Capítulo 16. SISTEMA CARDIOVASCULAR
DESARROLLO DEL SISTEMA VASCULAR
Hematopoyesis embrionaria
Aspectos celulares de la hematopoyesis
Síntesis de hemoglobina y su control
Formación de los vasos sanguíneos embrionarios
Desarrollo de las arterias. Arcos aórticos y sus derivados
Desarrollo de los conductos linfáticos
DESARROLLO Y TABICACION DEL CORAZON
Desarrollo inicial del corazón
Orígenes celulares
División inicial del canal aurículo-ventricular del corazón
Separación de las aurículas de los ventrículos
División del tracto de salida del corazón
COMIENZO DE LA FUNCIÓN CARDÍACA
CASO CLÍNICO
Malformaciones cardiacas(continuación: 16-1b 2)
CORTOCIRCUITOS DE CÁMARA A CÁMARA
Defectos del tabique interauricular
Atresia tricuspídea
Defectos del tabique interventricular
Malformaciones cardíacas (continuación 16-1c 3)
Transposición de los grandes vasos
CORRELACIÓN CLÍNICA 16-2
Malformaciones de los vasos sanguíneos
COARTACIÓN DE LA AORTA
Malformaciones del sistema linfático.
Este capítulo sigue el desarrollo del corazón desde una simple estructura tubular hasta el órgano de cuatro cámaras que puede asumir toda la responsabilidad de mantener una circulación independiente desde e! momento del nacimiento. De manera similar, seguiremos la formación de los vasos sanguíneos desde su aparición hasta la formación de un sistema integrado que lleve la sangre a todas las partes del embrión y la placenta (las primeras etapas en el establecimiento del corazón y los vasos sanguíneos se describen en el capitulo 5 [figuras 5-20, 21, 22, 23 y 5-24]. y el plan general de la circulación embrionaria se resume en la figura 5-31). Los aspectos celulares de la formación de la sangre también se describen brevemente. En las Correlaciones clínicas 16-1 y 16-2 que aparecen al final del capitulo se discuten las malformaciones del corazón y los vasos sanguineos, respectivamente. En la tabla 16-5 (no esta), también al final del capitulo, se resume la secuencia temporal del desarrollo cardiaco.
Desde el punto de vista funcional, el corazón embrionario sólo necesita funcionar como una bomba simple que mantiene el flujo de sangre a través del cuerpo del embrión y hacia la placenta, donde se intercambian los desechos fetales por oxigeno y nutrientes. Sin embargo, una función no menos importante es la previsión de los radicales cambios que tendrán lugar en la circulación al nacimiento como consecuencia de la súbita interrupción de la circulación placentaria y la iniciación de la respiración. Para satisfacer los complejos requerimientos del sistema circulatorio posnatal, el corazón embrionario tiene que desarrollar cuatro cámaras que puedan recibir o bombear todo el flujo de sangre que circula por el cuerpo. El corazón tiene también que adaptarse a la condición de los pulmones fetales, que tienen un desarrollo incompleto y que durante casi todo el periodo fetal no tienen una vascularización capaz de recibir un gran flujo de sangre. Este dilema fisiológico se resuelve gracias a la presencia de dos cortocircuitos que permiten que cada cámara cardiaca pueda trabajar con grandes cantidades de sangre a la vez que evita los vasos pulmonares subdesarrollados.
La morfogénesis cardiaca comprende interacciones celulares intrínsecas y moleculares, pero estas han de tener lugar con una función mecánica continua. Algunos de estos mecanismos aún se desconocen, pero otros se han venido definiendo mejor gracias a la investigación sobre el desarrollo cardíaco normal y anormal.
El desarrollo de la vascularización a nivel de los patrones macroscópicos de las arterias y las venas se ha entendido bastante bien desde hace muchos años. En época reciente los nuevos marcadores celulares y moleculares han permitido a los investigadores definir los orígenes celulares de las arterias y venas en órganos o en regiones específicos del organismo. La mayor parte de los estudios sobre los mecanismos de la diferenciación vascular aún se encuentran en sus inicios.
DESARROLLO DEL SISTEMA VASCULAR
El desarrollo del sistema vascular comienza. en la pared del saco vitelino durante la tercera semana de la gestación con la formación de islotes sanguíneos (figura 5-24). En este momento el embrión ha alcanzado un tamaño demasiado grande para que la distribución del oxigeno llegue a todos los tejidos sólo mediante difusión. Esto requiere el desarrollo muy precoz tanto del corazón como del sistema vascular. Puesto que los tejidos que normalmente producen las células de la sangre en el adulto todavía no han comenzado a formarse, la hematopoyesis del saco vitelino parece ser una adaptación transitoria para satisfacer las necesidades inmediatas del embrión.
Se piensa que las células que forman los islotes sanguíneos, llamadas hemangioblastos, tienen una capacidad de desarrollo bipotencial y pueden dar lugar tanto a las células endoteliales como a las hematopoyéticas. Una vez que las células se han comprometido con cualquiera de los dos linajes, las células hijas de los hemangioblastos pierden su capacidad de formar cualquier otro tipo de células.
Hematopoyesis embrionaria
Aunque la formación de las células sanguíneas (hematopoyesis) comienza en el saco vitelino. las evidencias obtenidas en experimentación animal sugieren que las células procedentes del saco vitelino pronto son remplazadas por otras células sanguíneas que se derivan de forma independiente a partir de focos intraembrionarios de hematopoyesis
Los islotes sanguíneos contienen células hematopoyéticas madre (stem), que pueden dar origen a todos los tipos de células que se encuentran en la sangre embrionaria. Los eritrocitos producidos en el saco vitelino son grandes células nucleadas que entran en el torrente circulatorio poco antes de que el tubo cardiaco comience a latir aproximadamente a los 22 días de gestación Durante las primeras seis semanas los eritrocitos circulantes son casi en su totalidad derivados del saco vitelino, pero durante ese tiempo está ocurriendo la preparación para las siguientes etapas de la hematopoyesis.
El análisis de los embriones de pollo y de ratón ha demostrado que la hematopoyesis intraembrionaria definitiva comienza en pequeños cúmulos de células (cúmulos paraórticos) situados en el mesodermo esplacnopleural asociado con la pared ventral de la aorta dorsal y poco después en la región aorta /cresta genital/mesonefros. Hacia las semanas 5 ó 6, los focos de hematopoyesis son muy importantes en el hígado.
Los eritrocitos producidos por el hígado son bastante diferentes de los derivados del saco vitelino. Aunque todavía son bastante más grandes que los glóbulos rojos del adulto. los eritrocitos hepáticos son anucleados y contienen diferentes tipos de hemoglobina. Hacia las semanas 6 a 8 de la gestación humana, el hígado sustituye al saco vitelino como la principal fuente de glóbulos rojos. Si bien el hígado continúa produciendo eritrocitos hasta el comienzo del periodo neonatal, su contribución comienza a disminuir durante el sexto mes del embarazo. En este momento la formación de las células sanguíneas se desplaza a la médula ósea, el lugar definitivo de la hematopoyesis adulta. Este desplazamiento se encuentra bajo el control del cortisol segregado por la corteza suprarrenal del feto. En ausencia de cortisol. la hematopoyesis se queda confinada al hígado. Antes de que la hematopoyesis se haya establecido del todo en la médula ósea, también puede haber la formación de pequeñas cantidades de sangre en el epiplón y posiblemente en el bazo.
Aspectos celulares de la hematopoyesis
Las primeras células madre {stem) hematopoyéticas que surgen en el embrión son pluripotenciales ya que pueden dar origen a todos los tipos de células que se encuentran en la sangre (figura 16-1). Estas células pluripotenciales. que en ocasiones se llaman hemocitoblastos, tienen una enorme capacidad de proliferación. Producen gran cantidad de células hijas, la mayoría de las cuales son células que se encuentran en la siguiente etapa de diferenciación, pero también producen pequeñas cantidades de células de su mismo tipo (stem), que actúan a manera de reserva capaz de reponer los linajes celulares individuales en caso de necesidad. Al comienzo del desarrollo el linaje celular primario se divide en dos linajes separados. Las células progenitoras linfoides terminan por formar las dos líneas de linfocitos: los linfocitos B (responsables de la producción de anticuerpos) y los linfocitos T (responsables de las reacciones de la inmunidad celular). Las células progenitoras mieloides son precursoras de otras líneas de células sanguíneas: eritrocitos, granulocitos (neutrófilos, eosinófilos y basófilos), monocitos v plaquetas. La segunda generación de células progenitoras (linfoides y mieloides) todavía son pluripotenciales, aunque su potencial está restringido, ya que ninguna de ellas puede formar la progenie del otro tipo.
