Tema 9
DESARROLLO DE LAS EXTREMIDADES
COMIENZO DEL DESARROLLO DE LA EXTREMIDAD
PROPIEDADES REGULADORAS Y DETERMINACIÓN AXIAL
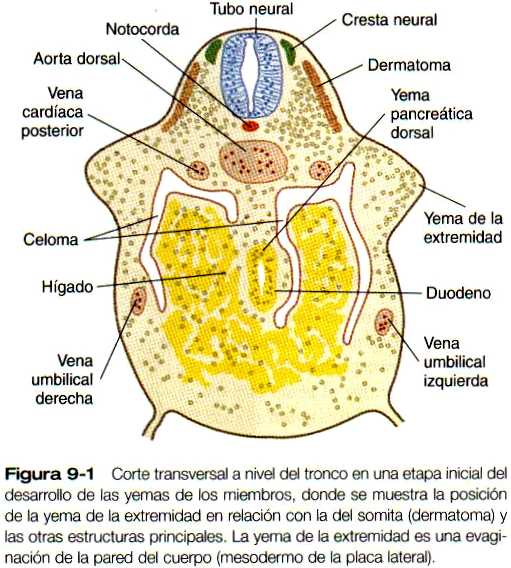
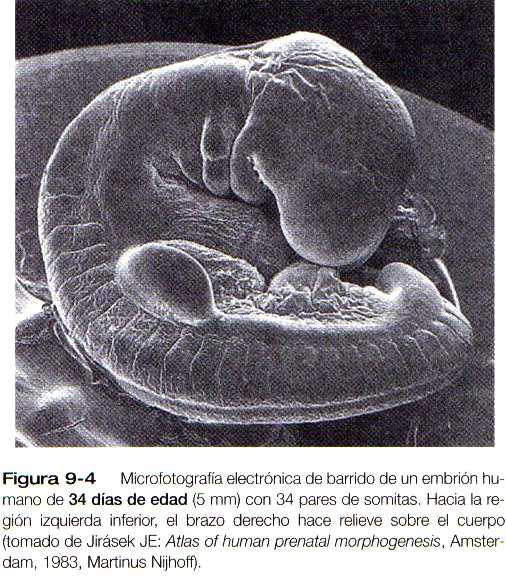
FORMACIÓN Y EVAGINACIÓN DE LA YEMA DE LA EXTREMIDAD
Mesodermo de la yema inicial del miembro
Interacciones mesodermo-ectodermo y función del mesodermo en la morfogénesis de las extremidades
Muerte celular y desarrollo de los dedos
Zona de actividad polarizante y señales morfogenéticas
DESARROLLO DE LOS TEJIDOS DE LAS EXTREMIDADES
Anomalías de las extremidades
PREGUNTAS DE REPASO
PROPIEDADES REGULADORAS Y DETERMINACIÓN AXIAL
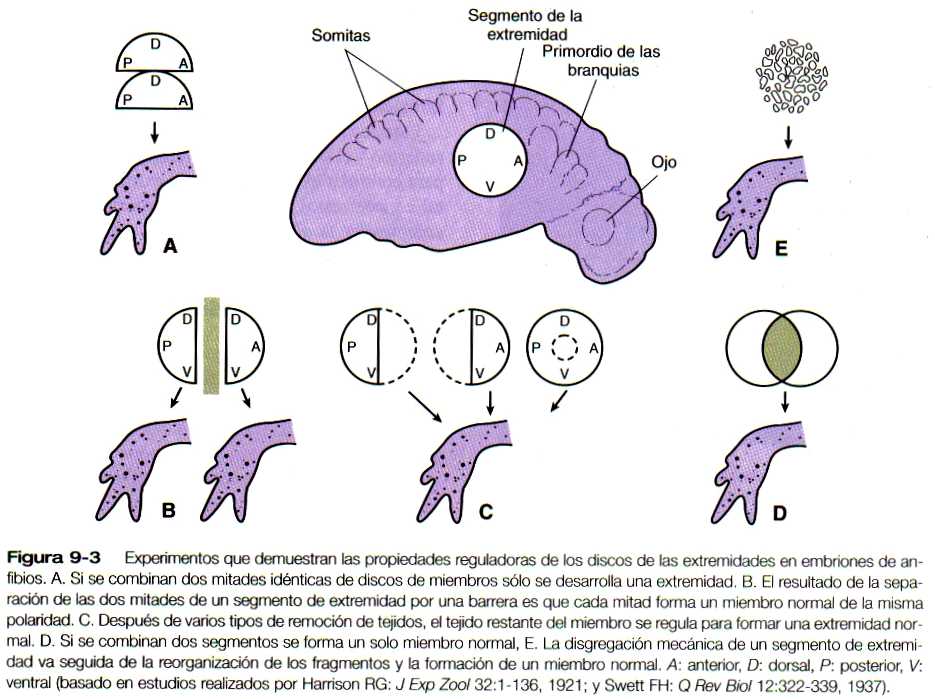
El primordio incipiente de la extremidad es un sistema de regulación, que tiene propiedades muy similares a las que se describieron para el embrión en segmentación (pág. 46). Estas propiedades pueden resumirse con los siguientes experimentos (figura 9-3):
1. Si se elimina una parte del primordio de una extremidad, el resto se reorganiza y forma la extremidad completa.
2. Si el primordio de una extremidad se divide en dos mitades y se impide que se fusionen, cada una da lugar a una extremidad completa (fenómeno de gemación).
3. Si se yuxtaponen dos mitades equivalentes de un primordio de extremidad, se forma un miembro completo.
4. Si se superponen dos segmentos equivalentes de extremidad, se reorganizan para formar un solo miembro (véase sección en Embriones tetraparentales, pág. 47).
* Debido a las diferentes utilizaciones de los nombres de los ejes, algunos embriólogos humanos estarían en desacuerdo con la terminología axial que se utiliza aquí. De acuerdo con la terminología estricta que se usa en la embriología humana, anterior significa "ventral", y posterior significa "dorsal". No obstante, la terminología axial que usamos en este capitulo (según la cual anterior significa "craneal" y posterior significa "caudal") se usa de forma tan uniforme en la bibliografía sobre embriología experimental y comparada de forma que el estudiante que consultase la bibliografía original sobre el desarrollo de los miembros encontraría muy contuso el empleo de la terminología axial humana.
FORMACIÓN Y EVAGINACIÓN DE LA YEMA DE LA EXTREMIDAD
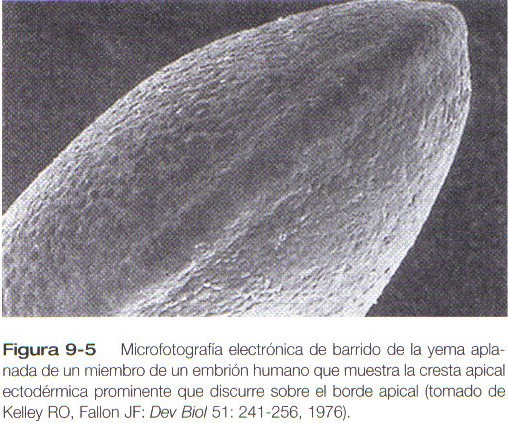
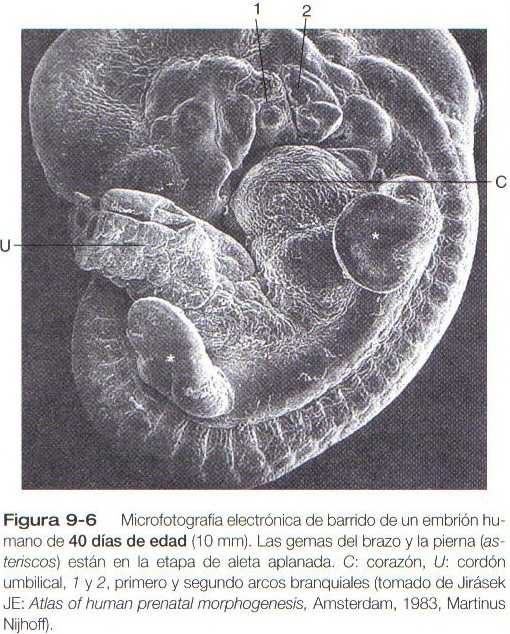
Una característica distinta es la presencia de una cresta de ectodermo engrosado (cresta apical ectodérmica) localizada a lo largo del plano anteroposterior del vértice de la yema del miembro (figura 9-5). Durante todo el tiempo en que la cresta apical ectodérmica está presente, las regiones de la yema a partir de las cuales se forman la mano y el pie tienen forma de aleta, con la cresta apical localizada a lo largo del borde de la misma (figura 9-6). Los experimentos han demostrado que la cresta ectodérmica apical interactúa con el mesodermo subyacente de la yema del miembro para promover el crecimiento de la extremidad en desarrollo. Otros aspectos del desarrollo del miembro, como la morfogénesis (el desarrollo de la forma), están bajo la dirección de la información contenida en el mesodermo.
Esta sección define muchas de las formas en que el mesodermo y el ectodermo de la yema del miembro interactúan para controlar el desarrollo de la extremidad. Es fundamental conocer estos mecanismos del desarrollo con el fin de comprender la génesis de diversas malformaciones de los miembros.
Cresta apical ectodérmica
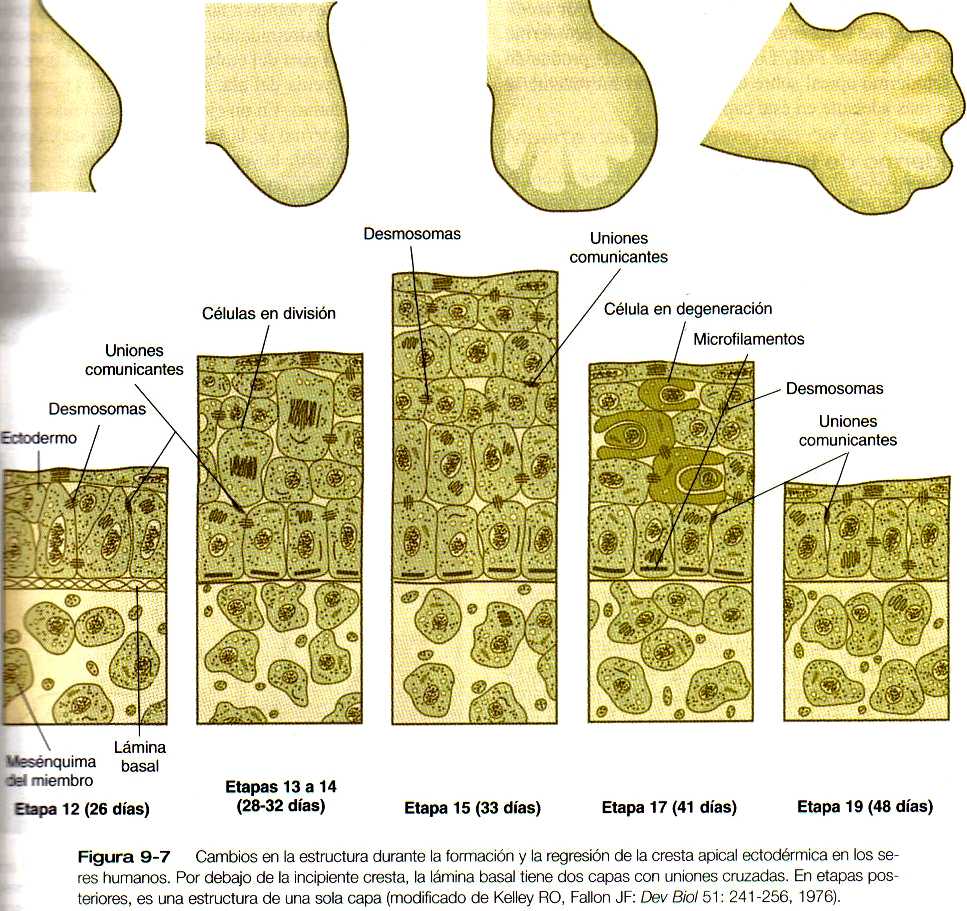
La cresta apical ectodérmica en los seres humanos es una formación epitelial de múltiples capas (figura 9-7) caracterizada por la presencia de numerosas uniones comunicantes (gap junctions) a través de las cuales las células se interconectan. Una lámina basal se interpone entre la cresta apical y las células del mesodermo subyacente.
