Capítulo 15. SISTEMA UROGENITAL
Primeras formas del riñón
Metanefros
Cambios posteriores en el desarrollo del riñón
Determinación genética del sexo
CORRELACION CLINICA 15-1
AGENESIA RENAL
CORRELACION CLINICA 15-1b
HIPOPLASIA RENAL
DUPLICACIONES RENALES
ANOMALIAS DE LA MIGRACION Y LA ROTACION RENAL
CORRELACION CLINICA 15-1c
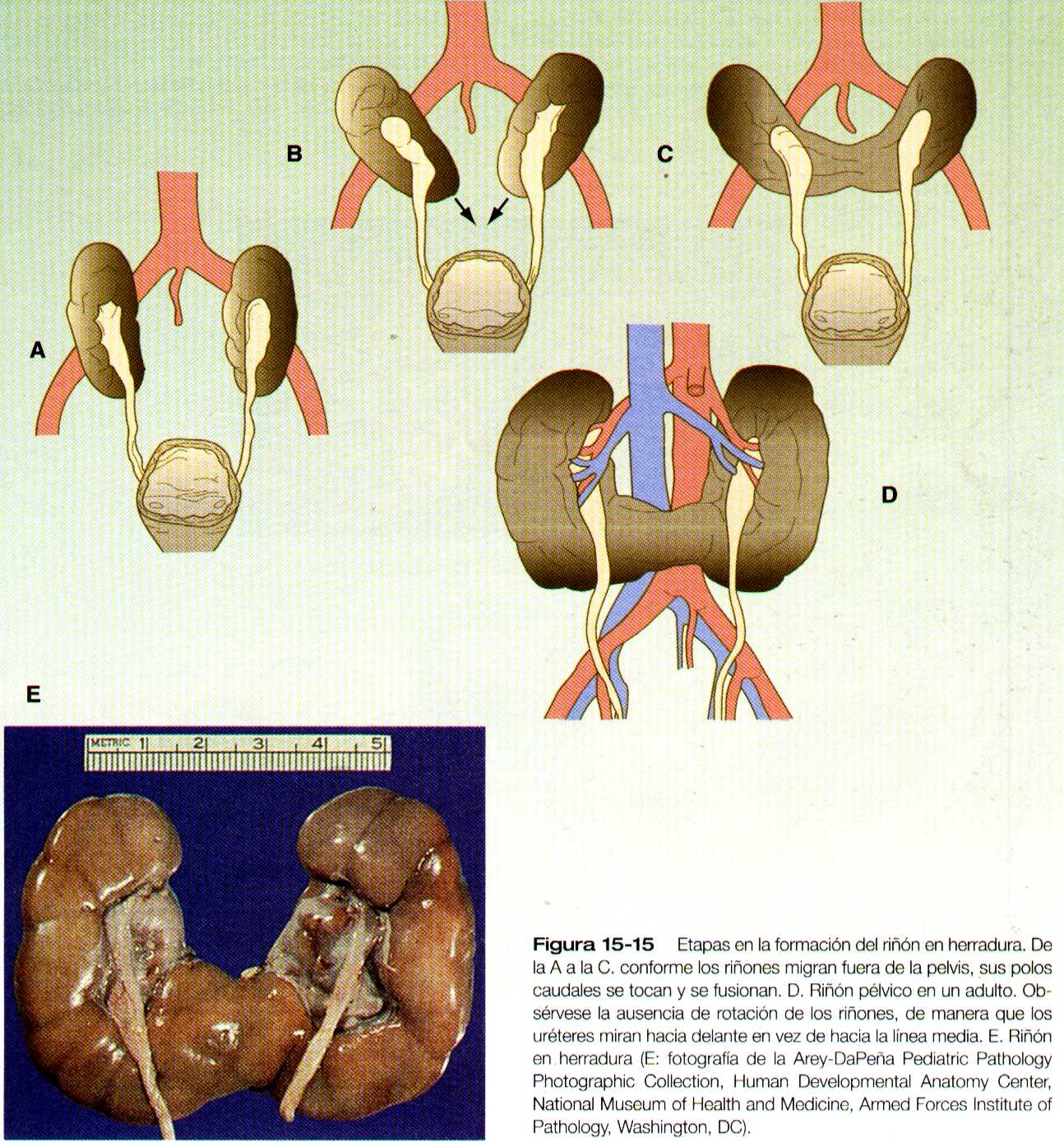
Riñón en herradura
CORRELACION CLINICA 15-1d
QUISTES, SENOS Y FÍSTULAS DEL URACO
Migración de las células germinales hacia las gónadas
Establecimiento del sexo gonadal
Diferenciación de los testículos
Fase indiferenciada del sistema de conductos sexuales
Sistema de conductos sexuales masculinos
Sistema de conductos sexuales femeninos
Descenso de las gónadas
Etapa indiferenciada
CORRELACION CLINICA 15-2a
Malformaciones del sistema genital
ANOMALIAS DE LA DIFERENCIACION SEXUAL
Síndrome de Turner (disgenesia gonadal)
Seudohermafroditismo masculino
Síndrome de feminización testicular (insensibilidad a los andrógenos)
ESTRUCTURAS VESTIGIALES DE LOS CONDUCTOS GENITALES EMBRIONARIOS
Restos de los conductos mesonéfricos
Restos del conducto paramesonéfrico
OTRAS ANOMALIAS DEL SISTEMA DE CONDUCTOS GENITALES
CORRELACION CLINICA 15-2b
Malformaciones del sistema genital (continuación)
ANOMALÍAS DEL DESCENSO TESTICULAR
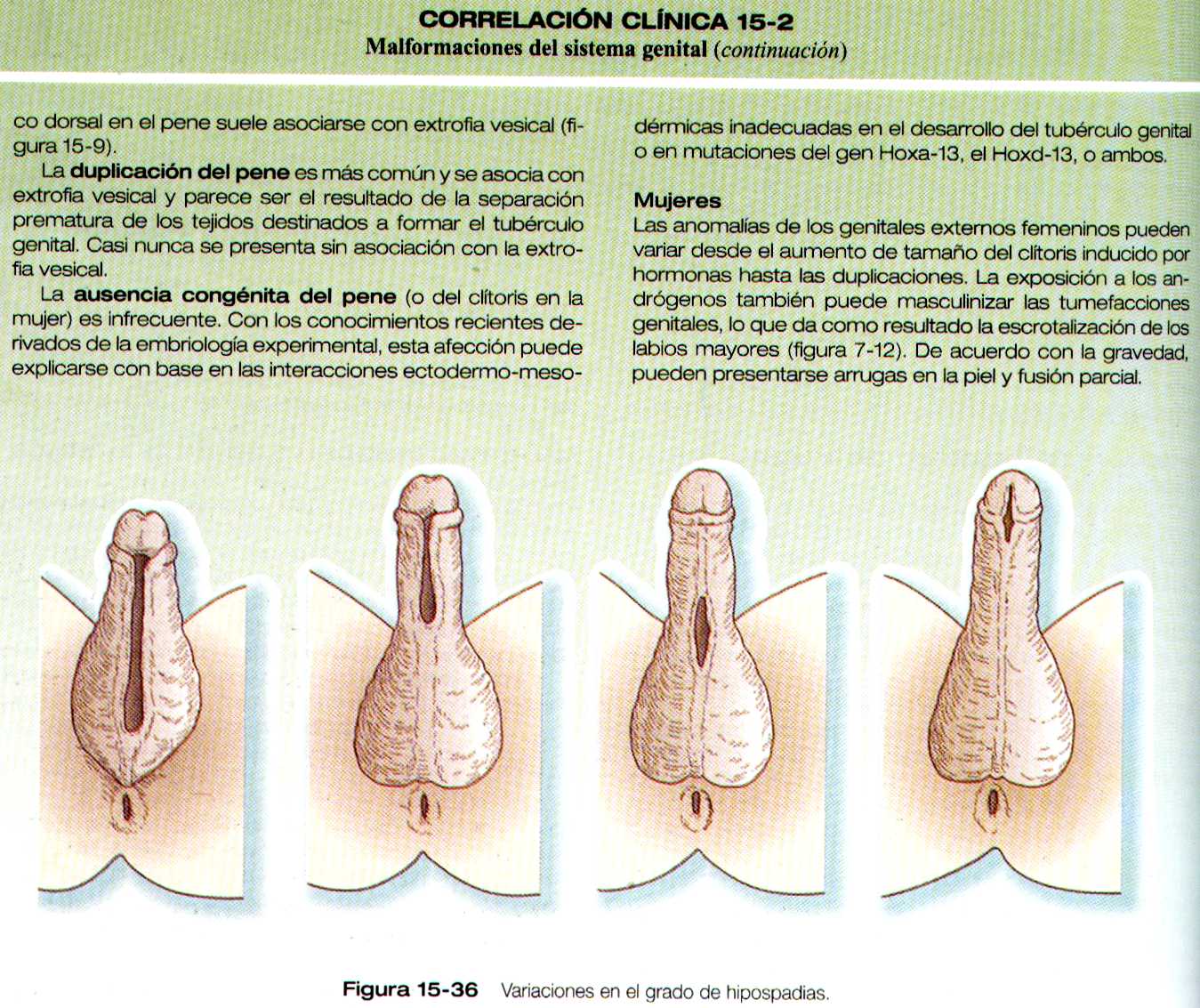
MALFORMACIONES DE LOS GENITALES EXTERNOS
Varones
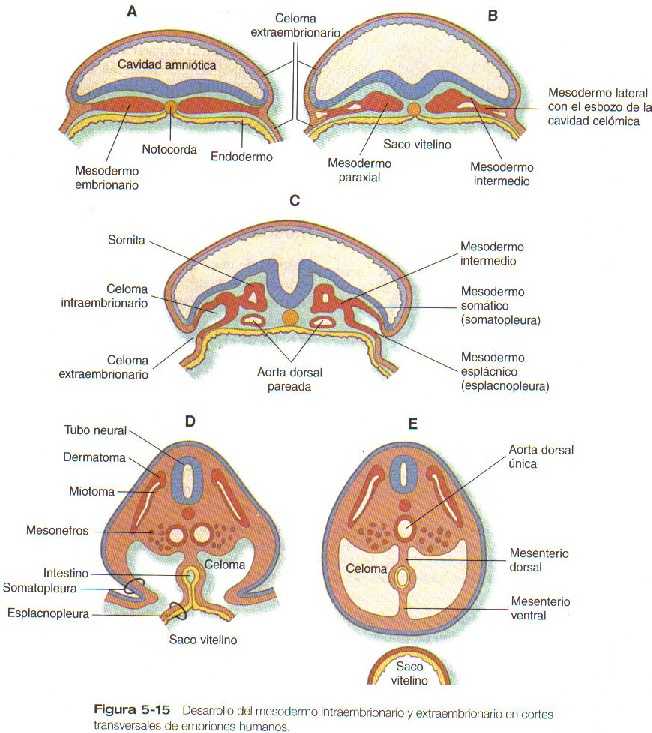
El sistema urogenital se origina a partir del mesodermo intermedio del embrión inicial (figura 5-15); varios temas se interrelacionan en el desarrollo de las estructuras urinarias y genitales a partir de este precursor común. El primero es la interconexión del desarrollo urinario con el genital, según la cual los componentes primarios de un sistema son sustituidos por el otro hacia el final del desarrollo. Un segundo tema es la recapitulación que se hace durante la ontogenia humana de los tipos renales (el equivalente de las isoformas del órgano) que son las formaciones renales definitivas de los vertebrados inferiores. Un tercer tema es la dependencia de la diferenciación y el mantenimiento de muchas estructuras del sistema urogenital por interacciones epitelio-mesénquima. Por ultimo, la diferenciación sexual de muchas estructuras pasa desde un estado indiferenciado, en el cual el macho y la hembra no se distinguen a simple vista, una vía masculina o femenina va a depender de la presencia de factores promotores o inhibidores específicos que actúen sobre la estructura. Aunque el sexo fenotípico se determina genéticamente, el genético puede ser modificado por factores ambientales, lo que puede llevar a una discordancia entre los dos. Las correlaciones clínicas 15-1 y 15-2 examinan anomalías de los sistemas urinario y genital, respectivamente.
SISTEMA URINARIO
El sistema urinario comienza a formarse antes de que se haga evidente cualquier desarrollo gonadal. La primera etapa en la embriogenesis del riñón es la formación de un par de órganos excretores similares en estructura y función a los riñones de los vertebrados inferiores. Estas formas incipientes de riñón mas tarde son remplazados por los riñones metanéfricos definitivos, pero conforme hacen su regresión, se mantiene algunos componentes que serán reutilizados por otras estructuras del sistema urogenital.
Primeras formas del riñón
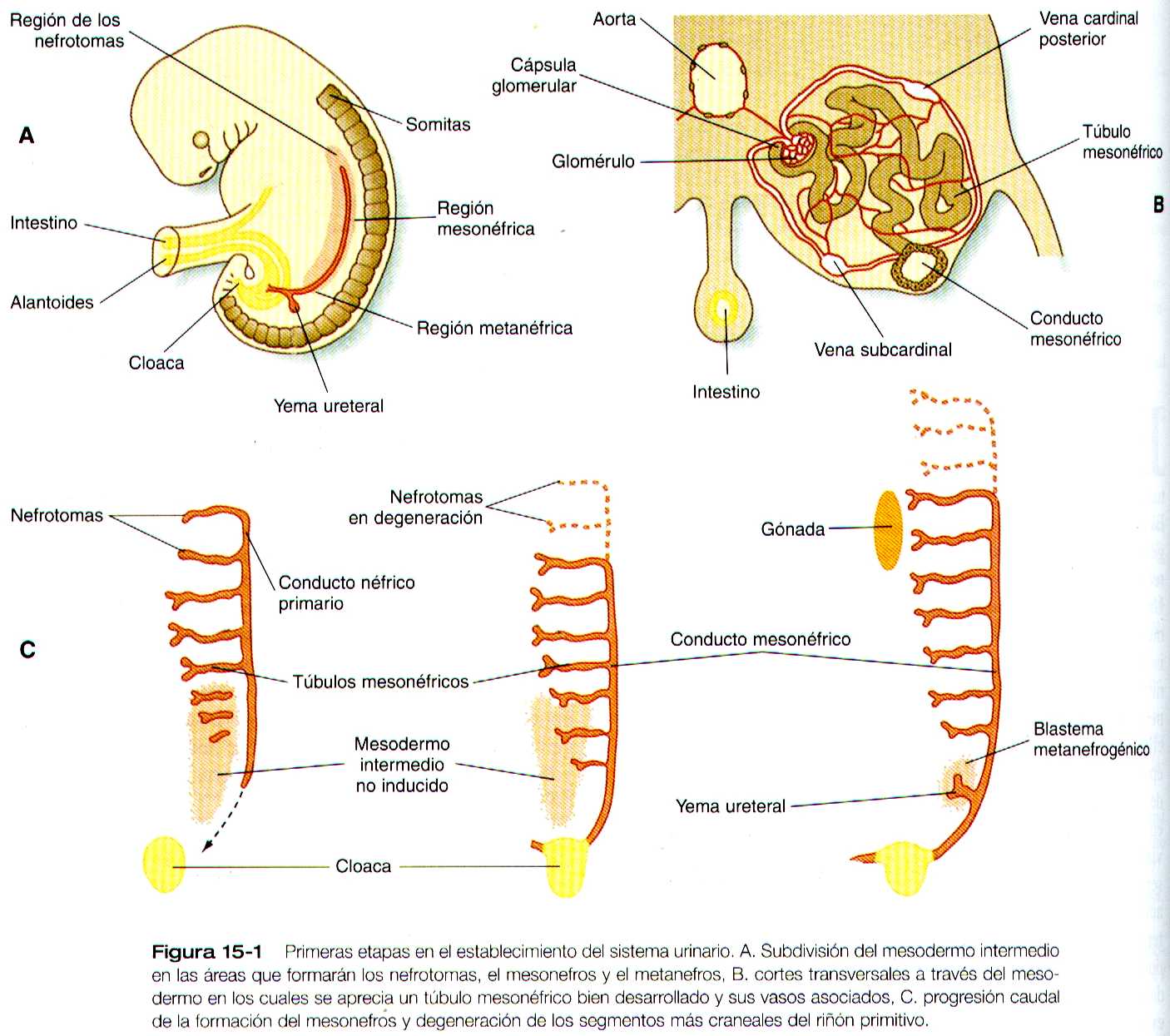
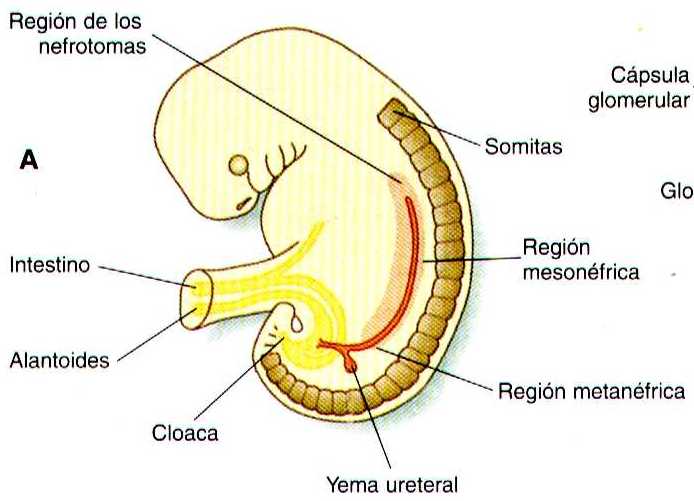
La representación común del desarrollo del riñón en los mamíferos comprende tres fases sucesivas que comienzan con la aparición del pronefros, el homologo del tipo de riñón que solo se encuentra en los vertebrados inferiores. En los embriones humanos la primera evidencia de un sistema urinario consiste en la aparición de unos pocos grupos de cordones epiteliales organizados de manera segmentaría que se diferencian a partir del mesodermo intermedio superior hacia los 22 días de gestación. Estas estructuras reciben el nombre mas apropiado de nefrotomas, las cuales se conectan hacia los lados con un par de conductos néfricos primarios (pronéfricos) que crecen hacia la cloaca (figura 15-1). La base molecular para la iniciación del desarrollo de los riñones aun no es clara, pero los experimentos de bloqueo de genes han sugerido que los genes de homeodominio Lim-1 y Pax-2 son importantes en las primeras etapas del desarrollo renal.
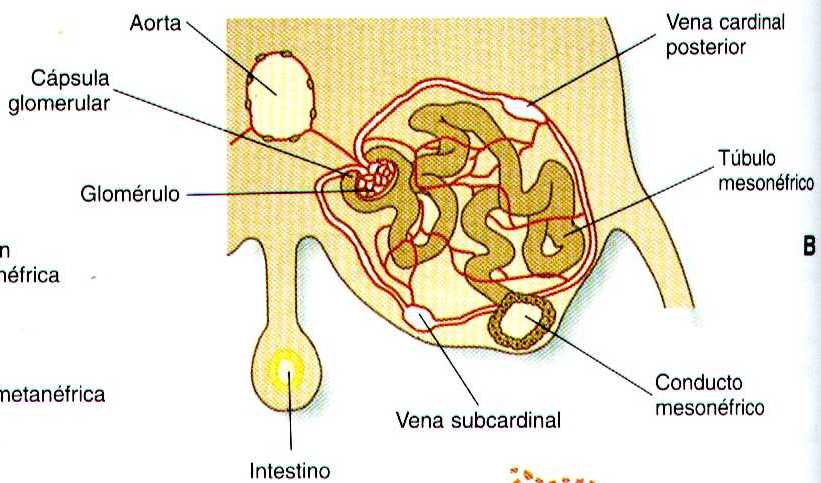
Conforme los conductos néfricos primarios se extienden en dirección caudal, estimulan al mesodermo intermedio PAA que forme grupos segmentarios adicionales de tubos. Estos tubos son los equivalentes estructurales de los túbulos mesonéfricos de los peces y los anfibios. Una unidad mesonefrica típica consta de un glomérulo vascular parcialmente rodeado por una cápsula epitelial, que se continua con un túbulo concentrado mesonéfrico. Este a su vez, esta rodeado por una red de capilares (figura 15-1 B). Cada túbulo mesonéfrico desemboca por separado en la continuación del conducto néfrico primario, que recibe entonces el nombre de conducto mesonéfrico (de Wolff).
La formación de pares de túbulos mesonéfricos se produce a lo largo de un gigante cráneo caudal. Aparecer los primeros cuatro a seis pares de túbulos mesonéfricos (así como los túbulos pronéfricos) surgen como excrecencias de los conductos néfricos primarios. En situación mas caudal, los túbulos mesonéfricos se forman individualmente en el mesodermo intermedio un poco por detrás de la extensión caudal de los conductos mesonéfricos. Hacia el final de la cuarta semana de gestación los conductos mesonéfricos se conecten con la cloaca, y hay una luz continua a lo largo de cada uno de ellos. Existe una diferencia entre los controles del desarrollo entre los cuatro a seis pares mas caudales de conductos mesonéfricos y los restantes caudales.