Con base en el comportamiento que muestran en ciertas situaciones experimentales, las células progenitoras hematopoyéticas se denominan en ocasiones unidades formadoras de colonias (UFC). La célula progenitora de primera generación se llama UFC-ML porque puede dar origen tanto a las líneas celulares mieloides como a las linfoides. Las células progenitoras de la segunda generación se llaman UFC-L (L. linfocitos) y UfC-S (S, bazo) (determinadas a partir de experimentos en los cuales se estudió la diferenciación de las células progenitoras en bazos irradiados). En todos los casos excepto uno, las descendientes de las UFC-L y las UFC-S son células progenitoras determinadas, ya que tienen la capacidad de formar sólo un tipo de célula sanguínea madura. En cada linaje los tipos celulares que se forman han de pasar por varias fases de diferenciación antes de alcanzar su fenotipo maduro.
¿Qué controla la diversificación de las células progenitoras en líneas celulares especificas? Los experimentos que comenzaron en el decenio de 1970 arrojaron pruebas de la existencia de factores estimulantes de colonias (FEC) específicos para cada línea de células sanguíneas. Los FEC son proteínas difusibles que estimulan la proliferación de las células progenitoras hematopoyétieas. Algunos FEC actúan sobre varios tipos de células progenitoras, mientras que otras estimulan sólo un tipo. Aunque queda mucho por conocer sobre los lugares de origen y los modos de acción de los FEC, muchos parecen producirse dentro del ámbito local de las células estromales de la médula ósea, y algunos pueden ser almacenados en la matriz extracelular local. Los FEC se Fijan a un pequeño número de receptores localizados en la superficie de sus células diana. Desde el punto de vista funcional, los FEC representan mecanismos para estimular la expansión de tipos específicos de células sanguíneas en caso de necesidad. El reconocimiento de la existencia de los FEC ha concitado enorme interés por su posible aplicación clínica en ciertas afecciones caracterizadas por una deficiencia de leucocitos (leucopenia).
Ciertos genes Hox, en especial los de las familias Hoxa y Hoxb, desempeñan un papel importante en algunos aspectos de la hematopoyesis. La exposición de la médula ósea a los oligonucleótidos contra los genes específicos Hox da como resultado la supresión de las líneas especificas de diferenciación de las células sanguíneas. Por el contrario, la sobreexpresión creada por ingeniería genética de los genes hoxb-8.hoxa-9 y Hoxa-10 causa leucemia en ratones. Hay evidencias crecientes que apuntan hacia la participación de genes Hox en la patogénesis de las leucemias humanas. Una función importante de los genes Hox en la hematopoyesis es la regulación de la proliferación.
El linaje de los eritrocitos representa una línea de descendientes de las células UFC-S. Si bien las células progenitoras de la línea eritroide (proeritrocitos) están restringidos a formar sólo glóbulos rojos, existen previamente muchas generaciones de células precursoras (figura 16-2). Algunas de ellas se han reconocido hace poco tiempo y las funciones de muchas de las etapas de las células precursoras están comenzando a dilucidarse.
Las primeras etapas de la eritropoyesis se reconocen por el comportamiento de las células precursoras en cultivo y no por diferencias morfológicas o bioquímicas. Estas se llaman unidades formadoras de brotes eritroides (UFB-E) y unidades formadoras de colonias eritroides (UFC-E). Cada una de ellas responde a diferentes factores estimuladores. Las UFC-S precursoras pluripotenciales (figura 16-1) responden a la interleucina-3, un producto de los macrófagos de la médula ósea del adulto. una hormona denominada burst-promoting activity (BPA) estimula la mitosis de los precursores UFB-E (figura 16-2). Una célula UFC-E, que tiene una capacidad de proliferación menor que la de la célula UFB-E, requiere la presencia de eritropoyetina como factor estimulante. La eritropoyetina es una glucoproteina que estimula la síntesis del ácido ribonucleico mensajero (RNA-m) de la globina y se produce primero en el hígado fetal. Más adelante en el curso del desarrollo, la síntesis pasa al riñón, que sigue siendo el lugar de la producción de la eritropoyetina en el adulto. En condiciones de hipoxia (por ejemplo, después de la pérdida de sangre o en las grandes alturas) aumenta la producción de eritropoyetina en los riñones, y esto estimula la producción de más glóbulos rojos para tratar de compensar el aumento de las necesidades. En la eritropoyesis del adulto la etapa de UFC-E parece ser la que más responde a las influencias ambientales. La placenta parece ser impermeable a la eritropoyetina, con lo que aísla al embrión del cambio de los niveles de eritropoyetina en la madre y también se evita la influencia de la eritropoyetina fetal en el sistema de formación sanguínea de la madre.
Una o dos generaciones después de la etapa de UFC-E, pueden reconocerse por su morfología sucesivas generaciones de células precursoras de los eritrocitos. La primera etapa reconocible es la de los proeritroblastos (figura 16-3), una célula grande y muy basófila que todavía no ha producido suficiente hemoglobina para que pueda ser detectada por análisis citoquímicos. Esta célula tiene un nucléolo grande, la cromatina nuclear muy poco condensada, numerosos ribosomas y una alta concentración de RNA-m de globina. Estas son las características citológicas clásicas de una célula indiferenciada.
Las sucesivas etapas de la diferenciación eritrocitaria (eritroblastos basofílicos, policromatófilos y ortocromáticos) se caracterizan por un cambio progresivo del equilibrio entre la acumulación de la hemoglobina recién sintetizada y la involución primero de la maquinaria de producción de RNA y después del aparato de síntesis proteica. El tamaño global de la célula disminuye y el núcleo se torna cada vez más picnótico (más pequeño y con la cromatina nuclear más condensada) hasta que por último es expulsado en la etapa del eritroblasto ortocromático. Después de perder el núcleo y la mayor parte de los organelos citoplásmicos, el glóbulo rojo inmaduro, que aún contiene un pequeño número de polisomas, es un reticulocito. Los reticulocitos pasan a la sangre circulante, donde continúan produciendo pequeñas cantidades de hemoglobina durante uno o dos días.
La etapa final de la eritropoyesis es el eritrocito maduro que es una célula terminal diferenciada a causa de la pérdida del núcleo y de la mayor parte de sus organelos citoplásmicos. Los eritrocitos embrionarios son más grandes que los del adulto y tienen una vida media más corta (50 a 70 días en el feto contra 120 días en el adulto).
Síntesis de hemoglobina y su control
Tanto los eritrocitos como la hemoglobina que están dentro de él experimentan transiciones isomórficas durante el desarrollo embrionario. La molécula de la hemoglobina es un complejo compuesto por el hem y cuatro cadenas de globina: dos cadenas alfa y dos beta. Tanto las subunidades a como las b son productos de genes localizados en los cromosomas 16 y 11, respectivamente (figura 16-4). Diferentes isoformas de las subunidades se codifican de forma lineal en estos cromosomas.