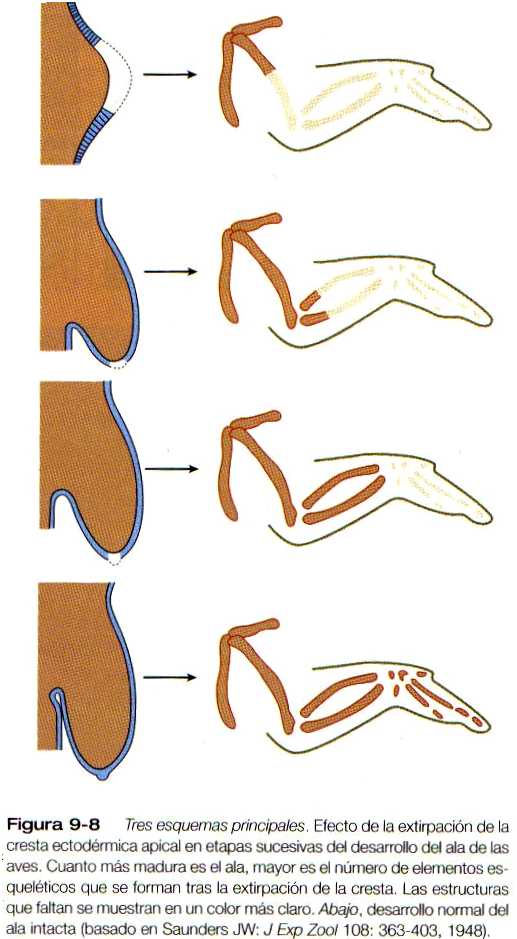
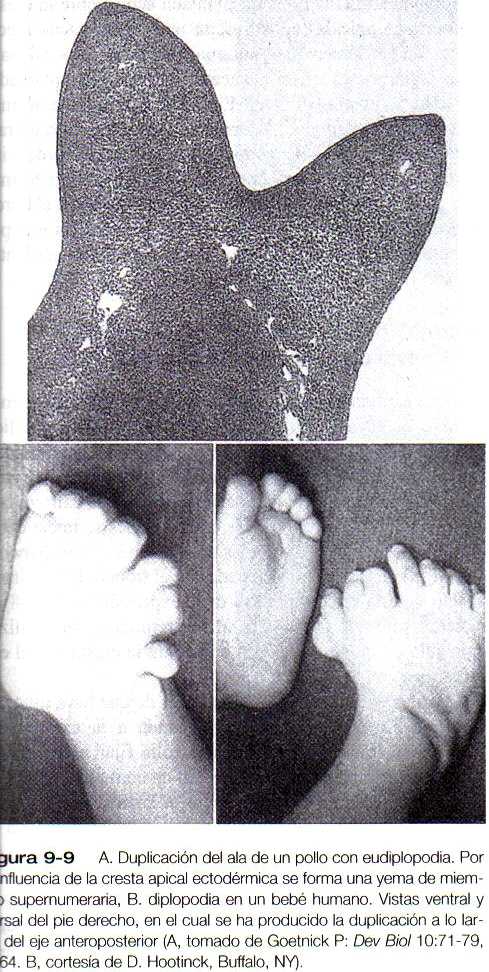
Aunque desde hace muchos años se ha conocido la morfología de la cresta apical ectodérmica, su papel en el desarrollo del miembro solo se comprendió cuando se la sometió a análisis experimental. La remoción de la cresta apical tiene como resultado una detención del desarrollo de la extremidad que lleva a que ésta quede bloqueada en su región distal (figura 9-8). En el pollo mutante amélico, el desarrollo inicial de los miembros es normal; más adelante la cresta apical ectodérmica desaparece y el desarrollo posterior del ala cesa. Si se coloca ectodermo del mutante sobre el mesodermo de la yema del ala de un embrión normal, el desarrollo del miembro se detiene, mientras que cuando se combina el mesodermo del imitante con ectodermo del embrión normal el resultado es el desarrollo de un miembro normal, lo que sugiere que el ectodermo es defectuoso en este mutante. Los estudios en el mutante amélico han demostrado que la cresta apical ectodérmica se forma en el punto de unión del ectodermo dorsal y el ventral y que en ausencia de yuxtaposición de ectodermo con propiedades dorsales y ventrales (por ejemplo, el mutante amélico), no puede mantenerse una cresta apical ectodérmica. Por el contrario, la presencia de una cresta apical ectodérmica adicional sobre la yema del miembro, ya sea debida a un trasplante experimental o como resultado de una mutación (por ejemplo, una diplopodia), da como resultado la formación de un miembro supernumerario.
Estudios recientes han demostrado que la señal de activación de la evaginación producida por la cresta apical ectodérmica es el FGF. En las primeras etapas de la formación del miembro, el ectodermo lateral comienza a producir FGF-8 a medida que se engruesa para formar la cresta apical ectodérmica. Conforme la yema del miembro empieza a crecer, la cresta apical también produce FGF-2 en toda su longitud y FGF-4 en su mitad posterior (figura 9-15). Si la cresta ectodérmica apical se extirpa, el crecimiento del mesodermo de la yema del miembro puede mantenerse con la aplicación local ya sea de FGF-2 o de FGF-4. Otros estudios han demostrado que en mutantes caracterizados por crecimiento deficiente o ausente de la extremidad, el ectodermo mutante no produce FGF. Los efectos del FGF producido por el ectodermo apical sobre el mesodermo subyacente se explican más adelante en este capitulo.
Mesodermo de la yema inicial del miembro
Estructura y composición
El mesodermo de la yema inicial de la extremidad está compuesto por células mesenquimatosas homogéneas irrigadas por una red vascular bien desarrollada. Las células mesenquimatosas están embebidas en una matriz formada por una malla laxa de fibras de colágeno y sustancia granular, en cuya composición entran grandes proporciones de ácido hialurónico y glucoproteínas. No hay nervios en la yema inicial de la extremidad.
No es posible distinguir los diferentes tipos celulares en el interior del mesénquima de la yema inicial sólo por su morfología. No obstante, están presentes células mesenquimatosas de diversas procedencias (figura 9-10). Al principio, el mesénquima de la yema de la extremidad está formado exclusivamente por células derivadas del mesodermo de la placa lateral. Estas células darán lugar al tejido esquelético, al tejido conectivo y algunos vasos sanguíneos. Las células mesenquimatosas derivadas de los somitas migran hacia la yema de la extremidad como precursores de las células musculares. Otra población de células migratorias es la de la cresta neural, que termina formando las células de Schwann de los nervios y las células pigmentarias (melanocitos).
Interacciones mesodermo-ectodermo y función del mesodermo en la morfogénesis de las extremidades
El desarrollo de las extremidades se produce como resultado de las continuas interacciones entre los componentes mesodérmico y ectodérmico de la yema. El ectodermo estimula el crecimiento de la yema del miembro porque promueve la mitosis e impide la diferenciación de las células del mesodermo distal situado debajo de él. Aunque la cresta apical promueve el crecimiento, su propia existencia está a su vez controlada por el mesodermo. Si se trasplanta una cresta apical de un miembro en desarrollo al mesodermo de una yema inicial del ala, la extremidad experimenta crecimiento normal hasta que se termina la morfogénesis. Sin embargo, si un mesodermo viejo de una extremidad en desarrollo se cubre con ectodermo apical de una yema joven, el desarrollo del miembro se detiene en el momento apropiado a la edad del mesodermo y no a la del ectodermo.
Se han efectuado experimentos de trasplantes recíprocos para demostrar que la forma general de la extremidad está determinada por el mesodermo y no por el ectodermo. Esto queda demostrado de manera evidente en los experimentos realizados en aves debido a las grandes diferencias que existen entre ambas extremidades. Por ejemplo, si el mesodermo de la yema de la pata del embrión del pollo se cubre con ectodermo de la yema del ala, se desarrolla una pata normal cubierta de escamas. En un ejemplo un poco más complejo, si se pone ectodermo de la yema de la pata de un pollo sobre el mesodermo de la yema del ala de un pato, se forma un ala de pato cubierta con plumas de pollo. Estos experimentos, que en ocasiones han implicado la realización de mosaicos de los componentes de las yemas de las extremidades de aves y mamíferos, muestran que la morfología general de la extremidad la determina el componente mesodérmico, no el ectodermo. Además, las características regionales de los derivados ectodérmicos (por ejemplo, pelo del cuero cabelludo en vez de pelo corporal en el caso de los mamíferos) también están bajo el dictado del mesodermo. Sin embargo, los experimentos de injertos cruzados entre especies muestran que la naturaleza de los derivados ectodérmicos formados (por ejemplo, pelo en vez de plumas) es la adecuada a la de la especie de la cual proviene el ectodermo.
La polidactilia es una afección caracterizada por la existencia de dedos supernumerarios y existe como mutante en las aves. Los experimentos de trasplantes recíprocos entre mesodermo y ectodermo han demostrado que el defecto es inherente al mesodermo y no al ectodermo. La polidactilia en seres humanos (figura 9-I I / 9_9) por lo general se hereda como un rasgo genético recesivo y es común encontrarla en poblaciones en las cuales la reserva genética total está relativamente restringida, como algunas comunidades Amish estadounidenses.