Los experimentos de bloqueo del gen WT-1 (supresor del tumor de Wilms) dan como resultado la ausencia de los túbulos mesonéfricos caudales, en tanto que los craneales que se desprenden del conducto pronéfrico se desarrollan normalmente. Al igual que en la formación del matanefros (véase mas adelante ), el WT-1 parece regular la transformación del mesénquima en epitelio durante el comienzo de la formación de los túbulos renales (mesonéfricos ). Muy cerca de su lugar de inserción en la cloaca, en el conducto mesonéfrico se desarrolla un excrecencia epitelial llamada yema ureteral (figura 15-1 A). Al principio de la quinta semana la yema ureteral comienza a crecer hacia la región mas caudal del mesodermo intermedio. A continuación activa una serie de interacciones inductivas continuas que llevan a la formación del riñón definitivo., el metanefros.
Aunque existen evidencias de función urinaria en el riñón mesonéfrico de los mamíferos, la fisiología del mesonefros no se han investigado en profundidad. La producción de orina en esta estructura comienza con un filtrado de sangre del glomérulo hacia la cápsula glomerular, este fluye después hacia la porción tubular del mesonefros, donde tiene lugar la reabsorción selectiva de los iones y otras sustancias . El regreso de los materiales reabsorbidos a la sangre se ve facilitado por la presencia de un plexo denso de capitales alrededor de los túbulos mesonéfricos.
La estructura del mesonefros del embrión humano es muy similar a la de los peces y los anfibios acuáticos adultos, y su función principal es filtrar y eliminar los desechos corporales. Puesto que estas especies y el embrión amniótico existen en un medio acuático, hay muy poca necesidad de conservar agua. En consecuencia, en el mesonefros no se desarrolla una región medular ni un sistema elaborado de concentración de la orina como si tiene que hacerlo el riñón humano adulto.
Los glomerulos mesonéfricos son mas voluminosos cuando el metanefros se esta formando. Si bien su regresión como unidad urinaria es rápida después de que los riñones metanéfricos se vuelven funcionales, los conductos mesonéfricos y algunos de los tubulos mesonéfricos persisten en el varón y acaban por incorporarse formando parte de las estructuras del sistema de conductos genitales.
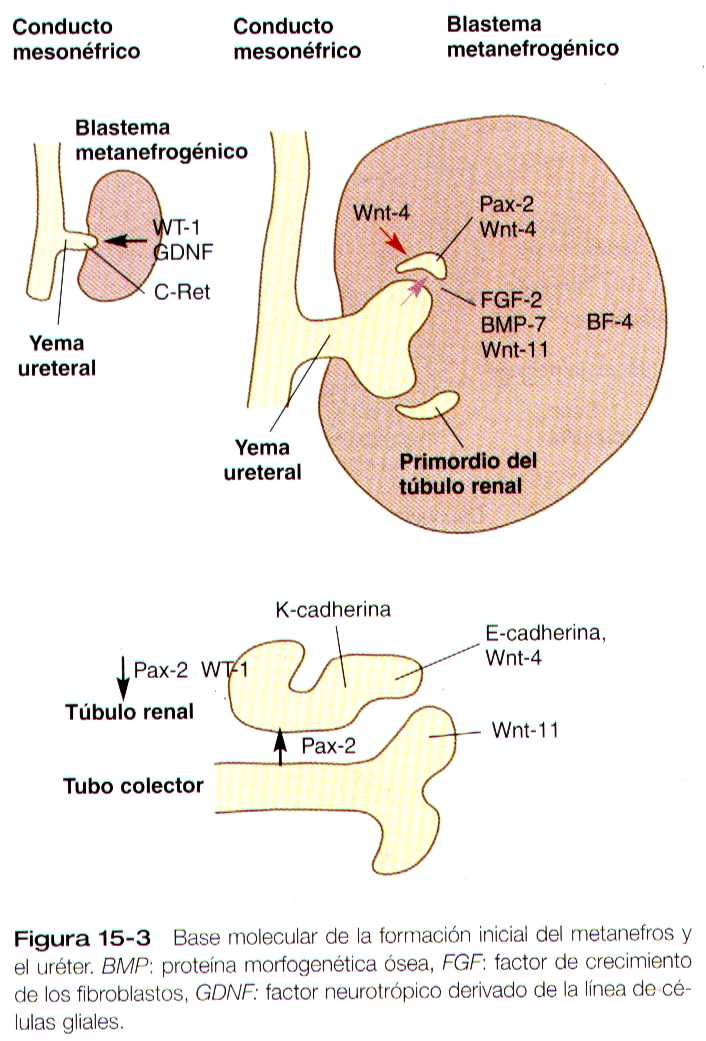
El desarrollo del metanefros se inicia al comienzo de la quinta semana, cuando la yema ureteral (divertículo metanéfrico) crece hacia la porción posterior del mesodermo intermedio. Las células mesenquimatosas del mesodermo intermedio se condensan alrededor del divertículo para formar el blastema metanefrogénico. El crecimiento de la yema ureteral a partir del conducto mesonéfrico es una respuesta a la secreción del factor neurotrópico derivado de la línea de las células gliales (GDNF) por parte de mesénquima diferenciado del blastema metanefrogénico. Esta señal inductora esta ligada al c-Ret, miembro de la super familia de receptores de la tirosina quinasa localizado en las membranas plasmáticas de las células epiteliales de la incipiente yema ureteral. La formación de GNDF del mesénquima metanéfrico es regulada por el WT-1.
La base molecular del desarrollo del riñón metanéfrico es el crecimiento y la ramificación (hasta a catorce a quince veces) de la yema ureteral, que se convierte en el sistema de conductos colectores (metanéfricos) del mesonefros, y la formación de los túbulos renales a partir de las condensaciones de mesénquima (blastema metanefrogénico) localizadas alrededor de las puntas de las ramificaciones ureterales
El mecanismo que subyace a estos sucesos es una serie de interacciones inductivas recíprocas entre las puntas de las ramificaciones del los conductos metanéfricos y las células del blastema metanéfrico que las rodea; sin el sistema de conductos metanéfricos, no se forman los túbulos. A la inversa el mesodermo metanefrogénico actúan sobre este sistema, en el cual inducen la ramificación característica. La respuesta a la señal inductiva del GDNF por parte del mesénquima metanéfrico, las células epiteliales de la yema ureteral produce factor de crecimiento de los fibroblastos (FGF-2), proteína mofo genética ósea (BMP-7) y WNt-11, que inducen al mesénquima metanéfrico circundante para que comience a formar los precursores epiteliales de los túbulos renales. El desarrollo de los túbulos también requiere una señal inductiva secuencial, el Wnt-11 que es producida por el mesénquima metanéfrico mismo. Esta inducción precoz moldea el mesénquima metanéfrico para que se convierta en un dominio epitelial tubular, en la cual las células expresan Wnt-4 y Pax-2, una región estromal, en la cual las células mesenquimatosas expresan un factor de trascripción de winged helix, BF-2.
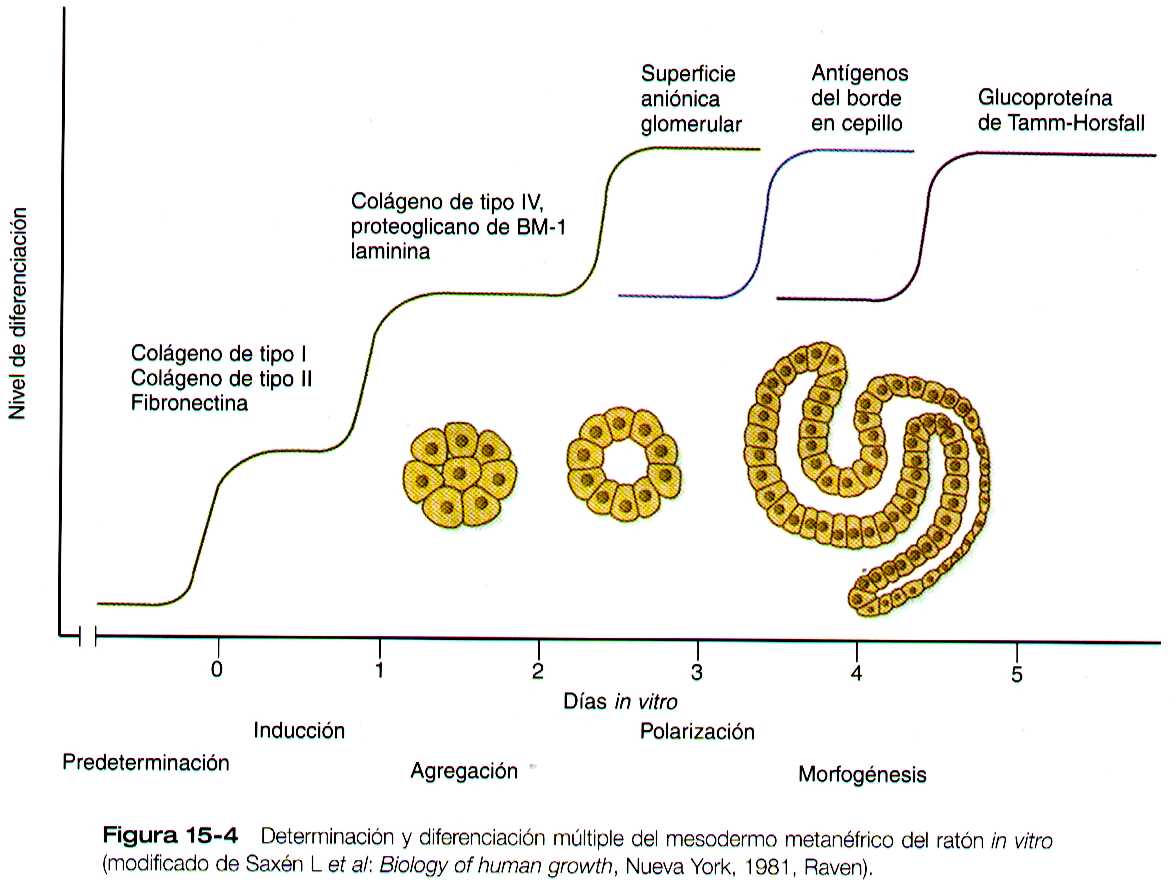
La formación de los túbulos individuales funcionales (nefronas) en el metanefros el desarrollo comprende tres linajes de células mesodérmicas: células epiteliales derivadas de la yema ureteral, células mesenquimatosas del blastema metanefrogénico y células endoteliales vasculares. La etapa mas temprana es la condensación de las células del blastema mesenquimatoso alrededor del brote terminando la yema ureteral ( que mas delante se convertirá en el conducto metanéfrico). El mesénquima preinducido contiene varias proteínas interticiales, como los colágenos de tipo I y III y fibronectina. Conforme las células mesenquimatosas se condensan después de la inducción local por parte de las puntas de ramificación de la yema ureteral, estas proteínas se pierden y son remplazadas por proteínas de tipo epitelial (colágeno tipo IV, sindecan t-1, laminina y el proteoglicano heparan sulfato), los cuales terminan por localizarse en las membranas básales. (figura 15-4)
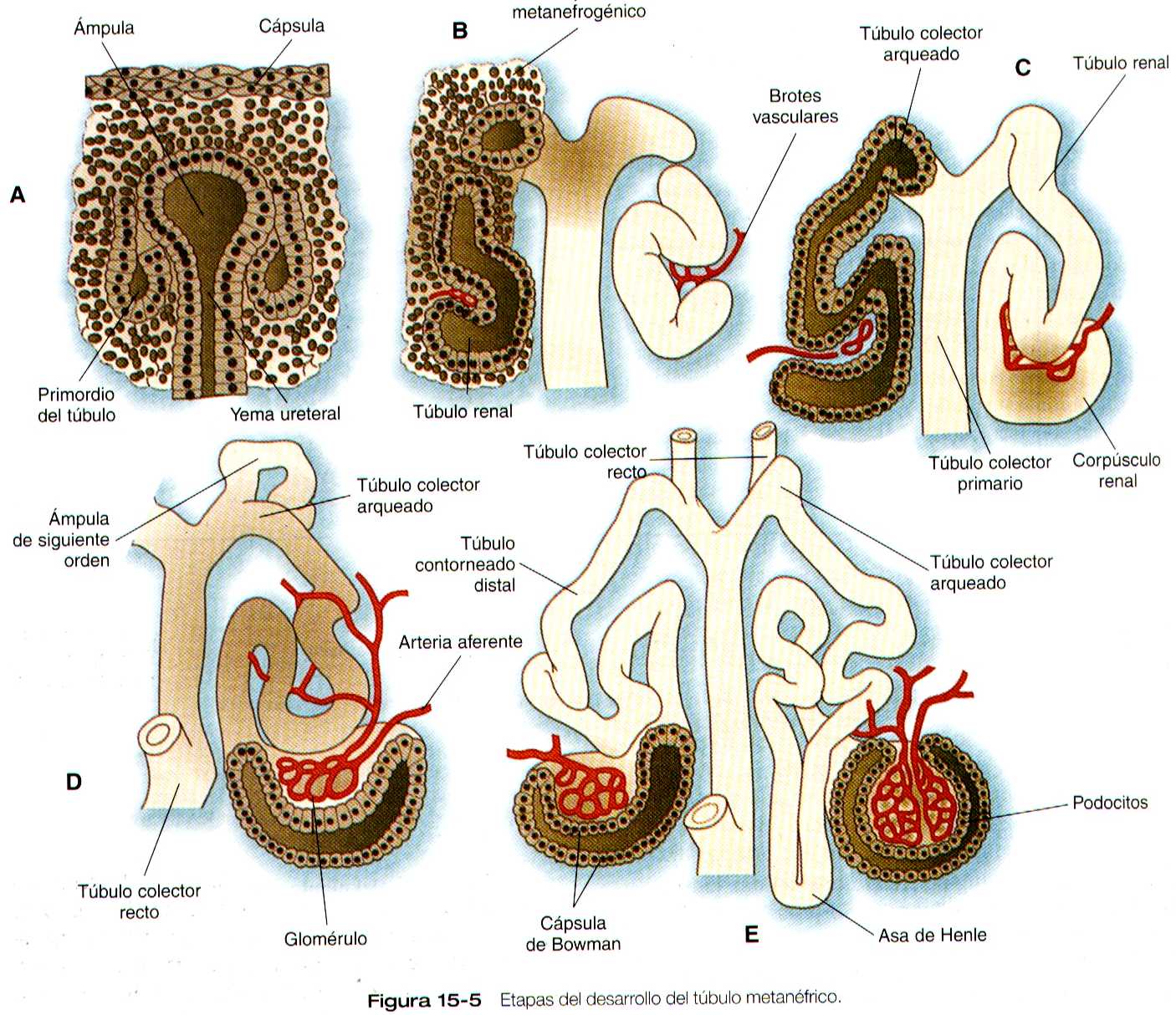
Conforme la yema terminal del conducto metanéfrico se ramifica, el mesénquima que la rodea se divide en dos partes. Una simple condensación y las células mesenquimatoas pasan por una serie de etapas hasta formar un túbulo renal, después de una fase de crecimiento, la actividad mitótica del interior del mesénquima del blastema disminuye, y el primordio del túbulo adopta la forma de una coma (figura 15-5 A). En ella, un grupo de células que están mas lejanas del extremo del conducto metanéfrico se polarizan, y forman una luz central con una lámina basal en la otra superficie. Esto indica la inducción de la transformación de las células mesenquimatosas hacia un epitelio especializado, específicamente de los podocitos, que al final rodean a endotelio vascular del glomérulo.
Una consecuencia de esta transformación epitelial en el primordio del túbulo es la formación de una hendidura por debajo de los precursores de los podocitos en desarrollo (figura 15-5 B). Los precursores de las células endoteliales vasculares crecen en esta hendidura, que al final se convierten en el glomérulo. La investigación con el uso de híbridos ínter específicos ha demostrado que el endotelio vascular migra desde fuera de la región del mesodermo intermedio hacia el metanefros en desarrollo. El mesénquima metanéfrico inducido estimula el crecimiento de las células endoteliales, posiblemente gracias a la liberación de un factor similar al FGF. El mesénquima no inducido no posee esta capacidad. Las células endoteliales están conectadas con ramas de la aorta dorsal, y dan lugar a una compleja estructura que forman asas y que al final se convertirá en el glomérulo renal. Las células del endotelio glomerular y el vecino epitelio de podocitos forman una gruesa membrana basal entre ellas, que mas tarde será un importante componente del aparato de la filtración renal.
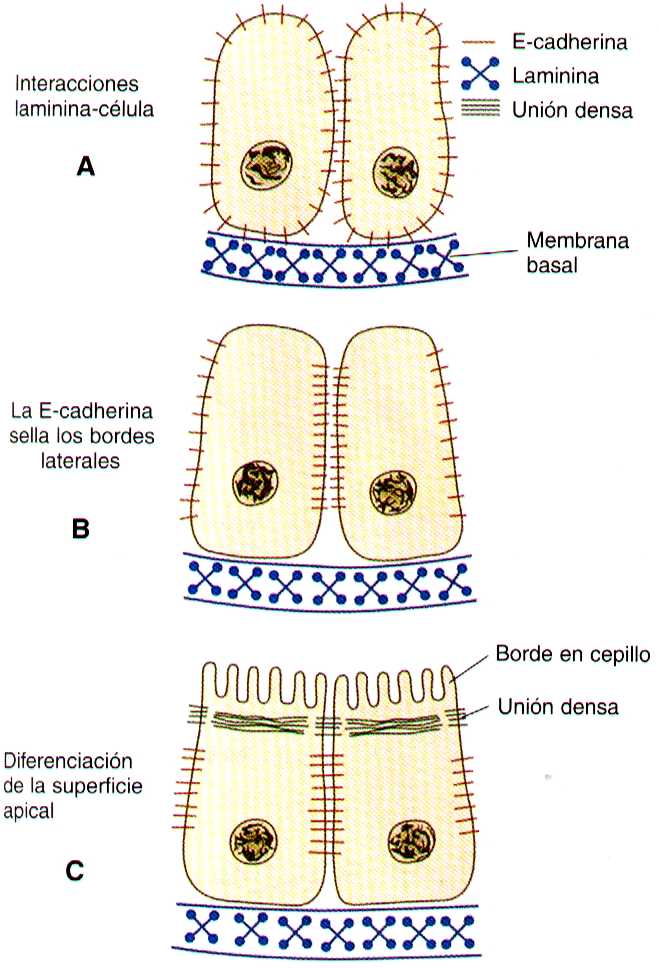
Conforme el aparato glomerular de la neurona toma forma, se desarrolla otra hendidura en el primordio tubular a manera de coma, así su aspecto cambia al de una S (figura 15-5 C). Las células del resto del túbulo primitivo también experimentan una transformación epitelial para formar el resto del túbulo renal. Esta transformación implica que al diferenciarse las células epiteliales adquieran polaridad. Este fenómeno se correlaciona con el depósito de láminina en la matriz extracelular a lo largo de la superficie basal de las células y con la concentración de la glucoproteina integral de la membrana uvomorulina (E- cadherina), que sella los bordes laterales de estas células (figura 15-6). Conforme el túbulo en proceso de diferenciación adopta la forma de una S, se observan diferentes patrones de expresión genética a lo largo de el. Cerca del futuro extremo glomerular, los niveles de expresión del Pax-2 caen mientras que comienza a expresarse niveles altos de WT-1 (figura 15-3). En el otro extremo del túbulo (futuro túbulo contorneado distal), el Wnt-4 y el E- cadherina siguen siendo importantes, en tanto que la mitad proximal (futuro túbulo contorneado proximal) la K-cadherina es un importante marcador celular. Muchas de las células mesenquimatosas no inducidas situadas entre los túbulos sufren apoptosis.
La diferenciación del túbulo renal avanza desde el glomérulo hasta el túbulo contorneado proximal y luego hasta el distal. Durante la diferenciación de la nefrona, una parte del túbulo se convierte en el asa de Henle, un asa alargada en forma de horquilla para el pelo que se extiende hasta la medula renal. Durante la diferenciación la células epiteliales desarrollaron rasgos moleculares característicos del riñón maduro (ejemplo, antígenos del borde en cepillo o la glucoproteinas de Tamm-Horsfall (figura 15-4).
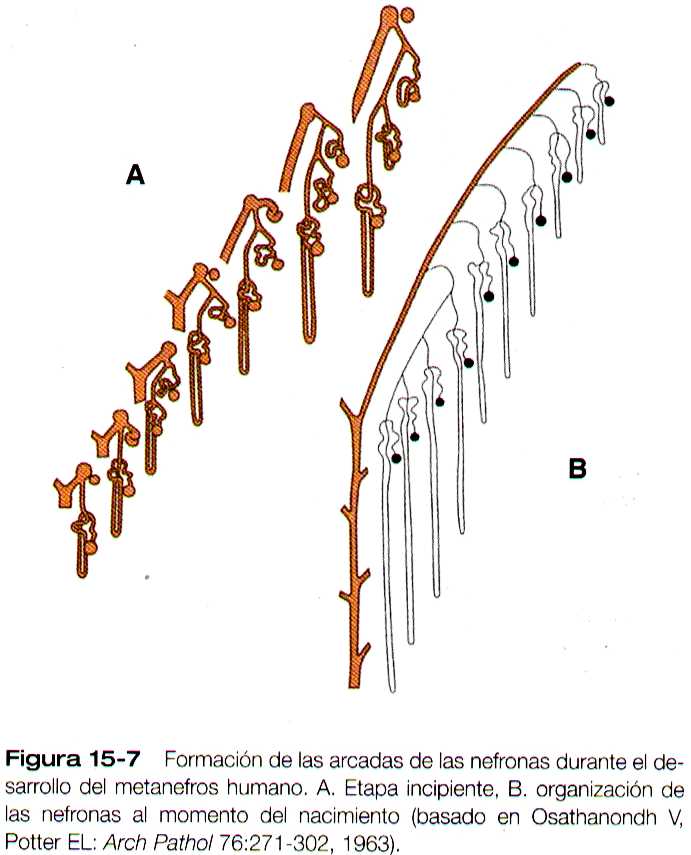
El crecimiento del riñón implica la formación de más de quince generaciones sucesivas de nefronas en su zona periférica; los más superficiales son menos maduros que los más interiores. El desarrollo de la estructura interna del riñón es muy complejo; e incluye la formación de arcadas de neuronas muy ordenadas (figura 15-7). Los detalles se encuentran fuera del alcance de este texto.
