PLACENTA y MEMBRANAS EXTRAEMBRIONARIAS
Situaciones relacionadas con el líquido amniótico
Formación de las vellosidades coriónicas
Relaciones macroscópicas de los tejidos coriónicos y deciduales
Formación y estructura de la placenta madura
Estructura de la placenta madura
Estructura de la vellosidad coriónica madura
Transferencia placentaria
Correlación CLÍNICA 6-2
Transferencia placentaria anómala
TRANSFERENCIA CELULAR E INCOMPATIBILIDAD Rh.
Síntesis y secreción de las hormonas placentarias
Inmunología placentaria
Explicaciones: Primera. Segunda y tercera.
LA PLACENTA DESPUÉS DEL NACIMIENTO
LA PLACENTA y LAS MEMBRANAS EN LOS EMBARAZOS MÚLTIPLES
CORRELACIÓN CLÍNICA 6-3
BIOPSIA DE LA VELLOSIDAD CORIÓNICA
PREGUNTAS DE REPASO
Uno de los rasgos más característicos y sobresalientes del desarrollo embrionario humano es la íntima relación entre el embrión y la madre. El óvulo fecundado de los mamíferos no tiene vítelo. Para sobrevivir y crecer durante la vida intrauterina, el embrión ha de mantener una relación en esencia parasitaria con el cuerpo de la madre para adquirir el oxígeno y los nutrientes y eliminar los desechos. También tiene que evitar que el sistema inmunológico de su receptora lo rechace como un cuerpo extraño. La placenta y las membranas extraembrionarias que rodean al embrión y sirven como interfase entre el embrión y la madre satisfacen estos exigentes requisitos.
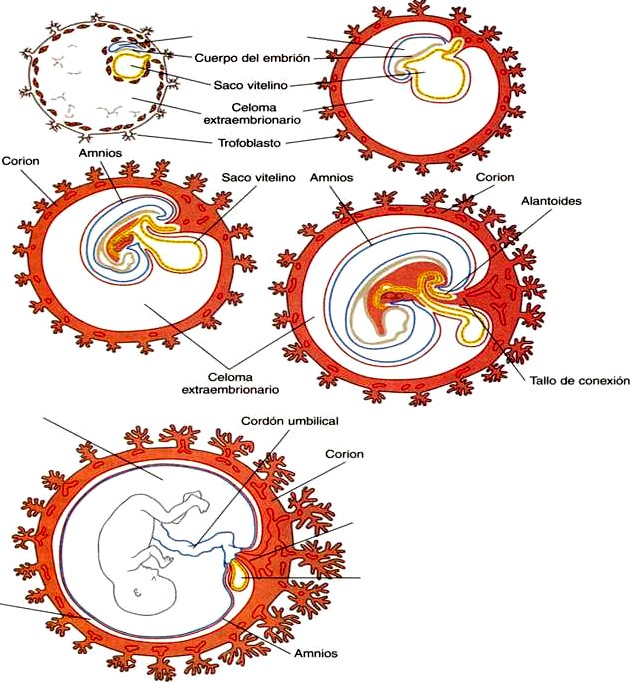
Los tejidos que componen la interfase fetomaterna (placenta y corion) son derivados del trofoblasto que se separa de la masa celular interna y rodea a los precursores celulares del embrión aun mientras el embrión está en segmentación y desciende por la trompa uterina hacia su lugar de implantación en la pared uterina (figura 3-17). Otros tejidos extraembrionarios se derivan de la masa celular interna. Éstos incluyen el amnios (un derivado ectodérmico), que forma una cápsula protectora llena de líquido alrededor del embrión; el saco vitelino (derivado del endodermo), que en los embriones de los mamíferos ya no cumple una función nutritiva primaria, la alantoides (derivado del endodermo) que se asocia con la eliminación de los desechos del embrión; y el mesodermo extraembrionario, que forma el cordón umbilical, el tejido conectivo que sustenta a las membranas extraembrionarias, y los vasos sanguíneos que las irrigan.
TEJIDO EXTRAEMBRIONARIO
El origen de la cavidad amniótica dentro del ectodermo de la masa celular interna en el embrión durante la fase de implantación se describió en el capítulo 4 (figuras 3-17 y 4-2). Conformen el embrión experimenta el plegamiento lateral y la incurvación cefalocaudal, la membrana amniótica que rodea el cuerpo del embrión como un globo lleno de líquido (figura 6-1), lo que permite que el embrión esté suspendido en un medio líquido durante todo el embarazo. El líquido amniótico sirve para amortiguar las presiones mecánicas a las que pudiese estar sometido el feto; además, el crecimiento, permite los movimientos fetales normales y protege al feto de las adherencias.
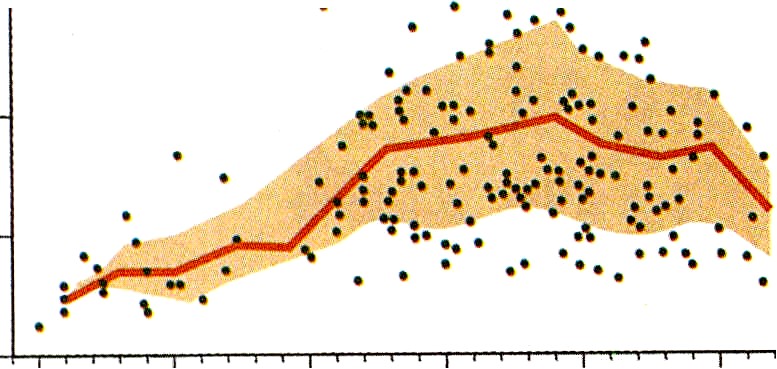
La delgada membrana amniótica consta de una única capa de células del ectodermo extraembrionario tapizadas por una capa no vascularizada de mesodermo extraembrionario. Al mismo ritmo que el crecimiento fetal, la cavidad amniótica se expande de manera continua hasta que su contenido líquido alcanza un volumen de casi 1.000 mL hacia las semanas 33 a 34 del embarazo (figura 6-2).
En muchos aspectos, el líquido amniótico puede considerarse como un trasudado diluido del plasma materno, pero el origen y la dinámica del intercambio del líquido amniótico son complejos y no han sido aclarados del todo. Parece haber dos fases en la producción del líquido amniótico. La primera fase comprende las 20 primeras semanas de embarazo, durante las cuales la composición del líquido amniótico es bastante similar a la de los líquidos fetales. Durante este período, la piel del feto no está queratinizada, y hay pruebas de que los líquidos y electrólitos pueden difundirse libremente a través de la piel del embrión de la piel. Además, la misma membrana amniótica segrega líquido y los componentes del suero materno pasan a través de la membrana amniótica.
Conforme avanza el embarazo (en especial después de la semana 20, cuando la epidermis fetal comienza a queratinizarse), se producen cambios en el origen del líquido amniótico. No hay datos concordantes en cuanto a los orígenes (y las contribuciones relativas) del líquido amniótico durante la segunda mitad del embarazo. En su formación, hay contribuciones crecientes a lo largo del embarazo procedentes de la orina fetal, filtración a partir de los vasos sanguíneos matemos situados cerca del corion leve (que está en estrecha aposición con la membrana amniótica en esta etapa), y posiblemente filtración a partir de los vasos fetales del cordón umbilical y la placa coriónica. En el tercer trimestre del embarazo el líquido amniótico se recambia por completo cada tres horas y, al final del embarazo, la tasa de intercambio de líquido puede aproximarse a los 500 rnL/h. Aunque gran parte del líquido amniótico se intercambia a través de la membrana amniótica, la deglución fetal es un mecanismo importante hacia el final del embarazo, en el cual el feto deglute unos 20 rnL/h de líquido. El líquido amniótico deglutido pasa al torrente circulatorio fetal después de ser absorbido por la pared del intestino. El agua ingerida puede salir de la circulación fetal a través de la placenta.
En la Correlación clínica 6-1 se examinan las situaciones relacionadas con la cantidad del líquido amniótico o las concentraciones de sustancias en el mismo.
Saco vitelino
El saco vitelino, que está tapizado por endodermo extraembrionario, se forma ventral al embrión bilaminar en contraposición al amnios que aparece en una localización dorsal al disco embrionario (figura 4-2). A diferencia de las aves y los reptiles, el saco vitelino de los mamíferos es pequeño y está desprovisto de yema. Aunque sus funciones originales como fuente de nutrición son escasas, sigue siendo vital para el embrión a causa de otras funciones que han llegado a desarrollarse en él.
Cuando aparece por primera vez, el saco vitelino tiene la forma de una hemiesfera que está limitada en la región ecuatorial por la pared dorsal del intestino primitivo (figura 6-1). Conforme el embrión crece y sufre el plegamiento lateral y la incurvación a lo largo del eje craneocaudal, la conexión entre el saco vitelino y el intestino en desarrollo disminuye y adquiere la forma de un tallo cilíndrico que se estrecha progresivamente y que en su extremo distal está unido a un saco vi te lino esférico. En las semanas siguientes el tallo vite lino se hace más largo y delgado y se sitúa en el interior del cordón umbilical. El saco vitelino se desplaza hacia la placa coriónica de la placenta (figura 6-3).
El endodermo del saco vitelino está revestido en la cara externa por un mesodermo extraembrionario bien vascularizado. Las células de cada una de estas capas contribuyen con algunos componentes al embrión. Durante la tercera semana, las células germinales primordiales, que han alcanzado el mesodermo extraembrionario cerca de la base de la alantoides, pueden reconocerse en el revestimiento del saco vitelino (figura 1-1). Pronto estas células migran hacia la pared del intestino y al mesenterio dorsal, en dirección hacia las gónadas, donde se diferenciarán en oogonias o espermatogonias.
Mientras tanto, otros grupos de células del mesodermo extraembrionario en la pared del saco vitelino forman los islotes sanguíneos (figura 5-24), y muchas de las células se diferencian en células sanguíneas primarias. La hematopoyesis extraembrionaria continúa en el saco vitelino hasta más o menos la sexta semana, cuando la actividad formadora de sangre se sitúa en localizaciones intraembrionarias, en especial el hígado.
A medida que se forma el tubo digestivo, la zona de anclaje del saco vitelino se hace cada vez más pequeña, hasta que hacia las seis semanas pierde del todo el contacto con el intestino. En un porcentaje pequeño de los adultos, los vestigios del saco vitelino persisten en la forma de un cordón fibroso o un divertículo en el intestino conocido como el divertículo de Meckel (figura 14-11 A). El saco vitelino puede persistir durante casi todo el embarazo, pero no se sabe si tiene una función específica en el período fetal. Las porciones proximales de los vasos sanguíneos del saco vitelino (el arco circulatorio vitelino) persisten y forman los vasos que irrigan la región del intestino medio.
La alantoides surge como una evaginación ventral del endodermo del intestino posterior (figura 6-1). En el embrión humano es tan sólo un vestigio de la gran estructura en forma de saco que usan los embriones de muchos mamíferos, aves y reptiles como órgano respiratorio principal y como depósito de los desechos urinarios. A semejanza del saco vitelino, la alantoides en los seres humanos tiene sólo una función secundaria, en este caso la respiración. En los seres humanos esta función la cumplen los vasos sanguíneos que se diferencian en la pared mesodérmica de la alantoides. Estos vasos conforman el arco circulatorio umbilical, que consta de las arterias y la vena que irrigan la placenta (figura 5-31) (el destino posnatal de estos vasos se explica en el capítulo 17).
CORION y PLACENTA
La formación del complejo placentario es el resultado de la interacción entre los tejidos extraembrionarios del embrión y los tejidos endometriales de la madre (las primeras etapas de la implantación del embrión y la reacción decidual del revestimiento uterino se describen en el capítulo 3). Después de que se ha completado la implantación, el trofoblasto inicial que rodea al embrión ha experimentado la diferenciación en dos capas: el citotrofoblasto y el sincitiotrofoblasto (figura 3-17 D). Las lagunas del trofoblasto, en rápido desarrollo, se han llenado de sangre materna y las células de tejido conectivo del endometrio han experimentado la reacción decidual (después de la cual contienen mayores cantidades de glucógeno y lípidos) como respuesta a la invasión del trofoblasto.
