Otro experimento a demostrado que si se añade medio de cultivo tisular expuesto a un inductor mesodérmico (células XTC) a un epiblasto aislado de ave, el resultado es la inducción del epiblasto a formar estructuras mesodérmicas axiales. La activina sola puede intuir estructuras axiales en el epiblasto de las aves. Existen razones fundadas para creer que estos resultados también se aplican a los embriones de los mamíferos (cuadros 4-1 y 4-2).
Formación inicial de la placa neural
La primera respuesta morfológica evidente del embrión a la inducción neural es la transformación del ectodermo dorsal que reviste al proceso notocordal en una zona alargada de células de epitelio engrosado llamado placa neural (figura 4-13).
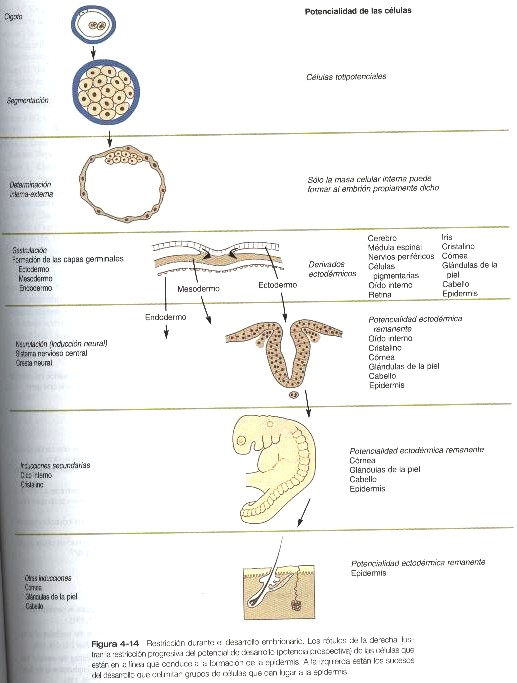
Con la formación de la placa neural, la capa germinal del ectodermo se subdivide en dos linajes de desarrollo: neural y no neural. Este ejemplo ilustra varios conceptos fundamentales del desarrollo: restricción, determinación y diferenciación. El cigoto y los blastómeros que resultan del primer par de divisiones de segmentación son totipotenciales (es decir, tienen capacidad de formar cualquier célula del organismo). A medida que el desarrollo avanza, se toman ciertas decisiones que restringen las opciones del desarrollo de las células (figuras 4-14). Por ejemplo, en una etapa inicial de la segmentación, algunas células quedan confinadas a la línea trofoblástica extraembrionaria y ya no pueden participar en la formación del embrión como tal. En el punto de decisión en el cual las células quedan confinadas a convertirse en trofoblasto, se ha producido un suceso de restricción.
Cuando una célula o un grupo de células ha pasado por su último punto de decisión (por ejemplo, la transición del citotrofoblasto a sincitiotrofoblasto), su destino queda sellado y se dice que están determinadas. Se entiende que estos términos, acuñados en los primeros días de la embriología experimental, reflejan actualmente las limitaciones en la expresión genética conforme los linajes celulares siguen el curso normal de su desarrollo. Los pocos casos en los cuales el curso de las células o tejidos se desvía mucho de su desarrollo normal –fenómeno llamado metaplasia- son de considerable interés para los patólogos y para los estudiosos del control de la expresión genética.
La restricción y la determinación significan la limitación progresiva de las capacidades del desarrollo en el embrión.
La diferenciación describe la expresión morfológica o funcional real de la porción del genoma que permanece disponible para una célula o un grupo de células definidas. La diferenciación suele connotar el curso de la especialización fenotípica de las células. Un ejemplo de diferenciación se produce en la espermatogénesis, cuando las espermatogonias, unas células de apariencia relativamente ordinaria, se transforman en los muy especializados espermatozoides.
MOLÉCULAS DE ADHESIÓN CELULAR.