Cambios posteriores en el desarrollo del riñón
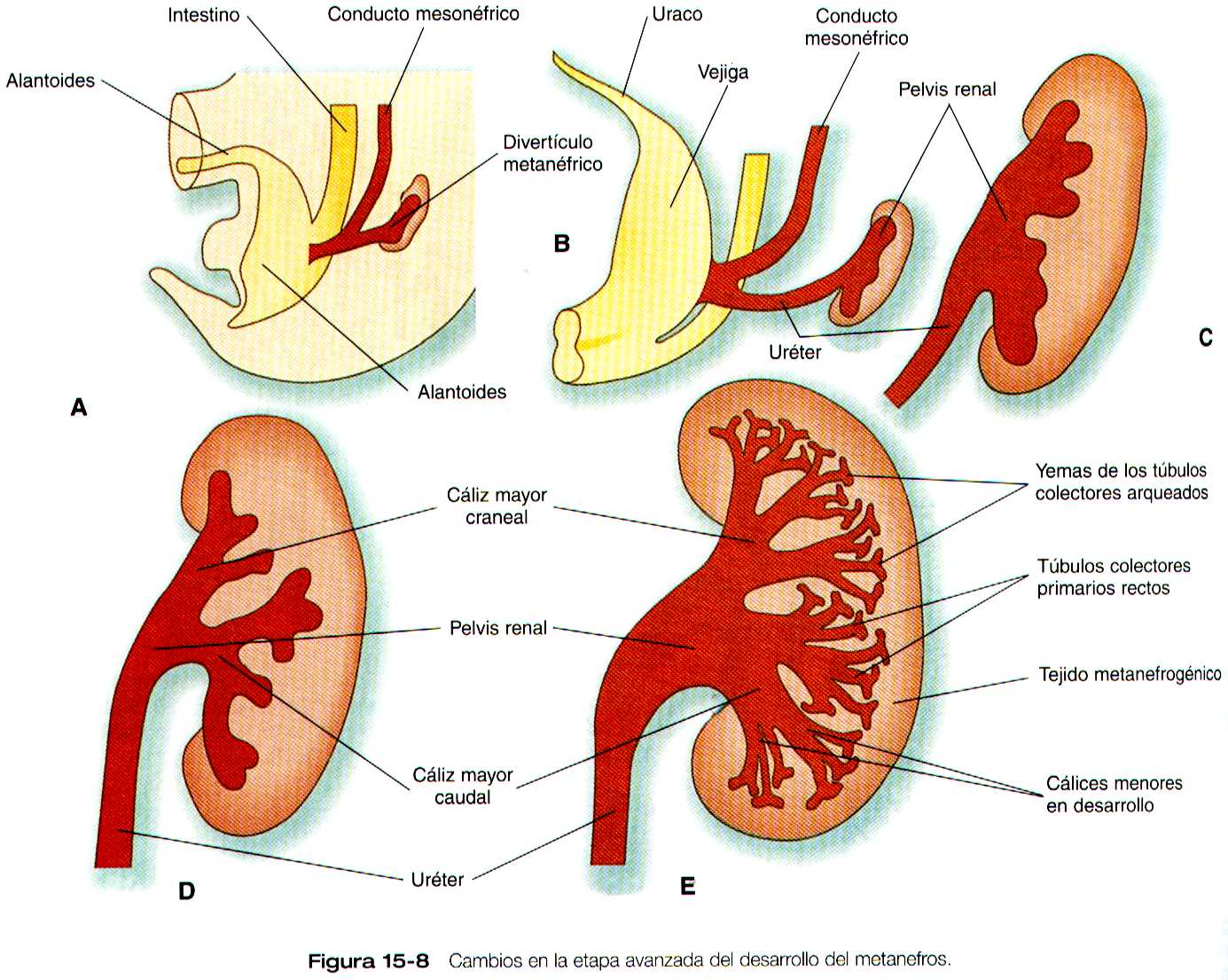
A medida que muchos grupos de nefronas están desarrollándose, el riñón crece cada vez más. El sistema ramificado de conductos también se vuelve mucho más grande y complejo, y da lugar a la pelvis y al sistema de cálices renales (figura 15-8).
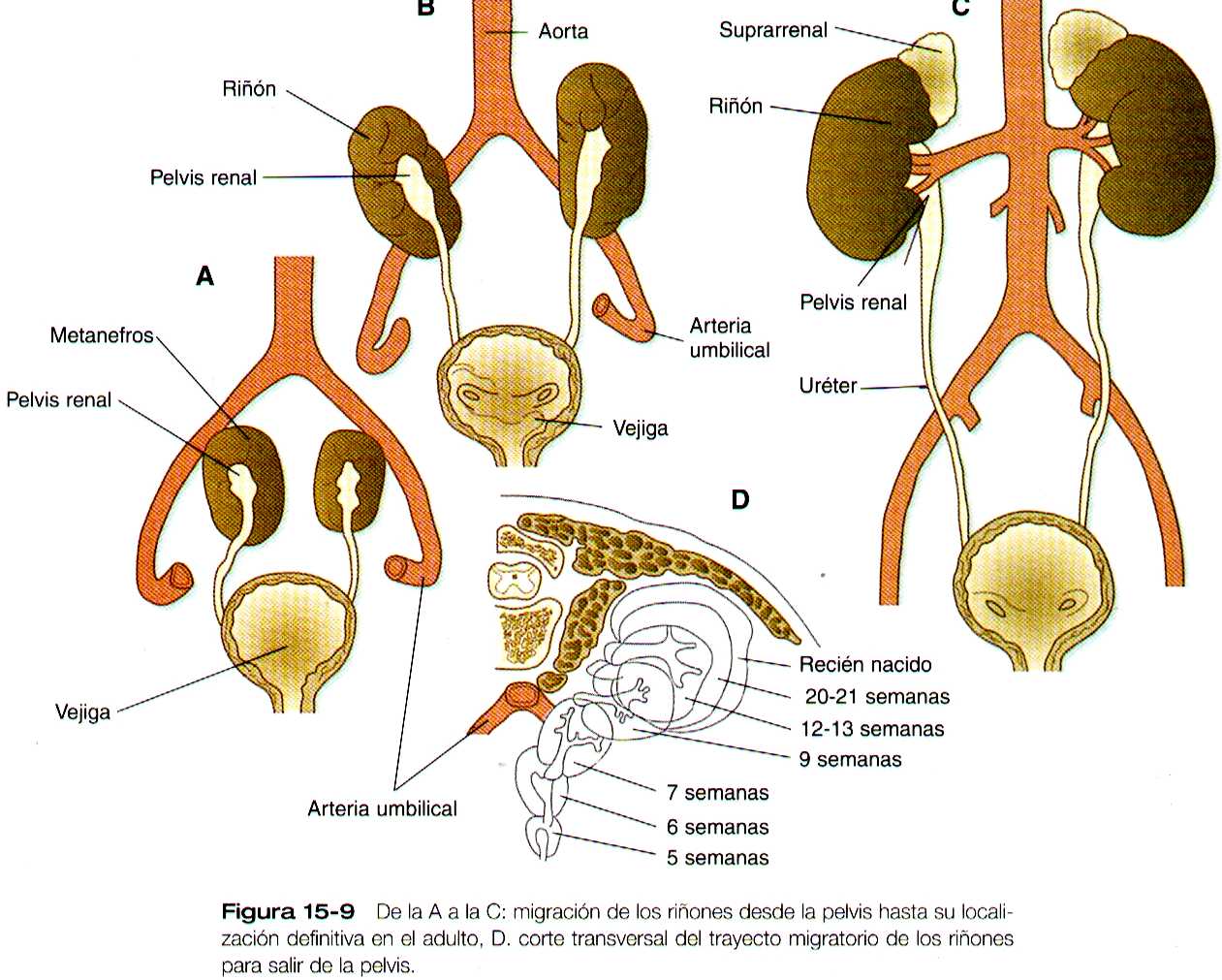
Estas estructuras recolectan la orina y la canalizan hacia los uréteres a la manera de un embudo. Durante gran parte del periodo fetal, los riñones están divididos en lóbulos visibles, que en el periodo neonatal, cuando se forman, los riñones metanéfricos están localizados en la profundidad de la región pélvica. Al final del periodo embrionario y al comienzo del fetal, experimentan un importante desplazamiento y se sitúan en la región abdominal. Este desplazamiento resulta en parte de una emigración renal y en parte de un marcado crecimiento de la región caudal del embrión. Se producen dos componentes concurrentes de la emigración: uno es un desplazamiento caudocraneal desde el nivel de la cuarta lumbar hasta la primera lumbar o incluso hasta la décimo segunda torácica. (figura 15-9); el otro un desplazamiento lateral. Estos cambios llevan a los riñones a contactar con las glándulas suprarrenales, que forman un casquete de tejido glandular en el polo craneal de cada riñón. Durante su migración, los riñones también experimentan una rotación de 90 grados, por la cual las pelvis terminan mirando hacia la línea media. Conforme migran fuera de la cavidad pélvica, los riñones se deslizan por encima de las grandes arterias umbilicales, que se ramifican del extremo caudal de la aorta. Todos estos cambios tienen lugar por detrás del peritoneo, ya que los riñones son órganos retroperitoneales. Durante las primeras fases de la migración de los riñones metanéfricos, los riñones mesonéfricos involucionan. No bastante, los conductos mesonéfricos se conservan ya que terminan asociándose estrechamente con las gónadas en desarrollo.
Aunque el riñón adulto normalmente es irrigado por la arteria renal grande que es rama directa de la aorta, presenta cinco lóbulos o ligamentos vasculares. Las arterias que irrigan cada uno de estos lóbulos al principio eran vasos segmentarios que irrigaban al mesonefros y que fueron reemplazadas por este en su migración. Sus orígenes aórticos se ven reducidos al par único de arterias renales, pero las variaciones anatómicas son frecuentes.
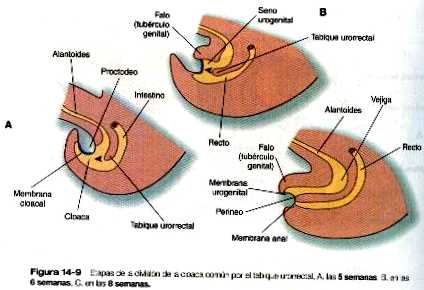
Formación de la vejiga
La división de la cloaca en el recto y la región del seno urogenital se expuso en el capitulo 14 (figura 14-9). El seno urogenital se continúa con la alantoides, la cual tiene una base amplia que se continúa con el seno urogenital, y un extremo tubular que se introduce el pedículo vitelino. La base ancha de la alantoides sigue creciendo y formara la vejiga; su extremo distal se solidificara y dará lugar al uraco, una estructura de forma cordonal que dará origen al ligamento umbilical mediano que va desde la vejiga hasta la región umbilical (figura 15-18).
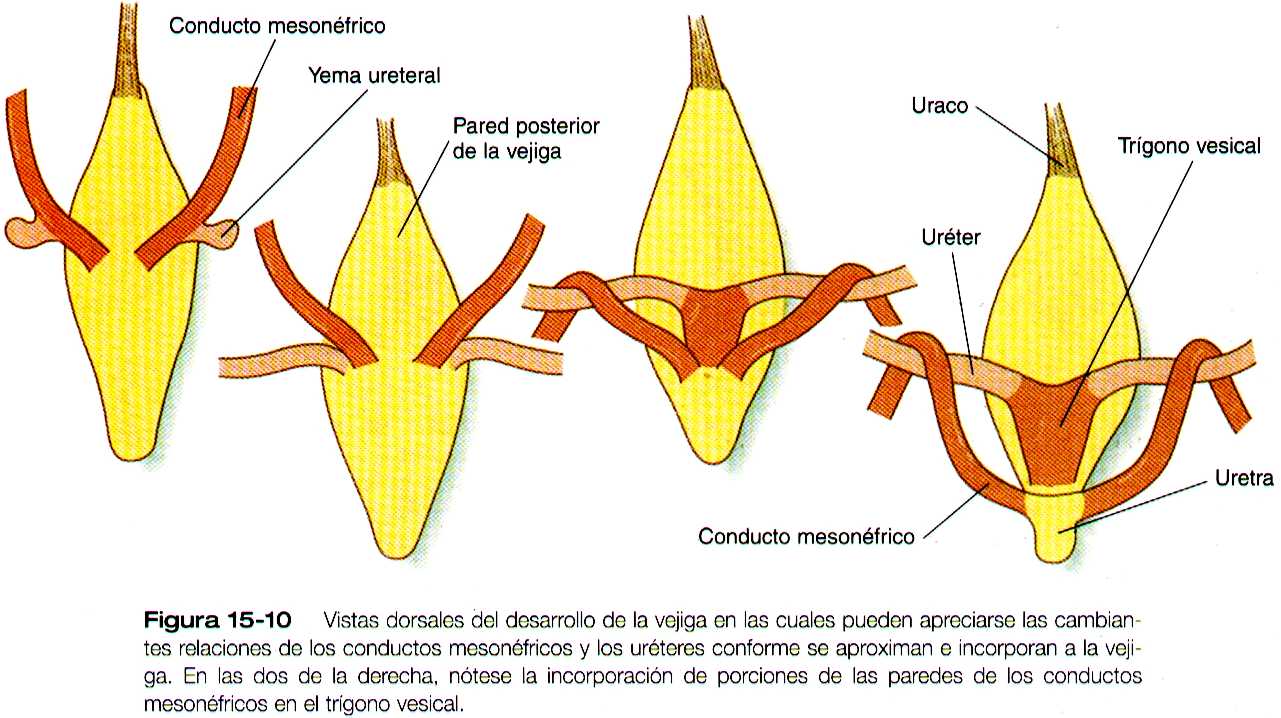
Conforme la vejiga va creciendo, su pared incorpora los conductos mesonéfricos y las yemas ureterales (figura 15-10). Como resultado, estas estructuras se abren por separado en la pared posterior de la vejiga. Mediante un mecanismo todavía no muy claro, que posiblemente implica la tensión mecánica ejercida por los riñones en su migración, los extremos de los uréteres se abren en la vejiga en situación lateral y cefálica a los conductos mesonéfricos. La región delimitada por estas estructuras se llama trígono vesical. En la región de entrada de los conductos mesonéfricos, la vejiga se reduce considerablemente. Esta región, que en su origen era parte del seno urogenital, forma la uretra que será la vía de salida de la vejiga (Pág. 387)
SISTEMA GENITAL
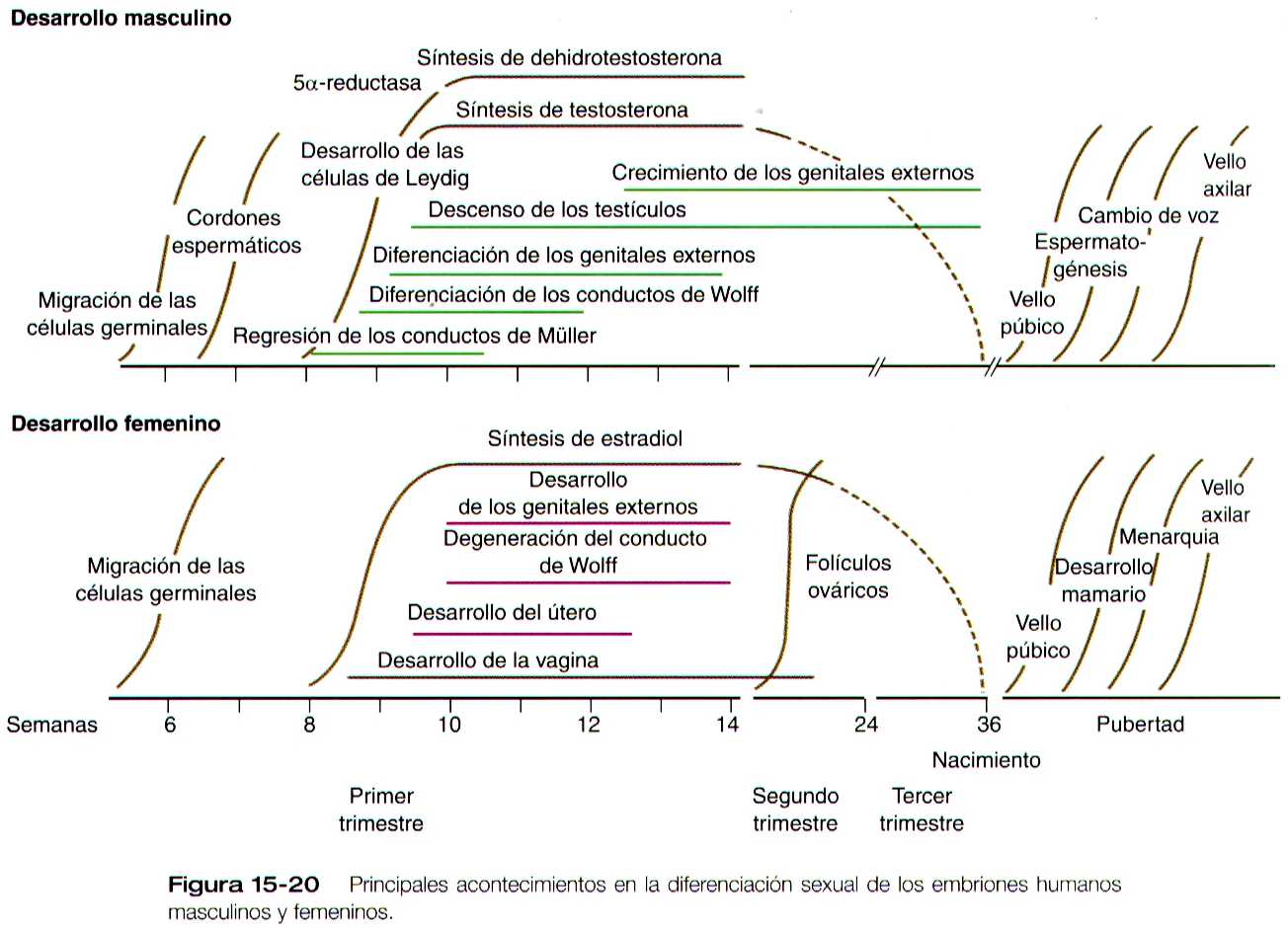
El desarrollo del sistema genital es una parte de la diferenciación sexual general de un individuo (figura 15-20). La determinación del sexo comienza en la fecundación, cuando un cromosoma Y o un cromosoma X adicional se unen al cromosoma X que ya existe en el óvulo. Esta fase es la determinación genética del género. Aunque el sexo genético del embrión se fija en el momento de la fecundación, el fenotípico viene a manifestarse hacia la séptima semana de desarrollo. Antes de ese momento, el principal indicador morfológico del sexo del embrión es la presencia o ausencia de la cromatina sexual (corpúsculo de Barr) en la mujer. El corpúsculo de Barr es el resultado de la inactivación de un cromosoma X. durante esta etapa de indiferenciación desde el punto de vista morfológico del desarrollo sexual. Los futuros gametos migran hacia los primordios gonadales desde el saco vitelino.
Tradicionalmente se ha considerado que la diferenciación fenotípica del genero comienza con las gónadas y avanza en virtud de las influencias gonadales sobre los sistemas de conductos sexuales. Unas influencias similares en la diferenciación de los genitales externos y por ultimo en el desarrollo de las características sexuales secundarias (por ejemplo, la configuración corporal, las mamas y los patrones de implantación del pelo) completan los sucesos que constituyen el proceso general de diferenciación sexual.
*sin embargo, las investigaciones recientes han demostrado diferencias sexuales ya desde antes de la implantación del embrión. Los genes Sry (véase mas adelante) ya se han transcrito antes de la implantación. Además previo a la implantación el embrión XY se desarrolla con más rapidez que el embrión XX. Los embriones masculino y femenino previos a la implantación son distinguidos desde el punto de vista antigénico. Esto sugiere diferencias en la expresión de genes. Continua en la Pág. 376
También da lugar la diferenciación sexual del cerebro, que influye en el comportamiento.
En algunas circunstancias el sexo genético de un individuo puede ser anulado por factores ambientales de manera que el genotipo y el fenotipo no se correspondan. Un principio general importante es que el desarrollo del fenotipo masculino requiere de la acción de sustancias producidas por el testículo. En ausencia de influencias testiculares específicas o de la capacidad de responder a ellas, el resultado es un fenotipo femenino. Este último es la condición basal (default), o por defecto, sobre la cual deben actuar las influencias masculinas para producir un fenotipo masculino.