Muerte celular y desarrollo de los dedos
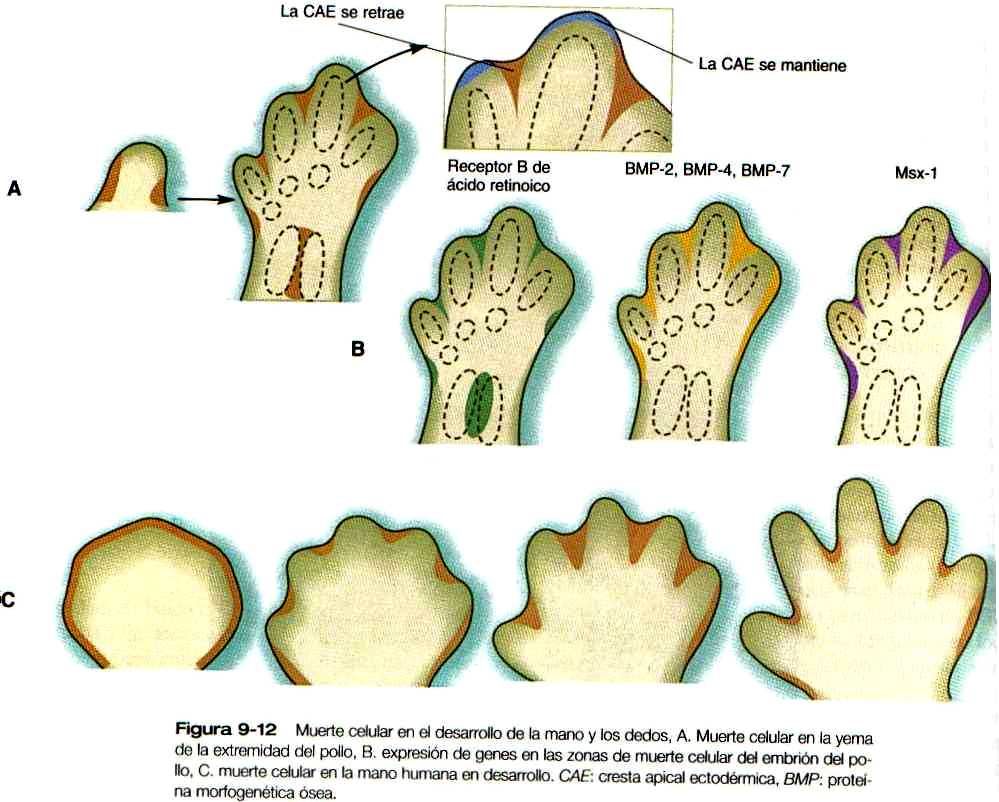
Aunque pueda parecer paradójico, la muerte celular programada genéticamente (apoptosis) es importante para el desarrollo de numerosas estructuras corporales. En el miembro superior se hace muy evidente en la futura región axilar, entre el radio y el cubito y en los espacios interdigitales (figura 9-12). Los experimentos en embriones de aves han mostrado que, hasta cierta etapa, las células mesodérmicas programadas para morir pueden no hacerlo si se las trasplanta a áreas en las cuales la muerte celular no suele suceder. No obstante, después de cierto tiempo, el "reloj de la muerte" se había fijado (un ejemplo de determinación) y ya no fue posible recuperar dichas células.
Conforme prosigue el desarrollo de la extremidad, aparecen cambios en la cresta ectodérmica apical. En vez de disponerse en continuidad alrededor de todo el vértice de la extremidad, la cresta comienza a fragmentarse, quedando segmentos de epitelio engrosado de la cresta en las zonas que cubren los radios digitales. Entre los dedos, la cresta regresa (figura 9-12 A). A medida que los primordios de los dedos continúan creciendo hacia fuera, la muerte celular esculpe los espacios interdigitales (figura 9-12 C). Numerosas moléculas importantes para el desarrollo, incluidas las proteínas morfogenéticas óseas 2 (BMP-2), BMP-4, y BMP-7. la Msx-1 y un receptor del ácido retinoico se expresan en las células mesodérmicas interdigitales. Aún no se conoce el mecanismo exacto de la muerte de las células interdigitales. pero están comenzando a aclararse algunos elementos del proceso. El adelgazamiento de la cresta apical ectodérmica que cubre las áreas interdigitales parece reducir la cantidad de FGF transmitida al mesénquima interdigital y sirve como un primer paso en el proceso de la apoptosis. Si se añaden FGF-2 o FGF-4 al mesénquima interdigital, se inhibe la muerte celular. Algunos investigadores afirman que las BMP, en especial la BMP-4, también forman parte de la señal que conduce a la muerte celular, no sólo en la región interdigital sino en otras partes del cuerpo.
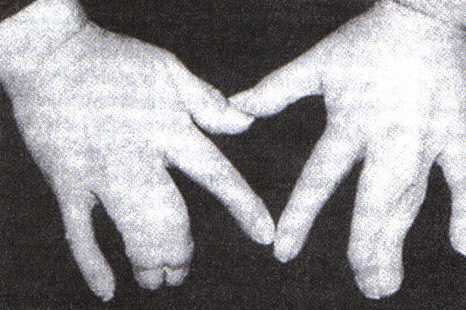
Si la muerte de las células interdigitales no se produce, una membrana de tejido blando conecta los dedos por ambos lados; ésta es la base del desarrollo normal de los pies palmeados en las aves palmípedas, como los patos, y de la formación anómala de la sindactilia (figura 9-13 A) en los seres humanos. No se encuentra BMP en el mesodermo interdigital de los pies de los patos en desarrollo, aunque si se encuentra muerte celular en otras zonas de la extremidad del pato.
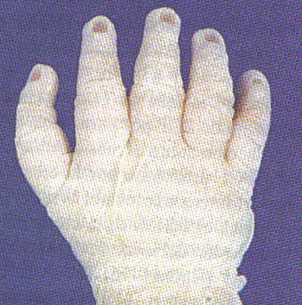
Todos los dedos humanos contienen tres segmentos falángicos con excepción de los primeros dedos (pulgares y dedos gordos), que sólo tienen dos segmentos. Algunos investigadores han atribuido el desarrollo de los primeros dedos bifalángicos a las acciones de una pequeña zona de muerte celular que probablemente existe en la punta del primordio del primer dedo. En raras ocasiones nace un individuo que tiene un pulgar con tres falanges (figura 9-13 B), que podría relacionarse con la ausencia de la muerte celular normal que tiene lugar en la punta del primordio del pulgar.
Zona de actividad polarizante y señales morfogenéticas
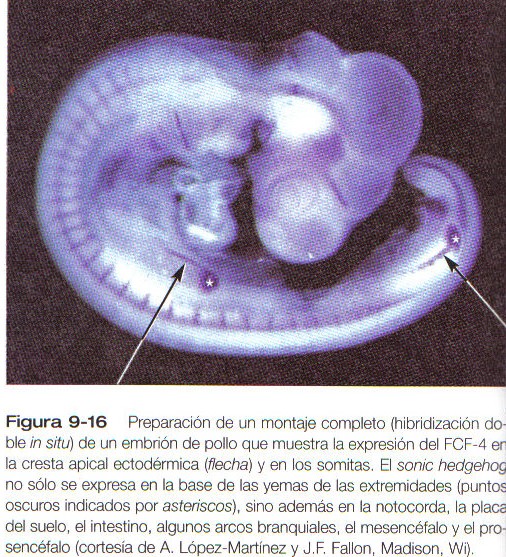
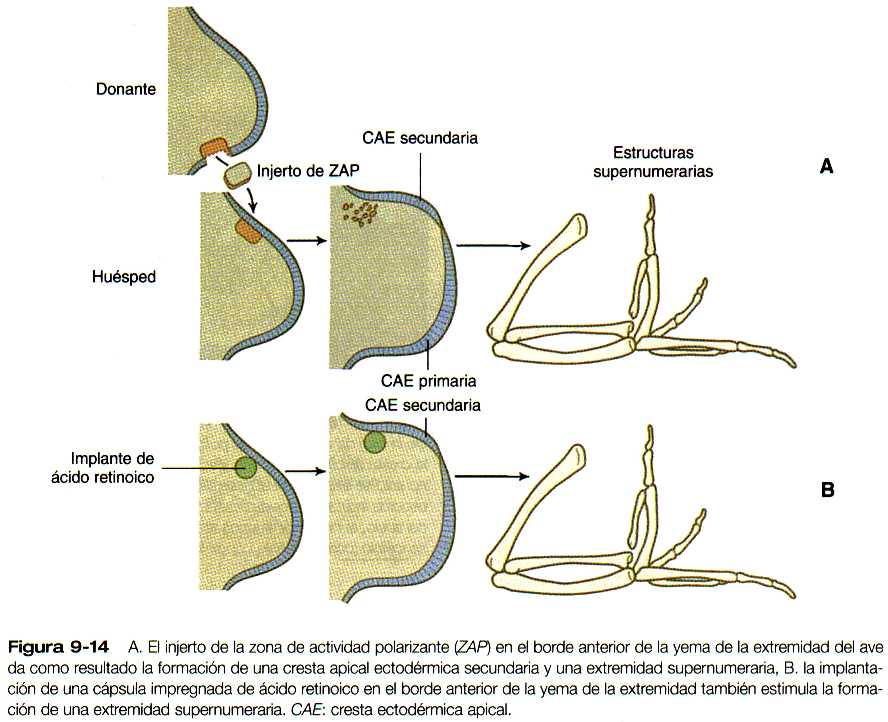
En los experimentos que han investigado la muerte celular morfogenética, los investigadores han injertado células mesodérmicas de la base posterior de la yema del ala de las aves en el borde anterior. El resultado de esta manipulación es la formación de un ala supernumeraria que era la imagen en espejo del ala normal (figura 9-14). Muchas investigaciones posteriores han demostrado que esta región posterior, llamada zona de actividad polarizante (ZAP), actúa como un centro emisor de señales y determina la organización de la extremidad a lo largo de su eje anteroposterior. Se ha demostrado que la señal es el sonic hedgehog, una molécula mediadora de varias interacciones tisulares en el embrión (tabla 5-3). Como se verá en la siguiente sección, el sonic hedgehog no sólo organiza los tejidos a lo largo del eje anteroposterior. sino que también mantiene la estructura y la función de la cresta apical ectodérmica. En ausencia de la ZAP o del sonic hedgehog la cresta apical involuciona.
Los experimentos de injertos cruzados entre especies han demostrado que las yemas de las extremidades de los mamíferos (incluidos los seres humanos) también contienen una ZAP funcional. Una ZAP trasplantada actúa sobre la cresta ectodérmica apical, donde suscita una respuesta de crecimiento en las células mesenquimatosas situadas por debajo de la región de la cresta adyacente a la ZAP trasplantada. Tan sólo 50 células de la ZAP pueden estimular la formación de una extremidad supernumeraria. Otras estructuras, como los fragmentos del nodo de Hensen, la notocorda e incluso las yemas de las plumas, pueden estimular la formación de miembros supernumerarios si se injertan en el borde anterior de la extremidad. Desde que se hicieron estos experimentos, se ha demostrado que todos los tejidos implantados son fuente de sonic hedgehog.
Modelo de control morfogenético de la extremidad en desarrollo En gran cantidad de datos sobre la expresión de los genes durante los últimos tres o cuatro años, junto con una sólida base de datos aportados por la embriología experimental, permite la construcción de un modelo tentativo para explicar e! control morfogenético del desarrollo de la extremidad. Al parecer, el desarrollo de las extremidades se inicia debido a la liberación de moléculas de activación (posiblemente el FGF-8) desde regiones situadas en el tronco. El resultado exacto de esta señal inductiva es la expresión de FGF-8 en el ectodermo que cubre la futura extremidad y su localización sólo en la región donde se formará la cresta apical ectodérmica.