Durante el período de la hematopoyesis en el saco vitelino, se producen isoformas de globina embrionaria. La hemoglobina embrionaria más primitiva, que en ocasiones se llama hemoglobina Gower-1, está compuesta por dos cadenas C (tipo a) y dos E (tipo b). Después de pasar por un par de formas transicionales (tabla 16-1 no está), hacia las doce semanas la síntesis de hemoglobina entra en la etapa fetal y viene a corresponder al desplazamiento del lugar de la eritropoyesis desde el saco vitelino hacia el hígado. La hemoglobina fetal está compuesta por dos cadenas a de tipo adulto, que se forman muy pronto en la embriogénesis, y dos cadenas y, la principal isoforma fetal de la cadena b. La hemoglobina fetal es la forma predominante durante el resto del embarazo. El principal valor adaptativo de la isoforma fetal de la hemoglobina es que su afinidad por el oxígeno es mayor que la de la hemoglobina adulta. Esto tiene ventajas para el feto, que depende de la concentración de oxigeno de la sangre materna. A partir de la semana 30 de embarazo, se produce un gradual desplazamiento desde el tipo fetal de hemoglobina hacia el adulto, de la que a2b2 es el tipo predominante. Una variante menor, pero similar desde el punto de vista funcional es a2b2
Formación de los vasos sanguíneos embrionarios
El embrión en su fase inicial está desprovisto de vasos sanguíneos. Aunque aparecen islotes sanguíneos en la pared del saco vitelino y asociados a ella se forman conductos vasculares extraembrionarios (figura 5-24), gran parte de la vascularización del cuerpo del embrión tienen un origen intraembrionario. Durante la formación inicial de los somitas, aparecen rápidamente redes de pequeños vasos en muchas regiones del cuerpo embrionario.
Algunos conductos vasculares confluyen para formar vasos más grandes, otros siguen siendo similares a los capilares y otros desaparecen. Un principio fundamental en la vasculogénesis inicial es el cambio dinámico relacionado con el crecimiento de las estructuras con las cuales están asociados los conductos vasculares. La denominación de los vasos iniciales como arterias y venas no está determinada en la fase inicial del desarrollo, ya que la dirección del flujo de la sangre en un canal dado puede invertirse con facilidad. Aunque el principal patrón de los conductos vasculares es reconocible en cada una de las especies, la forma aparentemente aleatoria de reclutamiento y absorción de nuevos capilares para que se conviertan en vasos más grandes explica en principio las frecuentes variaciones menores del patrón vascular de un individuo. La frecuencia de las variaciones anatómicas es muy importante en el sistema venoso.
Los estudios descriptivos detallados y los experimentos de trasplante en que se han usado marcadores intracelulares o marcadores de anticuerpos monoclonales específicos del injerto han demostrado que los angioblastos (precursores de las células endoteliales) se originan en los tejidos mesodérmicos del cuerpo con la excepción del mesodermo de la notocorda y del precordal (tabla 16-2 no está). Los vasos sanguíneos embrionarios se forman a partir de los angioblastos mediante tres mecanismos principales. Muchos de los vasos sanguíneos más grandes, como las aortas dorsales, se forman por la confluencia de los angioblastos in situ. Otros conductos igualmente grandes, como los del endocardio, se forman por angioblastos que migran hacia la región procedentes de otros lugares. Otros vasos, en especial los vasos íntersegmentarios del eje principal del cuerpo y los vasos del sistema nervioso central, se originan como brotes vasculares a partir de vasos más grandes ya formados. Muchos de los angioblastos del tronco al inicio proceden del mesodermo esplácnico.
Las paredes de los vasos sanguíneos de la mayor parte del tronco y de las extremidades derivan del mesodermo local que llega a continuarse con el endotelio de los vasos más grandes. En la cabeza y en muchas áreas del sistema de los arcos aórticos, el mesénquima derivado del ectodermo de la cresta neural constituye partes importantes de las paredes vasculares (por ejemplo, las células musculares lisas), pero la cresta neural no da origen a células endoteliales.
En la construcción de las paredes vasculares parece intervenir una señal molecular de doble vía. Según uno de los modelos, las células del mesénquima perivascular producen una molécula de señal llamada angiopoyetina-1, que se fija sobre las superficies de las células endoteliales a un receptor de tirosincinasa, el Tic-2. Esto estimula a las células endoteliales a liberar sus propias moléculas de señales, como el factor de crecimiento derivado de las plaquetas (PDGF), que estimula la migración de células mesenquimatosas hacia el endotelio vascular. Una vez establecido el contacto con las células mesenquimatosas, las células endoteliales liberan el factor b de transformación del crecimiento (TGF-b), que estimula la diferenciación de las células mesenquimatosas en células musculares lisas o pericitos.
Como en el caso con los mioblastos, los angioblastos parecen reaccionar a las claves ambientales locales, las cuales determinan un patrón morfológico especifico del vaso sanguíneo. Los estudios marcaje de angioblastos trasplantados han demostrado que algunos de ellos pueden migrar a largas distancias. Los angioblastos que han migrado lejos del lugar en el que fueron injertados terminan integrándose en vasos sanguíneos de morfología normal en las áreas donde se establecen.
Hay factores locales que también influyen sobre la iniciación de la vasculogénesis. En algunos órganos (por ejemplo, el hígado) o partes de órganos (por ejemplo, los bronquios del sistema respiratorio), los vasos arteriales que irrigan las regiones se originan en el mesodermo local, mientras que otros órganos (por ejemplo, los riñones metanéfricos) o partes de órganos (por ejemplo, los alvéolos pulmonares) son irrigados por vasos sanguíneos que crecen hacia el mesénquima procedentes de otros tejidos. En este último tipo de mecanismo de vascularización, hay pruebas crecientes de que los primordios de los órganos producen factores de angiogénesis que estimulan el crecimiento de los brotes vasculares (porque promueven la mitosis de las células endoteliales) hacia el mesénquima glandular. Muy pocos de los factores a los cuales se ha atribuido un papel angiogenético tienen determinada su composición molecular, pero se ha demostrado que algunas formas del factor de crecimiento de los fibroblastos y del factor de crecimiento del endotelio vascular tienen actividad angiogénica en varias estructuras en desarrollo.
Desarrollo de las arterias. Arcos aórticos y sus derivados
El sistema de los arcos aórticos en los estadios iniciales del embrión humano se organiza siguiendo los mismos principios del sistema arterial que lleva la sangre a las branquias de muchos de los vertebrados acuáticos inferiores. La sangre sale del ventrículo común del corazón hacia una raíz aórtica ventral, y llega a las arterias branquiales a través de pares de arcos aórticos (figura 16-5 A). En los vertebrados que tienen branquias las arterias de los arcos aórticos se ramifican en lechos capilares, donde la sangre se vuelve a oxigenar conforme pasa a través de las branquias. En los embriones de los mamíferos los arcos aórticos siguen siendo vasos gruesos porque el intercambio gaseoso tiene lugar en la placenta y no en los arcos faríngeos. Los arcos aórticos desembocan en el par de aortas dorsales en donde la sangre entra en la circulación sistémica regular. En los embriones humanos, no todos los arcos aórticos están presentes al mismo tiempo. Su formación y remodelación se produce según un gradiente craneocaudal muy marcado. La sangre proveniente del tracto de salida del corazón (la región troncoconal) fluye a continuación dentro del saco aórtico, que difiere de la región troncoconal en la constitución de su pared. Los arcos aórticos se originan a partir del saco aórtico.
La anatomía del desarrollo del sistema de los arcos aórticos ilustra bien el principio de la adaptación morfológica del lecho vascular durante las diferentes etapas de la embriogénesis (tabla 16-3 no está). El continuo desarrollo de las regiones craneal y cervical hace que los componentes de los tres primeros arcos y las raíces aórticas asociadas con ellos se remodelen y forman el sistema de la arteria carótida (figura 16-5). Con la remodelación del tubo cardiaco y la división interna del tracto de salida en los componentes aórtico y pulmonar, los cuartos arcos experimentan una adaptación asimétrica a la asimetría inicial del corazón. El cuarto arco aórtico izquierdo persiste formando un canal grande (arco de la aorta) que transporta todo el volumen del ventrículo izquierdo del corazón. El cuarto arco derecho se incorpora a la arteria subclavia derecha,
Los textos de embriología tradicionalmente han descrito el sistema de arcos aórticos como conformado por seis pares de arcos vasculares, pero el quinto y el sexto arcos nunca aparecen como conductos vasculares definidos similares a los cuatro primeros arcos. El quinto arco aórtico, si es que existe en realidad, se representa por unas pocas asas capilares. El sexto (arco pulmonar) surge como un plexo capilar asociado con la tráquea primitiva y las yemas pulmonares. El plexo capilar está irrigado por arterias segmentarias ventrales que provienen del par de aortas dorsales en esa región (figura 16-6). El equivalente del sexto arco se representa por un segmento distal definido (arteria segmentaria ventral) conectada con la aorta dorsal a través de un segmento proximal plexiforme que establece la conexión entre el saco aórtico a nivel del origen del cuarto arco y el componente segmentario distal. Conforme el divertículo respiratorio y las yemas pulmonares iniciales se alargan, partes de la red de capilares pulmonares se consolidan para formar un par de arterias pulmonares definidas que conectan con el denominado sexto arco. Aunque suele usarse el término sexto arco aórtico en la bibliografía anatómica y clínica, el término más apropiado es el de arco pulmonar ya que no implica equivalencia alguna con los demás arcos aórticos.