CORRELACiÓN CLíNICA 6-1
Situaciones relacionadas con el líquido amniótico
La cantidad normal de líquido amniótico al término del embarazo suele estar entre 500 y 1.000 mL. Una cantidad excesiva (más de 2.000 mL) recibe el nombre de hidramnios. Este fenómeno suele asociarse con embarazos múltiples y con atresia esofágica o anencefalia (anomalía congénita caracterizada por graves defectos de la cabeza y a menudo la incapacidad de deglutir [figura 7-4]). Estas pruebas circunstanciales apoyan el importante papel de la deglución fetal en el equilibrio total del intercambio de líquido amniótico. Muy poco líquido amniótico (menos de 500 mL) se denomina oligohidramnios. Esta alteración suele asociarse con la agenesia renal bilateral (ausencia de riñones) y apoya el papel de la excreción urinaria fetal en la dinámica del líquido amniótico. El oligohidramnios también puede ser la consecuencia de la rotura prematura de la membrana amniótica, que se presenta en aproximadamente el 10% de los embarazos.
Hay muchos componentes, tanto fetales como matemos, en el líquido amniótico; por ejemplo, en el líquido amniótico se han detectado más de 200 proteínas de origen matemo y fetal. Con las herramientas analíticas disponibles en la actualidad puede saberse mucho sobre el estado del feto mediante el análisis de la composición del líquido amniótico. La amniocentesis implica extraer una pequeña cantidad de líquido amniótico mediante la inserción de una aguja a través del abdomen de la madre y hasta la cavidad amniótica. A causa de la pequeña cantidad de líquido amniótico que tienen los embriones jóvenes, la amniocentesis no suele practicarse sino una vez cumplida la decimotercera o la decimocuarta semana del embarazo. El líquido amniótico tiene propiedades bacteriostáticas, lo cual puede explicar la baja incidencia de infecciones después de que se efectúa la amniocentesis.Las células fetales presentes en el líquido pueden cultivarse y detectar defectos cromosómicos y metOOólicos. Hay técnicas que permiten el examen de los cromosomas en las células fetales tan pronto como se obtienen, sin tener que esperar dos a tres semanas para que las células amnióticas cultivadas proliferen hasta el punto de ser apropiadas para el análisis genético. Además de la detección de los defectos cromosómicos (por ejemplo, las trisomías), es posible determinar el sexo del feto mediante el análisis directo de los cromosomas. Una elevada concentración de alfafetoproteína (una proteína del sistema nervioso central) en el líquido amniótico puede indicar un defecto del tubo neural. Puede evaluarse la madurez fetal mediante la determinación de la concentración de creatinina o la relación lecitina esfingomielina (que revela la madurez de los pulmones). La gravedad de la eritroblastosis fetal (enfermedad por Rh) también puede evaluarse examinando el líquido amniótico.
Formación de las vellosidades coriónicas
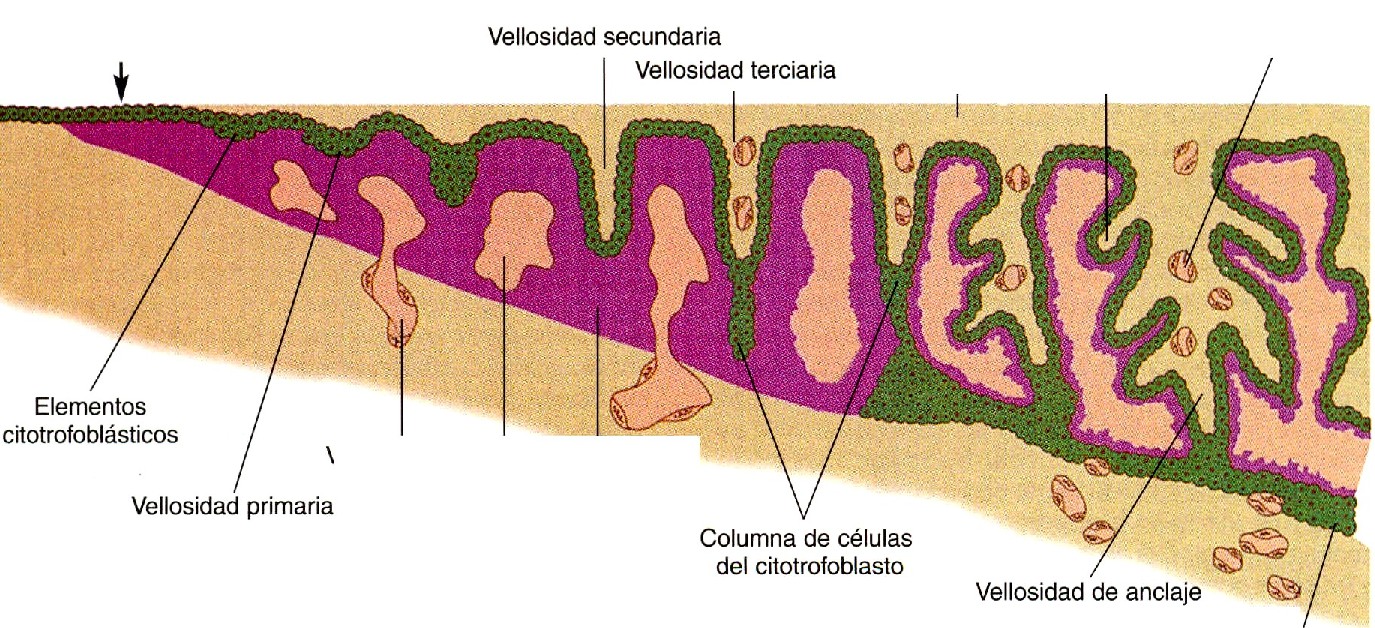
En el embrión en fase de implantación los tejidos trofoblásticos no tienen caracteristicas morfológicas macroscópicas homogéneas; en consecuencia, este período se llama del embrión prevelloso. Al final de la segunda semana comienzan a formarse unas proyecciones del citotrofoblasto llamadas vellosidades primarias (figura 4-2). Poco después, aparece una invasión mesenquimatosa en el interior de la vellosidad en crecimiento, que a partir de este momento se denomina vellosidad secundaria (figura 6-4). Rodeando al núcleo mesenquimatoso de la vellosidad secundaria se encuentra una capa completa de células del citotrofoblasto, y fuera de ellas está el sincitiotrofoblasto. Por definición, la vellosidad secundaria se convierte en vellosidad terciaria cuando en su núcleo mesenquimatoso entran vasos sanguíneos y se ramifican. Esto sucede hacia el final de la tercera semana del embarazo. Aunque cada vellosidad experimenta una considerable ramificación, la mayoria de ellas mantiene el mismo plan estructural básico durante todo el embarazo.
La porción terminal de la vellosidad sigue siendo trofoblástica, y consta de una masa sólida de citotrofoblasto llamada columna de células citotrofoblásticas (figura 6-4) y una cubierta relativamente delgada de sincitiotrofoblasto. La vellosidad está bañada por sangre materna. El desarrollo posterior del vértice de la vello si dad se produce cuando bajo la influencia del ambiente local hipóxico, la columna de células citotrofoblásticas crece en dirección distal, y penetra en la capa del sincitiotrofoblasto (figura 6-5). Estas células del citotrofoblasto se adosan directamente con las células de la decidua materna y se extienden sobre ellas para formar una capa completa de células denominada cápsula citotrofoblástica, que rodea al complejo embrionario. Las vellosidades que emiten las expansiones citotrofoblásticas se conocen como vellosidades de anclaje (figura 6-4) porque representan los verdaderos puntos de anclaje del complejo embrionario en los tejidos maternos.
Es importante entender las relaciones generales de los diversos tejidos embrionario s y maternos en esta etapa del desarrollo (figura 6-5). El embrión, conectado a través del tallo de conexión o cordón umbilical, está de hecho suspendido dentro de la cavidad coriónica. Esta cavidad estálimitada por la placa coriónica, que consta de mesodermo extraembrionario cubierto por trofoblasto. Las vellosidades coriónicas se extienden hacia fuera desde la placa coriónica y su cubierta de trofoblasto es continua con la de la placa coriónica. Las vellosidades y la superficie externa de la placa coriónica están bañadas por un mar de sangre materna en recambio continuo. A causa de ello, la placenta humana se clasifica como de tipo hemocorial. .
Aunque las vellosidades coriónicas tienen una estructura bastante compleja, conviene asimilar la estructura básica de un complejo vellositario al sistema de raíces de una planta. La vellosidad de anclaje es equivalente a la raíz central; por medio de las columnas de células citotrofoblásticas, se une al complejo vellositario con la cápsula citotrofoblástica externa. Las ramas no ancladas de las vellosidades flotantes (figura 6-12) flotan libres en la sangre materna que llena el espacio entre la placa coriónica y la cápsula citotrofoblástica externa. Todas las superficies de las vellosidades, la placa coriónica y la cápsula citotrofoblástica que están en contacto con la sangre materna están tapizadas por una capa continua de sincitiotrofoblasto.
Otros mamiferos tienen diferentes organizaciones de capas de tejido a través de las cuales los materiales tienen que pasar para intercambiarse entre la madre y el feto. Por ejemplo, en la placenta epiteliocorial, que se encuentra en el cerdo, el componente fetal de la placenta (corion) descansa sobre el epitelio uterino en vez de estar directamente bañado por la sangre materna.
La sangre materna entra en el espacio tapizado por el sincitiotrofoblasto a través de los extremos abiertos de las arterias espirales uterinas, que pasan a través de la cápsula citotrofoblástica externa. Las arterias han sido erosionadas por el trofoblasto invasor, pero sus luces de forma invariable están obstruidas parcialmente por un tapón de células del citotrofoblasto; se cree que esto es una adaptación para controlar el flujo sanguíneo. Durante las primeras doce semanas de vida embrionaria, el líquido que pasa a través de los espacios intervellosos es un filtrado de plasma materno y no contiene ninguna célula sanguínea. Durante este mismo período, los eritrocitos fetales contienen hemoglobina embrionaria que está adaptada para fijar oxígeno a tensiones bajas. Después de las doce semanas, los eritrocitos maternos aparecen en la sangre intervellosa y en una transición isomórfica los eritrocitos fetales comienzan a producir hemoglobina fetal, que requiere de una tensión más alta de oxígeno para fijar con efectividad el oxígeno. Después de dejar las arterias espirales a una presión relativamente alta, la sangre materna se filtra libremente a través de los espacios intervellosos y baña la superficie de las vellosidades. Luego la sangre, es recogida por los extremos abiertos de las venas uterinas, que también penetran en la cápsula citotrofoblástica (figura 6-10).
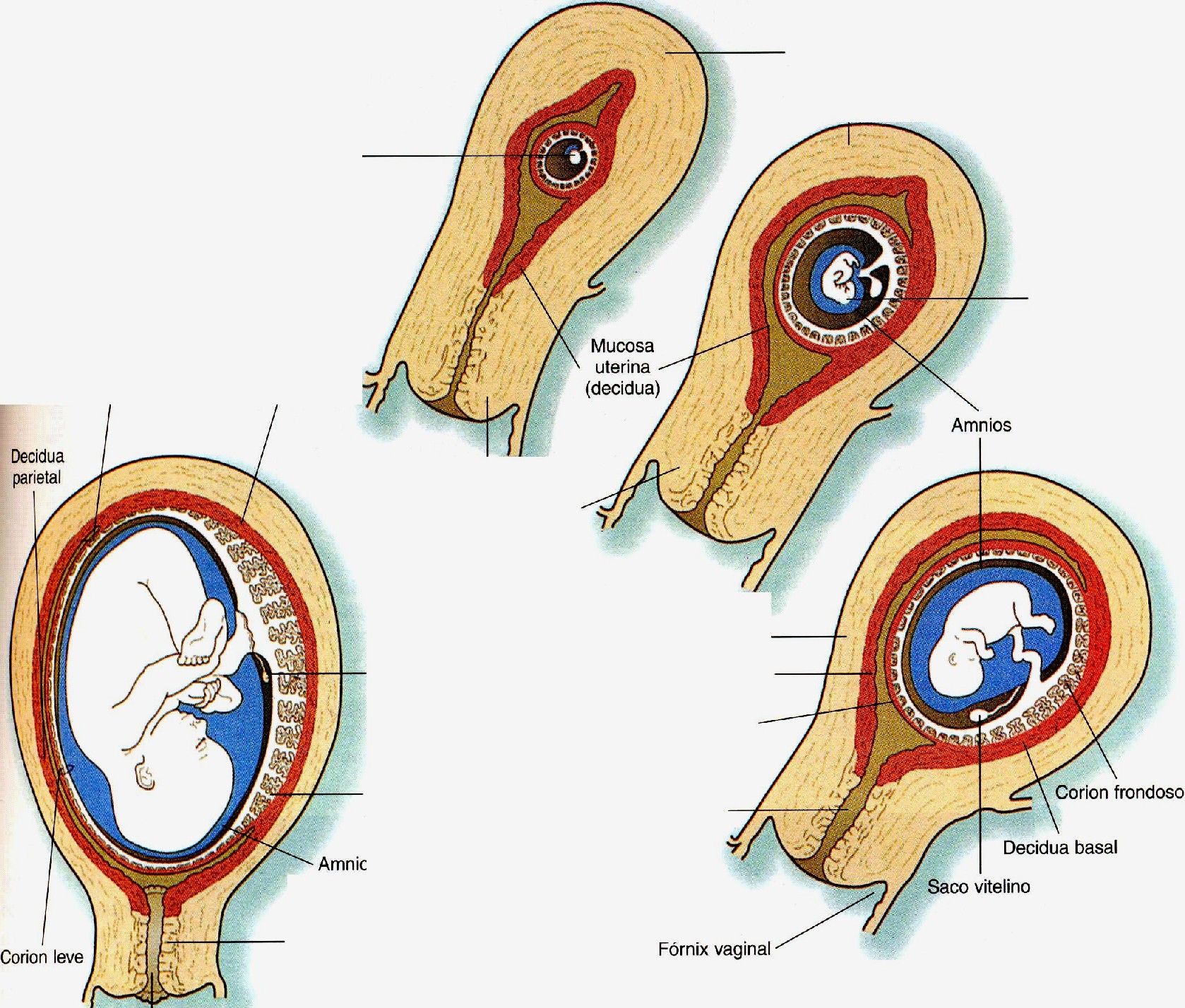
Relaciones macroscópicas de los tejidos coriónicos y deciduales
A los pocos días de la implantación del embrión, las células estromales del endometrio experimentan una importante transformación denominada reacción decidual*. Las células estromales se hipertrofian debido a la acumulación de glucógeno y lípidos en su citoplasma, y se les llama células deciduales (figura 6-6).La reacción decidual afecta a todas las células estromales de las capas superficiales del endometrío. La decidua materna recibe nombres topográfico s dependiendo de dónde esté localizada en relación con el embrión.