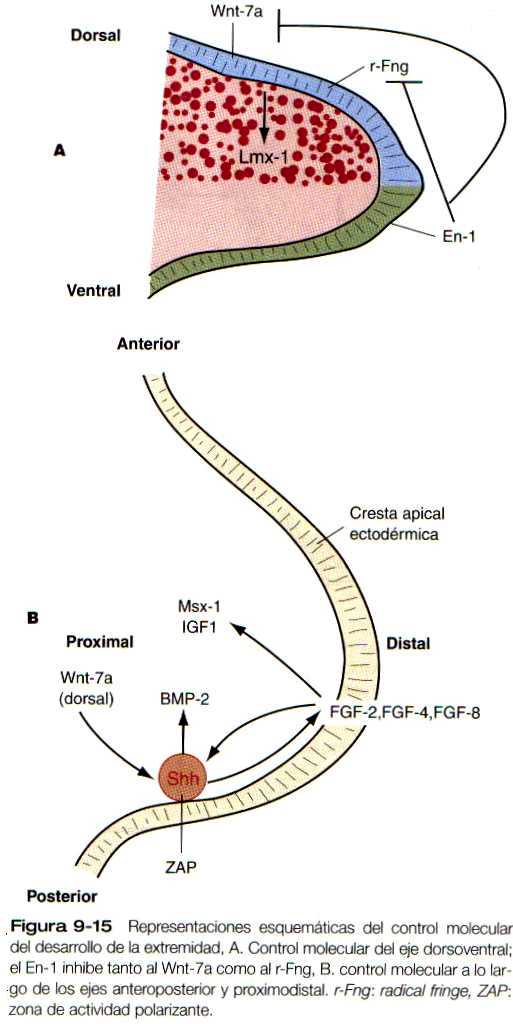
Durante estas primeras etapas, antes de que haya ninguna evaginación detectable que corresponda a la extremidad, unos hechos importantes en el desarrollo fijan el eje dorso-ventral del campo morfogenético. En un comienzo, el mesodermo contiene información sobre el patrón dorsoventral, pero ésta pronto se transfiere al ectodermo. Después, en el ectodermo del dorso del miembro se desarrolla un centro de señales morfogenéticas caracterizado por la expresión de Wnt-7a (tabla 9-1). El Wnt-7a induce la expresión del Lmx I, un gen portador de homeobox en el mesodermo dorsal subyacente y casi al mismo tiempo el ectodermo ventral comienza a expresar En-1 (figura 9-15 A). Esta combinación de acontecimientos moleculares instruye al mesodermo de la futura yema de la extremidad a que adopte una posición dorsal bajo la influencia del Wni-7a o ventral bajo la influencia del En-E Éste último parece inhibir la expresión del Wnt-7a en los tejidos ventrales.
De una forma aun no aclarada, el ectodermo situado en la frontera entre el ectodermo dorsal y el ventral de la extremidad se engruesa para formar la cresta apical ectodérmica. El ectodermo dorsal incipiente expresa un gen llamado radical fringe. La cresta apical ectodérmica forma el límite entre las células que expresan radical fringe y las que no lo hacen. Como sucede con el Wnt-7a. el En-1 inhibe la expresión del gen radical fringe en los tejidos ventrales. Estas células de la cresta apical ectodérmica precoz expresan varios genes, como FGF-2, FGF-4, FGF-8. BMP-2, BMP-4 v Msx-2. Se Considera que los acontecimientos que han tenido lugar hasta este punto pertenecen a la fase inicial del desarrollo de la extremidad.
A continuación, viene la fase de evaginación, que comienza con la secreción de FGF por parte de la cresta ectodérmica apical y su acción sobre el mesodermo subyacente (figura 9-15 B). El FGF producido por la cresta tiene dos efectos principales. Al interactuar con el Wnt-7a producido por el ectodermo dorsal, el FGF-8 de la cresta ectodérmica apical estimula la expresión del sonic hedgehog en el mesodermo posterior, la ZAP (figura 9-16). A su vez, bajo la influencia del sonic hedgehog, el FGF-4 comienza a expresarse en la mitad posterior de la cresta apical ectodérmica. Una vez que se ha inducido la producción del FGF-4. este y el sonic hedgehog conforman un circuito de retroah-mentación positiva, y cada uno estimula la formación del otro. El FGF segregado por la cresta ectodérmica apical, que actúa a través del factor 1 de crecimiento similar a la insulina, también estimula la proliferación de células mesodérmicas en toda la yema de la extremidad.
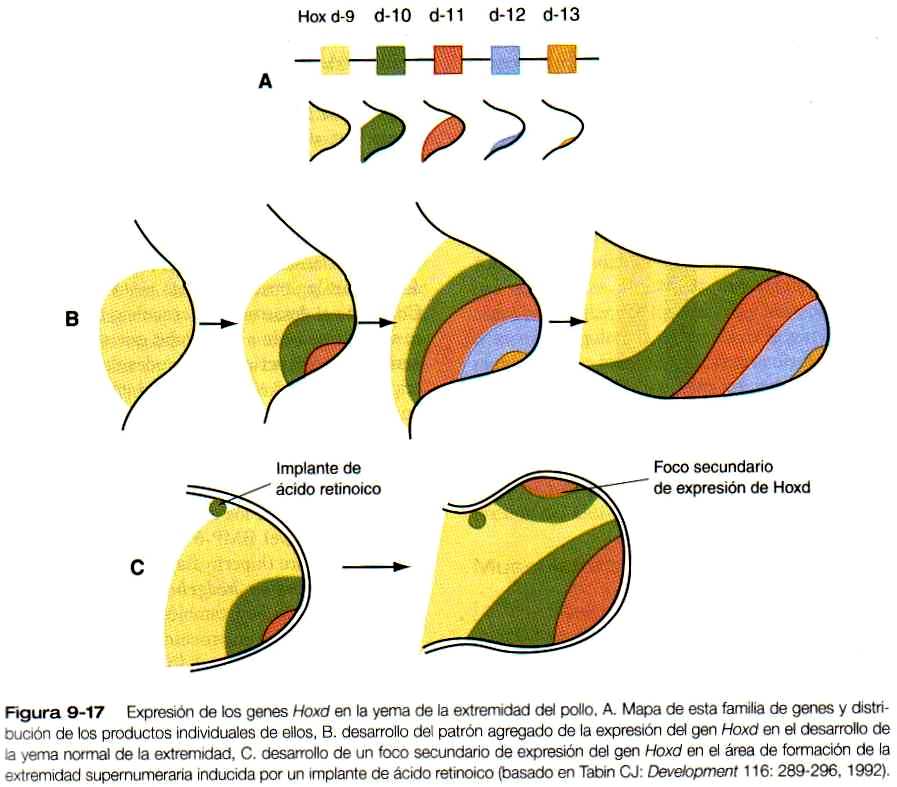
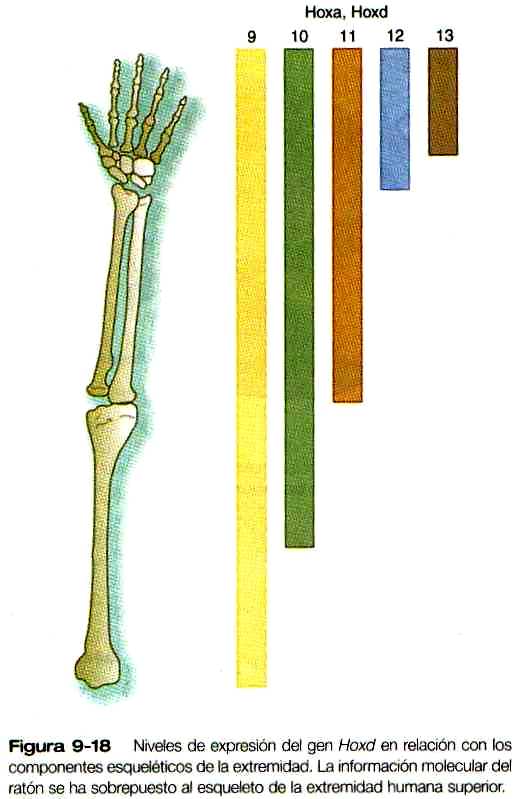
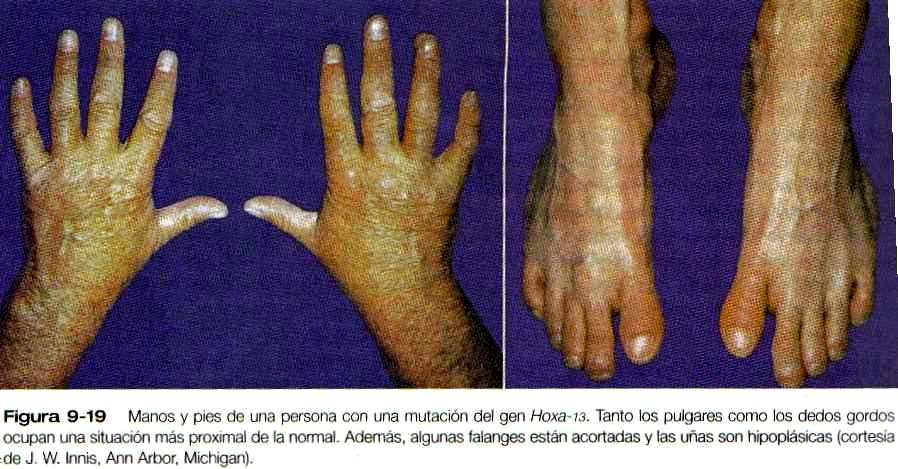
Al mismo tiempo que se estableció la ZAR se produce una secuencia ordenada de expresión de los genes que contienen homeobox Hoxd-9 a Hoxd-13 (figuras 9-17 A y B), asi como de algunos de los genes Hoxa, en la yema inicial de la extremidad. Todavía no se sabe qué estimula la expresión de los genes Hox. Las evidencias en algunos vertebrados sugieren que los genes Hoxa están implicados principalmente en el modelamiento del patrón a lo largo del eje próximo distal y que los genes Hoxci también tienen que ver con el modelamiento del patrón a lo largo del eje anteroposterior. Los estudios en ratones han mostrado defectos de las extremidades correspondientes a zonas de expresión de los genes Hoxa y Hoxti (figura 9-18). Las mutaciones del Hoxa-13 y de! Hoxd-13 causan patrones característicos de polisindactilia así como defectos de reducción de los dedos resultantes del acortamiento de las falanges (figura 9-19).