Al igual que el cuarto arco aórtico, el arco pulmonar se desarrolla de forma asimétrica. El lado izquierdo se convierte en un conducto grande. Su segmento distal, que se ha derivado de la arteria segmentaria ventral, persiste en la forma de un conducto grande (ductus arterioso) que desvía la sangre desde la arteria pulmonar izquierda hacia la aorta (figura 16-5 C). Gracias a esta derivación los pulmones están protegidos de un flujo de sangre más grande del que su red vascular puede soportar durante la mayor parte del periodo intrauterino. En el lado derecho el segmento distal del arco pulmonar involuciona y el segmento próxima (la base de la arteria pulmonar derecha) se ramifica a partir de! tronco pulmonar.
La asimetría de los derivados de los arcos pulmonares explica la diferencia en el trayecto de los nervios recurrentes laríngeos izquierdo y derecho, que son ramas del nervio vago (X par craneal). Estos nervios que inervan la laringe, forman un arco alrededor de los arcos pulmonares. Conforme el corazón desciende hacia la cavidad torácica desde la región cervical, la rama del vago y por ello cada nervio recurrente laríngeo se desplaza de igual manera. El nervio del lado izquierdo se asocia con el ductus arterioso (figura 16-5 C), que persiste durante toda la vida fetal, de modo que es llevado más profundo dentro de la cavidad torácica. En el lado derecho, en virtud de la regresión de la mayor parte del arco pulmonar de ese lado, el nervio asciende hasta el nivel del cuarto arco, que constituye una barrera anatómica. Las posiciones de los nervios recurrentes laríngeos derecho e izquierdo en el adulto reflejan esta asimetría, ya que el nervio derecho se incurva por debajo de la arteria subclavia derecha (cuarto arco) y el izquierdo lo hace alrededor del ligamento arterioso (el derivado adulto del ductus arterioso, el segmento distal del cuarto arco pulmonar izquierdo).
Ramas principales de la aorta
En el embrión joven, cuando las aortas dorsales son todavía vasos pares, se desprenden de ellas tres grupos de ramas arteriales: intersegmentaria dorsal, segmentaria lateral y segmentaria ventral (figura 16-7). Estas ramas experimentan diversas modificaciones en su forma antes de adoptar su configuración adulta (tabla 16-4 no está). Las arterias segmentarias ventrales se originan como un par de vasos que corren sobre las paredes dorsal y lateral del intestino y el saco vitelino. Con el cierre del intestino y el estrechamiento del mesenterio dorsal, ciertas ramas se fusionan en la línea media para formar el tronco celiaco y las mesentéricas superior e inferior.
Las arterias umbilicales comienzan como ramas segmentarias ventrales puras que irrigan al mesodermo de la alantoides, pero más adelante su raíz se conecta con los vasos intersegmentarios lumbares. Entonces los vasos umbilicales más proximales regresan y las ramas intersegmentarias se convierten en las ramas principales de la aorta. A semejanza de sus homólogas arterias subclavias en los brazos, las ramas arteriales, al comienzo pequeñas (arterias ilíacas). que irrigan las yemas de las piernas aparecen como ramas de la ramas intersegmentarias dorsales (lumbares) de la aorta. No obstante, después de que las arterias umbilicales engloban los segmentos proximales de los vasos intersegmentarios, las arterias iliacas parecen nacer como ramas de las arterias umbilicales.
Arterias de la cabeza
Las arterias que irrigan la cabeza se originan en dos fuentes. En situación ventral, el sistema de los arcos aórticos (primero a tercero y sus raíces correspondientes) da origen a las arterias que irrigan la cara (arterias carótidas externas) y la parte frontal de la base del cerebro (arterias carótidas internas) (figura 16-5). nivel de la médula espinal, las arterias vertebrales. que se forman a través de conexiones de las ramas laterales de las seis primeras arterias intersegmentarias dorsales, crecen en dirección al cerebro. Pronto giran hacia la línea media y se fusionan, dando lugar a la arteria basilar única (figura 16-8). Esta, corre a lo largo de la superficie ventral del tronco cerebral, al que irriga una serie de arterias pares. Conforme la arteria basilar y las carótidas internas se aproximan al nivel del diencéfalo, ramas de cada uno de estos vasos principales crecen y se fusionan, formando las arterias comunicantes posteriores, que unen las circulaciones de las arterias basilar y carótida interna. Otras dos pequeñas ramas del sistema de la carótida interna se anastomosan en la línea media para completar un anillo vascular (polígono de Willis), situado en la base del diencéfalo y que encierra al quiasma óptico y al tallo de la hipófisis. El polígono de Willis es una adaptación estructural que asegura un abastecimiento continuo de sangre en caso de que se ocluya alguna de las arterias principales que irrigan el cerebro.
Arterias coronarias
Aunque por intuición se esperaría que las arterias coronarias surgiesen como ramas de la aorta, los estudios experimentales han demostrado que las células endoteliales precursoras de las arterias coronarias surgen del mismo primordio celular que el futuro epicardio, migran hacia la aorta e invaden su pared. Las células musculares lisas de los vasos coronarios tienen un único origen mesodérmico puro en vez de un origen mixto de la cresta neural y del mesodermo como se observa en los derivados de los arcos aórticos. Los estudios en que se han marcado las células con marcadores retrovirales les confirman que las células progenitoras de los vasos coronarios penetran en la pared del corazón que ya late cuando la capa epicardica envuelve al miocardio y que después de formarse in situ, las arterias coronarias conectan secundariamente con la aorta.
Desarrollo de las venas
Las venas siguen un complejo patrón de desarrollo morfológico que se caracteriza por la formación de redes muy irregulares de capilares y el crecimiento posterior de ciertos conductos que se convierten en venas definitivas. A causa de los comienzos de múltiples conductos y las numerosas posibilidades de evolución (Figura 16-9), el sistema venoso del adulto se caracteriza por una mayor incidencia de variaciones anatómicas que el sistema arterial. Una descripción detallada del desarrollo de los vasos venosos sobrepasa los alcances de este libro.
Venas cardinales
Las venas cardinales constituyen la base de la circulación venosa intraembrionaria Aparecen varios grupos de venas cardinales en diferentes tiempos y lugares. En cualquier grupo de venas cardinales, algunos segmentos involucionan, y otros persisten ya sea como conductos independientes o como componentes de venas complejas que incluyen también porciones de otras venas cardinales.
El patrón mas primitivo de las venas cardinales consta de las venas cardinales anteriores y posteriores pares que drenan la sangre de la cabeza y del cuerpo hacia un par de cortas venas cardinales comunes (figura 16-9). Éstas últimas a su vez vacían su sangre en el seno venoso del corazón primitivo (figura 16-11).
En la región craneal las venas cardinales anteriores, que en sus orígenes eran simétricas, se transforman en las venas yugulares internas (figura 16-10). Conforme el corazón rota hacia la derecha, la base de la vena yugular interna izquierda se adelgaza. Al mismo tiempo un nuevo conducto anastomótico. que a la postre forma la vena braquicefálica izquierda, conecta la vena yugular interna izquierda con la derecha. A través de esta anastomosis la sangre que viene del lado izquierdo de la cabeza drena hacia la vena cardinal anterior derecha original, que acaba por convertirse en la vena cava superior, y que desemboca en la aurícula derecha del corazón. Entre tanto, la parte próximal de la vena cardinal y común izquierda persiste en la forma de un pequeño vaso, el seno coronario, que será la vía final de drenaje de las venas coronarias, hacía la aurícula derecha del corazón.