Determinación genética del sexo
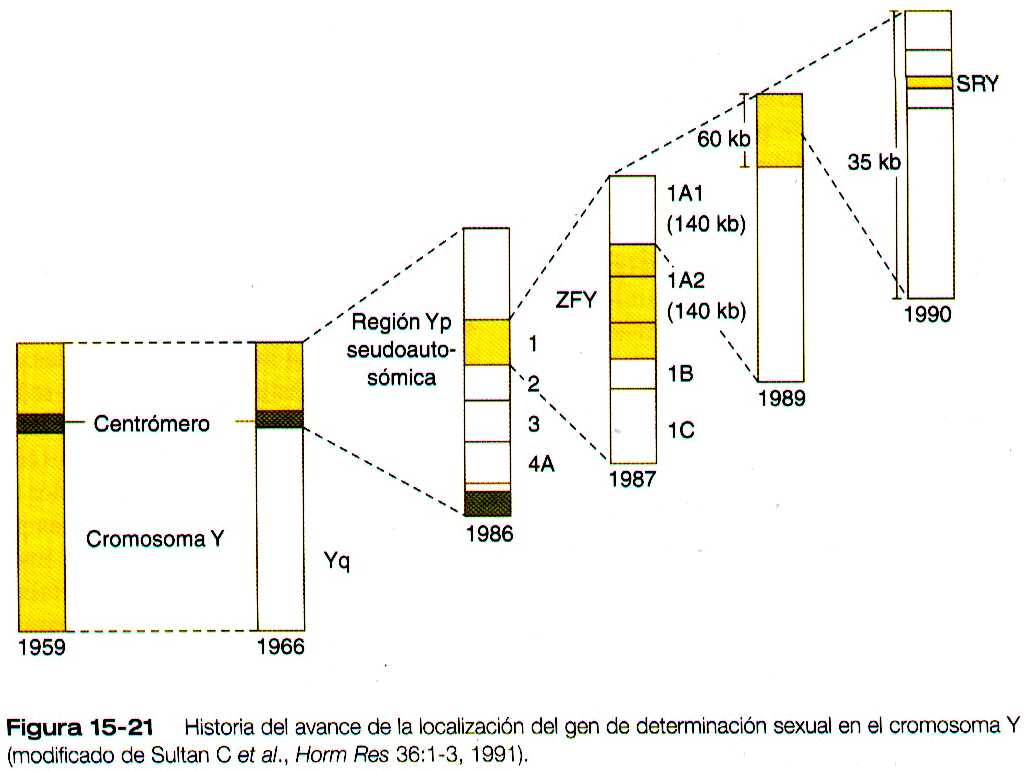
Desde 1923 los científicos han reconocido que los pares de cromosomas XX y XY representan la base genética de la feminidad y la masculinidad humanas. Durante muchos decenios se creyó que la presencia de dos cromosomas X era el factor que determinara el sexo, pero en 1959 se estableció que la diferenciación entre los seres humanos masculinos y femeninos depende del cromosoma Y.
No obstante, el vinculo entre el cromosoma Y y la determinación del testículo no se había descubierto. Durante las últimas décadas se han propuesto tres candidatos para el factor de determinación testicucular.
El primero fue el antígeno H-Y, un antígeno menor de histocompatibilidad presente en las células de los varones pero no en las de las mujeres. Este antígeno se ha localizado en el brazo corto del cromosoma Y humano. Se ha considerado que es un producto del gen determinante de los testículos en los mamíferos. No obstante, se encontró una cepa de ratones (Sxr) que reproduce machos en ausencia del antígeno H-Y. Se demostró que los ratones Sxr tienen una transposición de una región del cromosoma Y hacia el cromosoma X, pero el locus que codifica el antígeno H-Y no estaba incluido. Además, en ciertos varones humanos fenotípicos con genotipo XX se demostró la ausencia del material genético para el antígeno H-Y.
El siguiente candidato fue un locus del brazo del cromosoma Y llamado gen en Zinc finger Y (ZFY). Con las técnicas de hibridación del ácido desoxirribonucleico (DNA), se ha encontrado este gen tanto en los varones humanos XX como en los ratones en los que se traspusieron pequeños fragmentos de los cromosomas X y Y durante el entrecruzamiento de la meiosis.
A la inversa, este gen estaba ausente en algunos fenotipos femeninos XY. No obstante, se encontró que ciertos varones XX carecían del gen, y otros casos muy raros de anomalías en la diferenciación sexual no mostraban una correspondencia entre el fenotipo sexual y la presencia o ausencia del gen ZFY.
El candidato mas reciente para el gen de determinación testicular es el llamado Sry, que también esta localizado en el brazo corto del cromosoma Y en una región de 35 kilo bases (figura 15-21). El gen Sry codifica una proteína no-histona d 223 aminoácidos que pertenece a la familia de proteínas que contienen una región fijadora de DNA muy conservada de 59 aminoácidos llamada higt mobility group box. Después de que se clono el gen, se detecto en muchos casos de inversión de sexo, incluidos machos XX sin genes ZFY. El gen Sry esta localizado en el cromosoma Y humano cerca de la región homologa, lo que hace susceptible su traslocación al cromosoma X.
El gen Sry también esta ausente en una capa de ratones XY que tienen fenotipo femenino. Otras pruebas experimentales han constituido en producir ratones transgénicos con la inserción de un fragmento de 14 kilobases del cromosoma Y que tiene el gen Sry. Muchos de los ratones transgénicos XX se convierten en machos fenotipicos con testículos normales y comportamiento masculino. Los estudios de hibridación in situ en ratones han demostrado que la expresión del producto del gen Sry tiene lugar en el tejido gonadal en momento de la determinación sexual, pero no se expresa en las gónadas de los embriones femeninos.
CORRELACION CLINICA 15-1
Anomalías congénitas del sistema urinario
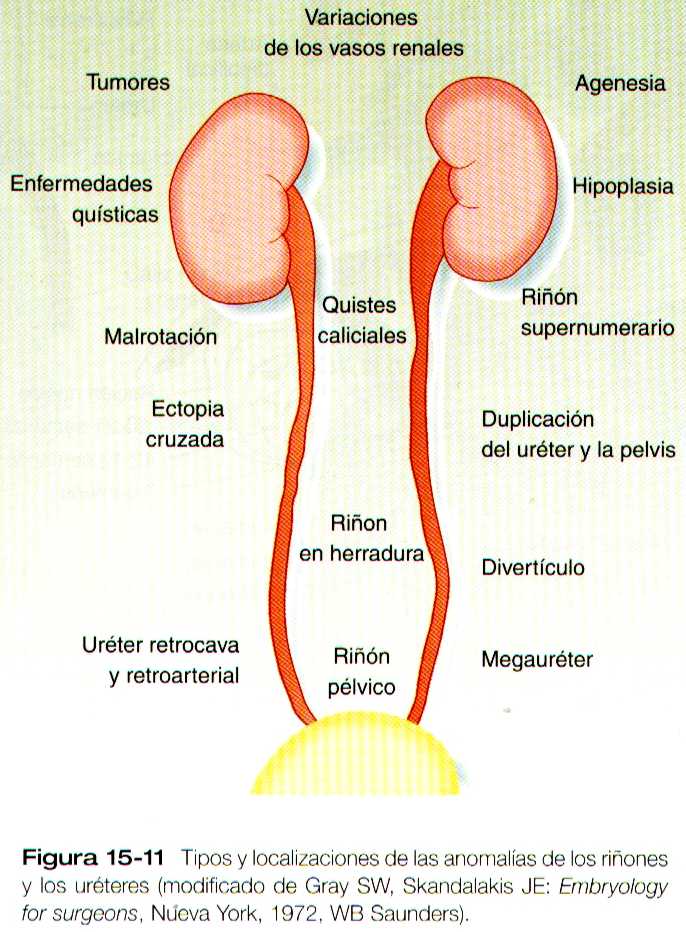
Las anomalías del sistema urinario son relativamente frecuentes (3 a 4% de los nacidos vivos). Muchas son asintomáticas y otra solo se manifiestan tardíamente. En la figura 15-11 se resumen las localizaciones de muchas de las malformaciones mas frecuentes del sistema urinario.
CORRELACION CLINICA 15-1a
Anomalías congénitas del sistema urinario (continuación)
AGENESIA RENAL
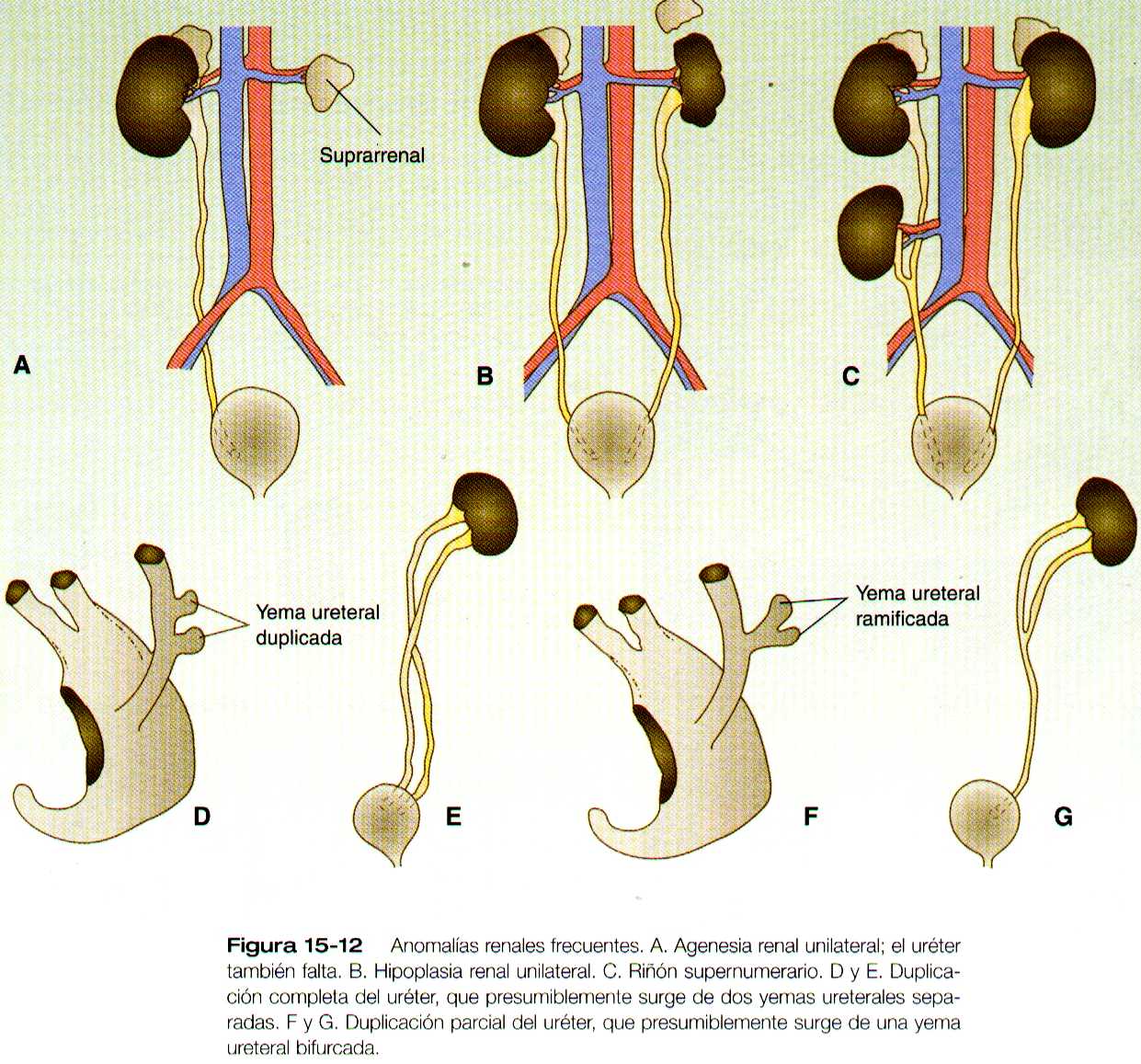
La agenesia renal es la ausencia uní o bilateral de cualquier tejido renal (figura 15-12 A). La agenesia renal unilateral se ve en cerca del 0,1% de los adultos, en tanto que la bilateral afecta a uno de cada 3,000 a 4,000 recién nacidos. El uréter puede estar presente. Esta anomalía se interpreta por lo general como un defecto de la interacción inductiva entre la yema ureteral y el mesénquima metanefrogénico. La expresión equivoca de las moléculas, como el Pax-2, el WT-1 o el Wnt-4, que son importantes en el desarrollo metanéfrico inicial, es probablemente una de las causas de algunos casos de agenesia renal. Los individuas afectados por agenesia renal unilateral a menudo son asintomáticos, pero es frecuente que el riñón único experimente una hipertrofia compensatoria para mantener una masa funcional normal de tejido renal.
Un bebe que nace con agenesia renal bilateral muere pocos días de nacer. A causa de la falta de diuresis, un rasgo asociado frecuentemente durante el embarazo es la reducción del liquido amniótico (oligohidramnios). Los bebes que nacen con agenesia renal bilateral muestran fascies de Potter, que se caracteriza por nariz achatada, espacio interpupilar amplio, micrognatia, dedos afilados y orejas grandes.
CORRELACION CLINICA 15-1b
Anomalías congénitas del sistema urinario (continuación)
HIPOPLASIA RENAL
Una afección intermedia entre la agenesia renal y el riñón normal es la hipoplasia renal (figura 15-12 B), en la cual un riñón, o con menor frecuencia los dos, es mucho mas pequeño de lo normal aunque puede conservar algo de su función. Aunque no se han identificado una causa especifica para la hopoplásia renal, algunos casos pueden relacionarse con diferencias en los factores de crecimiento o en sus receptores que son muy activos durante las ultimas fases criticas de la metanefrogénesis. Como el caso de la agenesia renal, es posible que el riñón normal opuesto al riñón hipoplásico experimente una hipertrofia compensatoria.
DUPLICACIONES RENALES
Las duplicaciones renales van desde una duplicación sencilla de la pelvis renal hasta un riñón supernumerario separado. Como sucede con los riñones hipoplásicos, las duplicaciones renales pueden ser asintomáticas, aunque la incidencia de infecciones renales puede ser mas alta. También se han descrito muchas variantes en la duplicación del uréter (figura 15-12), que suelen atribuirse a la división o a una amplia separación de las ramas de la yema ureteral.
ANOMALIAS DE LA MIGRACION Y LA ROTACION RENAL
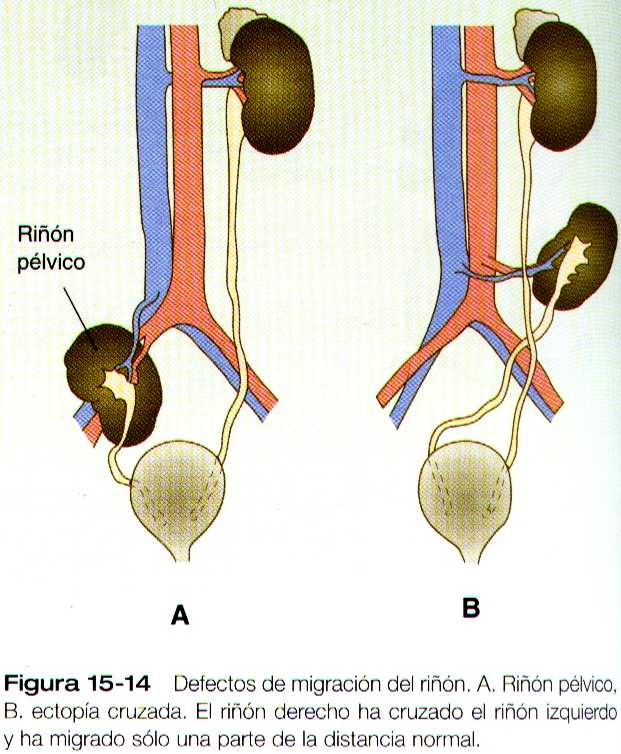
La alteración mas frecuente de la migración renal es que un riñón se quede en la cavidad pélvica (figura 15-14 A). Por lo general esta alteración también se asocia con la mal rotación del riñón, de manera que el hilio renal mira hacia delante en lugar de hacia la línea media. Otra categoría de malformaciones migratorias es la ectopia cruzada, en la cual el riñón y su uréter asociado se encuentran en el mismo lado del cuerpo que el otro riñón (figura 15-14 B). En esta alteración el riñón ectopíco puede estar funcionando con el normal.
CORRELACION CLINICA 15-1c
Anomalías congénitas del sistema urinario (continuación)
En la afección conocida como el riñón en herradura, que se presenta hasta en uno de cada 400 individuos, los riñones están fusionados por sus polos inferiores (figura 15-15). Los riñones en herradura no pueden migrar fuera de la pelvis porque la arteria mesentérica inferior, que se desprende de la aorta, los bloquea. En la mayoría de los casos esta afección es simbiótica, pero en ocasiones puede presentarse dolor u obstrucción de los uréteres. Esta enfermedad puede asociarse con malformaciones de otros órganos internos. Los riñones pélvicos están sujetos a una incidencia mayor de infecciones y de obstrucción ureteral.
CORRELACION CLINICA 15-1d
Anomalías congénitas del sistema urinario (continuación)
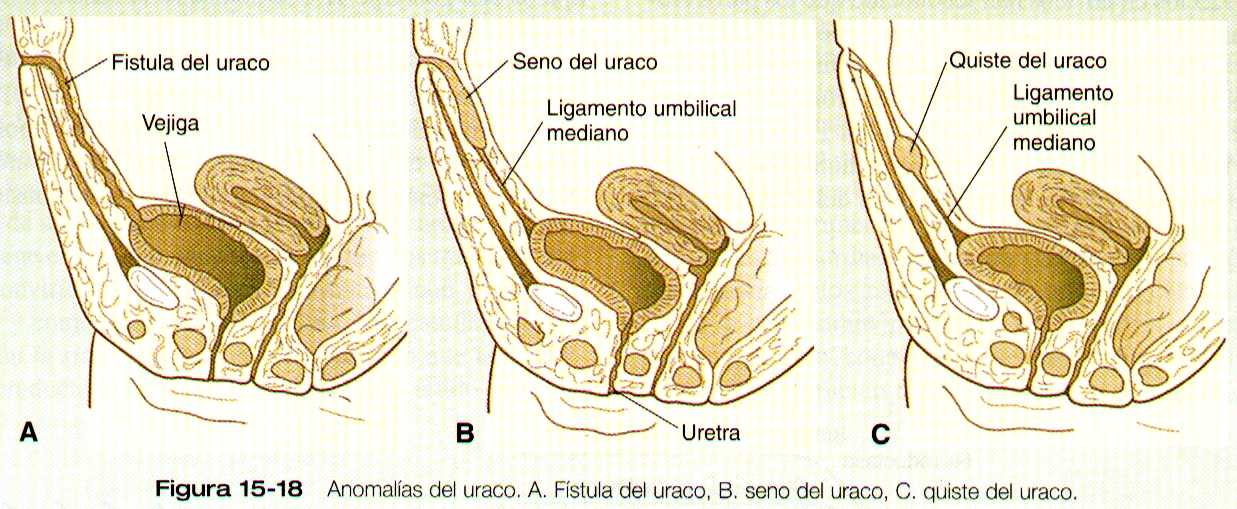
QUISTES, SENOS Y FISTICULAS DEL URACO
Si algunas parte de la luz de la alantoides no se obliteran, puede formarse quistes, senos o fístulas del uraco (figura 15-18). En el caso de la fístula del uraco, se filtra orina a través del ombligo. Los senos o quistes aunque no sean evidentes en la infancia pueden inflamarse mas tardíamente.
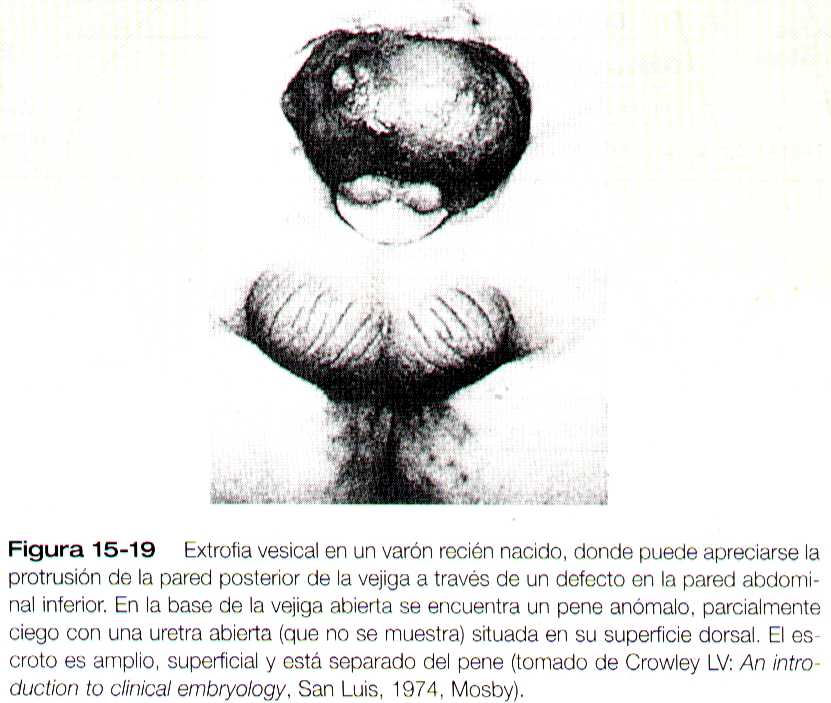
EXTROFIA VESICAL
La extrofia vesical es un grave defecto en el cual la vejiga se abre ampliamente en la pared abdominal (figura 15-19). Mas que un defecto primario del sistema urinario, suele atribuirse a insuficiencia del tejido mesodérmico en la formación de la pared ventral del abdomen. Aunque en un comienzo la pared ventral del cuerpo puede estar cubierta de ectodermo, si no tiene mesodermo se rompe, y a continuación degenera la pared anterior de la vejiga . En los varones, la extrofia vesical suele afectar al falo, de lo que resulta una afección denominada epispadias (Pág. 393).