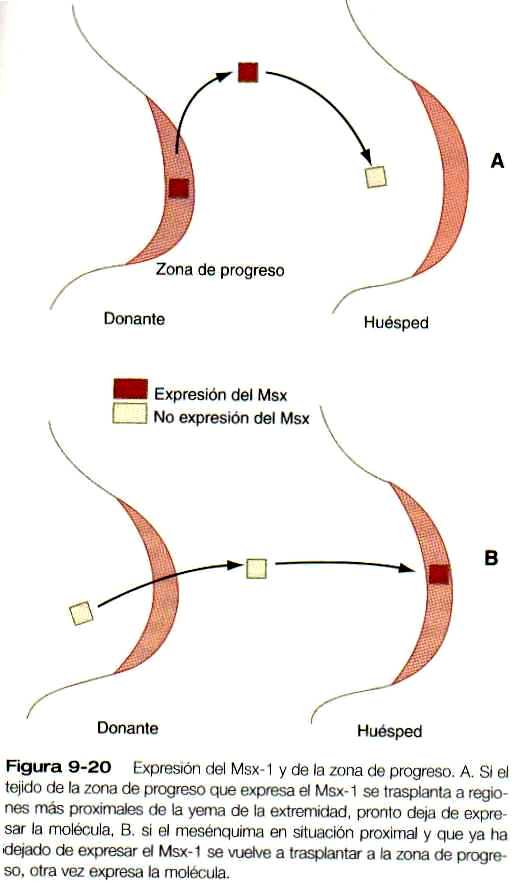
El establecimiento de la ZAP y la secreción de las proteínas del sonic hedgehog activan el centro de emisión de señales que organiza el desarrollo de las extremidades a lo largo del eje anteroposterior. El sonic hedgehog induce la expresión del BMP-2 en el mesodermo posterior. Además, actúa de forma recíproca sobre la cresta ectodérmica apical. En ausencia de la actividad del sonic hedgehog, la cresta apical no puede mantenerse. La cresta apical ectodérmica actúa como centro organizador para el eje proximodistal de la extremidad mediante la estimulación de las células del mesodermo. Justo debajo de la cresta apical ectodérmica se encuentra una región de mesodermo distal de varias mieras de espesor, conocida como zona de progreso (figura 9-20). Las células de la zona de progreso se dividen activamente y no están determinadas desde el punto de vista morfogenético. Sin embargo, a medida que la yema de la extremidad crece, las células mesodérmicas que están en la zona proximal de la extremidad pueden liberarse de la influencia de la cresta apical y quedan determinadas en su destino morfogenético, de manera que las células que abandonan primero la zona de progreso formarán elementos esqueléticos (como el húmero y el fémur) y las que lo hacen más tarde forman elementos del antebrazo o de la mano en el miembro superior. Las células de la zona de progreso se caracterizan por la expresión del gen Mv,v-/, y conforme van abandonando esta zona, la expresión de dicho gen cesa. Hay algo en el ambiente de la zona de progreso que estimula la expresión del Mv.v-/. puesto que si las células que ya han dejado la zona (y en consecuencia ya no están expresando dicho gen) vuelven a trasplantarse allí. expresan de nuevo el gen (figura 9 -20 B). Además, los valores de información de posición (véase más adelante) de las células trasplantadas vuelven a fijarse de manera que ahora pueden formar estructuras más dístales de lo que habrían hecho si no se hubieran trasplantado.
Una pregunta fundamental aún sin respuesta en el desarrollo de las extremidades es qué hace que un brazo se vuelva brazo y que una pierna se vuelva pierna. Está bastante claro que el desarrollo de las extremidades superiores e inferiores implica muchos mecanismos comunes, pero aún no se sabe qué determina la diferencia de forma entre los brazos y las piernas.
Los embriólogos moleculares están comenzando a identificar ciertos tipos de genes que contienen homeobox y que se expresan de forma diferencial entre los brazos y las piernas, pero aún no cuentan con una explicación coherente de las diferencias entre ambos.
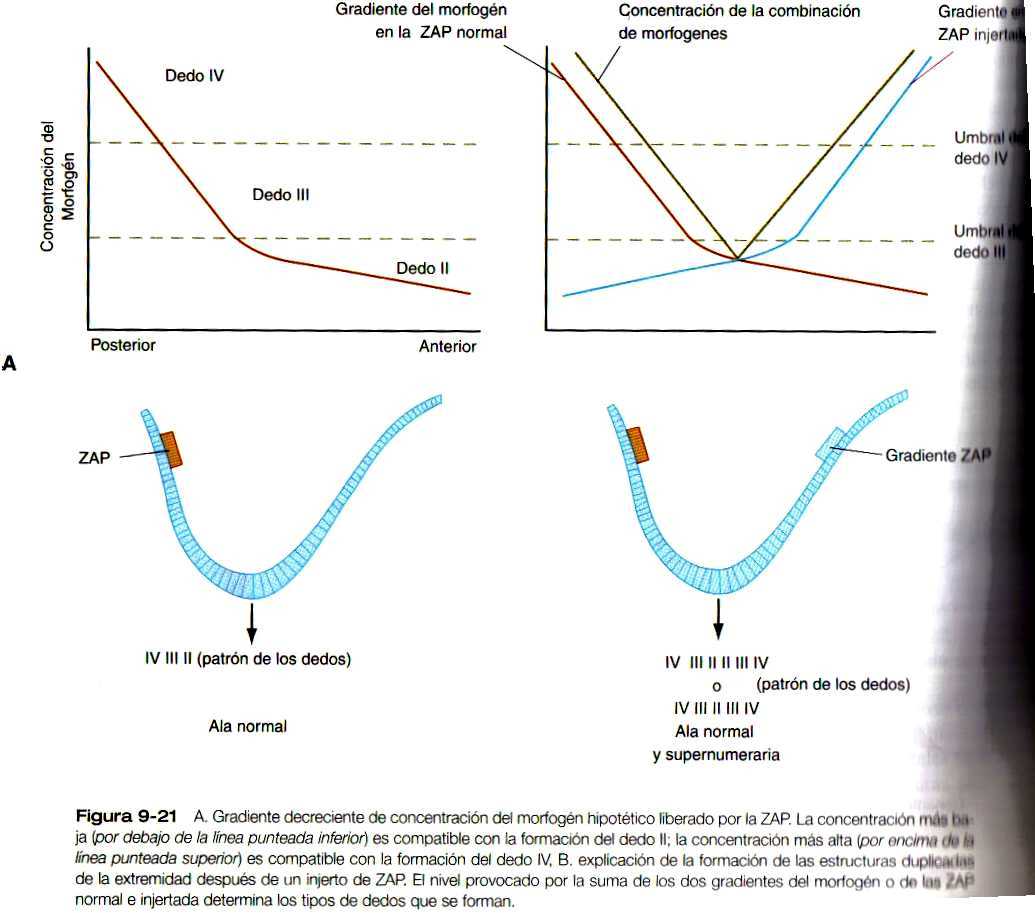
Muchos de los datos recientes aportados por la biología molecular reciente se ajustan al modelo antiguo de información posicional. Según este concepto, las células de una estructura en desarrollo, como el mesodermo de una extremidad, están expuestas a señales del ambiente. De alguna manera, estas células pueden recibir e interpretar las señales y luego se diferencian de acuerdo con esta información. Un ejemplo específico que guarda relación con el desarrollo de los miembros implica la influencia de la ZAP sobre la extremidad en desarrollo. Según la teoría clásica, la ZAP produce un morfogén difusible (una molécula capaz de influir sobre la morfogénesis); a medida que esto se difunde por el mesodermo. Su concentración disminuye con el tiempo y la distancia, con lo cual se establece un gradiente de concentración. Las células situadas a lo largo del eje anteroposterior de la yema de la extremidad pueden percibir las diversas concentraciones del morfogén (es de suponerse que sea una proteina sonic hedgehog o una molécula inducida por este gen) y se diferencia en función de ésta (figura 9-21). Una explicación de la formación de una extremidad supernumeraria cuando se trasplanta un fragmento del mesodermo posterior (o ácido retinoico) en el borde anterior de la yema de la extremidad (figura 9-14 A) es que las células del mesodermo anterior ahora están expuestas a una alta concentración del morfogén y se les hace creer que deberían convertirse en estructuras posteriores. Se induce una segunda cresta apical sobre el injerto y aparece una organización duplicada de expresión del gen Hoxd (figura 9-17 C) antes de la formación de una extremidad supernumeraria en espejo.
DESARROLLO DE LOS TEJIDOS DE LAS EXTREMIDADES
La mayor parte de los sucesos morfogenéticos que acaban de describirse tienen lugar durante las primeras etapas del desarrollo de la extremidad cuando la yema es una masa de apariencia homogénea de células mesodérmicas cubiertas de ectodermo. La diferenciación y la histogénesís de los componentes tisulares específicos de la extremidad son sucesos más tardíos del desarrollo que se construyen sobre el plan maestro morfogenético previamente establecido.
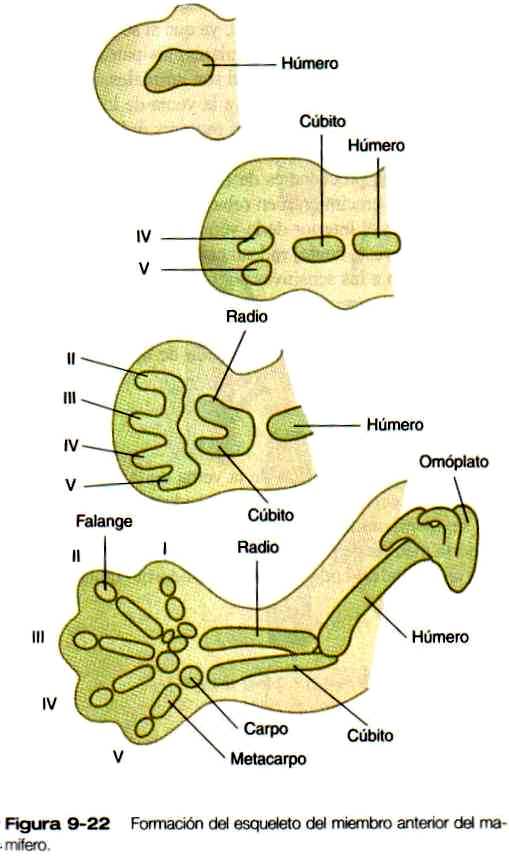
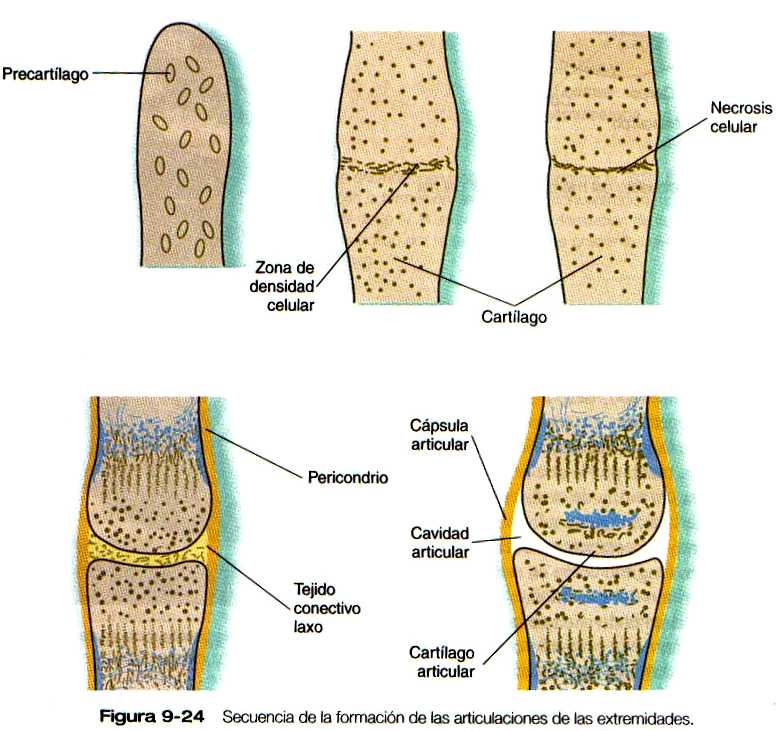
El esqueleto es el primero de los tejidos de la extremidad que muestra signos evidentes de diferenciación. Su morfología macroscópica, ya sea normal o anormal, refleja estrechamente los principales sucesos de formación del patrón maestro que "moldean" la extremidad como un todo. La formación del esqueleto puede verse primero como una condensación de células mesenquimatosas en la zona central de la parte próxima! de la yema de la extremidad. Aun antes de condensarse, estas células están determinadas a formar cartílago, y si se trasplantan a otros lugares o se cultivan, sólo se diferencian en cartílago. No obstante, otras células mesenquimatosas que normalmente formarían tejido conectivo mantienen la capacidad de convertirse en cartílago si se trasplantan en la región central de la yema de la extremidad.