En el tronco surge un par de venas subcardinales en asociación con el mesonefros en desarrollo. Las venas subcardinales están conectadas con las venas cardinales posteriores y entre si a través de numerosas anastomosis. Tanto las venas poscardinales como las subcardinales drenan los riñones mesonéfricos través de numerosas ramas laterales pequeñas. Conforme los riñones mesonéfricos comienzan su regresión. las venas que los drenan también comienzan a regresar. En este momento aparece un par de venas supracardinales en la pared del cuerpo situado dorsalmente a las venas subcardinales. Más adelante, los tres grupos de venas cardinales del cuerpo involucionan en grados variables según las zonas, y los restos que permanecen constituirán a la vena cava inferior. Esta última forma un vaso único asimétrico que corre paralelo y a la derecha de la aorta (figura 16-9). La mayoría de las venas de las cavidades abdominal y torácica se derivan de la persistencia de segmentos del sistema de las venas cardinales.
Venas umbilicales y vitelinas
Las venas umbilicales y vitelinas extraembrionarias se originan como unos pares de vasos simétricos que drenan por separado en el seno venoso del corazón (figura 16-11) Con el tiempo, estos vasos se asocian estrechamente con el hígado que está en su fase de crecimiento rápido. Las venas vitelinas, que drenan el saco vitelino, desarrollan grupos de conductos anastomóticos tanto dentro como fuera del hígado. Fuera de él. las dos venas vitelinas y sus conductos anastomóticos paralelos se relacionan con el duodeno. A través de la persistencia de algunos conductos y la desaparición de otros se forma la vena porta hepática, que drena los intestinos. Dentro del hígado el plexo vitelino se transforma en un lecho capilar, que permite una distribución amplia de los materiales alimenticios que son absorbidos en el intestino hacia las partes funcionales del hígado. Desde el lecho capilar hepático la sangre que llega del sistema de la vena porta pasa hacia un grupo de venas hepáticas que vacían la sangre en el seno venoso.
Las venas umbilicales, que en sus orígenes son simétricas, pronto pierden sus segmentos hepáticos y termina directamente en el hígado al combinarse con el plexo vascular intrahepático de las venas vitelinas. Pronto se forma un canal grande, el ductus venoso, que deriva gran parte de la sangre que entra por la vena umbilical izquierda directamente y a través del hígado hacia la vena cava inferior. El ductus venoso es una importante adaptación que permite el mantenimiento del patrón funcional embrionario de la circulación sanguínea. Pronto la vena umbilical derecha se degenera, lo cual conviene la vena umbilical izquierda en el único canal que lleva la sangre que se ha re oxigenado y purificado en la placenta de vuelta al cuerpo del embrión. El ductus venoso permite que la sangre placentaria oxigenada que entra pase de largo sin distribuirse por las redes capilares del hígado y que se distribuya en los órganos (por ejemplo, el cerebro y el corazón) que más la necesitan.
Las venas pulmonares desde el punto de vista filogenético son las estructuras más recientes y se forman independiente en vez de asumir porciones de los sistemas más antiguos de las venas cardinales. Provenientes de cada pulmón. los canales venosos de drenaje convergen hasta que terminan formando una sola vena pulmonar común de gran tamaño que desemboca en la aurícula izquierda del corazón. Conforme la aurícula se expande, la vena pulmonar común se incorpora en su pared (figura 16-12). La absorción incorpora a la aurícula las dos primeras ramas de las venas pulmonares originales, lo que da como resultado la entrada de cuatro venas independientes en la aurícula izquierda.
Desarrollo de los conductos linfáticos
El origen de los conductos linfáticos no se conoce del todo; una teoría sostiene que esta en el endotelio venoso y otros lo atribuyen al mesodermo local. Cualquiera que sea su origen. el sistema linfático hace su aparición en la forma de seis sacos linfáticos primarios al final de la sexta semana del embarazo (figura 16-13). En el ángulo formado por las venas precardinales (futura yugular interna) y las venas subclavias aparecen dos sacos linfáticos yugulares. En el abdomen se forma un saco linfático retroperitoneal en la pared posterior del cuerpo en la raíz del mesenterio durante la octava semana. Algo más tarde aparece la cisterna del quilo en el mismo nivel pero en situación dorsal a la aorta. Más o menos al mismo tiempo surge un par de sacos linfáticos posteriores en la bifurcación de las venas femoral y ciática. Hacia el final de la novena semana aparecen los vasos linfáticos que conectan estos sacos.
Dos vasos linfáticos principales conectan la cisterna del quilo con los sacos linfáticos yugulares. Se forma una anastomosis entre estos dos conductos. Un vaso linfático único compuesto por la parte caudal del canal derecho, el segmento anastomótico, y la parte craneal del conducto izquierdo acaba por convertirse en el conducto torácico definitivo del adulto. Este conducto drena la linfa de la mayor parte del cuerpo y del lado izquierdo de la cabeza en el sistema venoso a nivel de la unión de las venas yugular interna y subclavia. El conducto linfático derecho, que drena el lado derecho de la cabeza y la parte superior del tórax y el brazo derecho, también desemboca en el sistema venoso en la localización original del saco yugular derecho.
DESARROLLO Y TABICACIÓN DEL CORAZÓN
Desarrollo inicial del corazón
Orígenes celulares
El corazón tubular inicial (figura 5-23) se deriva de varias fuentes tisulares. El tejido miocardio auricular y ventricular se origina en un primordio bilateral de mesodermo esplácnico en forma de herradura (figura 5-21) que se fusiona en la línea media ventral para formar la masa inicial del tubo cardiaco primario. El tracto de salida tiene un origen celular bastante diferente, ya que los componentes endoteliales provienen del mesodermo cefálico paraxial y lateral de la región de la placoda ótica. Uno de los principales componentes celulares de la pared del tracto de salida deriva de la cresta neural craneal, más exactamente de la del nivel situado entre la mitad de la placoda ótica y el extremo caudal del tercer somita (figura 11-9). Estos componentes craneales se integran con los primordios cardiacos bilaterales mientras se encuentran en la región cervical. Conforme el corazón desciende hacia la cavidad torácica, las células del tracio de salida derivadas de la región craneal lo acompañan.
Formación del asa
Cuando al final de la tercera semana el tubo cardiaco se forma. tiene una simetría bilateral (figura 5-23). Pronto experimenta un plegamiento característico hacia la derecha, por lo cual es la primera estructura asimétrica que aparece en el cuerpo del embrión (la base molecular inicial de la asimetría del embrión se discute en la Pág.73) Además de estas cascadas moleculares, ciertos genes cardiacos específicos también parecen intervenir en la formación inicial del asa del tubo cardiaco. Los bloqueos de tres tipos de factores de transcripción cardiacos (Nkx-2. MEF-2 y d-HAND y eHAND) se caracterizan por la detención del desarrollo cardiaco en la etapa de formación del asa. La primera indicación molecular del desarrollo asimétrico del tubo cardiaco es el desplazamiento en la expresión del factor de transcripción e-Hand desde ambos lados hacia el lado caudal izquierdo del tubo cardiaco. El d-Hand se expresa de forma predominante en el primordio del ventrículo derecho. Las moléculas hand pueden desempeñar un papel en la interpretación de las señales moleculares asimétricas primitivas y en la expresión de esta información en las células, lo que da como resultado la formación del asa.
La caracterización de la base celular para la formación del asa cardiaca ha resultado muy difícil. Aunque se ha demostrado claramente que la formación del asa es una propiedad intrínseca del tubo cardiaco en desarrollo. para lograr este efecto se han postulado factores tan diversos como microtúbulos, haces de actina localizados asimétricamente, presión de la gelatina cardiaca y cambios individuales en la forma de las células miocárdicas.
El resultado de la formación del asa cardiaca es un corazón en forma de S, en el cual la parte del flujo de entrada que inicialmente es caudal (aurícula) termina colocada en la región situada dorsal del tracto de salida. Éste último recibe el nombre de bulbus cordis (figura 16-14). En la formación inicial del asa la rama craneal de la S representa el bulbus cordis y la rama media representa la parte ventricular del corazón, en la que el primordio del ventrículo derecho esta más próximo al tracto de salida y el del ventrículo izquierdo está próximo a la rama caudal. La rama caudal es la aurícula común, que algo más tarde crece hacia ambos lados (figura 16-15). y un tabique interno comienza a dividir el ventrículo común en una cámara derecha y una izquierda. El tracto de salida (bulbus cordis del corazón inicial) conserva su apariencia tubular macroscópica. Su parte distal. que se confirma con el sistema de los arcos aórticos, se denomina tronco arterioso. El corto segmento de transición situado entre el tronco y el ventrículo se llama cono arterioso. El cono está separado de los ventrículos por unos surcos poco marcados.