El tejido decidual que cubre al embrión y su vesícula coriónica es la decidua capsular, mientras que la decidua que se sitúa entre la vesícula coriónica y la pared uterina se llama decidua basal (figura 6-7). A consecuencia del crecimiento del embrión, la decidua basal se incorpora en el componente materno de la placenta definitiva. El resto de la decidua, que consta de tejido endometríal decidualizado en los lados del útero no ocupado por el embrión, es la decidua parietal.
En el embrión humano el corion está formado por el trofoblasto más el mesodermo extraembrionario subyacente (figura 6-1). El corion forma una cubierta completa (vesícula coriónica) que rodea al embrión, al amnios, al saco vitelino y al tallo de conexión.
* El término decidua (también llamada caduca) se refiere a los tejidos que se desprenden en el nacimiento, que incluyen los tejidos extraembrionarios más las capas superficiales del tejido conectivo y el epitelio endometrial..
Durante el período inicial posterior a la implantación, las vellosidades primarias y secundarias se evacuan de forma casi uniforme desde toda la superficie externa de la vesícula coriónica. La formación de las vellosidades terciarias es asimétrica, y la invasión del citotrofoblasto de las vellosidades primarias por el mesénquima y los vasos sanguíneos embrionario s se produce principalmente en las vellosidades primarias situadas más cerca de la decidua basa!. Conforme estas vellosidades siguen creciendo y ramificándose, las que están localizadas en el lado opuesto (el polo abembrionario) de la vesícula coriónica no se ramifican y terminan atrofiándose a medida que el saco embrionario en crecimiento protruye hacia la cavidad uterina (figura 6-7). La región que contiene las vellosidades coriónicas más activas y ramificadas y que se transformará en la placenta es el corion frondoso. El resto del corion, que termina siendo liso, es el corion leve o liso (figura 6-8).
El crecimiento general de la vesícula coriónica (figura 6-9), hacia la luz uterina, rechaza a la decidua capsular cada vez más lejos de los vasos sanguíneos endometriales. Hacia el final del primer trimestre, la propia decidua capsular experimenta una marcada atrofia. En el siguiente mes, algunas zonas de la decidua capsular atrófica comienzan a desaparecer, y dejan al corion leve en contacto directo con la decidua parietal del lado opuesto del útero (figura 6-7). Hacia la mitad de la gestación, el corion leve se ha fusionado con los tejidos de la decidua parietal; de este modo se oblitera por completo la cavidad uterina.
Mientras que el corion leve y la decidua capsular están sufriendo una atrofia progresiva, la placenta adquiere su forma definitiva y se configura como el único lugar de intercambio entre la madre y el embrión.
Formación y estructura de la placenta madura
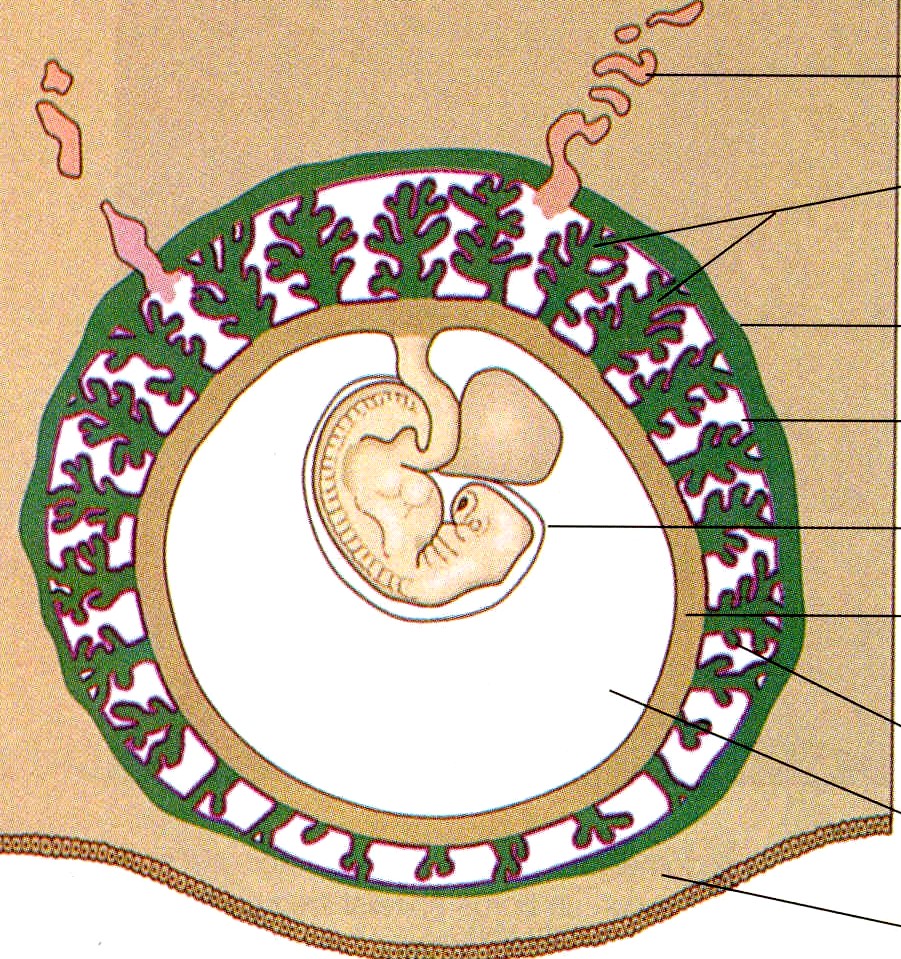
A medida que las diferencias entre el corion frondoso y el corion leve se hacen muy evidentes, se definen los límites de la placenta. Ésta consta de un componente materno y uno fetal (figura 6-10), el cual corresponde a la parte de la vesícula coriónica representada por el corion frondoso. Comprende la pared del corion, llamada placa coriónica, y las vellosidades coriónicas que surgen de esa región. El componente materno está representado por la decidua basal, pero lo que la cubre es la cápsula citotrofoblástica externa que se deriva del embrión. El espacio intervelloso entre los componentes fetal y materno de la placenta está ocupado por sangre materna que circula libremente. Para cumplir con su principal función como órgano que medio el intercambio entre los sistemas circulatorios materno y fetal, la morfología general de la placenta está estructurada para ofrecer una enorme superficie (más de10 m2) para dicho intercambio.
Estructura de la placenta madura
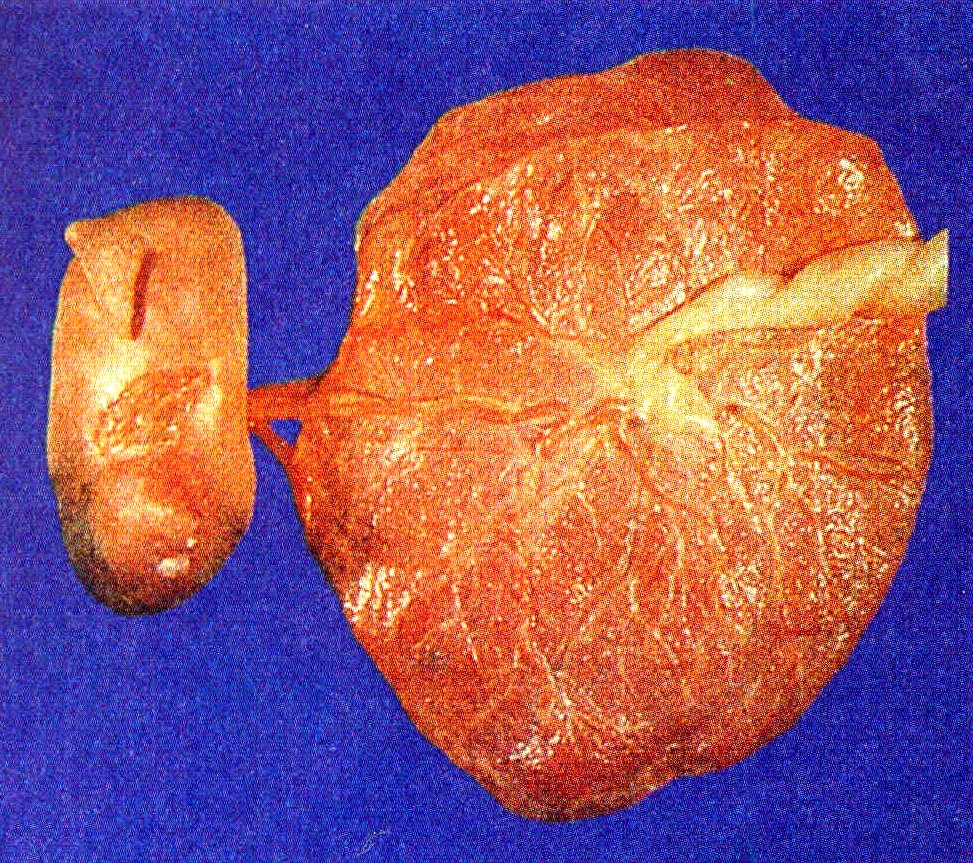
La placenta madura tiene forma de disco, con un grosor de 3 cm y un diámetro aproximado de 20 cm (tabla 6-1). Una placenta normal pesa unos 500 g. El lado fetal de la placenta es brillante a causa de la membrana amniótica que lo cubre. Desde el lado fetal pueden verse el punto de inserción del cordón umbilical en la placa coriónica y las grandes ramas placentarias de las arterias y la vena umbilicales que se irradian desde allí.
El lado materno de la placenta es opaco y está subdividido hasta 35 lóbulos. Los surcos entre los lóbulos están ocupados por tabiques placentarios, que surgen desde la decidua basal y se extienden hacia la placa basa!. En el interior de cada lóbulo placentario hay varios cotiledones, cada uno de los cuales consta de una vellosidad principal en forma de tronco y sus ramas. El espacio intervelloso de cada lóbulo representa un compartimiento casi aislado de circulación materna hacia la placenta.
Cordón umbilical
El tallo de conexión, que en un principio tenía una base amplia, se alarga y se adelgaza conforrnr avanza el embarazo. El cordón umbilical se convierte en el conducto de los vasos umbilicales, que discurren a todo lo largo de él entre el feto y la placenta (figura 6-10). Los vasos umbilicales están empotrados en un tejido conectivo mucoide que se llama gelatina de Wharton.
El cordón umbilical, que por lo general alcanza una longitud de 50 a 60 cm hacia el final del embarazo, suele estar retorcido muchas veces. La torsión puede observarse fácilmente en el examen macroscópico de los vasos sanguíneos umbilicales. En aproximadamente 1% de los embarazos a término se forman nudos verdaderos en el cordón. Si se aprietan como resultado de los movimientos fetal es, pueden causar anoxia e incluso la muerte del feto.