Migración de las células germinales hacia las gónadas
La detección precoz de las células germinales primordiales en el revestimiento del saco vitelino y su emigración hacia las gónadas en los embriones humanos se describen en el capitulo 1. los estudios descriptivos y experimentales han demostrado que las células germinales primordiales en el ratón puede detectarse primero en el epiblasto. Estas células pasan por la línea primitiva y a continuación se localizan como un pequeño cúmulo de células en el mesodermo de la pared posterior del saco vitelino (figura 1-1).
En los embriones humanos las células germinales migran desde la pared posterior del saco vitelino a lo largo de la pared del intestino posterior y a través del mesenterio dorsal hasta que alcanza la región de las crestas genitales recién formadas. Hay pruebas experimentales que sugieren que las primeras etapas de la migración de las células germinales primordiales hacia las gónadas son posibles gracias a movimientos ameboideos activos de las células, en respuesta a un sustrato permisivo de la matriz extracelular.
También pueden contribuir los desplazamientos tisulares gracias al crecimiento diferencial de la región posterior del embrión. Durante su migración muchas células germinales primordiales están unidas entre si mediante largas prolongaciones citoplasmáticas. Todavía no se ha determinado como estas interconexiones controlan la migración o el asentamiento en las gónadas. Las células germinales migratorias proliferan en respuesta a factores mitogénicos como el factor inhibidor de la leucemia y el factor de crecimiento de los mastocitos (factor de las células stem (madre) o factor de Steel).
Conforme la células germinales se aproximan a los crestas genitales al final de la quinta semana del desarrollo, pueden ser influenciadas por factores quimiotácticos segregados por las gónadas recién formadas. Estas influencias se han demostrado gracias al injerto de tejidos embrionarios (por ejemplo, intestino posterior, que contiene células germinales dispersas) en la cavidad corporal de un embrión huésped. Las células germinales primordiales del injerto se concentran mas cerca de las crestas genitales del huésped, en ocasiones migran desde allí hacia las crestas genitales. Entre mil y dos mil células germinales primordiales entran en las crestas genitales. Una vez allí, su comportamiento migratorio se detiene.
Algunas células germinales primordiales siguen trayectos migratorios incorporados, que las llevan a establecerse en localizaciones extragonadales. Estas células suelen comenzar a desarrollarse como oogonias, sin importar su genotipo, y después se degeneran. Sin embargo, en unos pocos casos pueden presentir en las localizaciones ectópicas, como el mediastino, y dar lugar a teratomas (capitulo I).
Establecimiento del sexo gonadal
Origen de las gónadas
las gónadas se originan en una región alargada de mesodermo esteroidogénico situado a los largo del bode ventromedial del mesonefros. Las células de la parte craneal de esta zona se condensan para formar los primordios adrenocorticales, y las de la parte caudal se convierten en las crestas genitales, identificables hacia la quinta semana. Las crestas genitales incipientes se componen de dos poblaciones principales de células: una derivada del epitelio celómico y la otra procedente de la cresta mesonefrica. La formación de las crestas genitales requiere de la formación de al menos dos genes: WT-1, que también es importante al comienzo de la formación de los riñones (figura 15-3) y el SF-1 (factor esteroidogénico-1), necesario para el desarrollo tanto de las gónadas como de las glándulas suprarrenales.
Diferenciación de los testículos
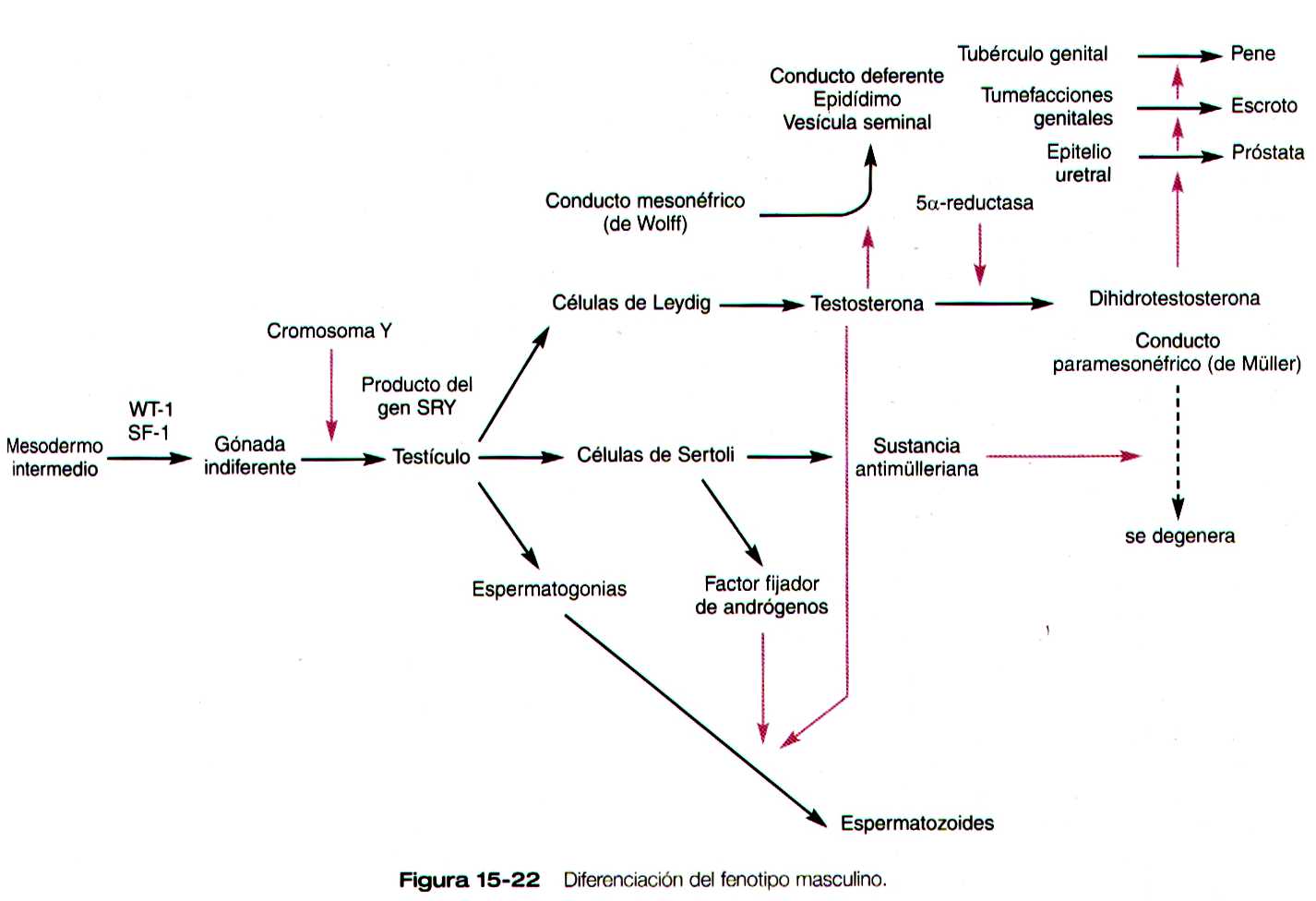
Cuando aparecen las crestas genitales, las de los machos y las hembras son distinguibles desde el punto de vista morfológico (etapa indiferenciada). El patrón general que controla la diferenciación gonadal es que bajo la influencia del gen Sry (factor de determinación testicular) en el cromosoma Y, la gónada indiferenciada se convierte en un testículo (figura 15-22). Si no se expresan los productos de este gen, la gónada se diferencia en un ovario.
En los machos, se detectan transcriptos del gen Sry solo en la cresta genital justo al comienzo de la diferenciación del testículo. Ni la expresión de este gen, ni la diferenciación tardía de los testículos dependen de la presencia de células germinales. Los genes que determinan el sexo actúan sobre la porción somática del testículo y no sobre estas células. Todavía queda mucho por averiguar sobre los acontecimientos que llevan desde la expresión del producto del gen Sry hasta la diferenciación de los testículos. Hasta ahora las pruebas sugieren que el producto del gen Sry actúan ante todo como un interruptor que activa otros genes necesarios para dirigir la diferenciación con sucesos morfológicos específicos.
El tiempo es importante en la diferenciación testicular. El testículo se desarrolla antes que el ovario. Las precursoras de las células de Sertoli deben estar preparadas para recibir la señales genéticas para la diferenciación testicular en un determinado momento; si esto no sucede, las células germinales primordiales comienzan a hacer meiosis y la gónada se convierte en un ovario. Al parecer, la diferenciación inicial del testículo depende de una señal procedente del mesonefros. Sin este, el desarrollo de las estructuras internas ( cordones espermáticos ) es deficiente.
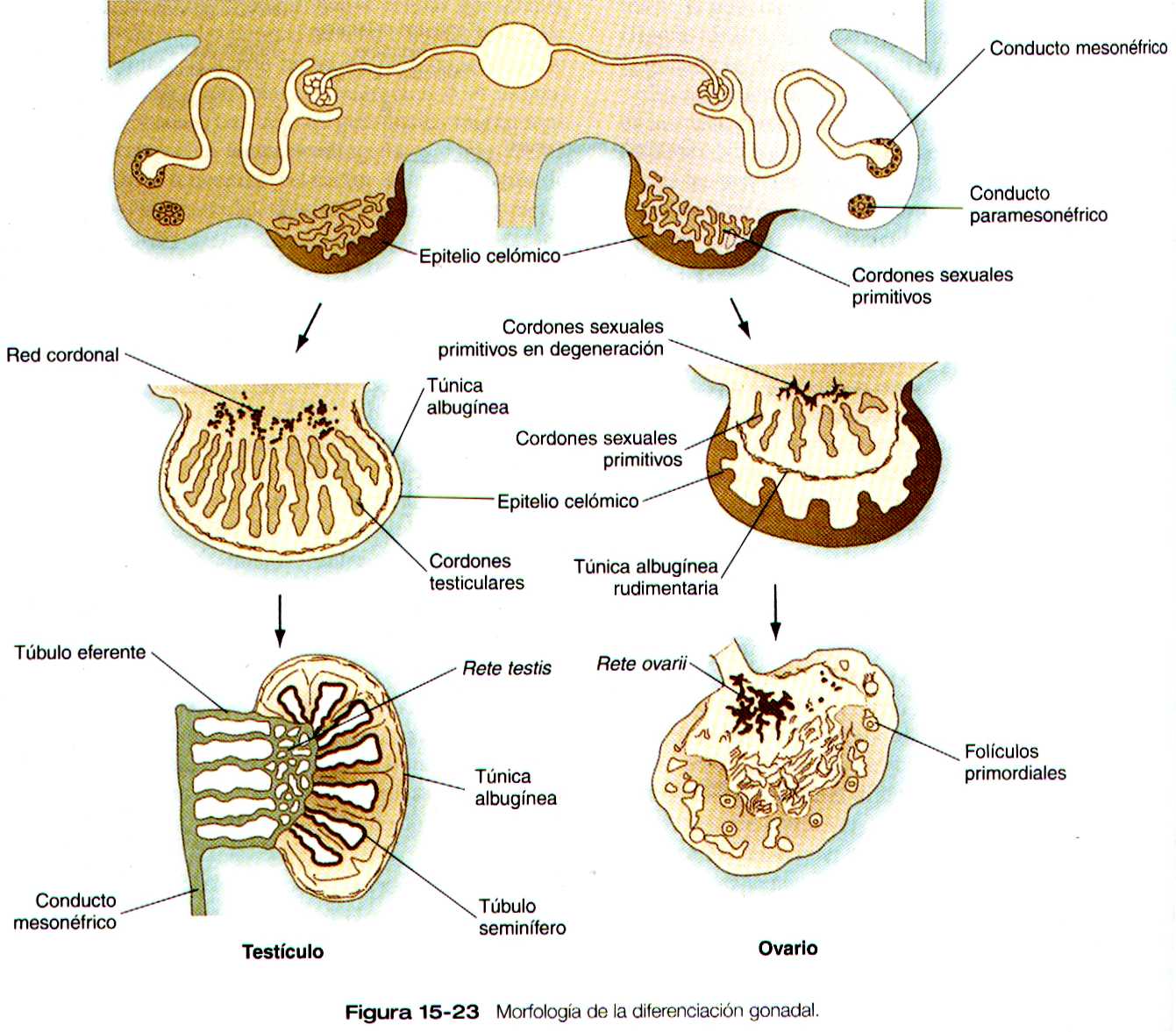
La morfología de la diferenciación gonadal inicial ha sido materia de controversia, ya que se ha propuesto varios tipos de linaje e interacciones celulares. Según datos morfológicos recientes, las crestas genitales aparecen a mitad de la quinta semana gracias a la proliferación de células del epitelio celómico a lo largo del borde interno de mesonefros (figura 15-23). Mas tarde en la quinta semana las células germinales primordiales entran en la cresta genital incipiente y el epitelio celómico emite unos cordones epiteliales cortos hacia el interior de la gónada. Al comienzo de la sexta semana un grupo de cordones sexuales primitivos se forman en la cresta genital, y las células germinales primordiales migran en el interior de ellos. Al parecer, la gónada masculina emite un agente quimioatrayente que estimula la migración desde el mesonefros de células mioides que rodean a los cordones sexuales. Además, las células endoteliales y las mío epiteliales que rodean a los vasos migran hacia el interior del testículo procedente del mesonefros. El tejido ovárico no atrae a estas células.
Hacia la sexta semana los testículos muestran signos de diferenciación. Los cordones sexuales primitivos se agrandan y se define mejor y se cree que sus celulas representan los precursores de las células de Sertoli. Conforme los cordones sexuales se diferencian, se separan del epitelio superficial (epitelio germinativo ) mediante una densa capa de tejido conectivo llamada túnica albugínea. Las porciones mas profundas de los cordones sexuales testiculares están en contacto con las formaciones quinta a decimosegunda de las nefronas mesonéfricas.
Las zonas mas periféricas de los cordones sexuales testiculares forman los túbulos seminíferos, y las porciones centrales adquieren la forma de red y por ultimo dan lugar a la rete testis. Al final esta se une a los conductillos eferentes, que derivan de los túbulos mesonéfricos.
Durante los primeros dos meses del desarrollo, las células de Leydig no son identificables en el testículo embrionario. Aparecen durante la octava semana y pronto comienzan a sintetizar hormonas androgénicas (testosterona y androstenediona). Esta actividad endocrina es importante porque la diferenciación del sistema de conductos masculinos y de los genitales externos depende de las hormonas sexuales producidas por el testículo fetal. Las células de Leydig fetales secretan sus productos hormonales justamente en el periodo durante el cual tiene lugar de la diferenciación de los conductos genitales y que son sensibles a las hormonas (semanas 9 a 14). Después de las semanas 17 y 18, las células de Leydig experimentan una involución gradual y solo reaparecen en la pubertad, cuando estimulan la espermatogénesis. Las células de Leydig fetales pueden considerarse como una isoforma celular que mas adelante es remplazada por la forma definitiva adulta de las células. Hacia las ocho semanas las células de Sertoli embrionarias producen la sustancia antimülleriana (Pág. 381), que también desempeña un importante papel en la formación del sistema de conductos sexuales porque provoca la involución de los precursores de los conductos genitales femeninos.
Al final de la vida embrionaria y en los periodos fetal y prenatal. Las células germinales primordiales del testículo se dividen lentamente por mitosis, pero las células de Sertoli fetales son insensibles a los andrógenos y no maduran. El testículo embrionario parece segregar un factor inhibidor de la meiosis, pero no se ha logrado aislar o caracterizar un factor de este tipo. El ambiente del testículo solo se vuelve favorable a la meiosis y la espermatogénesis después de la pubertad.
Diferenciación del ovario
En ausencia de las señales especificas de diferenciación testicular, las gónadas acaban convirtiéndose en ovarios. En contraste con los testículos, la presencia de células germinales visibles es esencial para la diferenciación ovárica; si tales células no llegan a las crestas genitales o si hay anomalías (por ejemplo, XO) y se degeneran, la gónada se retrae y el resultado son cintillas ováricas (ovarios vestigiales)
Después de que las células germinales primordiales han llegado al futuro ovario, permanecen concentradas en la región corticomedular externa o cerca del límite de la misma. Al igual que el testículo, el ovario contiene cordones sexuales primitivos en la región medular, pero están tan bien desarrollados como los de los testículos . El origen de las células que forman los folículos ováricos no se han determinado. Se han propuesto tres lugares de origen para las células del epitelio folicular: 1) el epitelio celómico ( cordones sexuales secundarios), 2) los cordones sexuales primitivos del origen mesonéfrico y 3) una combinación de los dos. La última posibilidad concuerda con la presencia de dos tipos diferentes de células-claras y obscuras en el epitelio folicular inicial.
Las células germinales primarias, ahora llamadas oogonias, proliferan por mitosis desde el momento que entran en la gónada hasta el inicio del cuarto mes de gestación. Entonces, algunas oogonias de la región medular interna del ovario entran en la profase de la primera división meiótica, posiblemente bajo la influencia de un factor estimulante de la meiosis que procede del mesonefros. Esta influencia puede estar asociada con cúmulos de epitelio derivados del mesonefros llamados rete ovarii medular. La oogonias en meiosis, ahora llamadas oocitos, se asocian con las células foliculares y se forman los folículos primordiales (figura 1-5). Entre tanto, las oogonias de la región cortical del ovario continúan dividiéndose por mitosis. Las oogonias y los oocitos primarios están unidos por puentes citoplásmicos intercelulares que pueden intervenir en la sincronización de su desarrollo. Hacia la semana 22, el desarrollo folicular esta bastante avanzado en todo el ovario. Los oocitos continúan en meiosis hasta que alcanza la etapa de diplotene de la profase de la primera división meiótica. Entonces se detiene la meiosis y los oocitos permanecen en esta etapa hasta que desaparece el bloqueo.
En el adulto esto sucede en los oocitos individuales unos pocos días antes de la ovulación . En las mujeres premenopáusicas, pueden haber pasado hasta 50 años desde que estos oocitos entraron en el bloqueo meiótico durante la vida embrionaria.
En el ovario fetal se forma una delgada túnica albugínea, casi imperceptible, en la unión corticomedular. La corteza del ovario es el componente dominante y contiene la mayoría de los oocitos. La medula llena con tejido conectivo y vasos sanguíneos que proceden del mesonefros. El testículo, por otra parte , se caracteriza por el predomino del componente medular situado bajo una evidente túnica albugínea.
El ovario en desarrollo no mantiene una relación con el mesonefros. Normalmente, los túbulos mesonéfricos del embrión femenino se degeneran, y solo dejan pocos restos (tabla 15-1).