División inicial del canal aurículo-ventricular del corazón
Al comienzo del desarrollo cardiaco la aurícula se separa parcialmente del ventrículo mediante la formación de unos cojinetes engrosados aurículo-ventriculares. En la unión entre el ventrículo y el tracto de salida se forma un engrosamiento similar pero menos pronunciado (figura 16-16). En estas áreas la gelatina cardiaca, que esta organizada como una membrana basal gruesa, se hernia hacia el canal aurículo-ventricular. Los cojinetes endocárdicos funcionan como válvulas primitivas que ayudan en la propulsión de la sangre hacia delante.
En respuesta a una acción inductiva ejercida por el miocardio subyacente. algunas células procedentes de dos áreas circunscritas del endocardio pierden su carácter epitelial y se transforman en células mesenquimatosas que migran hacia la gelatina cardiaca. Sólo en la región aurículo-ventricular y en el tracto de salida proximal las células endocardiacas responden a la inducción y se transforman en mesenquimatosas. Las células endocárdicas auriculares y ventriculares no pierden su carácter epitelial. El corazón primitivo está segmentado desde el punto de vista molecular. Los segmentos del endocardio que experimentan una transformación mesenquimatosa expresan el gen Msx-1, mientras que las células endocárdicas de la aurícula y el ventrículo no lo hacen. Las células endocárdicas contienen la molécula de adhesión celular N-CAM en sus superficies. Aquellas células que se transforman en mesénquima disminuyen la producción de la N-CAM. Se supone que esto facilita su conversión en células móviles.
los estudios in vivo han demostrado que poco antes de la transformación de las células endocárdicas en mesénquima, aparecen en la gelatina cardiaca unas partículas de 20 a 50 nm de diámetro por debajo de estas células pero no bajo otras áreas del endocardio (figura 16-16). Estas partículas, que son producidas por las células miocárdicas subyacentes y que se llaman adherones. contienen un complejo molecular de proteoglicano. fibronectina y varias proteínas de la matriz. Una de estas proteínas,. la ES-30 también se expresa en otros tejidos (por ejemplo, la notocorda, el ectodermo del tubo neural. La cresta ectodérmica apical), que son los únicos otros tejidos que pueden sustituir al miocardio e inducir a la transformación de epitelio del endocardio en mesénquima. Estas partículas pueden inducir in Vitro a las células endoteliales aurículo-ventriculares a transformarse en mesénquima. La inducción de la invasión de la gelatina cardiaca por las células del endotelio aurículo-ventricular también se acompaña de la actividad del TGF-b1 y el TGF-b3. Si este factor de crecimiento se inactiva. la transformación no se produce. Las células mesenquimatosas transformadas secretan proteasas, que pueden destruir los adherones inductivamente activos y restablecer la estabilidad morfogenética de las regiones de los cojinetes endocárdicos.
Estos acontecimientos celulares y moleculares constituyen la base inicial de la formación de las principales válvulas cardiacas. Los trastornos en estos procesos pueden explicar muchas de las malformaciones cardiacas.
Tabicación tardía del corazón
Separación de las aurículas de los ventrículos
Los cojinetes endocárdicos (figura 16-16) que posteriormente se convertirán en tejido conectivo denso. se forman en las paredes dorsal y ventral del canal aurículo-ventricular. Conforme crecen hacia el canal. Los dos cojinetes se unen y separan el canal aurículo-ventricular en canales derecho e izquierdo (figuras 16-17 y 16-18). Los cojinetes endocárdicos en su fase inicial sirven como válvulas primitivas que ayudan a propulsar la sangre hacia delante a través del corazón. Posteriormente en el desarrollo. En el canal aurículo-ventricular aparecen unas delgadas hojas de las válvulas anatómicas. Las hojas valvulares definitivas no parecen provenir del tejido del cojinete endocárdico tanto como de una investigación de los tejidos superficiales derivados del epicardio del surco aurículo-ventricular. La válvula que protege el canal aurículo-ventricular derecho desarrolla tres hojas (válvula tricúspide) pero la válvula del canal izquierdo (mitral o bicúspide) solo desarrolla dos hojas.
División de las aurículas.
Mientras que los canales aurículo-ventriculares se forman, una serie de cambios estructurales dividen la aurícula común en dos cámaras independientes, derecha e izquierda. La división comienza en la quinta semana con el crecimiento descendente de un septum primum interauricular en forma de medialuna desde la pared craneal de la aurícula común (figura 16-18). Las puntas de la medialuna del septum primum se dirigen hacia el canal auriculo-ventricular y se fusionan con los cojinetes endocárdicos. El espacio que queda entre el borde libre del septum primum y los cojinetes endocárdicos se llama foramen (orificio) primum interauricular. Este orificio sirve como un atajo que permite el paso directo de la sangre desde la aurícula derecha a la izquierda.
Los cortocircuitos circulatorios presentes en el corazón en desarrollo satisfacen una necesidad muy práctica. Toda la sangre que llega entra en el lado derecho del septum primum interauricular. No obstante, a causa del desarrollo más tardío de los pulmones y de la deficiente capacidad de transporte los vasos pulmonares durante la mayor parte del periodo fetal. la circulación pulmonar no puede tolerar toda la carga sanguínea Si el corazón tuviese que formar cuatro cámaras separadas por completo desde el comienzo, la circulación pulmonar estaría sujeta a un esfuerzo excesivo y el lado izquierdo del corazón no estaría bombeando sangre suficiente para favorecer el desarrollo normal de otras estructuras, en especial durante las primeras semanas.
El problema de mantener una carga circulatoria equilibrada en todas las cámaras del corazón se satisface mediante la existencia de dos cortocircuitos que permiten que la mayoría de la sangre circulante no tenga que pasar por los pulmones. Un cortocircuito es la conexión directa entre las dos aurículas que permite que la sangre entre en la aurícula derecha para eludir por completo la circulación pulmonar pasando de forma directa a la aurícula izquierda. Este cortocircuito permite el desarrollo funcional normal de la aurícula izquierda. Sin embargo, si toda la sangre que entra en la aurícula derecha pasase directamente a la izquierda. el ventrículo derecho no tendría resistencia contra la cual bombear y se volvería hipoplásico. con la modificación de orificios de los canales vasculares en la aurícula derecha, una importante cantidad de sangre también entra en el ventrículo derecho y sale de esa cámara a través del tracto de salida de la arteria pulmonar. La mayor parte de la sangre que sale del ventrículo derecho, que es demasiada para caber en la circulación pulmonar evita a los pulmones a través del dactus arterioso y pasa directamente en la aorta descendente. Mediante estos dos mecanismos, el corazón funciona en su totalidad y se protege la circulación pulmonar Cuando el septum primum interauricular está a punto de fusionarse con los cojinetes endocárdicos, un área de muerte celular genéticamente programada ocasiona la aparición de múltiples perforaciones cerca de su extremo craneal (figura I6-18). Conforme el borde libre del septum primum se fusiona con los cojinetes cndocárdicos, obliterando el foramen primum. las perforaciones craneales del septum primum constituyen y dan lugar al foramen (orificio) interauricular secundum. Este nuevo orificio preserva la conexión directa entre las dos aurículas.