En ocasiones el cordón umbilical contiene dos venas si la vena umbilical derecha no sufre su degeneración normal (figura 16-11). Aproximadamente 0,5% de los cordones umbilicales maduros contienen una sola arteria umbilical. Este fenómeno se asocia con una incidencia del 15 al 20% de defectos cardiovasculares en el feto.
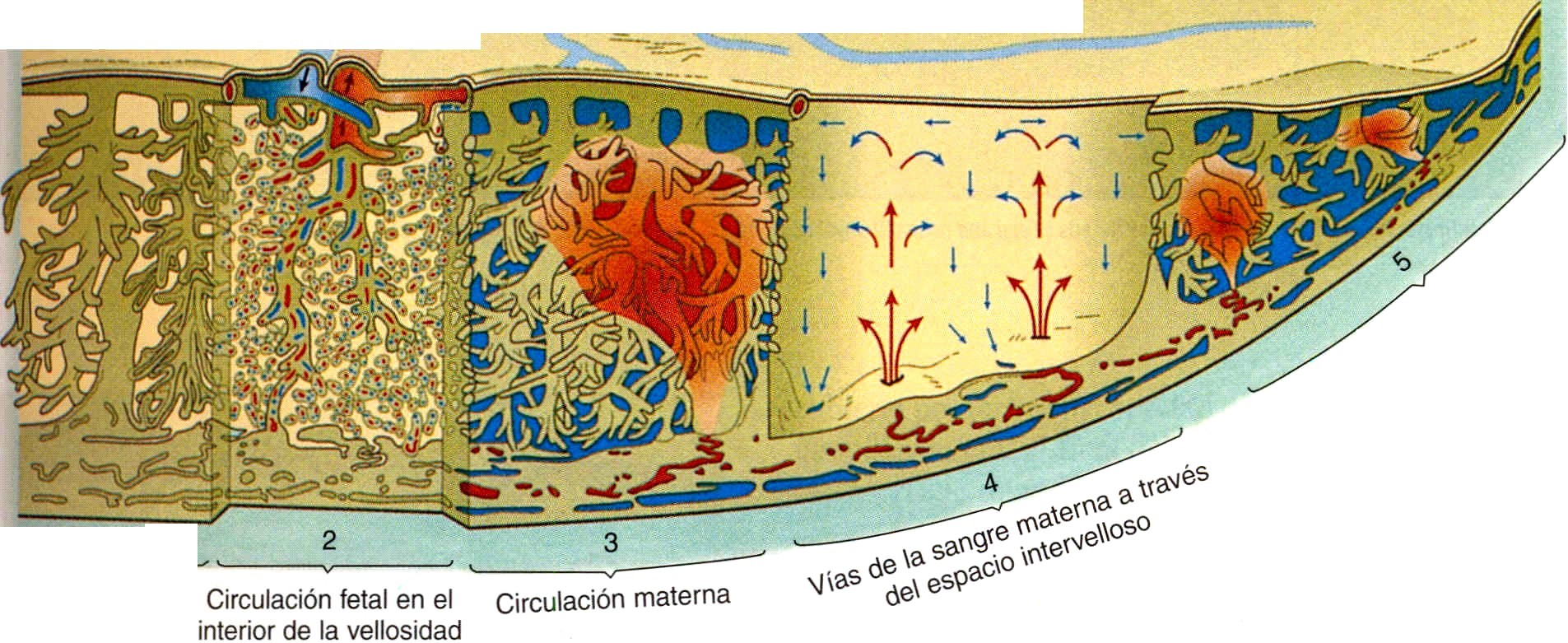
Tanto el feto como la madre contribuyen a la circulación placentaria (figura 6-10). La circulación fetal está formada por el sistema de los vasos umbilicales y placentarios. La sangre fetal llega a la placenta a través de las dos arterias umbilicales, que se ramifican por toda la placa coriónica. Las ramas, más pequeñas de estas arterias entran en las vellosidades coriónicas y después se dividen en redes capilares en las ramas ¡ terminales de las vellosidades coriónicas, donde tiene lugar el intercambio de materiales con la sangre materna (figura 6-14 F). Desde el lecho capilar velloso, los vasos sanguíneos se reúnen en ramas venosas cada vez más gruesas. Éstas recorren inversamente el camino a través de la placa coriónica hacia la gran vena umbilical única y de vuelta al feto.
En contraste con la circulación fetal, que discurre siempre dentro de los vasos sanguíneos, el abastecimiento de sangre materna a la placenta es un lago de flujo libre que no está limitado por paredes vasculares. Como resultado de las actividades invasoras del trofoblasto, entre 80 y 100 arterias espirales uterinas se abren directamente a los espacios intervellosos y bañan las vellosidades con unos 150 mL de sangre materna, que se recambia de tres a cuatro veces por minuto.
La sangre materna entra en el espacio intervelloso a presión reducida por los tapones de citotrofoblasto que ocluyen parcialmente las luces de las arterias espirales. No obstante, la presión de la sangre materna es suficiente para forzar el paso de la sangre arterial materna oxigenada hacia las bases de las ramificaciones vellosas en la placa coriónica (figura 6-10). La presión global de la sangre materna en la placenta es de unos 10 mmHg cuando el útero está relajado. Desde la placa coriónica, la sangre se filtra sobre la vellosidad terminal a medida que regresa hacia las vías de drenaje venoso localizadas en la placa decidual (materna) de la placenta. Un flujo adecuado de sangre materna a la placenta es vital para el crecimiento y el desarrollo del feto, y un aporte reducido de sangre a la placenta conduce a un feto pequeño.
En las vellosidades terminales (flotantes), los capilares fetales están localizados cerca de la superficie trofoblástica para facilitar el intercambio entre la sangre fetal y la materna (figura 6-11). La barrera placentaria de la placenta madura está compuesta por el sincitiotrofoblasto, su lámina basal, la lámina basal de los capilares fetales y el endotelio A menudo las dos láminas basales parecen haberse, fusionado. En los embriones más pequeños hay una capa de ciotrofoblasto en la barrera placentaria, pero hacia los cuatro meses esta capa comienza a atrofiarse y a los cinco meses prácticamente ha desaparecido (la transferencia placentaria describe en la pág. 121).
Estructura de la vellosidad coriónica madura
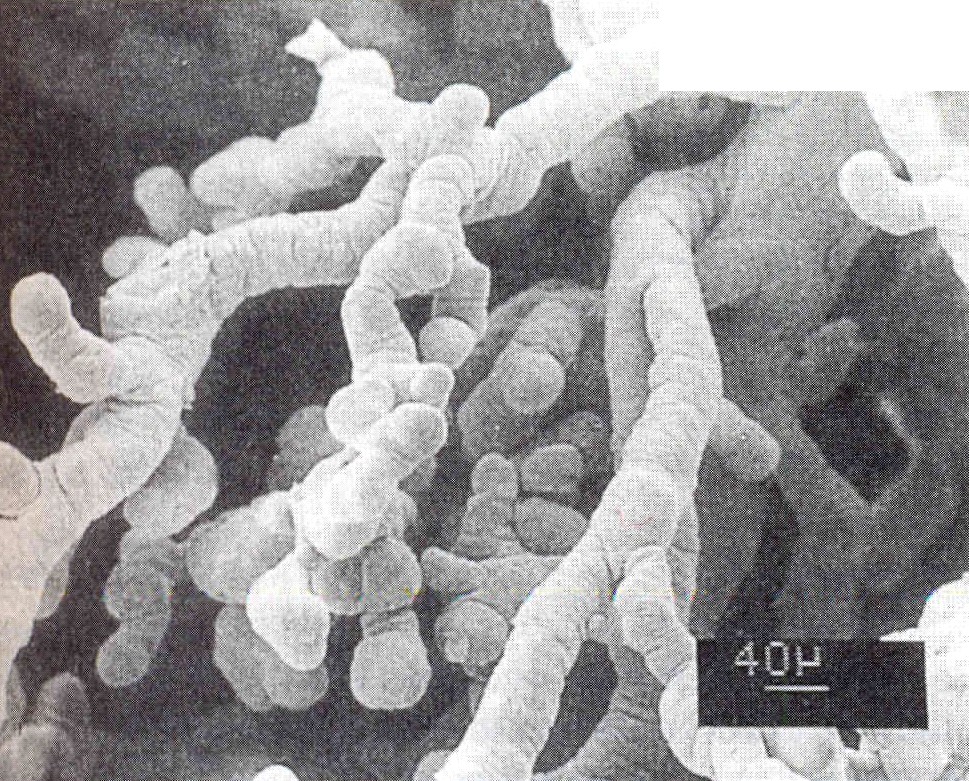
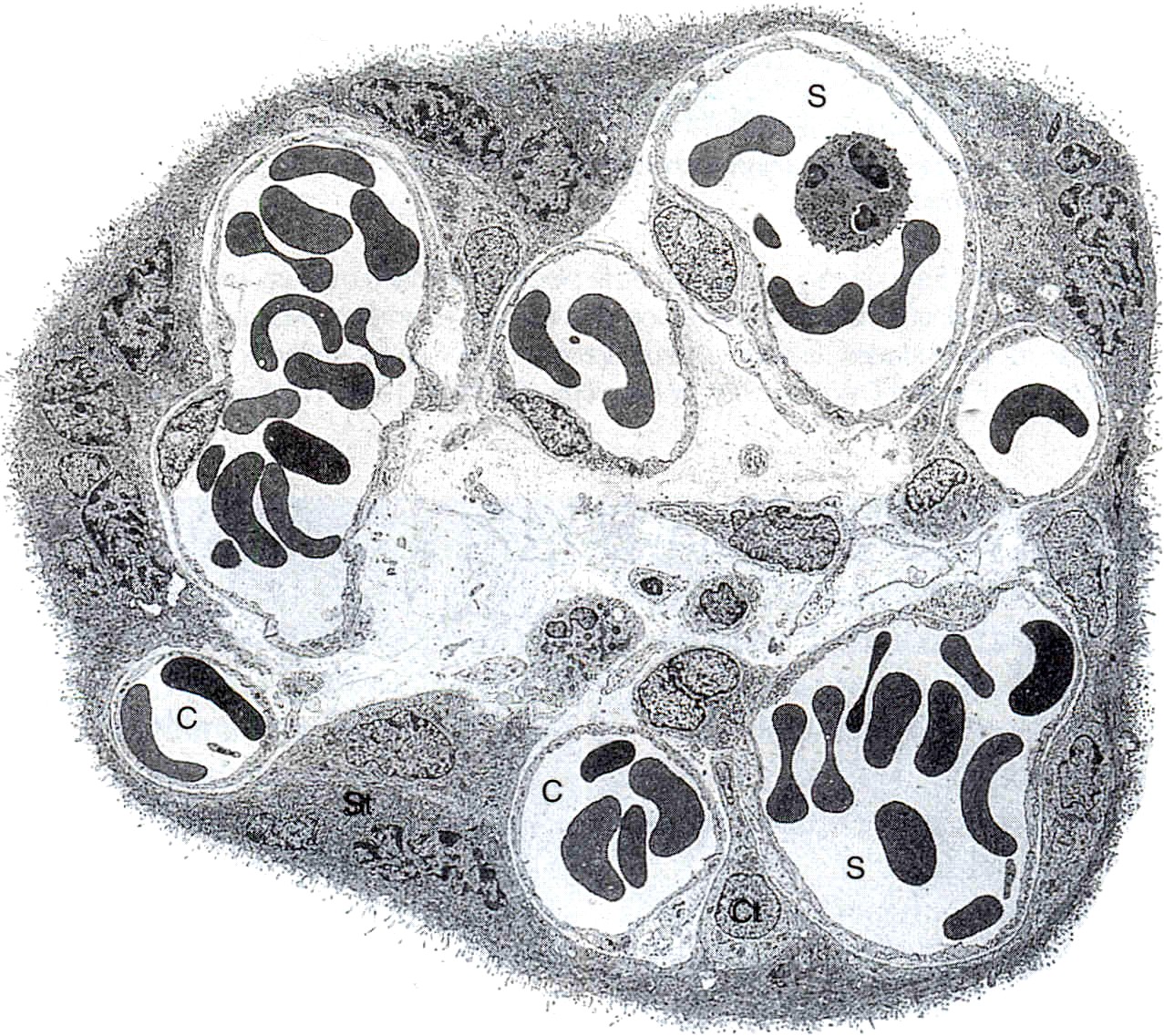
Las vellosidades coriónicas maduras constituyen una masa muy compleja de ramas aparentemente entrecruzadas (figura 6-12). El núcleo de la vellosidad está compuesto por vasos sanguíneos y mesénquima cuya composición es similar a la del mesénquima del cordón umbilical (figura 6-11). Dispersas entre las células mesenquimatosas se encuentran las grandes células de Hoftbauer que funcionan como macrófagos fetales.