El ectodermo del vértice de la extremidad ejerce un efecto inhibitorio sobre la diferenciación del cartílago, de manera que no se forma cartílago en la región situada inmediatamente por debajo del ectodermo. Los estudios in vitro sugieren que la inhibición está mediada por un material difusible segregado por el ectodermo; está por averiguarse si éste es el mismo mecanismo que mantiene la integridad de la zona de progreso.
Las condensaciones celulares que componen los agregados precartilaginosos expresan tanto BMP-2 como BMP-4. Conforme el desarrollo del esqueleto continúa, su expresión se va restringiendo a las células que van a convertirse en el pericondrio o el periostio que rodean a los huesos. Los transcriptos de BMP-3 se detectan por primera vez en e! cartílago, y no en el precartílago. pero este factor de crecimiento también termina por aparecer en el pericondrio. La traslocación de la expresión de estas moléculas de BMP en el pericondrio refleja su papel continuado en las primeras fases de la diferenciación de los tejidos esqueléticos.
En contraste, el BMP-6 sólo se expresa en las áreas de cartílago maduro (hipertrofia) dentro de los huesos de la extremidad. El indian hedgehog, una molécula relacionada con el sonic hedgehog, también se expresa en las mismas rexxxx del cartílago en proceso de hipertrofia (en el que además existe colágeno tipo X) y es esta molécula de señal la que puede inducir la expresión del BMP-6.
La diferenciación del esqueleto cartilaginoso tiene lugar según una secuencia proximodistal, y en los mamíferos las estructuras post axiales de los segmentos distales de la extremidad se diferencian antes que las estructuras preaxiales.
La musculatura de la extremidad se deriva de las células miogénicas que migran hacia la yema inicial del miembro desde la parte ventral del dermomiotoma del somita. Estas células son inducidas a abandonar el somita y migrar hacia la extremidad gracias a la acción del factor de dispersión (factor de crecimiento hepático), que es segregado por las células proximales del área prospectiva de la futura extremidad. Antes de migrar, las células premusculares del somita expresan el c-met, que es un receptor específico del factor de dispersión. Las células premiogénicas, cuya morfología es indistinguible de la de las otras células mesenquimatosas pero que ya expresan Pax-3, se diseminan por toda la yema de la extremidad. En el mutante splotch. que se caracteriza porque no se expresa el Pax-3, las células musculares no invaden la yema de la extremidad. La migración de las células premusculares se mantiene al mismo ritmo que la elongación de la yema de la extremidad y aunque las células expresan moléculas musculares características (por ejemplo, la Myo-D) no se detectan en la zona de progreso. Algunas pruebas experimentales sugieren que las células premiogénicas no están presentes en la zona de progreso; otros experimentos han demostrado que el ambiente de esta zona suprime la expresión de la Myo-D y de otras moléculas especificas del músculo, quizá a través de la influencia promotora de la mitosis de los FGF en esa área.
Poco después de que las condensaciones de los elementos esqueléticos se forman, las células miogénicas comienzan a condensarse en dos masas musculares comunes: una es la masa precursora de los músculos flexores y la otra dará lugar a los músculos extensores. La siguiente etapa en la formación del músculo es la división de las masas musculares comunes. La fusión de mioblastos en miotubos incipientes comienza durante estas primeras etapas del desarrollo muscular.
Hay bastantes datos que sugieren que las células miogénicas precursoras no poseen información intrínseca que guie su morfogénesis. En cambio, parece que las células miogénicas son guiadas por las células del tejido conectivo, que son los portadores y los efectores de la información morfologenética requerida para formar músculos correctos desde el punto de vista anatómico. En experimentos en los cuales se extirpan los somitas que normalmente se asocian con una yema de extremidad y se remplazan por somitas de cualquier otro lugar del eje corporal, las células miogénicas se comportan indiferentemente en el aspecto morfogenético, la morfogénesis del músculo es normal aunque los precursores de las fibras musculares se deriven de fuentes anormales. Por medio de la extirpación de los somitas, el uso de los rayos X y el análisis de algunos mutantes como el wingless, se ha demostrado que los componentes tendinosos y el tejido conectivo del músculo se derivan del mesodermo de la yema de la extremidad, en tanto que las fibras musculares se derivan de los somitas. En situaciones en las cuales se impide que las células miogénicas invadan la yema de la extremidad, se forman tendones de morfología normal, pero no están unidos a "músculos". En las mismas extremidades, otras células del tejido conectivo forman un "molde del músculo", aunque no contiene fibras musculares.
En función del músculo específico; a formar la migración, la fusión o el desplazamiento de los primordios musculares pueden estar implicados en la génesis de su forma final. Por ejemplo, la muerte celular genéticamente programada la apoptosis es responsable de la desaparición de una capa muscular completa (los músculos contrahentes) en el lado flexor de la mano humana. Las células miogénicas se diferencian en la etapa de miotubo; después, acumulan el glucógeno y muy pronto degeneran. La capa de los músculos contrahentes está conservada en la mayoría de los grandes simios. No se conoce la razón por la que degenera en la mano humana en una etapa tan tardía de su diferenciación. Si bien los músculos de la extremidad adoptan su forma definitiva en el embrión muy precozmente, deben crecer de forma considerable tanto en longitud como en sentido transversal para mantenerse al ritmo del crecimiento general del embrión. Esto se logra mediante la división de las células satélites (capitulo 8) y la fusión de su progenie con las fibras musculares. Los núcleos añadidos de las células satélites aumentan el potencial de la fibra muscular para producir proteínas estructurales v contráctiles, las cuales aumentan el área de la sección transversa de cada fibra muscular. La suma de núcleos compleméntanos a las fibras musculares se acompaña de un alargamiento secundario debido a la agregación de más sarcómeros, por lo general en los extremos de las fibras musculares.
Es característico que la formación de nuevas fibras musculares se detenga poco después del nacimiento. Aunque los músculos pueden contraerse al comienzo del período fetal, sus propiedades fisiológicas continúan diferenciándose hasta después del nacimiento.
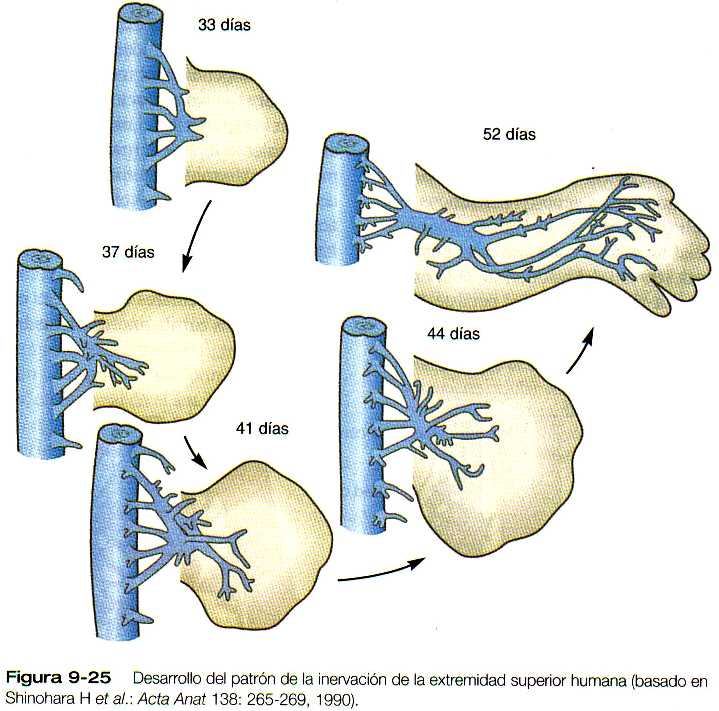
Los axones motores que se originan en la médula espinal entran en la yema de la extremidad en una fase inicial del desarrollo (durante la quinta semana) y comienzan a introducirse en las masas musculares ventrales y dorsales antes de que las dorsales se hayan dividido en los primordios individuales de los músculos (figura 9-25). Los estudios con trazadores han demostrado un alto grado de ordenación en la proyección de las motoneuronas en la extremidad. Las neuronas ubicadas en posiciones mediales en la médula espinal envían sus axones a la masa muscular ventral, en tanto que los que están en una localización más lateral en la médula inervan la masa muscular dorsal. De igual forma, existe una correlación entre la posición craneocaudal de las neuronas en la médula y el patrón de inervación anteroposterior de los músculos de las extremidades en el interior de las masas musculares iniciales. Por ejemplo, las neuronas más rostrales inervan los primordios musculares más anteriores.
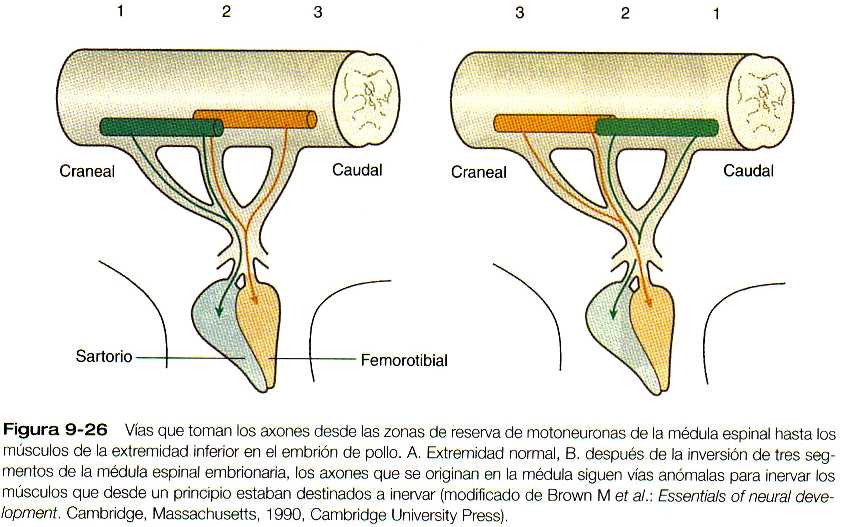
Parece ser que unas señales locales en la base de la yema de la extremidad son las guías de la vía de entrada de las fibras musculares a la yema. Si se invierte en sentido craneocaudal opuesto al área de evaginación de la yema de la extremidad un segmento de la médula espinal, las motoneuronas cambian la dirección de su crecimiento y entran a la yema en sus posiciones normales (figura 9-26). Si se invierten segmentos más largos de la médula y las motoneuronas se encuentran a considerable distancia del nivel de la yema de la extremidad, sus axones no encuentran el camino hacia sus localizaciones normales en la extremidad. En apariencia los propios músculos no emiten señales específicas para atraer a los axones que crecen hacia ellos, ya que si se impide la formación de los primordios musculares, los patrones básicos de la inervación de la extremidad son normales.