Poco después de la aparición del foramen secundum, comienza a formarse un tabique a la derecha del septum primum. el septum secundum que tiene forma de medialuna. Esta estructura, que crece desde la parte dorsal hacia la ventral de la aurícula, delimita el foramen oval. La posición del foramen oval permite que casi toda la sangre que entra en la aurícula derecha proveniente de la vena cava inferior pase directamente a la aurícula derecha través de él y del foramen secundum. Sin embargo, la disposición de los dos tabiques interauriculares les permite actuar como una válvula de una sola vía, es decir, que la sangre puede cruzar de la aurícula derecha hacia la izquierda pero no en el sentido contrario
Durante la etapa del corazón tubular recto el seno venoso es una cámara simétrica bilateral en la cual desembocan las principales venas del cuerpo(figura 16-11)Conforme se desarrolla la incurvación del corazón y se forman los tabiques interauriculares. La entrada del seno venoso en el corazón se desplaza por completo hacia la aurícula derecha (figura 16-15 y 16-18). Cuando esto sucede, el cuerno derecho del seno venoso se incorpora cada vez mas a la pared de la aurícula derecha. De forma que el cuerno derecho o seno coronario(que es el canal de drenaje común para las venas coronarias) ahora muy reducido, desemboca directamente en la aurícula derecha (figura 16-12). También en la aurícula derecha se forman unos repliegues de tejido similares a unas válvulas (válvulas venosas) a la entrada de las venas cavas superior e inferior. A causa de la orientación del orificio y de su posesión. la sangre que entra en la aurícula derecha por la vena cava inferior pasa casi toda a través del cortocircuito auricular hacia la aurícula derecha. En tanto que la sangre que viene de la vena cava superior y el seno coronario fluye a través de la válvula tricúspide hacia el ventrículo derecho.
Cuando los tabiques interauriculares están comenzando a formarse. comienza a crecer un tabique interventricular desde el vértice del ventrículo común hacia los cojinetes endocárdicos auriculo-ventriculares. La división inicial del ventrículo común también se ve reflejada en la superficie externa del corazón por la presencia de un surco (figura 16-19). Aunque al comienzo existe un foramen interventricular, al final se oblitera. Esto se logra mediante: 1) el crecimiento del tabique interventricular muscular. 2) una contribución del tejido de la cresta troncoconal que divide el tracto de salida del corazón, y 3) un componente membranoso derivado del tejido conectivo del cojinete endocárdico.
Desarrollo y absorción del seno venoso y de la aurícula derecha
División del tracto de salida del corazón
En el corazón tubular inicial el tracto de salida es un tubo sencillo, el bulbo cardiaco. Cuando comienza a formarse el tabique interventricular, el bulbo se alarga v puede dividirse en un cono arterioso proximal y un tronco arterioso distal (figura 16-14). Si bien al comienzo es un canal único, el tracto de salida se divide en canales aórticos y pulmonares separados gracias a la aparición de dos crestas troncoconales espirales que derivan en su mayor parte del mesénquima de la cresta neural. Estas crestas hacen relieve hacia la luz y acaban por fusionarse, dividiéndola en dos canales. El saco aórtico. que está localizado en situación distal a la región troncoconal. no contiene crestas. La partición del tracto de salida comienza cerca de la raíz aórtica ventral entre el cuarto y el sexto arcos y se extiende hacia los ventrículos haciendo un giro en espiral conforme avanza (figura 16-20). Esto explica la espiral parcial entre la aorta y la arteria pulmonar en corazón adulto.
Antes y durante el proceso de partición, las células derivadas de la cresta neural de la pared del tracto de salida comienzan a producir fibras elásticas que aportan la elasticidad requerida por la aorta y otros grandes vasos. La elastogénesis sigue un gradiente, primero a través del tracto de salida, luego en la aorta misma y por último en las ramas arteriales más pequeñas que se originan de la aorta.
En la base del cono, donde se forma el tejido de los rodetes endocárdicos de la misma forma que sucede en el canal aurículo-ventricular, se forman dos nuevos juegos de válvulas semilunares (figura 16-21). Estas válvulas, cada una de las cuales tiene tres valvas, impiden que la sangre inyectada se devuelva hacia los ventrículos. Se dice que hay células de la cresta neural craneal y del mesodermo cardiaco que contribuyen a la formación de las válvulas semilunares, aunque no todos los investigadores están de acuerdo en la contribución de la cresta neural. Como ya se dijo, las extensiones más proximales de las crestas troncoconales contribuyen a la formación del tabique interventricular. Inmediatamente después en el lado aórtico de la válvula semilunar aórtica, las dos arterias coronarias se unen a la aorta para irrigar el corazón.
Inervación del corazón
Aunque el desarrollo inicial del corazón tiene lugar con independencia de los nervios, tres tipos de fibras nerviosas inervan el corazón (figura 16-22). Las fibras nerviosas simpáticas (adrenérgicas), cuya acción es acelerar el latido cardiaco. llegan como proyecciones de los ganglios simpáticos del tronco. Estas fibras nerviosas se derivan de la cresta neural del tronco. La inervación parasimpática (colinérgica) deriva del componente cardiaco de la cresta neural craneal. Las neuronas de los ganglios cardiacos, que son las neuronas parasimpáticas de segundo orden, migran directamente hacia el corazón desde la cresta neural. Estas hacen sinapsis con los axones de las neuronas parasimpáticas de primer orden que llegan al corazón a través del nervio vago. La innervación sensitiva del corazón también llega a través del nervio vago. pero las neuronas sensitivas se originan en el ectodermo de las plácodas (plácoda nodosa) (figura 5-14). Así. la inervación del corazón tiene tres orígenes separados.
Si se extirpa la cresta neural cardiaca de un embrión joven de pollo, siguen formándose los ganglios colinérgicos cardiacos. Las investigaciones han determinado que las placodas nodosas compensan la pérdida de la cresta neural mediante el aporte de neuronas que toman el lugar de las parasimpáticas normales.
Sistema de conducción del corazón
El latido cardiaco normal es el resultado de un sistema complejo de marcapasos internos y de un sistema de conducción. que distribuye rápidamente el estimulo contráctil a todo el corazón. También participa la inervación del sistema nervioso autónomo. que modula el latido cardiaco a una frecuencia más alta (nervios simpáticos) o más baja (nervios parasimpáticos).
En el desarrollo muy precoz del corazón la localización del marcapasos se desplaza desde el extremo mas caudal del tubo izquierdo del corazón no fusionado hasta el seno venoso. Conforme el seno venoso se incorpora a la aurícula derecha, el marcapasos. que ahora se llama nodo sinoauricular, se sitúa en la parte alta de la aurícula derecha, cerca de la entrada de la vena cava superior. Un poco más tarde, se forma el nodo aurículo-ventricular en el área del tabique interauricular situado precisamente por encima del tejido del cojinete endocárdico. Estos dos nodos están conectados por varias vías de células musculares cardiacas muy modificadas que conducen el estímulo contráctil desde el nodo sinoauricular hasta el auriculo-ventricular. Desde este último. un haz auriculo-ventricular bien definido pasa desde la aurícula hasta el ventrículo y se divide en dos ramas, derecha e izquierda. Estas ramas a continuación distribuyen el tejido de conducción (fibras de Purkinje) por todo el miocardio ventricular.
Aun se desconocen muchos aspectos de la embriología del sistema de conducción. Los haces de conducción maduros están compuestos por miocitos cardiacos muy modificados que contienen cantidades abundantes de glucógeno. Hay pruebas de que las células del haz aurículo-ventricular se diferencian a partir de células precursoras independientes. posiblemente de los anillos de tejido que al final formaran el esqueleto cardíaco, y que las células de Purkinje se derivan de un linaje separado que se diferencia a partir las células que dan lugar a los miocitos ventriculares Estas mismas células expresan el factor de trascripción Msx-2 durante la mayor parte de su desarrollo. Aunque el sistema de conducción tiene una rica inervación, se forma antes de que los nervios entren en el corazón. Por ello. puede descartarse la inervación como factor causal de su desarrollo.
COMIENZO DE LA FUNCIÓN CARDÍACA
Debido a su fácil acceso, el embrión de pollo ha enseñado a los científicos casi todo lo que se conoce sobre las primeras funciones. del corazón embrionario. En años recientes la aplicación de técnicas de ultrasonido ha permitido investigar algunos aspectos de la función cardiaca en los embriones humanos a las 4 ó 5 semanas de gestación.
Las regiones bilaterales del mesodermo precardiaco en el embrión de pollo se diferencian en los tubos cardiacos derecho e izquierdo con regiones funcionales ventricular. auricular y sinoauricular. La contracción del corazón embrionario; comienza cuando los tubos cardiacos derecho e izquierdo comienzan a fusionarse, y la circulación sanguínea comienza poco después. En el embrión humano esto sucede entre los 21 y los 23 días de la gestación.