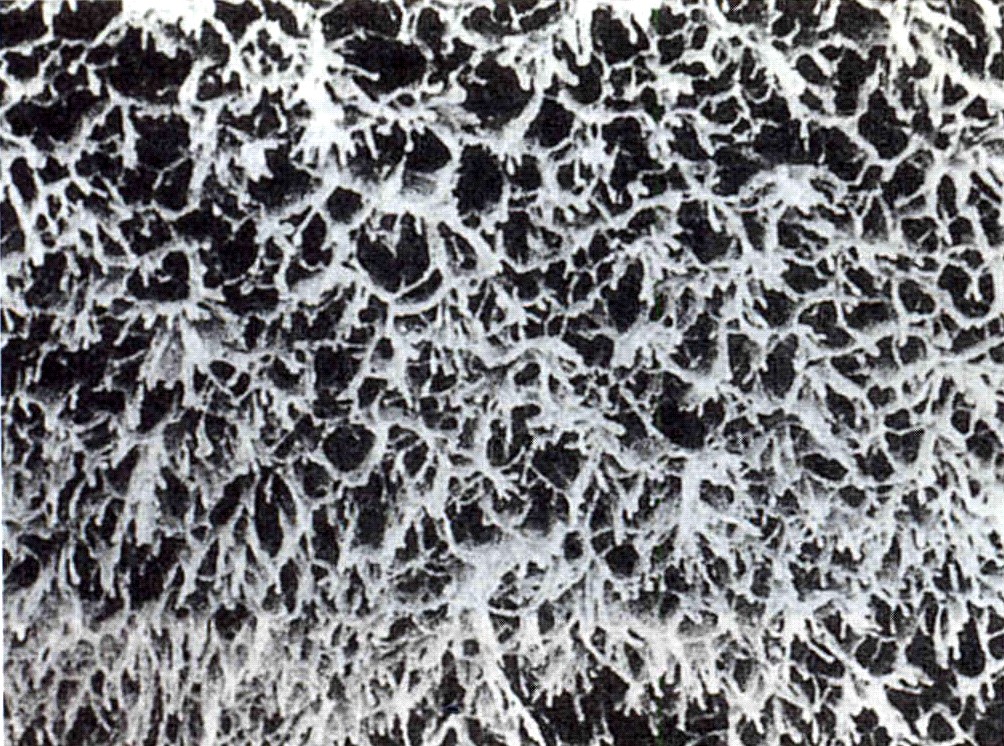
El núcleo de la vellosidad está cubierto por una capa continua de sincitiotrofoblasto, con pocas células de citotrofoblasto debajo de él. La superficie del sincitiotrofoblasto está cubierta por una gran cantidad de microvellosidades (más de un millardo/cm2 al final del embarazo) que aumentan mucho la superficie total de la placenta (figura 6-13). El tamaño y la densidad de las microvellosidades no son constantes sino que cambian con la edad de la placenta y en función de las distintas condiciones ambientales. Por ejemplo, en casos de mala nutrición materna o transporte deficiente de oxígeno, el número de las microvellosidades aumenta. Una mala adaptación de éstas a las condiciones adversas puede producir recién nacidos de bajo peso al nacer (crecimiento intrautero retrasado).
La superficie del trofoblasto no es homogénea, sino que parece estar organizada en territorios. Entre los distintos componentes funcionales de la superficie de microvellosidades se encuentran: 1) una gran variedad de sistemas de transporte de sustancias que van desde iones hasta macromoléculas, 2) receptores de hormonas y factores de crecimiento, 3) enzimas y 4) numerosas proteínas con funciones hasta ahora poco claras. La superficie placentaria es deficiente o carece de antígenos de histocompatibilidad importantes, cuya ausencia se presume que influye en la protección del feto y las membranas contra el rechazo inmunológico por la madre. Para poder cumplir con su papel activo tanto en la síntesis como en el transporte, el sincitiotrofoblasto tiene una alta densidad y amplia diversidad de orgánulos subcelulares.
Fisiología placentaria
Transferencia placentaria
El transporte de sustancias en doble vía entre la placenta y la sangre materna que la baña se facilita gracias a la gran superficie de la placenta, que crece desde 5 m2 a las 28 semanas, hasta casi 11 m2 al término. Aproximadamente del 5 al 10% de la superficie de la placenta humana consta de áreas dispersas donde la barrera entre la sangre fetal y la materna es muy delgada, ya que mide sólo unas pocas micras. Estas áreas, que en ocasiones se llaman placas epiteliales, parecen ser adaptaciones morfogenéticas cuyo fin es facilitar la difusión de sustancias entre las circulaciones materna y fetal (figura 6-14 F).
La transferencia de sustancias tiene lugar en ambas direcciones a través de la placenta. La mayor parte de las que se transfieren de la madre al feto son oxígeno y nutrientes. La placenta constituye el medio para la eliminación final del dióxido de carbono y otros materiales fetales de desecho hacia la circulación materna. En algunas circunstancias, otras sustancias, algunas de ellas nocivas, pueden transferirse a través de la placenta. La Correlación clínica 6-2 describe una transferencia placentaria anormal.
Los gases, principalmente oxígeno de la madre y dióxido de carbono del feto, cruzan con facilidad la barrera placentaria por simple difusión. El límite para la cantidad de intercambio es más el flujo sanguíneo que la eficiencia de la difusión. La placenta también es permeable al monóxido de carbono y a muchos anestésicos inhalatorios. Estos últimos pueden entorpecer la adaptación del recién nacido a un funcionamiento autónomo (por ejemplo, la respiración) si se usan sin precaución durante el parto.
A semejanza de los gases, el agua y los electrólitos se transfieren fácilmente a través de la placenta. Las tasas de transferencia se modifican debido a la presión coloidosmótica en el caso del agua y a la función de los canales iónicos en el caso de los electrólitos. Los desechos fetales (por ejemplo, la urea, la creatinina y la bilirrubina) se transfieren con rapidez a través de la placenta desde la circulación fetal hacia la sangre materna que baña las vellosidades.
La placenta tiene una alta permeabilidad a ciertos nutrientes como la glucosa; por otra parte, es mucho menos permeable a la fructosa y a los disacáridos más comunes. Los aminoácidos se transportan a través de la placenta gracias a la acción de receptores específicos. Existe un cierto grado de transferencia de ácidos grasos libres de la madre al feto, pero queda mucho por averiguar sobre el mecanismo de esta transferencia. Las vitaminas, en especial las hidrosolubles, se transfieren de la circulación materna a la fetal.
Las hormonas esteroideas atraviesan la barrera placentaria desde la sangre materna. Los neonatos varones muestran evidencias de los efectos de exposición a las hormonas sexuales maternas; por ejemplo, el utrículo prostático -el vestigio de la formación del útero (conductos müllerianos fusionados [capítulo 15]- está levemente aumentado. Por el contrario, los fetos femeninos expuestos a la testosterona o a ciertos progestágenos sintéticos (en especial durante las décadas de 1950 y 1960, antes de que fuesen conocidos sus efectos) sufren masculinización de los genitales externos. Las hormonas proteicas, por lo general, se transportan poco a través de la placenta, aunque los síntomas de la diabetes materna pueden reducirse hacia el final del embarazo gracias a la insulina producida por el feto. La hormona tiroidea materna pasa en escasa cantidad al feto.
Algunas proteínas se transfieren muy lentamente a través de la placenta, y lo hacen sobre todo por medio de la pinocitosis (captación por vesículas rodeadas por la membrana en las células). La transferencia de anticuerpos maternos, sobre todo las inmunoglobulinas de clase G, reviste gran importancia. Debido a la inmadurez de su sistema inmunológico, el feto sólo produce pequeñas cantidades de anticuerpos. La transferencia de anticuerpos de la madre confiere al recién nacido una inmunidad pasiva contra ciertas enfermedades comunes de la infancia como la viruela, la difteria y el sarampión hasta que el sistema inmuno1ógico del bebé comienza a funcionar con más efectividad.
Otra proteína materna, la transferrina, es importante porque, como su nombre lo indica, transporta hierro hacia el feto. La superficie placentaria contiene receptores específicos para esta proteína. Parece que el hierro se disocia de su transportador, la transferrina, en la superficie p1acentaria y después se transporta activamente a los tejidos fetales.
Correlación CLÍNICA 6-2
Transferencia placentaria anómala
Infortunadamente, la placenta es permeable a sustancias que pueden ser dañinas para el embrión. Numerosos medicamentos ingeridos por la madre cruzan con facilidad la barrera placentaria. Ciertos medicamentos pueden ocasionar graves malformaciones congénitas si alcanzan al embrión durante los períodos críticos de la morfogénesis (en el capítulo 7 se describen varios ejemplos clásicos de esta situación). La placenta es muy permeable al alcohol; la ingestión excesiva de esta sustancia por parte de la madre puede producir el síndrome alcohólico fetal (pág. 143). La tragedia de los neonatos que nacen adictos a la heroína o al "crack" es muy común en la sociedad contemporánea.
Además de los medicamentos, ciertos agentes infecciosos pueden penetrar la barrera placentaria e infectar al feto. Algunos (como el virus de la rubéola) pueden causar malformaciones si infectan al embrión en períodos criticas del desarrollo. Normalmente las bacterias no pueden cruzar la barrera placentaria. Los virus que pueden infectar al feto son el de la rubéola, el citomegalovirus, el virus de la polio, el de la varicela, el de la viruela, el de inmunodeficiencia humana y los coxsackie. La espiroqueta causante de la sífilis, el Treponema pallidum, puede ocasionar devastadoras infecciones fetales. El Toxoplasma gondii puede atravesar la barrera placentaria y causar defectos congénitos.
TRANSFERENCIA CELULAR E INCOMPATIBILIDAD Rh.
Pequeñas cantidades de sangre fetal suelen escapar hacia la circulación materna, ya sea a través de pequeños defectos en la red vascular placentaria, o por la hemorragia durante el parto. Si los eritrocitos fetales son positivos para el antígeno Rh y la madre es Rh-negativa, la presencia de eritrocitos fetales en la circulación materna puede estimular al sistema inmunológico de la madre a que forme anticuerpos anti-Rh. El feto primogénito por lo general se salva de los efectos de los anticuerpos maternos (a menudo porque todavía no se han formado en cantidades suficientes), pero en los embarazos posteriores los fetos Rh-positivos son atacados por los anticuerpos anti-Rh maternos, que pasan a la circulación fetal. Estos anticuerpos provocan la hemólisis de los eritrocitos fetales Rh-positivos, y el feto sufre eritroblastosis fetal que también se conoce como enfermedad hemolítica. En los casos graves, la bilirrubina liberada por los glóbulos rojos hemolizados causa ictericia y daño cerebral además de la anemia. Cuando se diagnostica, la enfermedad se trata con exsanguino transfusiones de sangre de donante Rh-negativo ya sea al feto o al recién nacido. Mediante el análisis del líquido amniótico puede obtenerse una indicación sobre la gravedad de la enfermedad.
Síntesis y secreción de las hormonas placentarias
La placenta, específicamente el sincitiotrofoblasto, es un importante órgano endocrino durante la mayor parte del embarazo. Produce tanto hormonas esteroideas como proteicas.
La primera proteohormona en producirse es la gonadotropina coriónica humana (HCG), que es responsable de mantener al cuerpo lúteo y su producción de progesterona y estrógenos. Puesto que su síntesis comienza incluso antes de la implantación, la presencia de esta hormona en la orina materna constituye la base de muchas de las pruebas de diagnóstico de embarazo. La producción de HCG es máxima aproximadamente en la octava semana de embarazo y después se reduce de forma gradual. Hacia el final del primer trimestre, la placenta produce estrógenos y progesterona en cantidad suficiente para que pueda mantenerse el embarazo incluso si el cuerpo lúteo se extirpa quirúrgicamente. La placenta puede sintetizar progesterona en forma independiente a partir de precursores de acetato o colesterol, pero no contiene el aparato enzimático completo para la síntesis de los estrógenos. Para poder sintetizar estrógenos la placenta debe operar en armonía con la glándula suprarrenal y posiblemente el hígado fetal; estas estructuras poseen las enzimas de las cuales carece la placenta.