A comienzos del siglo XX, los investigadores determinaron que las suspensiones de células de un mismo tipo tienen una fuerte tendencia a agregarse. Si se mezclan juntos diferentes tipos de células embrionarias, lo característico es que se ordenen de acuerdo con el tipo del cual proceden. Sus patrones de ordenamiento incluso dan claves sobre sus propiedades y su comportamiento en el organismo maduro. Por ejemplo, si se mezclan dos células de ectodermo y mesodermo embrionarios, se reúnen en un agregado que tiene una capa superficial de células ectodérmicas que rodean a un agregado central de células mesodérmicas.
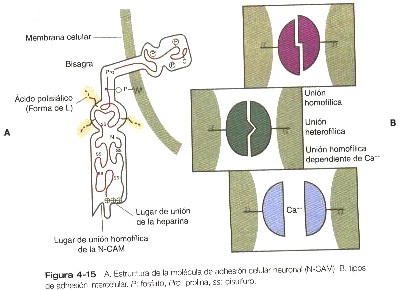
Las investigaciones actuales han comenzado a encontrar una base molecular para muchos de los fenómenos de agregación y ordenamiento celular descritos por los primeros embriólogos. Se han caracterizado varias familias de moléculas de adhesión celular (CAM). Las CAM se han estudiado ampliamente en relación con los primeros sucesos del desarrollo. En los embriones de aves previos a la gastrulación, las células tanto del epiblasto como del hipoblasto contienen dos CAM (N-CAM y L-CAM) en sus superficies. Se produce un cambio importante cuando las células del epiblasto migran por la estría primitiva; ellas pierden la expresión de ambas CAM durante la fase migratoria y mientras que están comenzando a formar un mesodermo organizado. Mas adelante, ciertos derivados de las células mesodérmicas vuelven a expresar las CAM. En general, cuando un tipo celular epitelial se transforma en una célula mesenquimatosa, se pierden sus CAM de superficie.
La expresión de las CAM es un excelente indicador de la inducción primaria en el embrión joven. Antes de la inducción, el epiblasto expresa tanto la N-CAM como la L-CAM (también llamada E-cadherina). Después de la inducción primaria del sistema nervios, las células de la placa neural conservan la N-CAM pero pierden la expresión de la L-CAM. Por el contrario, en el ectodermo no neural, se pierde la expresión N-CAM pero se conserva la L-CAM.
Algunas de las CAM requieren de Ca para funcionar, en tanto que otras son independientes del Ca. La N-CAM, una CAM independiente del Ca, se une directamente con otras moléculas de N-CAM de las células vecinas del mismo tipo (figura 4-15). La N-CAM es inusual porque tiene una alta concentración de grupos de ácido siálico de carga negativa en el componente de carbohidrato de la molécula, y las formas embrionarias de N-CAM tienen tres veces más ácido siálico que la forma adulta de la molécula.
RESUMEN
PREGUNTAS DE REPASO
1. El principal inductor de la inducción neural primaria es:
2. ¿Cuál de los siguientes tejidos surge a partir de las células que pasan por la estría primitiva?
3. ¿Las células de qué capa germinal no están presentes en la membrana orofaríngea?
4. La lámina procordal desempeña un papel importante en la regionalización de:
5. ¿La braquiuria, una deficiencia de los tejidos caudales del cuerpo, es causada por una mutación de que gen?
6. ¿Cuál capa del embrión bilaminar (de dos capas) da lugar a todo el tejido embrionario propiamente dicho?
7. ¿Qué importancia tiene el nódulo primitivo en el desarrollo embrionario?
8. ¿La migración de las células mesodérmicas a partir de la estría primitiva es facilitada por la presencia de que moléculas de la matriz extracelular?
9. ¿Qué moléculas pueden dar lugar a la inducción mesodérmica en un embrión joven?
10. ¿En qué etapa de la historia vital de muchas células se pierden las moléculas de adhesión celular?
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}