Los axones sensitivos entran a la yema de la extremidad después de que lo han hecho los motores, y los usan como guía. De forma similar, los precursores celulares de las células de Schwann procedentes de la cresta neural se retrasan un poco en su crecimiento en relación con el de los axones motores, hacia el interior de la yema de la extremidad. Las células de la cresta neural rodean tanto a las fibras nerviosas motoras como a las sensitivas para formar las vainas de los nervios de ¡as extremidades. En el momento en que se han formado los dedos en los miembros en desarrollo, el patrón macroscópico básico de la inervación de la extremidad del adulto ya está establecido.
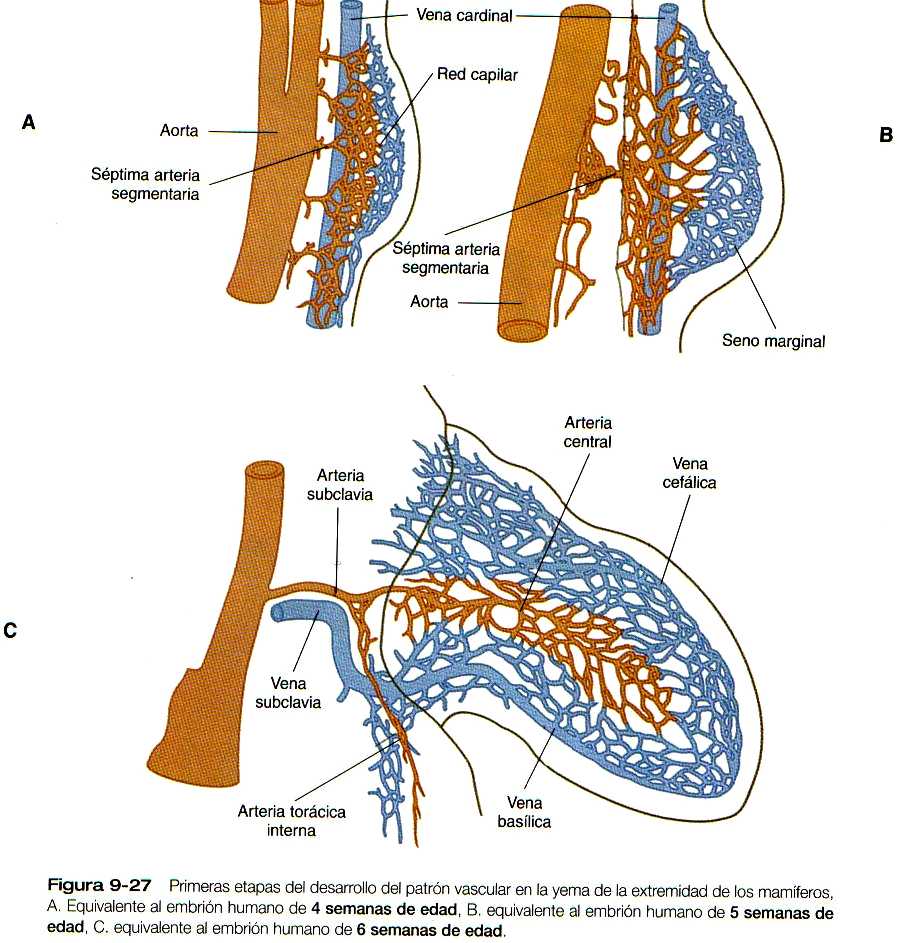
La primera vascularización de la yema de la extremidad se deriva de las células endoteliales procedentes tanto de las ramas segmentarias de la aorta como de las venas cardinales y en cierta medida de los angioblastos (precursores de las células endoteliales ) endógenos del mesodermo de la yema de la extremidad. Al comienzo, la vascularización de la extremidad se compone de una fina red de capilares, pero pronto algunos de ellos experimentan un desarrollo preferente y, cuyo resultado es una arteria central grande que irriga la yema de la extremidad (figura 9-27). Desde la arteria central, la sangre se distribuye hacia la periferia a través de una red de capilares y después se recoge en un seno marginal, localizado por debajo de la cresta apical ectodérmica. La sangre del seno marginal drena en canales venosos periféricos que la transportan fuera de la yema de la extremidad.
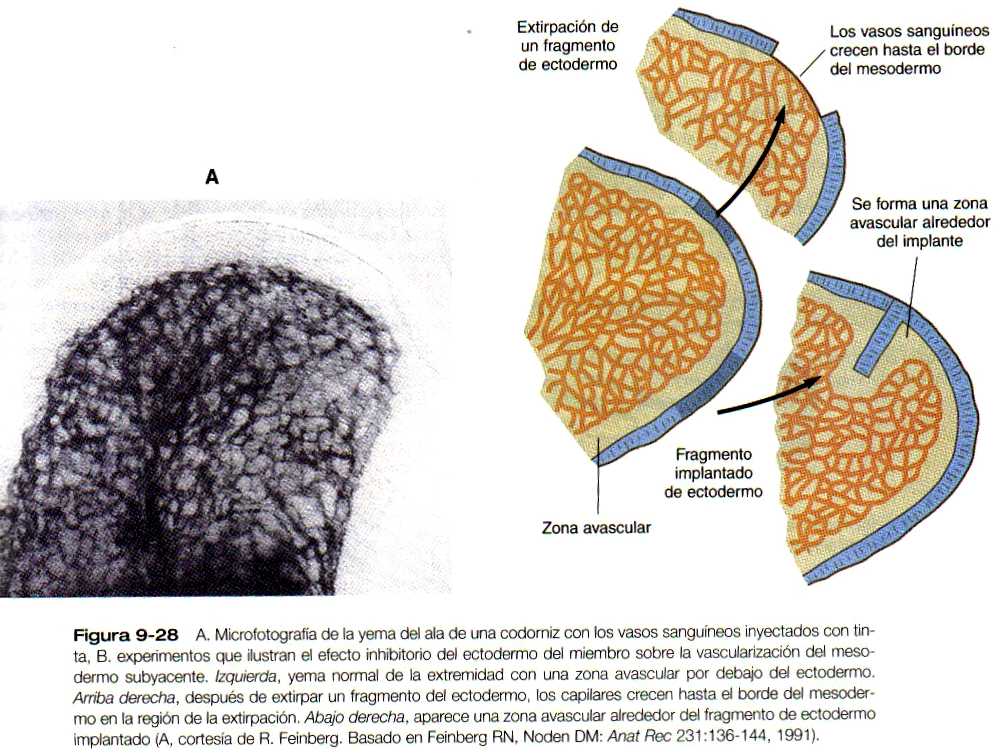
Aun en la yema inicial existe una zona avascular periférica de mesodermo a unas 100 (чm del ectodermo de la extremidad (figura 9-28 A). La región avascular persiste hasta que los dedos han comenzado a formarse. Hay angioblastos en la zona avascular, pero están aislados de los capilares funcionales. Los estudios experimentales han demostrado con claridad que la proximidad del ectodermo es inhibidora para la vasculogénesis del mesodermo de la yema de la extremidad. Si se extirpa el ectodermo, se forman canales vasculares en la superficie del mesodermo de la yema, y si se pone un fragmento de ectodermo en la profundidad del mesodermo de la extremidad, se forma una zona avascular a su alrededor (figura 9-28 B). Los productos de la degradación del ácido hialurónico, secretado por el ectodermo, parecen ser los agentes inhibidores.
Poco antes de que el esqueleto comience a formarse, aparecen zonas avasculares en las áreas en las cuales se formarán los moldes cartilaginosos de los huesos. Hasta ahora no se conocen ni el estímulo que da lugar a la desaparición de los vasos sanguíneos ni el destino dé las células endoteliales que estaban presentes en esa región.
El patrón de la red vascular está sujeto a constantes cambios conforme se desarrolla el miembro, en especial por la formación de evaginaciones y brotes en los canales existentes, la regresión de los canales originales y la fusión de los brotes de los nuevos vasos. Este tipo de mecanismo explica la progresión distal del seno marginal. Con el establecimiento de los radios digitales, la porción apical del seno marginal se fragmenta, pero los canales proximales del seno marginal persisten hasta la edad adulta como las venas basílica y cefálica del brazo (figura 9-27 C).
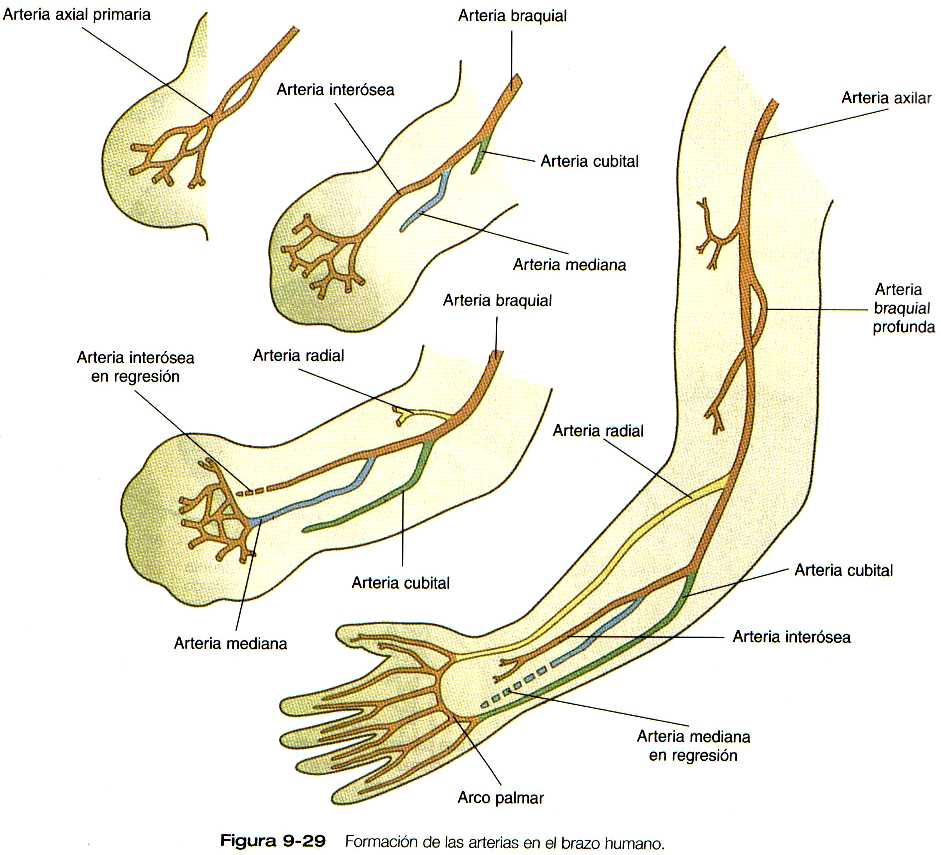
Unos cambios similares tienen lugar en los canales arteriales. que recorren la extremidad en desarrollo (figura 9-29). Las ramas que se desprenden de la arteria axial primaria terminan por hacerse dominantes, en especial en el antebrazo, y dejan a la arteria axial primaria original convertida en un vaso relativamente menor (la arteria interósea) en el antebrazo.