Otra proteohormona placentaria es la somatomamotropina coriónica también llamada lactógeno placentario humano. Similar en estructura a la hormona de crecimiento humana, influye sobre el crecimiento, la lactancia y el metabolismo de los carbohidratos y los lípidos. La placenta también produce pequeñas cantidades de tirotropina coriónica y de corticotropina coriónica. Cuando se segregan hacia la circulación materna, algunas de las hormonas placentarias estimulan cambios en el metabolismo y la función cardiovascular de la madre, los cuales aseguran que los tipos y las cantidades apropiadas de nutrientes y sustratos fundamentales alcancen la placenta y se transporten al feto.
Un buen ejemplo de una hormona placentaria que influye sobre la madre es la hormona de crecimiento placentaria humana. Esta hormona, que difiere en trece aminoácidos de la hormona de crecimiento hipofisiaria, se produce en el sincitiotrofoblasto. La hormona placentaria de crecimiento no es detectable en el suero fetal, aunque al parecer influye sobre el crecimiento de la placenta de manera paracrina. Esta hormona fetal ejerce un profundo efecto en la madre. Durante las primeras quince a 20 semanas del embarazo la hormona de crecimiento hipofisiaria materna es la principal forma hormonal presente en la circulación de la madre, pero a partir de las quince semanas hasta el término, esta hormona es remplazada gradualmente por la hormona de crecimiento placentaria hasta el punto que la hormona materna deja de ser detectab1e en el suero materno. Una de las funciones principales de esta hormona parece ser la regulación de los niveles de glucosa en sangre de la madre para asegurar al feto un aporte adecuado de nutrientes. La secreción de la hormona placentaria de crecimiento se estimula por los niveles bajos de glucemia. El aumento de los niveles de la hormona estimula la gluconeogénesis en el hígado y en otros órganos de la madre, aumentando así los niveles de glucosa disponibles para el uso del feto.
En ciertos aspectos, la placenta duplica el sistema de control multinive1 que regula la producción de hormonas en el recién nacido. Las células del citotrofoblasto producen un homólogo de la hormona liberadora de gonadotropinas (GnRH), similar a como 10 hace el hipotálamo. La GnRH pasa entonces al interior del sincitiotrofob1asto en donde, con ciertos péptidos opiáceos y sus receptores (que han sido identificados en el sincitiotrofoblasto), estimulan la liberación de la HCG a partir del sincitiotrofoblasto. Los péptidos opiáceos y sus receptores también intervienen en la liberación de la somatomamotropina coriónica a partir del sincitiotrofoblasto. Por último, parece que la HCG interviene en la regulación de la síntesis de los esteroides placentarios en el sincitiotrofoblasto.
Además de las hormonas, la p1acenta produce una gran diversidad de otras proteínas que en principio se han identificado con técnicas inmunológicas. Las funciones de las docenas de proteínas placentarias que se han descubierto aún no son claras.
Inmunología placentaria
Uno de los mayores misterios del embarazo es por qué el feto y la placenta, que desde el punto de vista inmunológico son diferentes de la madre, no son reconocidos como tejidos extraños y rechazados por el sistema inmunológico de ella (el rechazo inmunológico de los tejidos extraños normalmente ocurre debido a la activación de los linfocitos citotóxicos, pero también son posibles las respuestas humorales inmunes). A pesar de la multitud de investigaciones que se han dedicado a este tema, la respuesta a esta pregunta aún se desconoce. Se han sugerido varias explicaciones amplias para la especial tolerancia de la madre a la presencia prolongada del embrión, inmunológicamente diferente, durante el embarazo.
La primera posibilidad es que los tejidos fetales, en especial los de la placenta, que constituyen la interfase directa entre el feto y la madre, no presenten antígenos extraños al sistema inmunológico de la madre. Hasta cierto punto, esta hipótesis es verdadera porque ni el sincitiotrofoblasto ni el citotrofoblasto no velloso (cápsula citotrofoblástica) expresan los dos tipos principales de antígenos mayores de histocompatibilidad que desencadenan la respuesta inmunológica del huésped en el rechazo de los injertos de tejidos (por ejemplo, un trasplante de riñón). Sin embargo, estos antígenos están presentes en las células del feto y en los tejidos estromales de la placenta. La expresión de los antígenos menores de histocompatibilidad (por ejemplo, el antígeno HY en fetos masculinos [capítulo 15]) sigue un patrón similar. No obstante, los tejidos trofoblásticos expresan otros antígenos menores. Además, a causa de las brechas existentes en la barrera placentaria, es frecuente encontrar glóbulos fetales rojos y blancos circulando en la sangre materna. Estas células deberían poder sensibilizar al sistema de defensas de la madre.
La segunda explicación es que el sistema inmunológico de la madre sufre una especie de bloqueo durante el embarazo, de manera que no reacciona a los antígenos totales a los cuales se expone. Pese a ello, la madre es capaz de poner en marcha una respuesta inmunológica a la infección o al injerto de un tejido extraño. Parece probable que exista una represión selectiva del sistema inmunológico ante los antígenos fetales, aunque la respuesta de incompatibilidad Rh muestra que ésta no debe de ser total.
Una tercera posibilidad es que la barrera decidual local impida el reconocimiento inmunológico del feto por parte de la madre o el paso de células inmunocompetentes de la madre al feto. Hay pruebas de la existencia de una barrera decidual funcional, pero en un número importante de casos se sabe que esa barrera se rompe debido a traumatismos o enfermedades.
En la actualidad, los estudios están dirigidos a situaciones como el aborto espontáneo de repetición, con la esperanza de encontrar otras claves sobre las complejas interrelaciones inmunológicas entre el feto y la madre. En todo caso, es evidente que no se trata de una relación sencilla. Sin embargo, la solución a este problema puede aportar datos que podrían aplicarse a la problemática del rechazo de trasplantes de órganos y tejidos.
LA PLACENTA DESPUÉS DEL NACIMIENTO
Unos 30 minutos después del parto, la placenta, las membranas embrionarias y el resto del cordón umbilical, junto con gran parte de la decidua materna, son expulsados del útero en lo que se denomina alumbramiento o cuarto período del parto. La superficie fetal de la placenta es lisa, brillante y grisácea a causa del amnios que cubre el lado fetal de la placa coriónica. La superficie materna es de un rojo mate y puede tener coágulos de sangre; es necesario examinada con cuidado porque si falta un cotiledón y éste se queda retenido en la pared uterina, puede causar una seria hemorragia en el posparto. Conocer algunos tipos de patologías placentarias puede arrojar información sobre los factores intrauterinos susceptibles de afectar el bienestar del recién nacido (Correlación clínica 6-3).
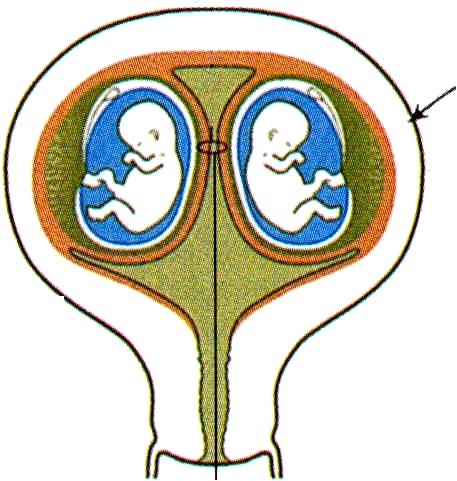
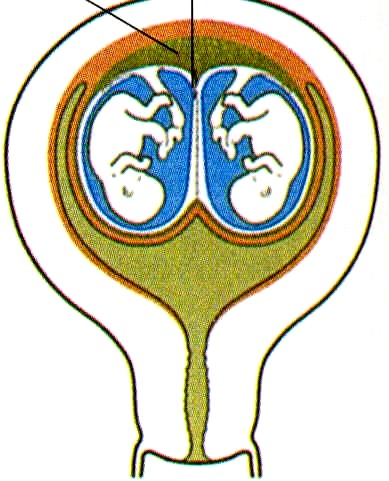
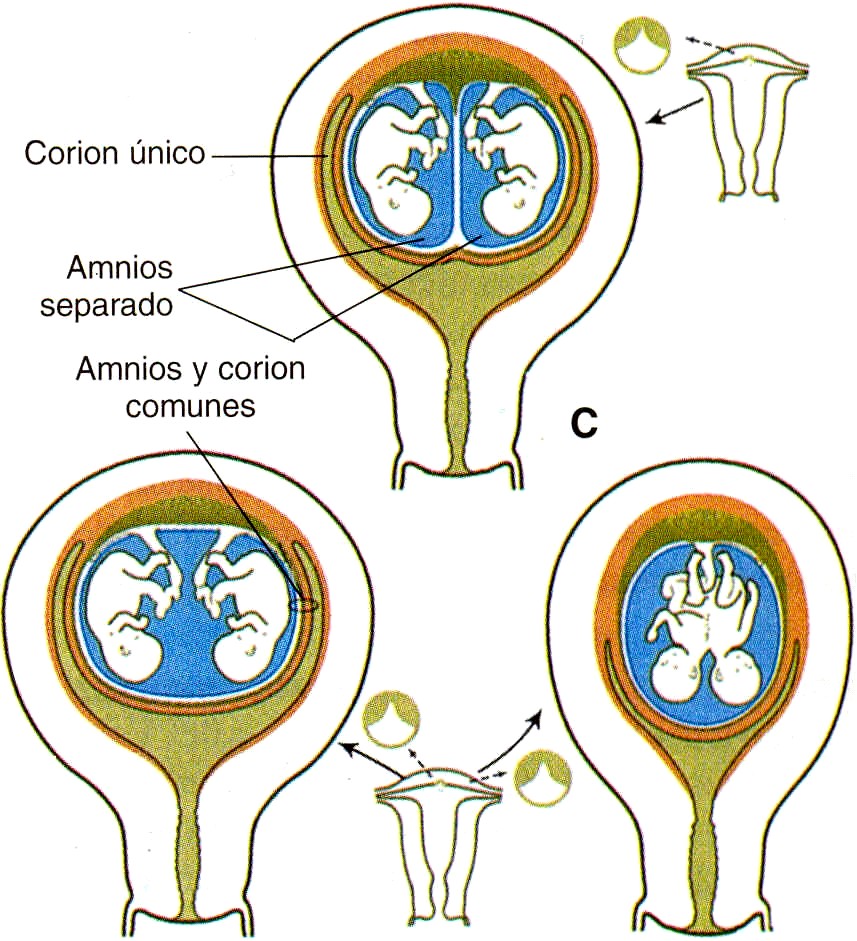
LA PLACENTA y LAS MEMBRANAS EN LOS EMBARAZOS MÚLTIPLES
Durante los embarazos múltiples son posibles varias configuraciones diferentes de la placenta y las membranas extraembrionarias. Los gemelos dicigóticos o monocigóticos que resultan de la separación completa de los blastómeros muy al comienzo de la segmentación pueden tener placentas y membranas separadas si los dos embriones se implantan en lugares distantes en la pared uterina (figura 6-15-A). En contraste, si los lugares de implantación están muy cerca uno del otro, las placentas y los corion (que al comienzo de la implantación estaban separados) pueden fusionarse, aunque los sistemas vasculares de los dos embriones permanecen separados (figura 6-15 B). Cuando los gemelos monocigóticos se forman por la separación de la masa celular interna del blastocisto, lo frecuente es que tengan una placenta y un corion común, pero en el interior del mismo cada embrión se desarrolla dentro de una bolsa amniótica adherente (figura 6-15 C). En este caso, los sistemas vasculares pueden estar separados o fusionados en la placenta común. Cuando los sistemas vasculares están fusionados, un gemelo puede recibir una proporción mayor del flujo sanguíneo placentario que el otro; esto puede redundar en un retraso leve a severo del crecimiento del embrión que recibe menos sangre de la placenta. El gemelo del cual la sangre es transfundida al otro suele estar gravemente malformado y recibe el nombre de monstruo acárdiaco (figura 6-16).
CORRELACIÓN CLÍNICA 6-3
Afecciones placentarias
Las afecciones placentarias cubren un amplio espectro, que va desde las anomalías del lugar de implantación hasta las neoplasias y las infecciones bacterianas. Puede aprenderse mucho de la historia pasada y de las perspectivas del recién nacido cuando se examina la placenta. Esta correlación sólo tiene que ver con aquellos aspectos de la patología placentaria que son importantes para los mecanismos del desarrollo.