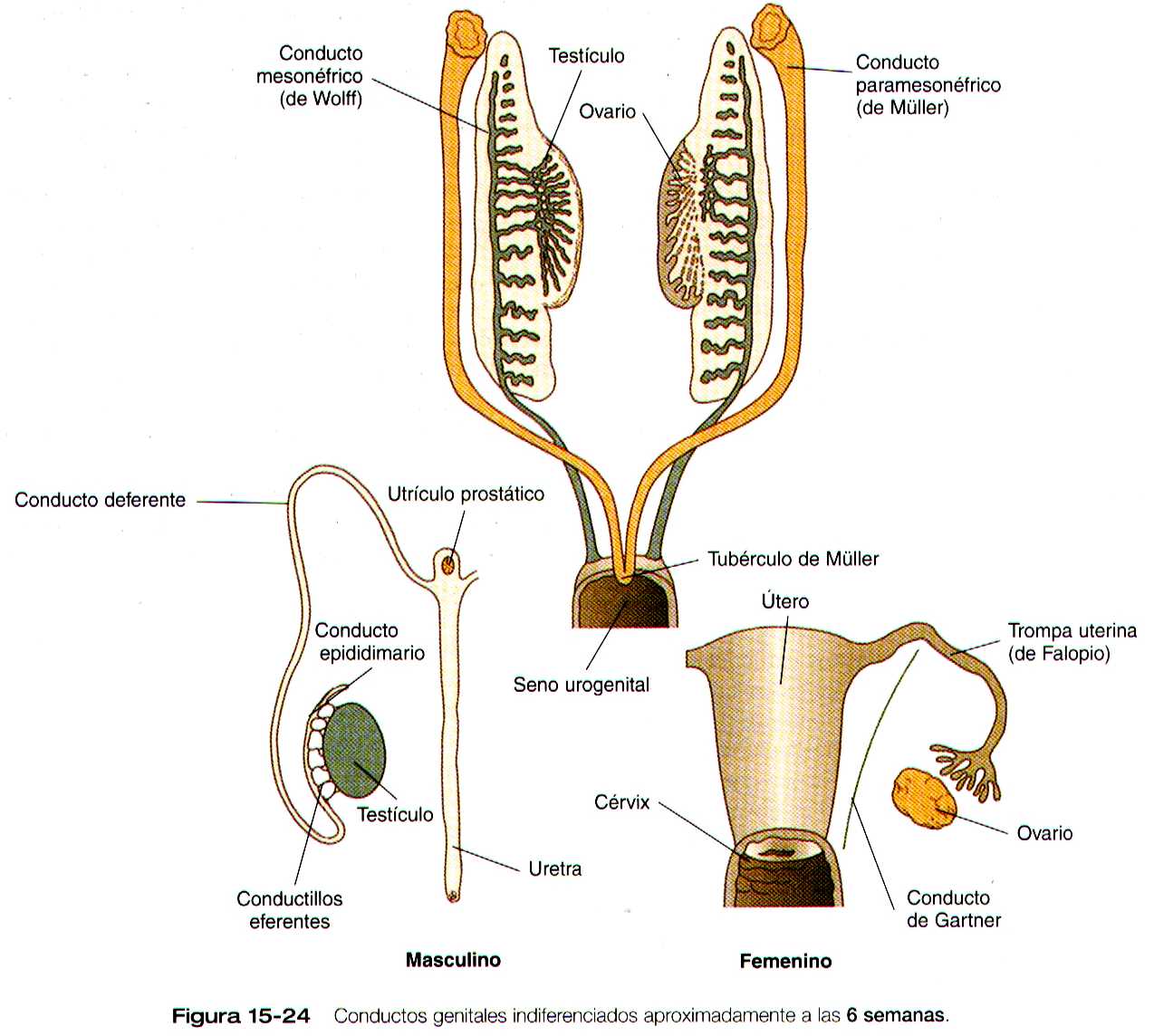
Fase indiferenciada del sistema de conductos sexuales
El sistema indiferente de conductos sexuales consta de los conductos mesonéfricos (de Wolff) y los conductos paramesonefricos (de Müller) (figura 15-24). Los conductos paramesonefricos aparecen entre los 44 y los 48 días de gestación como invaginaciones longitudinales del epitelio de superficie a lo largo de la cresta mesonefrica lateral a los conductos mesonéfricos. Las investigaciones pronto se convierten en cordones epiteliales, que crecen en dirección caudal y terminan en el seno urogenital entre los extremos de los conductos mesonéfricos sin abrirse en el seno. A continuación, estos cordones adquieren luz en dirección cráneo caudal. El extremo craneal de cada conducto paramesonefrico se abre a la cavidad celómica como una estructura parecida a un embudo. El destino de los conductos genitales indifenciadas depende del genero de la gónada.
Sistema de conductos sexuales masculinos
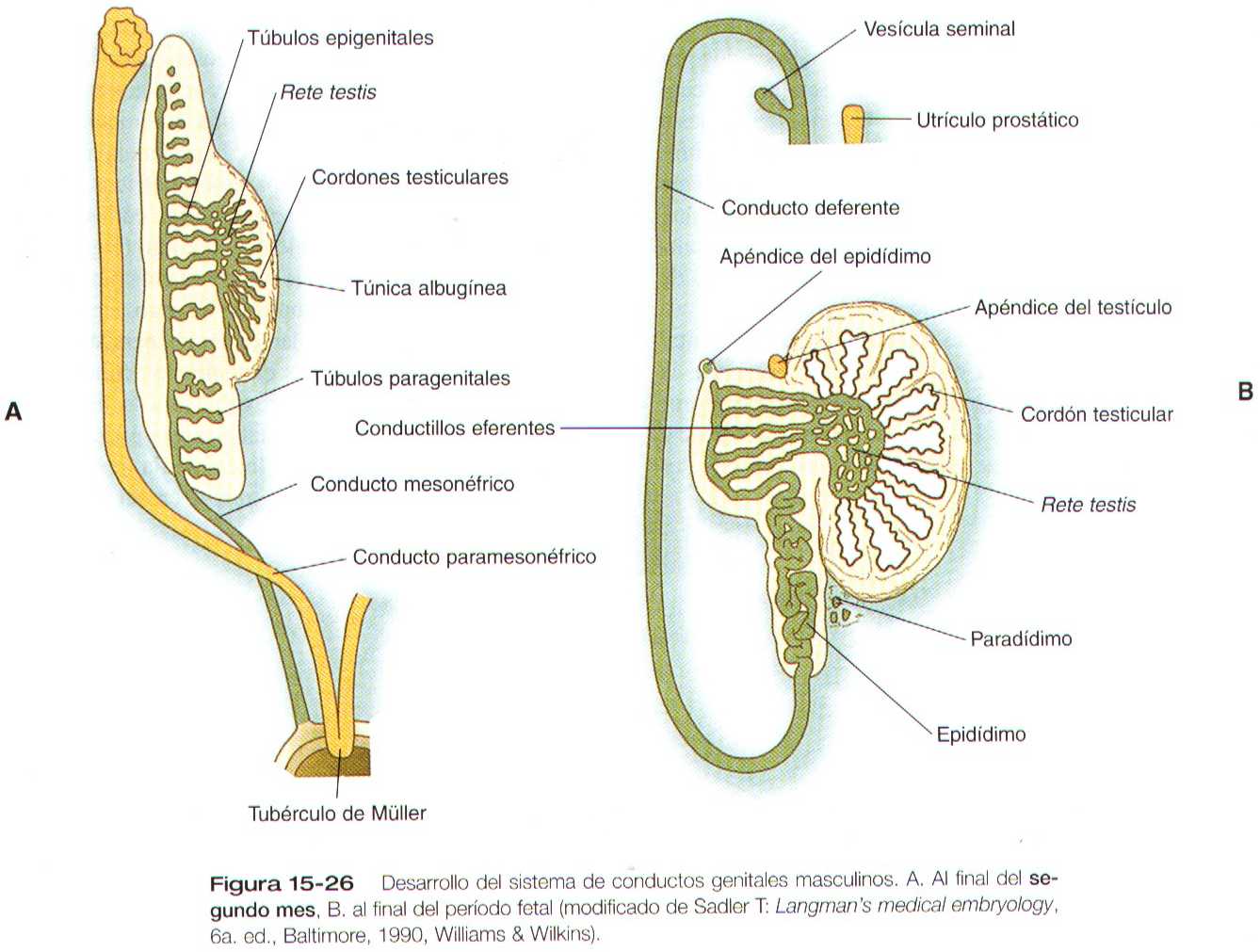
El desarrollo del sistema de conductos sexuales en el varón depende de la secreción del testículo. Bajo la influencia de la sustancia antimülleriana, una glucoproteina de la familia del factor B de transformación del crecimiento segregada por las células de Sertoli del testículo a las ocho semanas de gestación, los conductos paramesonefricos degeneran y solo quedan unos restos de sus extremos caudales y craneales (figuras 15-1 y 15-26 y tabla 15-1 no está). La sustancia antimülleriana no parece actuar directamente sobre el epitelio de los conductos paramesonefricos sino que actúan sobre el mesénquima que los rodea. Estas células mesenquimatosas expresan un gen que codifica un receptor fijo de membrana de serina/treonincinasa que fija la sustancia inhibitoria antimülleriana. Bajo su influencia, las células del mesénquima circundante instruyen a las células epiteliales del conducto de Müller para que involucionen.
Debido a la acción de la testosterona, que es producida por las células de Leydig de los testículos, los conductos mesonéfricos continúan su desarrollo aunque los riñones mesonéfricos estén degenerando. Los conductos mesonéfricos se convierten en los conductos deferentes, que constituyen la vía de paso para el transporte de los espermatozoides desde los testículos hasta la uretra. Pueden persistir porciones de los testículos mesonéfricos en degeneración cerca del testículo formando el paradidimo.
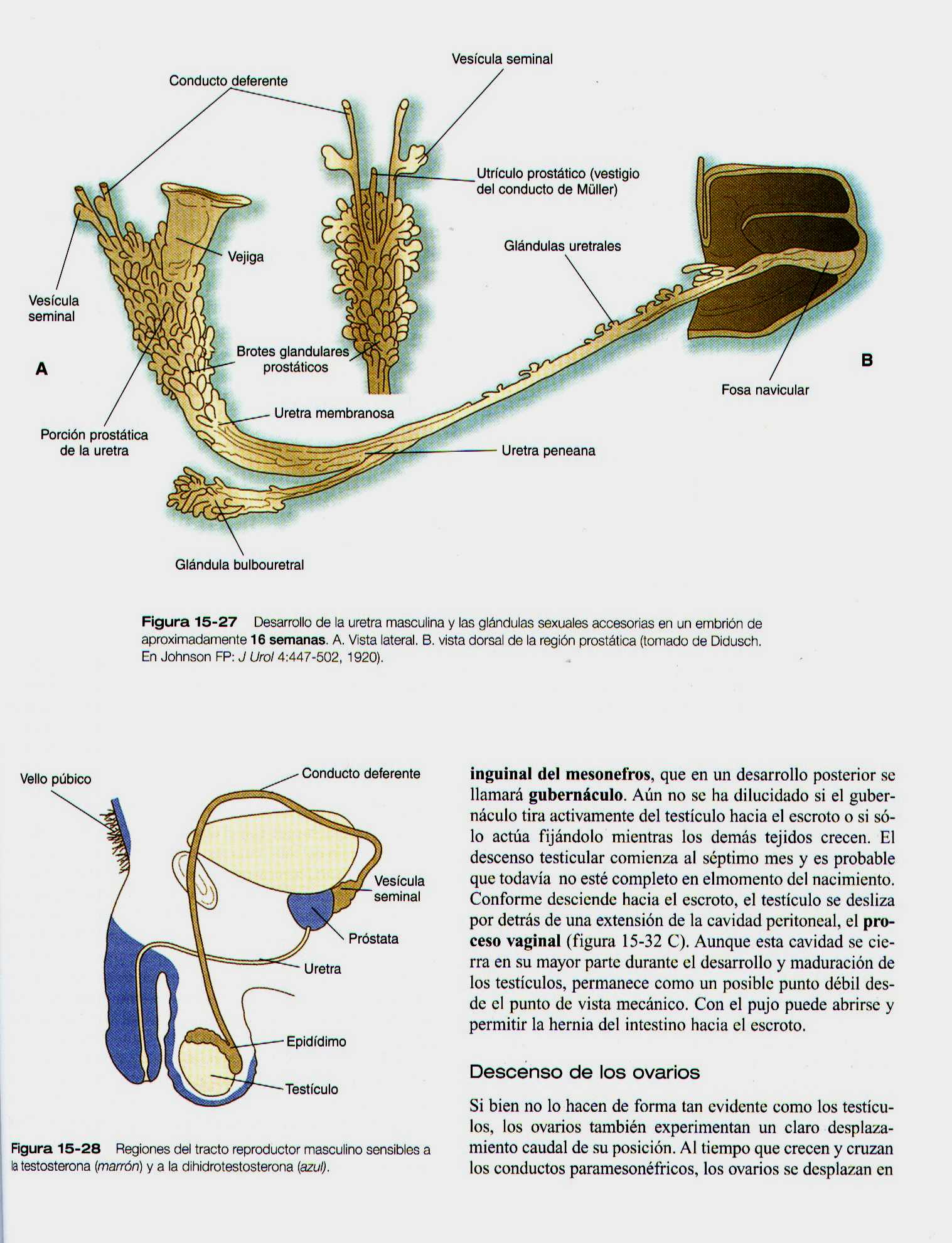
La formación de las glándulas sexuales accesorias las vesículas seminales, la próstata y las glándulas bulboureterales (figura 15-27) esta asociada con el desarrollo del sistema de conductos genitales masculinos (tanto el conducto deferente como la uretra). Estas glándulas surgen como evaginaciones epiteliales que se desprenden de sus respectivos conductos (las vesículas seminales del conducto deferente y las otras de la uretra), y su formación implica interacciones epitelio-mesénquima similares a las de otras glándulas. Además, estas glándulas dependen de la estimulación androgénica para su desarrollo. En las células mesenquimatosas aparecen receptores a los andrógenos que parecen ser las dianas primarias de las hormonas andrógenas circulantes (en esta etapa las células epiteliales no tiene receptores de andrógenos). Después de ser estimuladas por los andrógenos las células mesenquimatosas actúan sobre el epitelio asociado mediante un efecto paracrino local de los factores de crecimiento, que hacen que se diferencien con sus características glandulares específicas.
En la próstata en desarrollo el mesénquima urogenital actúa sobre los brotes epiteliales del endodermo del seno urogenital situados justo por debajo de la vejiga. El mesénquima induce la formación de conductos epiteliales, regula la proliferación del epitelio y la expresión de los receptores de andrógenos en este, y estimula la síntesis de proteínas especificas de la próstata. A la inversa, el epitelio prostático en desarrollo induce al mesénquima que los rodea a que se diferencie en células musculares lisas.
En experimentos de recombinación de tejidos en los que se combino mesodermo glandular de ratones afectados por el síndrome de feminización testicular (ausencia de receptores de testosterona cuyo resultado es la falta de respuesta a esta hormona ) con epitelio normal, no se produjo la diferenciación del epitelio; se demostró así que el componente mesodérmico del primordio glandular es la diana hormonal. Al contrario, cuando se combino mesodermo glandular normal con epitelio de animales afectados por el síndrome de feminización testicular, el desarrollo fue normal.
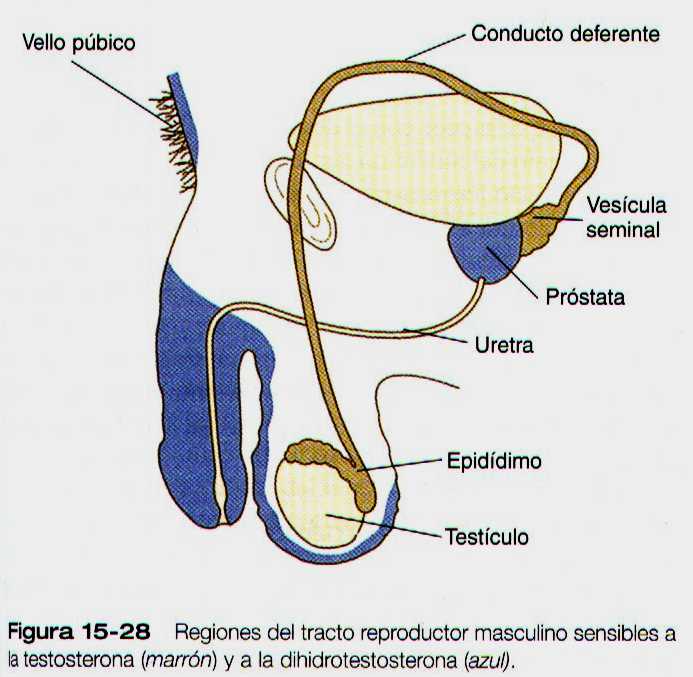
En el embrión los tejidos que rodean al seno urogenital sintetizan una enzima (5-alfa- reductasa) que convierte la testosterona en dihidrotestosterona. Mediante los receptores apropiados de cualquiera de las formas de las testosterona, estas actúan y diferencian los tejidos del tracto reproductor masculino (figura 15-28).
Sistema de conductos sexuales femeninos
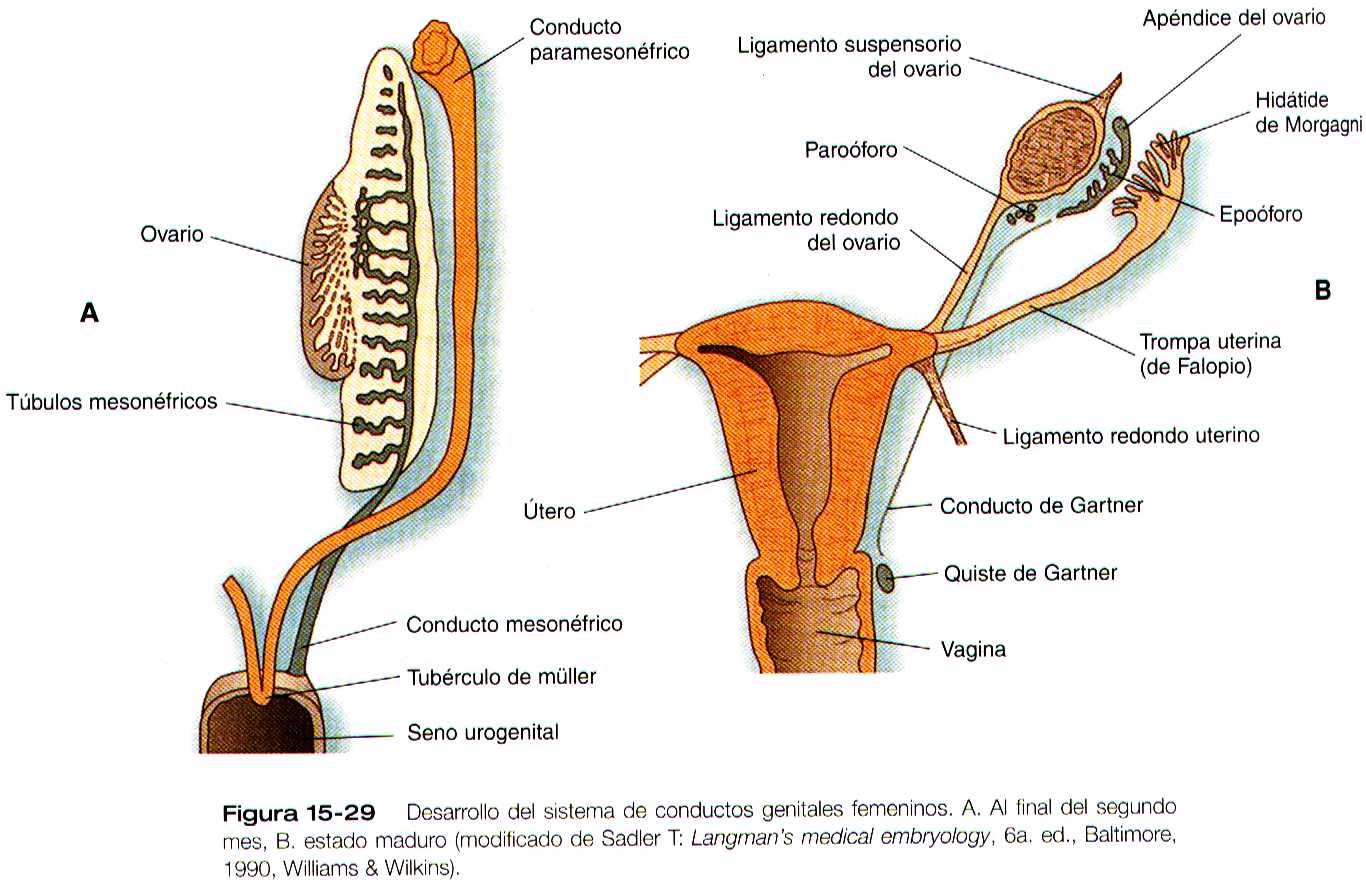
Si hay ovarios presentes o si las gónadas están ausentes o si son disgénicas, el sistema de conductos sexuales se diferencia hacia el fenotipo femenino. En ausencia de la testosterona producida por los testículos, los conductos mesonéfricos involucionan y solo dejan vestigios rudimentarios (tabla 15-1 no está). En contraste, la ausencia de la sustancia antimülleriana permite que los conductos paramesonéfricos (de Müller) continúen desarrollándose y se conviertan en las principales estructuras del tracto reproductor femenino (figura 15-29).
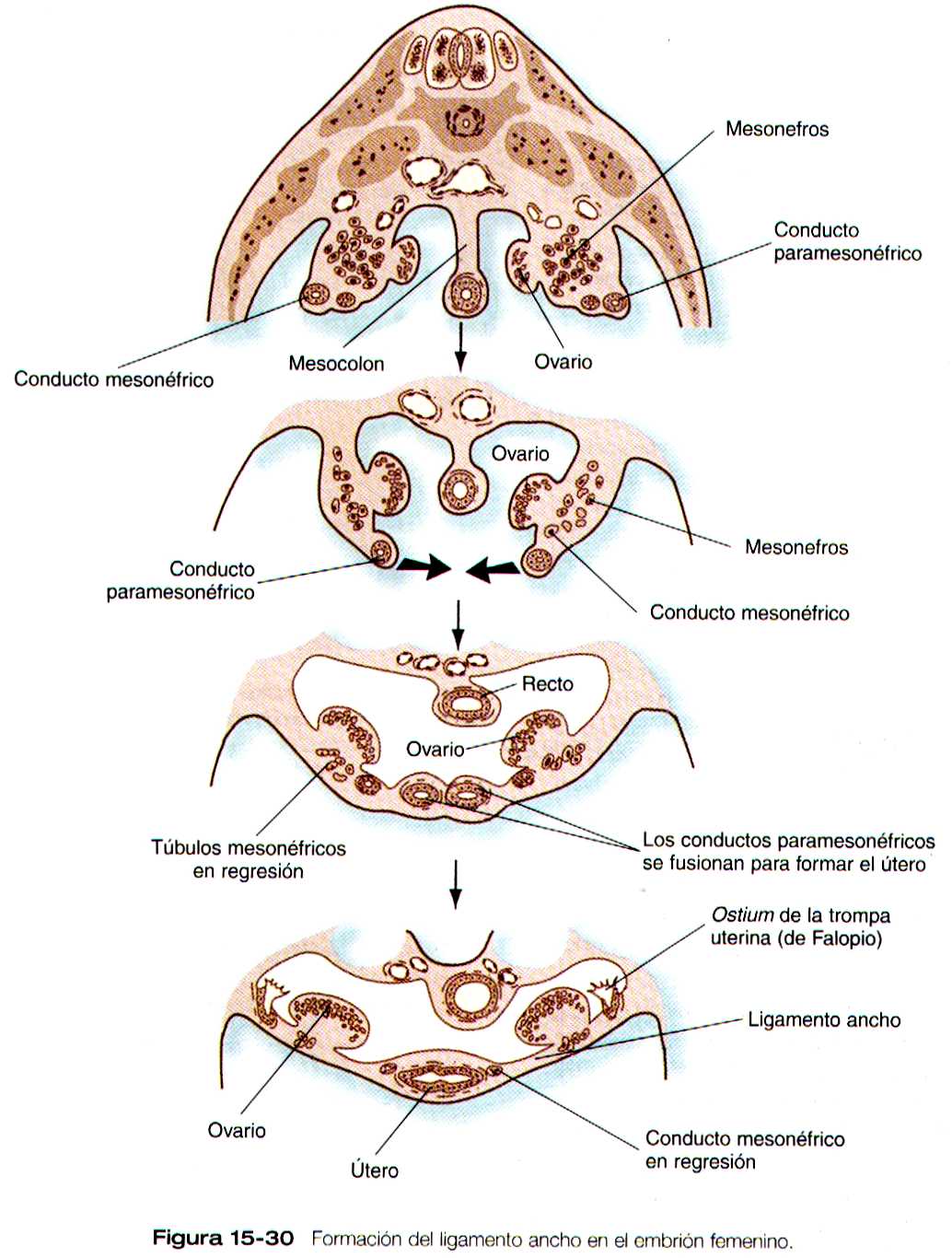
Las porciones craneales de los conductos paramesonéfricos se convierten en las trompas de Falopio, cuyos orificios craneales abiertos en la cavidad celómica persisten como las fimbrias. Hacia sus extremos caudales a la línea media cruzan ventralmente a los conductos mesonéfricos. Este cruce y y la posterior fusión en la línea media tiene sus origen en el desplazamiento medial de toda la cresta urogenital (figura 15-30). La zona de fusión de los conductos paramesonefricos en la línea media da lugar al útero, y el tejido de la cresta que se desplaza con ellos forma el ligamento redondo del útero.