• Las extremidades surgen del mesodermo de la lámina lateral y del ectodermo que lo cubre bajo la influencia de un inductor (posiblemente el FGF-8 del mesodermo intermedio). La yema inicia] de la extremidad es un sistema de alta regulación que puede compensar numerosas perturbaciones quirúrgicas y formar un miembro normal. Los ejes de la extremidad se fijan en la secuencia anteroposterior, dorsoventral y proximodistal.
• El mesodermo de la yema inicial de la extremidad estimula al ectodermo que lo cubre para que allí se forme una cresta apical ectodérmica que activa el crecimiento de la extremidad mediante la proliferación de las células del mesodermo subyacente. La cresta apical segrega FGF-2 y FGF-4 que pueden inducir el crecimiento en ausencia de la cresta. La morfogénesis general del miembro está determinada por las propiedades del mesodermo, mientras que el ectodermo actúa en un sentido más permisivo.
• La muerte celular es un mecanismo importante en el desarrollo normal de la extremidad. Las regiones de muerte celular programada incluyen la región axilar y los espacios interdigitales. Si las células interdigitales no mueren, se produce la sindactilia.
Una ZAP localizada en el mesodermo posterior actúa como centro biológico de emisión de señales y desempeña un papel importante en la organización anteroposterior de la extremidad porque emite sanie hedgehog. El Wnt-7a, liberado por el ectodermo dorsal, es el organizador de la orientación dorsoventral de la yema de la extremidad. Según la teoría de la información posicional, las células de la extremidad en desarrollo están expuestas a informaciones posiciónales (como las señales de la ZAP) que les permiten determinar su posición relativa en el interior de la yema de la extremidad. Las células procesan esta información y se diferencian de conformidad con ella. El control proximodistal de la morfogénesis puede residir en la zona de progreso, una delgada capa de mesodermo que se encuentra por debajo de la cresta apical ectodérmica.
El ácido retinoico ejerce un importante papel en la morfogénesis de las extremidades y puede causar la formación de una extremidad supernumeraria si se injerta en el borde anterior de la yema de la extremidad, pero aún no se ha definido con claridad su función exacta en el desarrollo de los miembros. La expresión de varios genes que contienen homeobox sigue un patrón bien definido en el desarrollo normal de la extremidad. Algunos de los patrones de expresión de genes sufren profundas alteraciones en las extremidades tratadas con ácido retinoico. El esqueleto de la extremidad surge del mesodermo de la lámina lateral. El ectodermo de la yema de la extremidad inhibe la formación de cartílago en las células del mesodermo situadas justo por debajo de él: esto podría explicar por qué el esqueleto de las extremidades de los vertebrados se forma en posición central.
Los músculos de las extremidades se forman a partir de células derivadas del mesodermo de los semitas. Estas células expresan el gen Pax-3 durante su migración hacia la yema de la extremidad. Las células miogénicas primero forman las masas musculares primarias dorsal y ventral, que más adelante se dividen en los primordios individuales de cada músculo.
El control morfogenético de los músculos reside en el tejido conectivo asociado y no en las células musculares mismas. Las etapas más tardías del desarrollo muscular pueden implicar muerte celular, fusión de los primordios de los músculos y desplazamiento de los primordios musculares hacia otras áreas.
Los nervios crecen hacia la yema de la extremidad en desarrollo y se introducen en las masas musculares primarias conforme éstas se dividen para formar los músculos individuales. La emisión de señales locales son importantes para guiar a los axones en crecimiento hacia la extremidad en desarrollo. La vascularización de la yema de la extremidad surge de células que se desprenden de la aorta y de las venas cardinales por gemación, así como a partir de células mesodérmicas locales.
El patrón vascular primario consta de una arteria central que drena en el seno marginal periférico y éste, a su vez, en los canales venosos periféricos. Los vasos sanguíneos no se forman por debajo del ectodermo ni en las regiones centrales formadoras de cartílago.
CORRELACIÓN CLÍNICA 9-1
Anomalías de las extremidades
Debido a que son tan evidentes, las anomalías de las extremidades se describen con frecuencia en la bibliografía. No obstante, la etiología de gran número de ellas aún se desconoce, en especial cuando se consideran los casos individuales. Algunas anomalías de las extremidades que afectan a los seres humanos se asemejan a las que se han producido por manipulaciones experimentales específicas, muchas otras en cambio no pueden atribuirse a trastornos de mecanismos específicos.
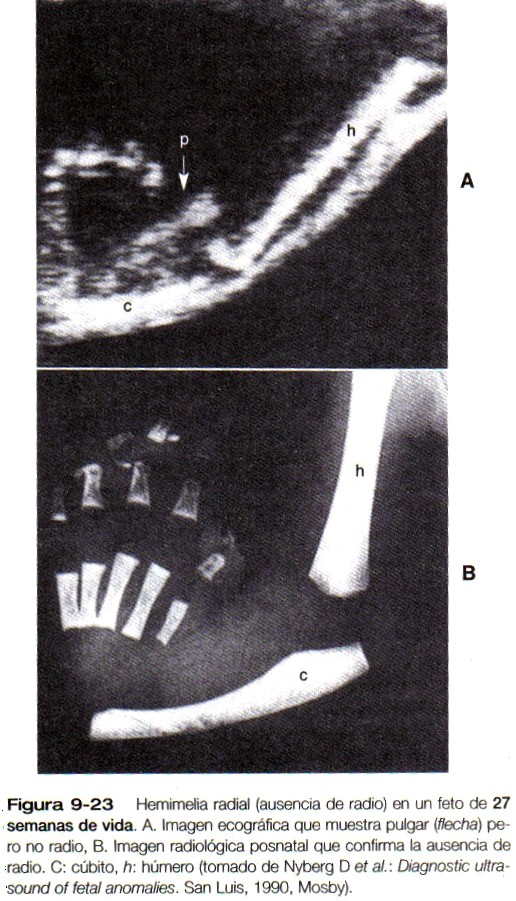
Diversas anomalías de las extremidades se han atribuido a malformaciones vasculares, pero no siempre es fácil saber si éstas últimas precedieron o acompañaron al desarrollo general anómalo de la extremidad. Una de las explicaciones que han persistido para entender el defecto de reducción proximal en la focomelia (figura 7-2), en la cual las estructuras dístales de la extremidad están bien formadas, es el daño secundario de los vasos sanguíneos en la parte proximal de la yema de la extremidad después de que se ha establecido el patrón de las estructuras distales de la mano o del pie. Esto provocaría entonces la regresión de las estructuras próximas de la extremidad.
Algunos de los defectos más frecuentes de los miembros pueden atribuirse a causas mecánicas. Las amputaciones intrauterinas causadas por bandas o bridas amnióticas, presumiblemente originadas en desgarros del amnios, pueden consistir en la pérdida de partes de los dedos o incluso de las manos o los pies (figura 7-15). Otras deformidades, como el pie zambo (tatípes equinovarus) y algunos casos de luxaciones congénitas, han sido atribuidas a presiones mecánicas mantenidas de la pared uterina sobre el feto, en especial en los casos de oligohidramnios (capítulo 6).
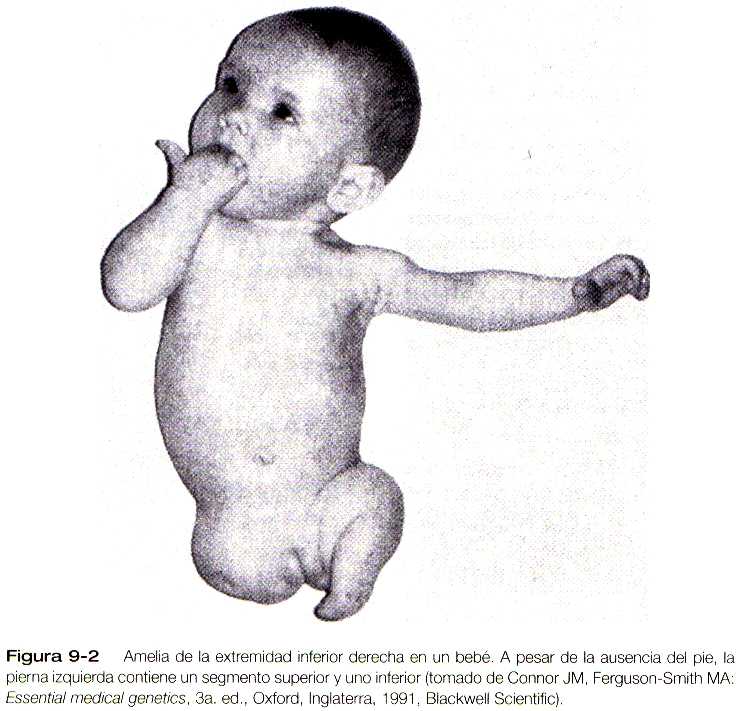
Otras anomalías de las extremidades son familiares y tienen base genética ya sea en la forma de rasgos dominantes o recesivos. Aunque se sabe que algunas anomalías como la Amelia y la polidactilia existen como mutaciones en animales de laboratorio, el defecto del desarrollo que subyace a la génesis de un defecto humano en particular puede no ser el mismo que el del modelo animal; varios mecanismos defectuosos pueden dar lugar al mismo fenotipo.
Una deformidad muy poco frecuente es la macromeiia (o macrodactilia), en la cual una extremidad o un dedo son considerablemente más grandes de lo normal. Estas anomalías se han asociado en ocasiones con neurofibromatosis, y quizá la cresta neural tenga que ver con este defecto.
Las anomalías de los miembros pueden ser el resultado de mutaciones genéticas, efectos farmacológicos, perturbación de las interacciones titulares o efectos puramente mecánicos.
CASO CLÍNICO
Después de un embarazo y un parto normales, una mujer de 32 años dio a luz a un niño que pesó 3.400 g y que tenía una duplicación del pie izquierdo a lo largo del eje anteroposterior. Hacia la izquierda de un dedo gordo único (hallux) había cuatro dedos adicionales organizados en forma simétrica en espejo, de manera que el patrón de los dedos de ese pie era 543212345 siendo el 1 el dedo gordo común. El bebé tenía otros dos hermanos mayores normales.
¿ Como explicaría esta malformación con base en los mecanismos del desarrollo que aprendió en este capítulo?
PREGUNTAS DE REPASO
i ¿Cuál de las siguientes moléculas desempeña un papel importante en la determinación del eje dorsoventral de la extremidad en desarrollo? A. Msx-1 B- Wnt-7a C Hoxd-13 D. Pax-1 E. FGF-8
2. ¿Qué molécula se asocia con la migración de las células miogénicas desde los somitas hasta la yema de la extremidad? A. Shh B. BMP-7 C. FGF-4 D. Pax-3 E En-1
3 ¿Cuál es la principal función de la cresta apical ectodérmica ? A Estimular el crecimiento de la yema de la extremidad
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}