LUGARES DE IMPLANTACIÓN ANÓMALOS
Un lugar anómalo de implantación dentro de la cavidad uterina se conoce como placenta previa (el embarazo ectópico se explica en el capitulo 3). Cuando una parte de la placenta cubre el orificio cervical de la cavidad uterina, su presencia es un obstáculo mecánico en el canal del parto. Además, la hemorragia, que puede ser mortal para el feto e incluso para la madre, es una consecuencia habitual de la placenta previa como resultado de la separación prematura de parte de la placenta del útero.
CORRELACIÓN CLÍNICA 6-3
Afecciones placentarias (continuación)
ANOMALíAS PLACENTARIAS MACROSCÓPICAS
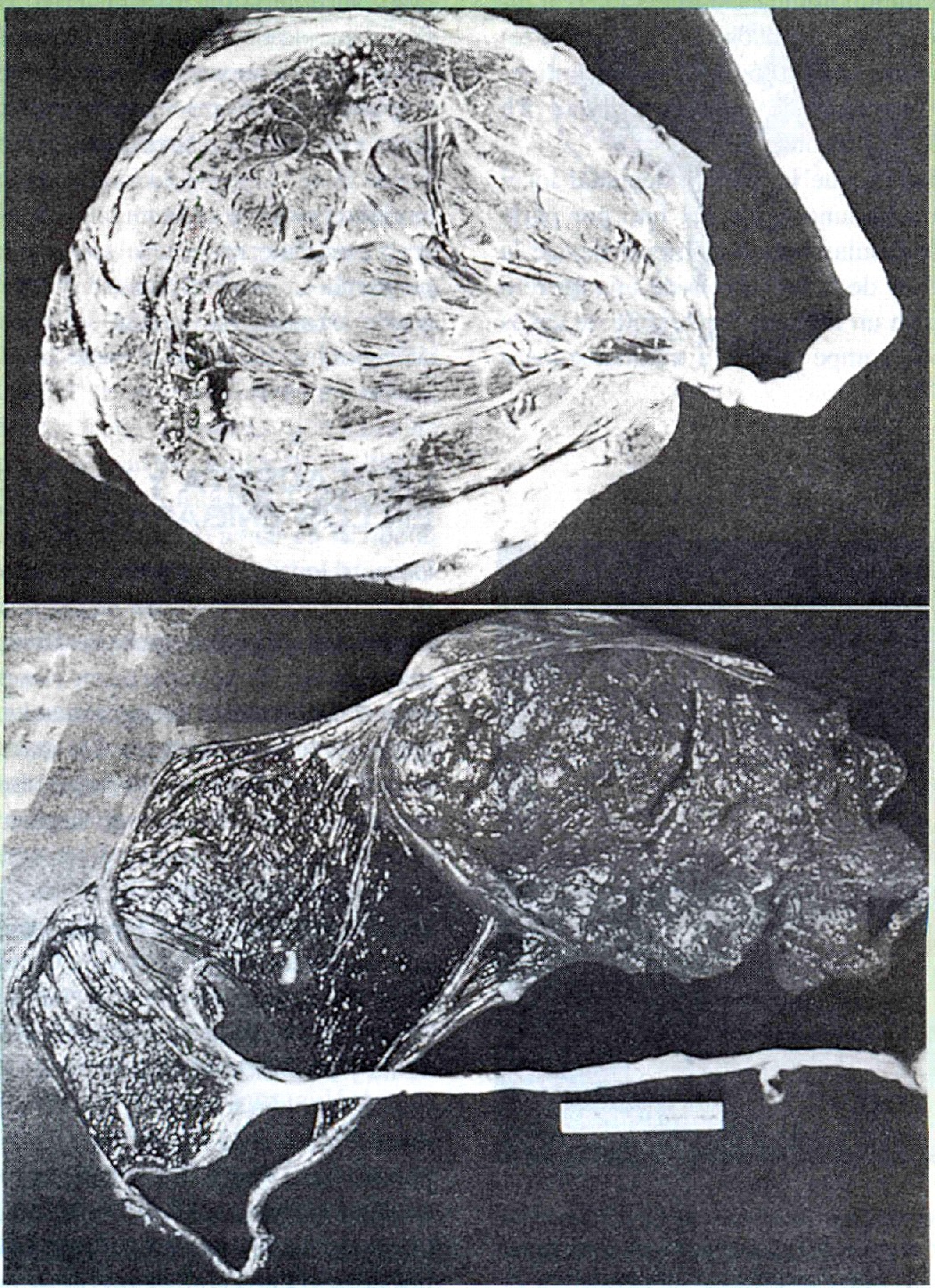
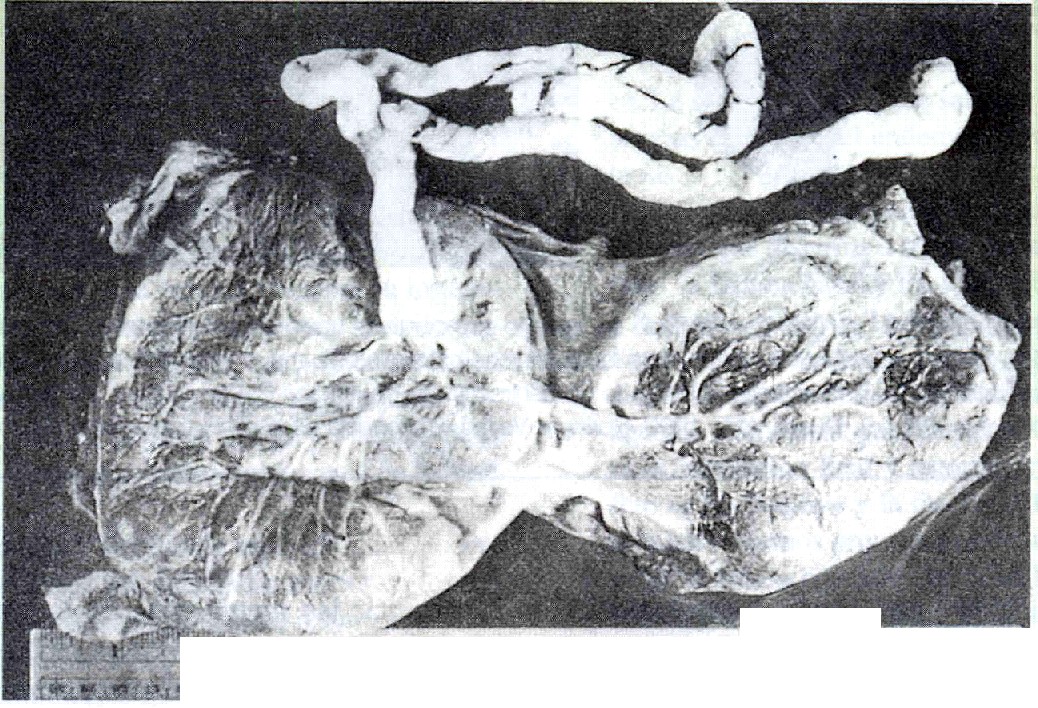
Se han descrito muchas variaciones en la forma de la placenta, pero pocas parecen tener alguna importancia funcional. Una de ellas es la inserción marginal en lugar de central del cordón umbilical (figura 6-17 A). Si el cordón umbilical se inserta en las membranas por fuera de los límites de la placenta misma, se conoce como inserción velamentosa del cordón umbilical (figura 6-17 B).
La placenta propiamente dicha puede subdividirse en lóbulos accesorios (figura 6-17 C), o estar dividida por completo en dos partes por una membrana lisa en medio (figura 6-17 d).
MOLA HIDATlDIFORME
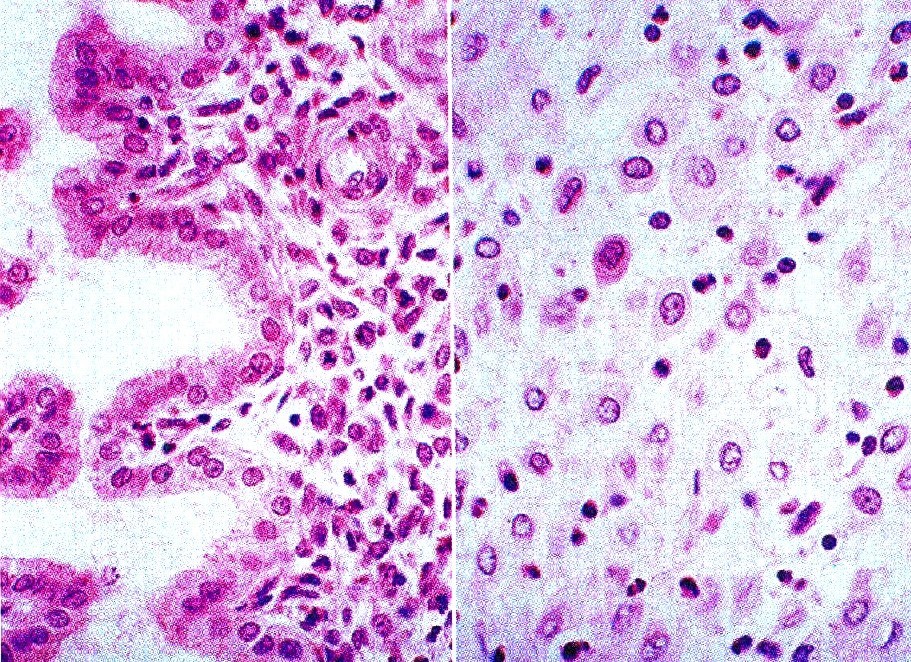
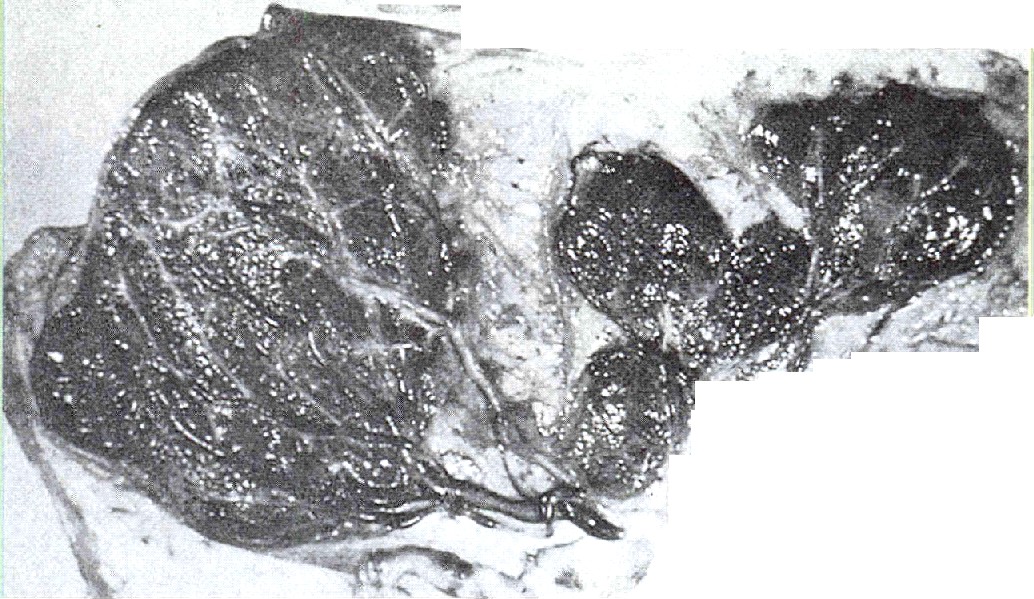
La mola hidatidiforme es una afección no invasiva en la cual muchas de las vellosidades coriónicas se caracterizan por tumefacciones nodulares, que les dan una apariencia similar a racimos de uvas. Por lo común, gran parte de la superficie de la placenta adopta este aspecto; además, el embrión está ausente o es inviable (figura 6-18). Las vellosidades no muestran signos de vascularización.
El análisis genético ha demostrado que las molas hidatidiformes representan los resultados de la impronta patema en la cual el pronúcleo femenino del ovulo no participa en el desarrollo (capítulo 3). En vez de ello, el material cromosómico se deriva de dos espermatozoides que han penetrado en el óvulo, o de la duplicación de un pronúcleo espermático único en el interior del óvulo. Los cromosomas de las molas hidatidiformes son derivados paternos, 46 XX, ya que el número de genes letales en los embriones 46 YY es incompatible con su supervivencia.
CORRELACiÓN CLÍNICA 6-3
Afecciones placentarias (continuación)
Los coriocarcinomas son tumores malignos derivados del citotrofoblasto y el sincitiotrofoblasto embrionarios. Estos tumores son muy invasivos de los tejidos deciduales y de los vasos maternos. A semejanza de la mola hidatidiforme, la mayoría de los coriocarcinomas sólo contiene cromosomas derivados del padre y por tanto son producto de la impronta paterna.