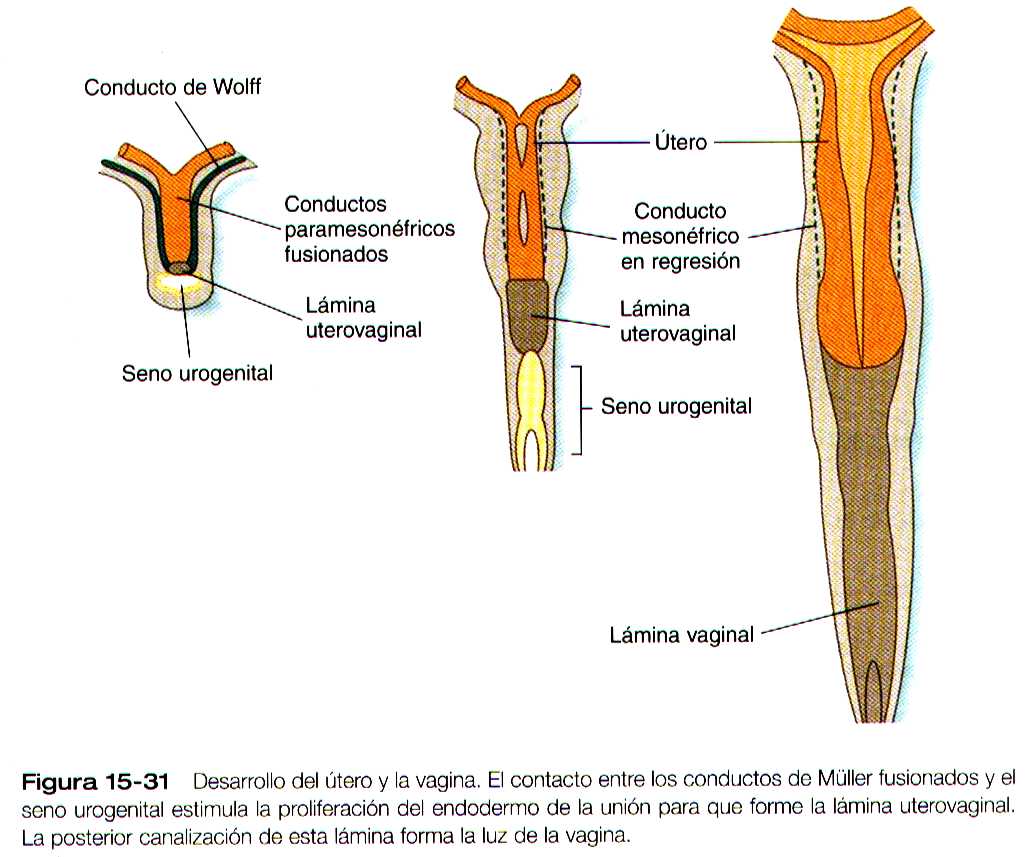
La formación de la vagina no está aclarada, y se han propuesto varias explicaciones para su origen. Una hipótesis muy común, plantea que los conductos paramesonéfricos al fusionarse dan lugar al tercio superior de la vagina, y el tejido epitelial del tubérculo de Müller (lamina útero vaginal) se ahueca para formar los dos tercios inferiores (figura 15-31). En época mas reciente, varias investigaciones han sugerido que las porciones mas caudales de los conductos mesonéfricos participan en la formación de la vagina ya sea mediante una contribución directa de las células a sus pares, o mediante una acción inductiva sobre el tejido paramesonefrico. El desarrollo completo del tracto reproductor femenino depende de las hormonas estrogénicas segregadas por los ovarios fetales.
Descenso de las gónadas
Descenso de los testículos
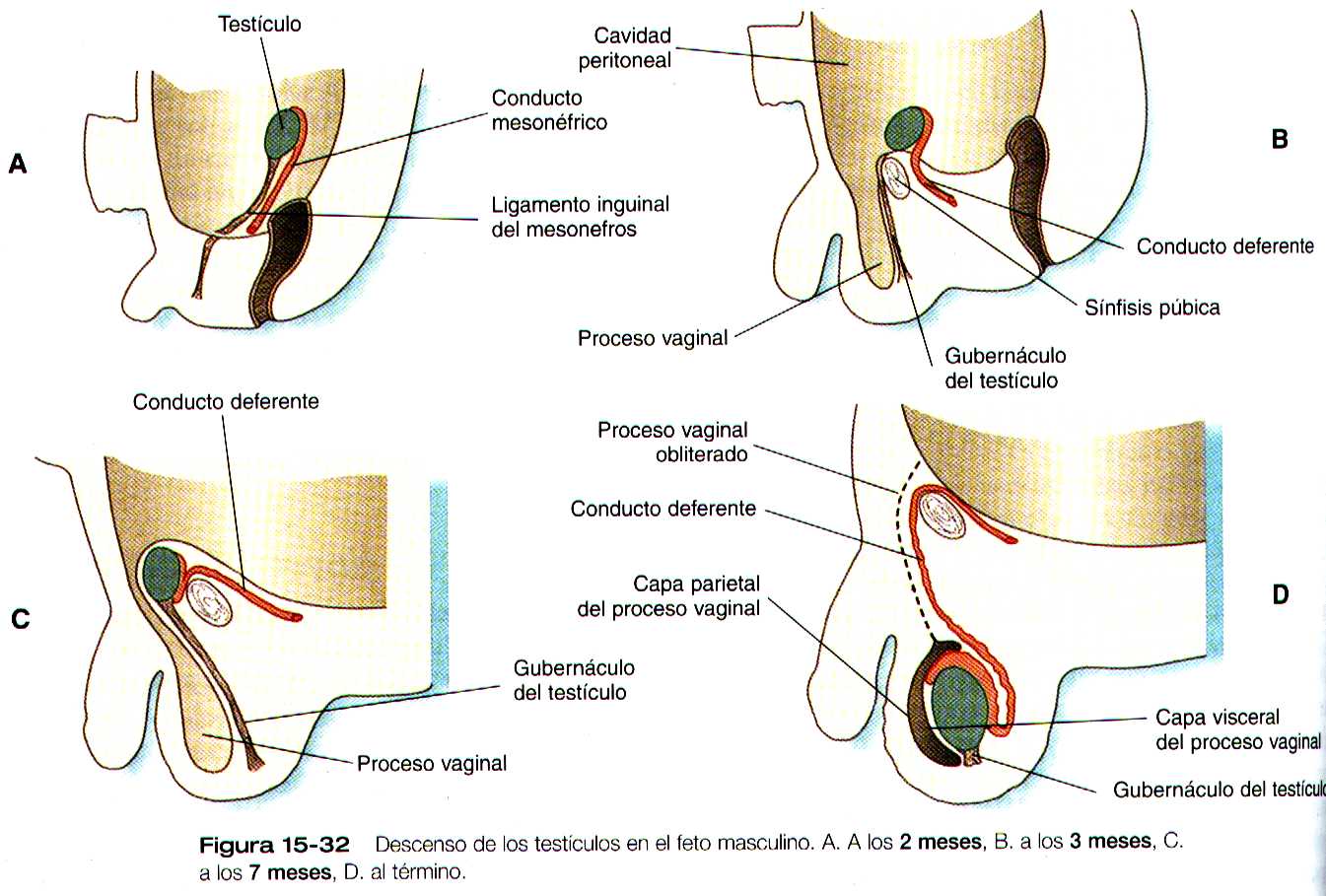
Los testículos no permanecen en su lugar de origen migran desde se localización intrabdominal hacia el escroto (figura 15-32). A semejanza de los riñones, los testículos son estructuras retroperitoneales y su descenso se produce por detrás del epitelio peritoneal.
El descenso testicular se ha dividido en tres fases : la primera esta asociada con el crecimiento de los testículos y la regresión concomitante de los riñones mesonéfricos. Esto causa algo de desplazamiento caudal de los testículos. La segunda fase, que suele denominarse descenso transabdominal, lleva los testículos por debajo del nivel del anillo inguinal pero no hasta el escroto. El control de esta fase se ha distribuido a la sustancia antimülleriana y a la regresión de los conductos paramesonefricos. Esta fase implica tanto la acción de la testosterona como la dirección del ligamento inguinal del mesonefros, que en un desarrollo posterior se llamara gubernáculum (gubernáculo).
Aun no se ha dilucidado si el gubernáculo tira activamente del testículo hacia el escroto o si solo actúa fijándolo mientras los demás tejido crecen. El descenso testicular comienza al séptimo mes y es probable que todavía no este completo en el momento del nacimiento. Conforme desciende al escroto, el testículo se desliza por detrás de una extensión de la cavidad peritoneal, el proceso vaginal (figura 15-32 C). Aunque esta cavidad se cierre en su mayor parte durante el desarrollo y maduración de los testículos, permanece como un posible punto débil desde el punto de vista mecánico. Con el pujo puede abrirse y permitir la hernia del intestino hacia el escroto.
Descenso de los ovarios
Si bien no lo hacen de forma tan evidente como los testículos, los ovarios también experimentan un claro desplazamiento caudal desde su posición. Al tiempo que crecen y cruzan los conductos paramesonéfricos, los ovarios se desplazan en dirección caudal y lateral.
Su posición es mantenida por dos ligamentos, que son restos de estructuras asociadas con el mesonefros. En dirección craneal se encuentra el ligamento diafragmático del mesonefros, que se convierten en el ligamento suspensorio del ovario (infundíbulo-pélvico). La porción superior del ligamento inguinal (a la cual algunos autores llaman ligamento gonadal caudal) se convierte en el ligamento redondo del ovario (útero-ovárico), y la porción inferior del ligamento inguinal se transforma en el ligamento redondo del útero ( figura 15-29). Los extremos mas caudales de los ligamentos redondos del útero terminan confundidos con el tejido conectivo denso de la fascia de los labios mayores.
GENITALES EXTERNOS
Etapa indiferenciada
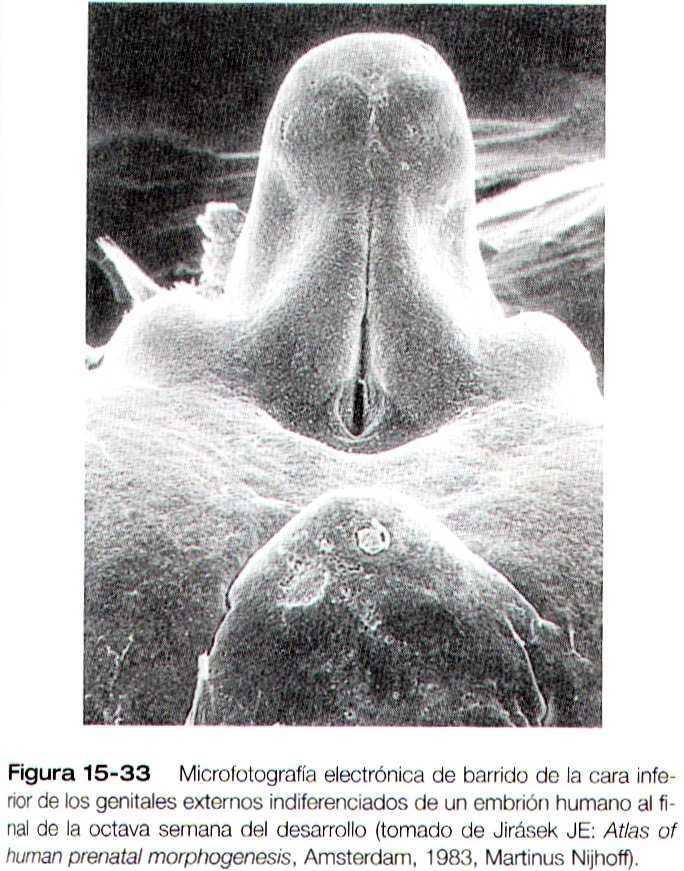
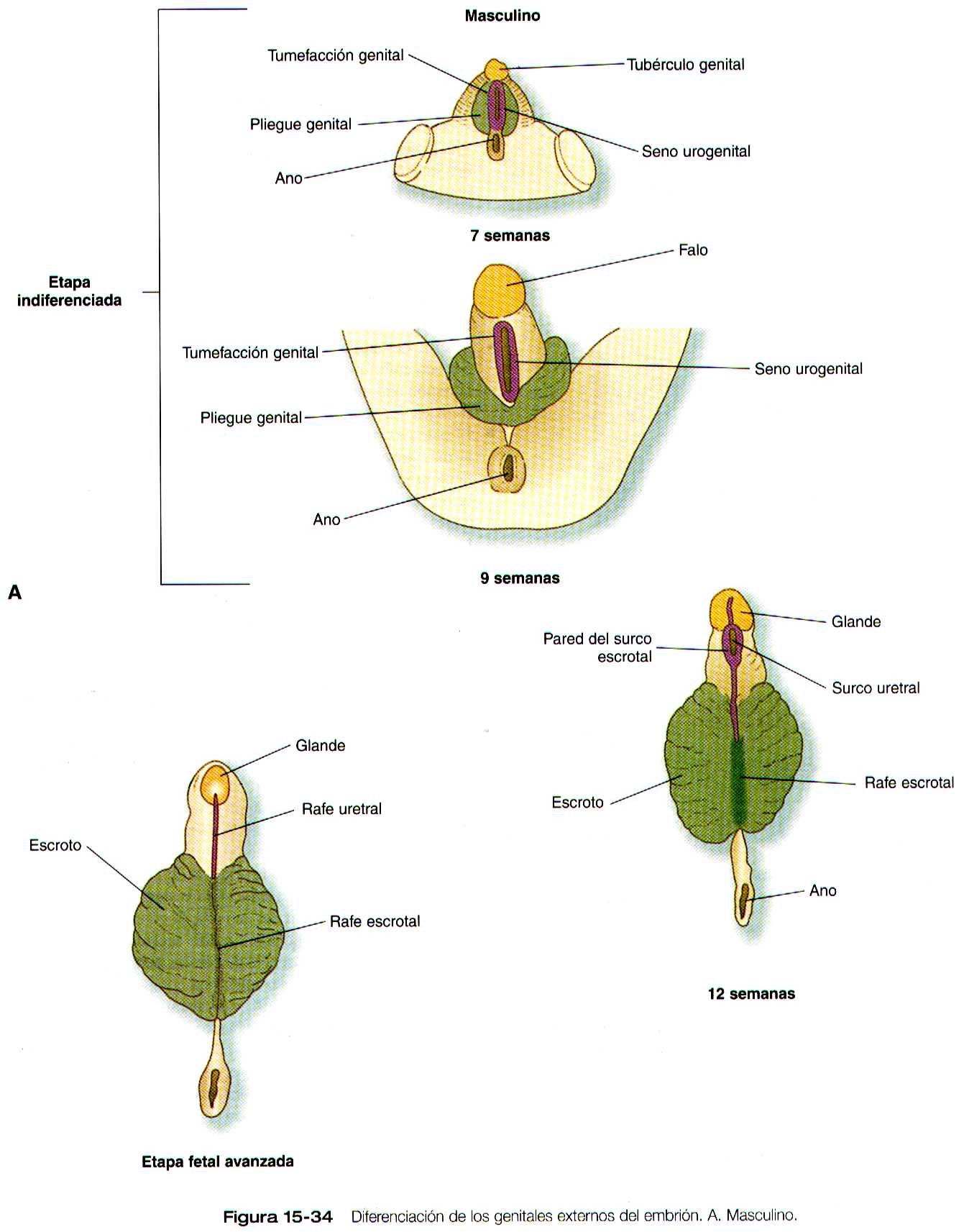
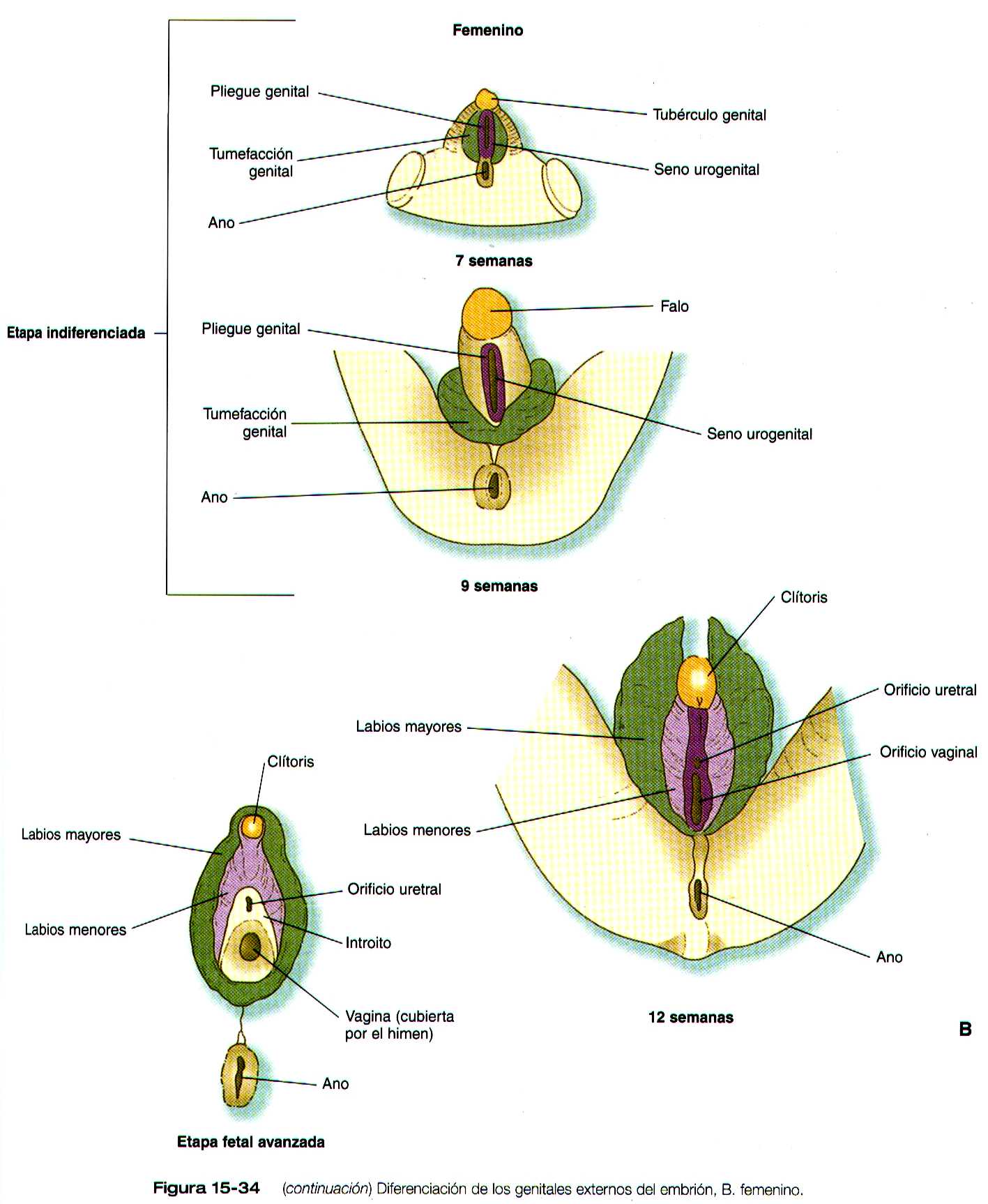
Los genitales externos se derivan de un complejo de tejido mesodérmico localizado alrededor de la cloaca. Precozmente, por delante de la depresión del proctodeo y en la línea media se forma una elevación denominada eminencia genital. Esta estructura pronto se transforma en el predomínante (denominado) tubérculo genital (figura 15-33), que esta flanqueado por un par de pliegues genitales que se extienden hacia el proctodeo. En situación algo lateral a estos se encuentran el par de engrosamientos genitales (figura 15-34a y b). Cuando la membrana genital se rompe durante la octava semana, el seno urogenital se abre directamente al exterior entre los pliegues genitales. Estas estructuras, que son prácticamente idénticas en los embriones masculinos y femeninos durante la etapa indiferenciada, constituye la base para el desarrollo de los genitales externos.