BIOPSIA DE LA VELLOSIDAD CORIÓNICA
Recientemente se ha empezado a practicar las biopsias de ia vellosidad coriónica en la segunda mitad del segundo mes de vida embrionaria, en lugar de la obtención de la muestra de líquido amniótico. Este procedimiento se realiza bajo guía ecográfica y se obtienen biopsias de vellosidades coriónicas para el análisis de posibles anomalías cromosómicas o para el diagnóstico de ciertos problemas metabólicos.
En los gemelos fusionados y rara vez en los gemelos dicigóticos con mínima separación de la masa celular interna, los embriones se desarrollan en el interior de un anmios y un corion único y tienen una placenta común con una irrigación sanguínea también común (figura 6-15 D Y E). Esta situación, y las ya descritas, pueden determinarse con facilidad al examinar las membranas de la placenta. En una época se creía que podía determinarse si los gemelos eran monocigóticos o dicigóticos con sólo examinar las membranas. Aunque en la mayoría de los casos es posible hacer la deducción correcta, este método no es infalible. Para hacer el diagnóstico definitivo deben usarse otros métodos que van desde la observación simple del sexo, el color de los ojos y los patrones de las huellas digitales hasta la determinación del tipo sanguíneo o incluso de la composición del ácido desoxirribonucleico (DNA). En la actualidad, con los trasplantes de células y de órganos puede ser vital saber si los gemelos son monocigóticos en el caso de que uno de ellos desarrolle una afección que pueda tratarse con un trasplante.
CASO CLÍNICO
El obstetra de una mujer de 32 años de edad observa que su aumento de peso hacia el final del embarazo es excesivo. Al menos parte de dicho aumento parece ser el resultado de un volumen de líquido amniótico mayor del normal. Ella reside en una remota área rural lejos de un centro de diagnóstico ecográfico. Se le hace una amniocentesis y el laboratorio informa acerca de la presencia de un nivel elevado de alfa-fetoproteína en el líquido amniótico. El médico está preocupado de que este embarazo no tenga como resultado un parto normal.
¿Qué afección sospecha el médico y por qué?
A. Atresia esofágica
B. Agenesia renal
C. Trillizos
D. Anencefalia
E. Placenta previa.
. Las membranas extraembrionarias comprenden el corion (la combinación del trofoblasto más el mesodermo extraembrionario subyacente), el amnios, el saco vitelino y la alantoides.
. El amnios, una delgada membrana ectodérmica tapizada con mesodermo, crece hasta encerrar al embrión a la manera de un globo. Está lleno de un líquido transparente que es generado en muchas partes como la piel del feto, el mismo amnios, los riñones fetales y posiblemente los vasos fetales. Al término del embarazo el volumen del líquido amniótico se aproxima a 1 litro. El líquido amniótico se elimina mediante intercambio a través de la membrana amniótica y deglución fetal.
. El saco vitelino es una estructura de situación ventral tapizada por endodermo y que no tiene una función nutritiva en los embriones de los mamíferos. Los islotes sanguíneos mesodérmicos en la pared del saco vitelino forman las primeras células sanguíneas y los primeros vasos. Las células germinales primordiales son reconocibles en la pared del saco vitelino, pero se originan en el mesodermo extraembrionario cerca de la base de la alantoides.
. La alantoides es un pequeño divertículo tapizado de endodermo, situado en el lado ventral del intestino posterior en los seres humanos. No tiene una función directa de respiración ni de almacenamiento de desechos. Estas funciones las cumplen la placenta y los vasos umbilicales que surgen al mismo tiempo que la alantoides.
. Las vellosidades coriónicas se forman como evaginaciones externas del trofoblasto. Las vellosidades primarias son proyecciones de solo trofoblasto. Cuando se forma una zona mesenquimatosa en el interior de una vellosidad, se convierte en veIlosidad secundaria y cuando el mesénquima se vasculariza, la vellosidad es terciaria. Conforme maduran las vellosidades, el citotrofoblasto de algunas de ellas crece a través del sincitiotrofoblasto en forma de columnas de células citotrofoblásticas y establece contacto con el tejido endometrial materno. El citotrofoblasto continúa creciendo alrededor del espacio lleno de sangre que rodea al corion hasta formar una cápsula citotrofoblástica, que es la interfase directa entre los tejidos matemos y fetales. Las vellosidades que hacen contacto directo con los tejidos matemos se llaman de anclaje; las que no lo hacen se llaman flotantes. Puesto que las vellosidades coriónicas flotan en un lago de sangre materna, la placenta humana se clasifica como placenta hemocorial.
. Estimuladas por el embrión al implantarse, las células del estroma endometrial experimentan la reacción decidual. Los tejidos matemos que se pierden en el parto reciben el nombre global de decidua. La decidua basal subyace a la placenta; la capsular, engloba al resto del corion como una cápsula; las partes de la pared uterina que no están ocupadas por el corion fetal constituyen la decidua parietal. Conforme el corion fetal madura, se subdivide en un corion leve, en el cual las vellosidades sufren regresión, y el corion frondoso, que es la región del corion mas próxima a los tejidos basales del endometrio. El corion frondoso dará lugar a la placenta.
. La placenta madura consta de la pared del corion (la placa coriónica) y numerosas vellosidades que se proyectan desde ella. La superficie fetal de la placenta es lisa y brillante a causa de la aposición de la membrana amniótica. La superficie materna es mate y lobulada, con cotiledones de numerosas vellosidades placentarias y sus ramificaciones. El cordón umbilical (que en un principio fue el tallo de conexión) se inserta en el centro de la placenta. La sangre procedente del feto llega a la placenta a través de las arterias umbilicales. Éstas se ramifican en numerosos vasos pequeños, que terminan en asas capilares en los extremos de las vellosidades placentarias. Allí se intercambian el oxígeno, los nutrientes y los desechos entre la sangre materna y la fetal, que baña a las vellosidades. La sangre fetal regresa al cuerpo del feto maduro a través de una vena umbilical única. La sangre materna que sale por los extremos abiertos de las arterias espirales del endometrio baña las vellosidades placentarias.
. La transferencia de sustancias desde la sangre fetal a la materna tiene que discurrir a través del endotelio de los capilares fetales, la lámina basal y los tejidos trofoblásticos antes de llegar a la sangre materna. La transferencia de sustancias se logra en virtud de mecanismos activos y pasivos. Además de las sustancias normales, el alcohol, ciertos medicamentos y algunos
agentes infecciosos pueden pasar desde la sangre materna hasta la circulación fetal y entorpecer el desarrollo normal. Si un feto es Rh-positivo y la madre es Rh-negativa, los anticuerpos anti-Rh maternos procedentes de un embarazo anterior pueden pasar al feto y causarle una eritroblastosis fetal o enfermedad hemolítica.
. La placenta produce una gran variedad de hormonas, muchas de las cuales suelen sintetizarse en el hipotálamo y la adenohipófisis. La primera hormona en liberarse es la HCG, que sirve de base a muchas de las pruebas de embarazo. Otras hormonas placentarias son la somatomamotropina coriónica (lactógeno
placentario humano), las hormonas esteroideas y la tirotropina y corticotropina coriónicas.
Los tejidos fetales y placentarios son diferentes de los de la madre desde el punto de vista inmunológico, pero el sistema inmunológico de la madre no rechaza a la placenta y al feto. Las razones aún no están del todo claras, pero algunas explicaciones hacen referencia a la reducida antigenicidad de los tejidos del trofoblasto, el bloqueo del sistema inmunológico materno durante el embarazo y a barreras inmunológicas locales entre el feto y la madre.
La placenta con membranas fetales se expulsa unos 30 minutos después del feto. La inspección de la placenta puede revelar anomalías placentarias, falta de cotiledones o la disposición de las membranas en los embarazos múltiples. Este último dato puede ayudar a determinar si el origen de un embarazo múltiple es o no monocigótico. Los hallazgos patológicos de la placenta incluyen las anomalías morfológicas macroscópicas, la mola hidatidiforrne benigna y los coriocarcinomas malignos.
PREGUNTAS DE REPASO
1. En la placenta madura, ¿qué tejido fetal está en relación directa con el tejido conectivo uterino materno?
A. El citotrofoblasto
B. El sincitiotrofoblasto
C. El mesodermo extraembrionario
D. Las células deciduales
E. Nil29una de las anteriores
2. ¿Que afección se relaciona con la impronta paterna?
A. Lóbulos placentarios accesorios
B. Placenta previa
C. Oligohidramnios
D. Arteria umbilical única
E. Mola hidatidiforme
3. ¿Los vasos sanguíneos con qué estructura entran en el componente fetal de la placenta?
A. Decidua basal
B. Alantoides
C. Amnios
D. Saco vitelino
E. Decidua parietal
4. ¿Qué tipo de células invaden las arterias espirales maternas y reduce el flujo de sangre a través de sus extremos abiertos?
A. Células de Hoffbauer
B. Sincitiotrofoblasto
C. Eritrocitos fetales
D. Citotrofoblasto
E. Epitelio amniótico
5. ¿Qué disposición de las membranas extraembrionarias puede encontrarse en el útero que contiene gemelos idénticos?
A. Placenta y membrana amniótica comunes
B. Placenta y corion comunes, amnios separados
C; PIFlr.AntFls V mAmhmnFlsAxtmAmhrinnFlriFls,sAORrFldFlSc
D. Placenta común, corion parcialmente fusionado
E. Todas las anteriores
6. El segundo hijo de una mujer Rh-negativa de 28 años nace con una grave ictericia. ¿Cuál de las siguientes afirmaciones caracteriza con mayor exactitud a su primer hijo?
A. Varón
B. Mujer
C. Rh-positivo
D. Rh-negativo
E. Hidramnios
7. ¿Por qué se llama a la placenta humana placenta de tipo hemocorial?
8. ¿A través de qué capas de una vellosidad placentaria debe pasar una molécula de oxígeno para ir desde la sangre materna hasta la circulación embrionaria?
9. ¿Cuál hormona embrionaria ha servido de base a muchas pruebas estándar de embarazo y por qué?
10. ¿Por qué la mujer embarazada tiene que tener mucho cuidado con lo que bebe o come?
Ahmed MS, Cemerikic B, AgLas A: Properties and functions of hu
man placental opioid system, Life Sci 50:83-97, 1991.
AIsat E et al.: Human placental growth hormone, Am J Obstet Gyne
col 177:1526-1534, 1997.
Aplin JD: Implantation, trophoblast differentiation and haemochorial
placentation mechanistic evidence in vivo and in vitro, J Cell Sci
99:681-692, 1991.
Benirschke K, Kaufmann P: Pathology of the human placenta, 2a. ed.,
Nueva Yórk; 1990; spririger- Vérlág.
Billingham RE, Beer AE: Reproductive immunology: past, present
and future, Prospect Biol Med 27:259-275, 1984.
Bohn H, Dati F, Lueben G: Human trophoblast specific products other
than hormones. En Loke YW, Whyte A, eds.: Biology of tropho
blast, Amsterdam, 1983, EIsevier Science, págs. 317-352.
Bohn H, Winckler W, Grundmann U: Immunochemically detected pla
cental proteins and their biological functions, Arch Gynecol Obstet
249:1O7-118,199l.
Boyd ID, Hamilton WJ: The human placenta, Cambridge, Inglaterra,
1970, Heffer & Sons.
Chamberlain GVP, Wilkinson AW, eds.: Placen tal transjer Tunbridge
Wells, Kent, Inglaterra, 1979, Pitman Medical.
Cullen TS: Embryology, anatomy and diseases of the umbilicus, to
gether with diseases of the urachus, Filadelfia, 1916, WB Saunders.
Dallaire L, Potier M: Amniotic fluido En Milunsky A, ed: Genetic disorders and thefetus, Nueva York, 1986, Plenum, págs. 53-97.
Dearden L, Ockleford CD: Structure ofhuman trophoblast correlation
with function. En Loke YW, Whyte A, eds.: Biology of trophoblast,
Amsterdam, 1983, EIsevier Science, págs. 69-110.
Demir R et al.: Classification of human placental stem villi review of
structural and functional aspects, Micros Res Techn 38:29-41,
1997.
Enders AC: Trophoblast differentiation during the transition from trop
hoblastic plate to lacunar stage of implantation in the rhesus mon
key and human, Am J Anat 186:85-98, 1989.
IOn i>"""U :iIJ\-."rnhirouJof;T >-v>P,'<:;QcP.! -T'lm!-whufopriyJ lfflU~Y,' ''IUCM--dnK) 99fh, Raven.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}