Al igual que sucede con el desarrollo de las extremidades, el crecimiento del tubérculo genital depende de una continua interacción ectodérmica-mesodérmica, aunque no existe una cresta ectodérmica apical en el tubérculo genital. En otra similitud con la yema de la extremidad, existe un gradiente de productos del gen Hoxd en el tubérculo genital y en los conductos genitales de los ratones; se supone que esto también ocurre en los seres humanos. Por ultimo, fragmentos de mesénquima de tubérculo genital de ratones muestran actividad polarizante cuando se injertan en yemas de la extremidad de embriones de pollo (capitulo 9).
Genitales externos masculinos
Debido a la acción de la dihidrotestosterona (figura 15-28), el tubérculo genital experimenta un considerable aumento de tamaño para formar el pene y las tumefacciones genitales se agrandan para dar lugar a las bolsas escrotales (figura 15-34 A ).
Conforme se produce este crecimiento, el seno urogenital establece continuidad con el surco que se ha desarrollado a lo largo de la cara caudal del tubérculo genital. Este surco se cierra y se convierte en la parte peneana de la uretra, y el seno urogenital cerrado se transforma en la porción prostática de la uretra. En la parte mas distal del pene un cordón sólido de células epiteliales crece y se evagina a partir del glande para encontrarse con la uretra peneana. Cuando se canaliza, se completa la formación de la uretra masculina. La línea de fusión a lo largo de la uretra y que pasa por las tumefacciones escrotales se conoce como rafe mediano.
Genitales externos femeninos
En las mujeres el patrón de los genitales externos es similar al de la etapa indiferenciada (figura 15-34 B ). El tubérculo genital se convierte en el clítoris, los pliegues genitales dan lugar a los labios menores, y las tumefacciones genitales se transforman en los labios mayores.
El seno urogenital permanece abierto y forma el intróito o vestíbulo, en el cual desembocan la uretra y la vagina. La uretra femenina, que se desarrolla desde la parte mas craneal del seno urogenital, equivale a la uretra prostática del varón.
El sistema urogenital se origina en el mesodermo intermedio.
El sistema urinario surge antes del desarrollo gonadal.
El desarrollo de los riñones comienza con la formación de pares de nefrotomas que conectan con un par de conductos néfricos. En dirección mas caudal a los nefrotomas se desarrollan pares de túbulos mesonéfricos en una secuencia cráneo caudal y se conectan con los conductos néfricos primarios, que se conocen como conductos mesonéfricos. En la parte caudal de cada conducto mesonéfrico crece una yema ureteral que induce al mesodermo circundante a trasformarse en metanefros.
En el interior del metanefros en desarrollo, las nefronas (unidades funcionales de riñón) se forman a partir de tres fuentes: el blastema metanefrogénico, el divertículo metanefrogénico y las células endoteliales vasculares en crecimiento. Las nefronas continúan formándose durante toda la vida fetal; su inducción comprende inducciones reciprocas colectores (yema ureteral) y el mesodermo metanefrogénico. Muchas interacciones moleculares median estas inducciones.
Los riñones surgen en la cavidad pélvica y, durante el final del periodo embrionario y el comienzo del fetal se desplazan hacia la región abdominal, donde se asocian con las glándulas suprarrenales. La vejiga surge a partir de la base de la alantoides.
El sistema urinario esta expuesto a numerosas malformaciones, la mas grave de las cuales es la agenesia, que probablemente tiene origen en una inducción defectuosa en el embrión primitivo . La migración anómala puede dar como resultado los riñones pélvicos, otros a riñones ectópicos o el riñón en herradura. La enfermedad del riñón poliquistico se asocia con la presencia de quistes en otros órganos internos. El defecto de cierre de las alantoides da como resultado quistes, senos o fístulas del uraco.
La determinación del sexo comienza en el momento de la fecundación en virtud del aporte de un cromosoma X o Y que el espermatozoide hace al óvulo. El sexo del embrión es indiferenciado. A través de la acción de gen Sry, la gónada indiferente del varón se convierte en testículo. En ausencia de este gen, la gónada se transforma en ovario.
La diferenciación gonadal comienza después de la migración de las células germinales primordiales hacia el interior de las gónadas indiferenciadas. Bajo la influencia del producto del gen Sry (factor de determinación testicular), los testículos comienzan a diferenciarse. No se requiere la presencia de las células germinales para la diferenciación de los cordones testiculares. En el testículo embrionario las células de Leydig segregan testosterona y las de Sertoli producen la sustancia antimülleriana. En ausencia de expresión del gen Sry, la gónada se trasforma en ovario y contiene folículos. La diferenciación folicular ovárica no se produce en ausencia de las células germinales.
El sistema de los conductos sexuales esta compuesto por los conductos mesonéfricos (de Wolff) y los paramesonefricos (de Müller); al comienzo, es indiferenciado. En el varón la sustancia antimülleriana causa la regresión del sistema de conductos paramesonefricos, y la testosterona causa el posterior desarrollo del sistema de conductos mesonéfricos. En la mujer, los conductos mesonéfricos involucionan en ausencia de testosterona y los conductos paramesonefricos persisten si no hay sustancia antimülleriana.
En los varones, los conductos mesonéfricos dan origen al conducto deferente y a las glándulas sexuales masculinas accesorias. En las mujeres, los conductos paramesonefricos forman las trompas de Falopio, el útero y el tercio superior de la vagina.
Los testículos descienden desde la cavidad abdominal hacia el escroto al final del desarrollo. Los ovarios también se desplazan hacia una posición mas caudal. La falta de descenso de los testículos da como resultado la criptorquidia y se asocia con probable torsión, esterilidad y tumores testiculares malignos.
CORRELACION CLINICA 15-2a
Malformaciones del sistema genital
ANOMALIAS DE LA DIFERENCIACION SEXUAL
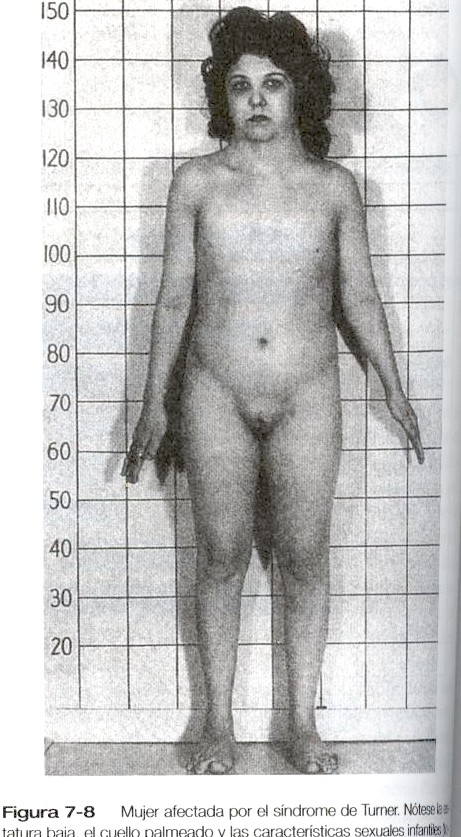
Síndrome de Turner (disgenesia gonadal)
El síndrome de Turner es el resultado de una anomalía cromosómica (45, XO) (Pág. 138). Los individuos afectados tiene células germinales primordiales que deberán involucionar al poco tiempo de llegar a las gónadas; en consecuencia no se produce la diferenciación de estos órganos, lo cual lleva a la formación de una cintilla gonadal. En ausencia de las hormonas gonadales los genitales se forman siguiendo la línea femenina, pero permanecen en estado infantil. El sistema de los conductos mesonéfricos involucionan a falta de estimulación androgénica.
Hermafroditismo verdadero
Los individuos con hermafroditismo verdadero, una afección muy rara, tienen tejido testicular y ovario. En los casos de mosaicismo genético, puede haber ovario y testículo presentes; en otros casos, los tejidos ovárico y testicular están presentes en la misma gónada (ovotestis). La mayoría de los hermafroditas verdaderos tiene una constitución cromosómica 46, XX, y los genitales externos son básicamente femeninos, aunque es característico que tengan un clítoris hipertrofico. Por los general a estos individuos los crían como mujeres.
Seudohermafroditismo masculino
Los seudohermafroditas masculinos son negativos para la cromatina sexual (46, XY). Dado que esta afección suele ser el resultado de la producción inadecuada de hormonas por los testículos fetales, el fenotipo puede variar. A menudo se asocia con hipoplasia del falo, y puede haber grados variables de persistencia de las estructuras del conducto paramesonefrico.
Síndrome de feminización testicular (insensibilidad a los andrógenos)
Los individuos afectados por este síndrome son varones desde el punto de vista genético (46, XY) y tienen testículos internos, pero es característico que tenga una apariencia fenotípica externa femenina y que sean criados como mujeres (figura 8-12). A menudo, la feminización testicular solo viene a descubrirse cuando la mujer busca tratamiento por la amenorrea o se le determina la cromatina sexual antes de competencias deportivas. Los testículos producen testosterona pero debido a la deficiencia de los receptores causada por una mutación del cromosoma X, la testosterona no puede actuar sobres los tejidos apropiados. puesto que la sustancia antimülleriana es un producto de los testículos, el útero y los dos tercios superiores de la vejiga están ausentes.
ESTRUCTURAS VESTIGIALES DE LOS CONDUCTOS GENITALES EMBRIONARIOS
Las estructuras vestigiales son los restos de la regresión de los conductos genitales embrionarios, que rara vez es completa. Son tan frecuentes que no siempre se las considera como malformaciones, aunque pueden volverse quísticas.
Restos de los conductos mesonéfricos
En los varones la persistencia del extremo craneal ciego del conducto mesonéfrico puede aparecer como el apéndice del epidídimo (figura 15-26). Los restos de pocos túbulos mesonéfricos es situación caudal a los conductillos eferentes aparecen en ocasiones como el paradídimo.
En las mujeres, los restos de la parte craneal del mesonefros pueden persistir como el Epooforo, o el paroóforo (figura 15-29). La parte caudal de conducto mesonéfrico se puede ver a menudo en los cortes histológicos a lo largo del útero o del tercio superior de la vagina como los conductos de Gartner. En ocasiones los restos de estos conductos se agrandan y forman quistes.
Restos del conducto paramesonéfrico
El extremo craneal puede persistir como un pequeño apéndice del testículo (figura 15-26). Los extremos caudales fusionados de los conductos paramesonéfricos se ven en la glándula prostática como una pequeña estructura situada en la línea media llamada utrículo prostático, que representa un rudimentario primordio del útero. En los neonatos de sexo masculino el utrículo suele estar un poco agrandado debido a la influencia de las hormonas estrogénicas maternas durante el embarazo, pero involuciona poco después del parto. Esta estructura puede aumentar de tamaño para formar una estructura similar al útero en algunos casos de seudohermafroditismo masculino.
OTRAS ANOMALIAS DEL SISTEMA DE CONDUCTOS GENITALES
Las anomalías del sistema de conductos mesonéfricos son relativamente raras, pero pueden presentarse duplicaciones o divertículos de conducto deferente. Hay una interesante relación de conducto deferente rudimentario o ausente en los varones afectados por fibrosis quística. Puede ser el resultado de un defecto en un gen situado a lo largo del gen causante de la fibrosis quística.
El síndrome del conducto mülleriano persistente, caracterizado por la formación de útero y trompas de Falopio, se ha descrito en algunos varones con fenotipo 46, XY. No existe una causa única para esta afección, y se han demostrado mutaciones de genes tanto para la sustancia antimülleriana como para su receptor.
CORRELACION CLINICA 15-2b
Malformaciones del sistema genital (continuación)
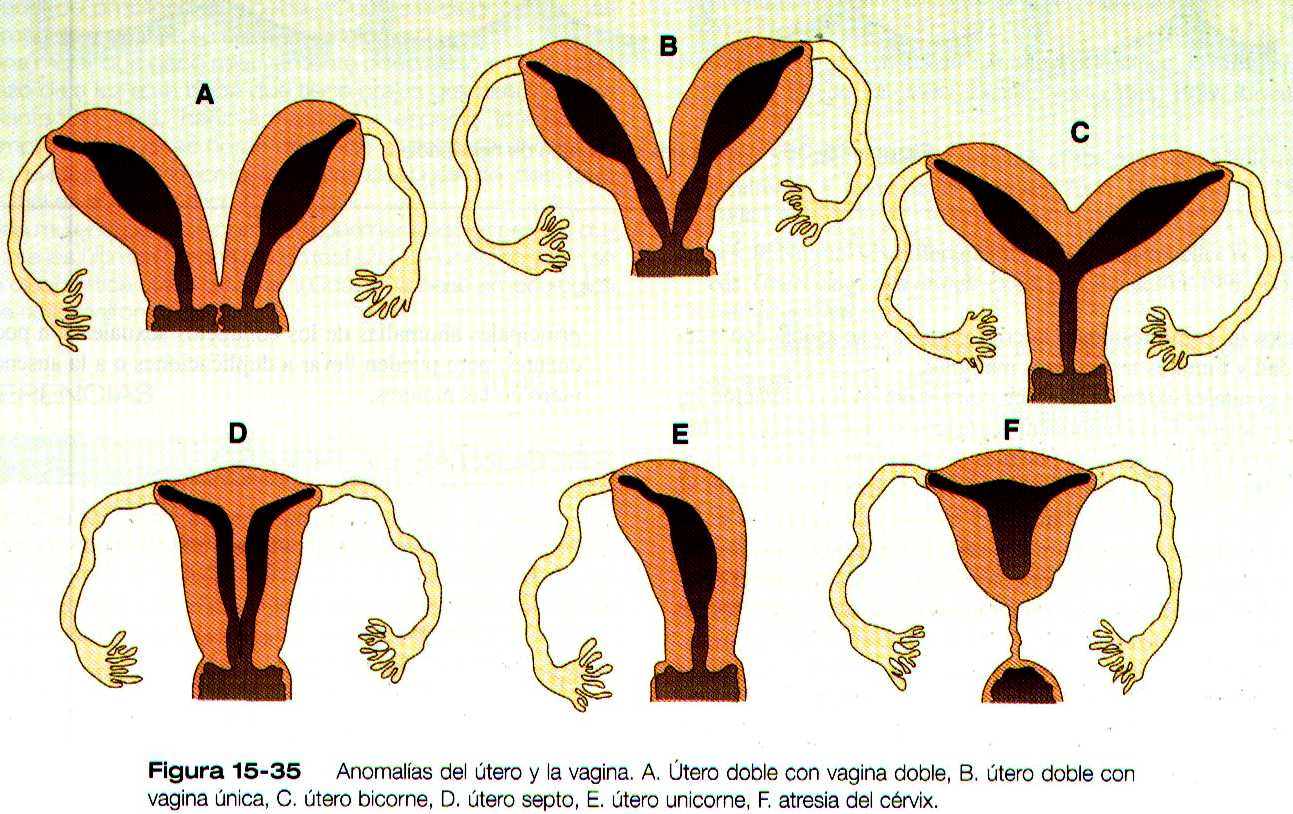
Las malformaciones del útero o de la vagina se atribuyen a anomalías de la fusión o de la regresión de los extremos caudales de los conductos paramesonéfricos (figura 15-35 ). Las anomalías uterinas van desde la existencia de un pequeño tabique que se extiende desde la pared dorsal del útero, hasta la duplicación completa del útero y el cervix. Se han registrado numerosos embarazos normales y con éxito en mujeres con úteros malformados. La agenesia de la vagina se ha atribuido a la ausencia de formación de la lamina vaginal epitelial en el lugar donde se unen el tubérculo de Müller y el seno urogenital.
ANOMALIAS DEL DESCENSO TESTICULAR
La falta de descenso testicular es frecuente en los varones prematuros y se observa en cerca del 3% de los varones a termino. Por lo general, los testículos de estos individuos descienden hacia el escroto pocos meses después del parto. Si no lo hacen, el resultado es la criptorquidia. Esta afección se acompaña de infertilidad porque la espermatogénesis no tiene lugar a la temperatura de la cavidad corporal. También existe una incidencia cincuenta veces mayor de tumores malignos en los testículos no descendidos.
Un testículo ocasionalmente puede migrar hacia un lugar distinto del escroto, incluidos el muslo, el perine y la pared ventral del abdomen. Debido a la elevada temperatura de los tejidos circundantes, el numero de espermatozoides viables que producen los testículos ectópicos es reducido.
Hernia inguinal congénita
Si el canal peritoneal que llega hasta el escroto fetal no se cierra, se presenta una malformación llamada persistencia del proceso vaginal. Este espacio puede llegar a ocuparlo el intestino que se hernia hacia el escroto.
MALFORMACIONES DE LOS GENITALES EXTERNOS
Varones
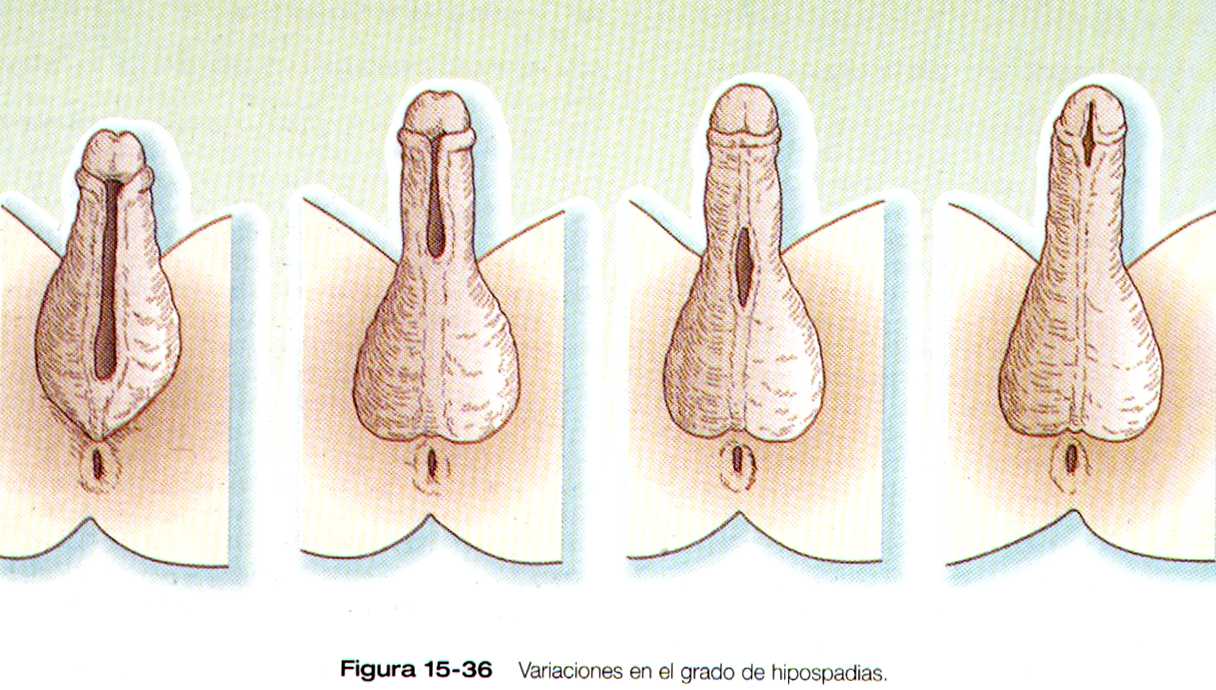
Las malformaciones mas frecuentes del pene es la Hipospadias, en la cual la uretra desemboca en la superficie ventral del pene en vez de en la punta del glande (figura 15-36 ). El grado de Hipospadias puede ir desde una leve desviación ventral del meato uretral hasta un orificio alargado que presenta la porción no fusionada del seno urogenital. En variedades mas graves el pene suele estar arqueado hacia abajo.
Los genitales externos también comienza en una situación indiferenciada. Los componentes básicos de los genitales externos son el tubérculo. Los pliegues y los rodetes genitales. Bajo la influencia de la dihidrotestosterona el tubérculo genital se alarga y se convierte en un falo y los pliegues genitales se fusionan para formar la uretra peneana. Los rodetes genitales dan lugar al escroto. En la mujer el tubérculo genital forma el clítoris, los pliegues dan lugar a los labios menores y los rodetes genitales a los labios mayores.
Si un individuo tiene solo un cromosoma X (XO) el resultado es el síndrome de Turner. Estas personas tiene fenotipo femenino con una estría gonadal. El hermafroditismo verdadero o el seudohermafroditismo pueden tener varias causas. La feminización testicular se encuentra en varones genotípicos que no tiene receptores de testosterona; su fenotipo es femenino. Las principales anomalías de los conductos sexuales son poco frecuentes, pero pueden llevar a duplicaciones o a la ausencia del útero en las mujeres.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}