Las distintas partes del tubo cardiaco tienen diferentes latidos intrínsecos, y los experimentos de injerto han demostrado que las características funcionales de las regiones del corazón dependen de claves extracelulares. Por ejemplo, si se injerta tejido preauricular en un área preventricular, adquiere las características funcionales del tejido ventricular.
Conforme los tubos cardíacos del embrión de pollo comienzan a fusionarse a nivel de la región ventricular. los ventrículos fusionados tienen un latido intrínseco con una frecuencia de 25 latidos por minuto, y las aurículas no fusionadas no laten. Unas pocas horas mas tarde, cuando las partes auriculares de los tubos cardiacos se han fusionado, la región auricular late a una frecuencia de 62 latidos por minuto. El latido auricular actúa como un marcapasos. y hace que el ventrículo lata a la misma frecuencia. No obstante, si se separa la aurícula del ventrículo, el latido del ventrículo desciende a una frecuencia intrínseca de sólo 24 por minuto. Por último, cuando se forma el seno venoso, su latido intrínseco de 140 por minuto marca el paso del latido de todo el corazón.
En el corazón en proceso de fusión se localiza en la región del seno venoso una zona marcapasos bien definida. El marcapasos esta compuesto por un cúmulo de 60 a 150 células aproximadamente que emiten una señal como si fuera una sola célula. El marcapasos comienza su onda de excitación que se propaga por todo el corazón en 0.5 segundos. Conforme el corazón madura y se agranda, la velocidad de la propagación aumenta en proporción al tamaño del corazón. Así pues, la velocidad de conducción del estimulo excitatorio se multiplica por cien. pero el tiempo real de conducción no cambia mucho a pesar del aumento de mil veces en la masa del corazón del embrión de pollo. Los números crecientes de uniones comunicantes (gap-junctions) entre los miocitos cardiacos en desarrollo pueden explicar en parte el aumento de la velocidad de conducción del corazón en desarrollo.
No se conoce el mecanismo por el cual el tubo cardiaco es capaz de bombear la sangre en ausencia de válvulas. Una vez que la aurícula se llena de sangre proveniente del seno venoso, se contrae y envía la sangre al ventrículo. Entonces un movimiento parecido al peristáltico desplaza la sangre hacia la región del tronco, desde la que sale del corazón y entra en el saco aórtico. La contracción activa de la región del tronco impide el reflujo de sangre desde la nariz aórtica hacia el corazón. Incluso en el corazón joven en los cojinetes endocárdicos del canal aurículo-ventricular y del tracto de salida tienen una función similar a las válvulas. En ocasiones se ha pensado que los patrones del flujo sanguíneo en el Corazón inicial son responsables de los patrones de la tabicación interna del corazón. No obstante, se ha demostrado que muchos aspectos de la morfogénesis cardiaca están separados de esta función. La presión sanguínea del embrión temprano es muy baja (0.61 0.3 mml hg en el embrión de pollo de 3 días checar dato). Conforme el embrión crece, la presión de la sangre aumenta de forma exponencial y luego el aumento se detiene poco antes del nacimiento
En muchos aspectos el plan general de la circulación embrionaria parece ser ineficiente y mas complejo de lo que se necesita para mantener el desarrollo y el crecimiento del feto. Sin embargo, el embrión tiene que prepararse para el momento en que se produce un cambio súbito hacia un patrón completamente diferente de oxigenación de la sangre por los pulmones en vez de serlo por la placenta, y esto hace que las modificaciones del patrón de la circulación fetal sean imprescindibles.
La sangre bien oxigenada proveniente de la placenta entra en la vena umbilical como un gran chorro que en ocasiones esta bajo una gran presión causada por las contracciones uterinas el hígado, la sangre de la vena umbilical que tiene una presión alta pasa directamente por el
conducto venoso lo que le permite eludir los pequeños capilares circulatorios del hígado y fluir directamente hacia la vena cava inferior (figura 16-23). Una vez en la vena cava, alcanza inmediatamente el corazón. La sangre poco oxigenada que fluye por la vena cava inferior puede regurgitar en cierto grado a causa de la fuerza del flujo sanguíneo umbilical.
Hay pruebas funcionales de la existencia de un esfínter fisiológico en el conducto venoso, que obliga a gran parte de la sangre umbilical a pasar por los canales capilares hepáticos y entrar en la vena cava inferior a través de las venas hepáticas cuando se han desarrollado.
Esto reduce de forma considerable la presión de la sangre umbilical y permite que la sangre sistémica poco oxigenada que discurre por la vena cava inferior entre en la aurícula derecha con una presión baja. La sangre que discurre a alta presión y que entra en la vena umbilical procedente de la placenta también tiende a impedir que la sangre de la vena porta hepática entre en el conducto venoso. Cuando el útero está relajado y la sangre venosa umbilical discurre a baja presión, la sangre portal poco oxigenada se mezcla con la sangre umbilical en el conducto venoso. En la vena cava inferior también se produce mezcla de la sangre sistémica con la umbilical.
En la aurícula derecha la orientación de la entrada de la vena cava inferior permite que un chorro de sangre a una presión ligeramente elevada pase directamente a través del foramen oval y de! foramen secundum hacia la aurícula derecha (figura 16-23). Ésta es la ruta que suele tomar la sangre umbilical bien oxigenada que entra en el organismo a alta presión Puesto que la comunicación interauricular en el feto es mas pequeña que el orificio de la vena cava inferior. Parte de la sangre bien oxigenada de la cava pasa de la aurícula derecha al ventrículo derecho Cuando la sangre a baja presión (o sea la sangre sistémica poco oxigenada) entra en la aurícula derecha, se mezcla con la sangre venosa que viene de la cabeza por la vena cava superior y del corazón por el seno coronario y su mayor parte se dirige por la válvula tricúspide hacia: el ventrículo derecho.
Toda la sangre que entra en el ventrículo derecho sale por la arteria pulmonar y pasa hacia los pulmones. Incluso hacia el final de la vida fetal, la vascularización pulmonar no esta preparada para recibir todo el volumen de sangre que entra en la arteria pulmonar. La sangre que no cabe en las arterias pulmonares se deriva hacia la aorta a través del ductus arterioso. Esta estructura protege a los pulmones de la sobrecarga Circulatoria. y sin embargo permite que el ventrículo derecho se desarrolle en preparación para su funcionamiento total en el momento del nacimiento. El control de la permeabilidad del ductus ha sido sujeto de gran controversia. La permeabilidad del ductus arterioso como del conjunto venoso en el feto se mantiene de forma activa a través de la acción de las prostaglandinas (prostaglandina E2 y prostaglandina I2 respectivamente) La aurícula izquierda recibe un chorro de sangre umbilical bien oxigenada a través de la comunicación interauricular pequeña cantidad de sangre poco oxigenada desde las venas pulmonares. Esta sangre, que en su conjunto esta relativamente bien oxigenada, pasa hacia el ventrículo izquierdo y sale del corazón a través de la aorta. Algunas de las Primeras ramas arteriales que se desprenden de la aorta irrigan el corazón y el cerebro, los dos órganos que requieren mayor concentración de oxigeno para su desarrollo normal.
aporta sangre poco oxigenada a su torrente. Esta mezcla A nivel donde el cayado de la aorta comienza a descender. El ductus Arterioso de sangre bien oxigenada y poco oxigenada se distribuye entonces a los tejidos y órganos irrigados por las ramas torácicas y abdomen
ales de la aorta. Cerca del extremo caudal de la aorta se desprenden las dos grandes arterias umbilicales. Que transportan la sangre a la placenta para su renovación (checar de donde es)
CASO CLÍNICO
Un niño de 8 años es traído al médico aquejado de fatiga excesiva y de molestias en las piernas al caminar o al correr. En el examen físico, el médico observa una reducción del pulso pedio dorsal y algunos signos de cianosis en los dedos de los pies. Las manos del niño no muestran signos de cianosis.
¿Con base en el examen físico el médico ha sospechado la presencia de qué malformación? ¿Por qué?
* Durante la hematopoyesis, los hemocitoblastos dan lugar a las células precursoras linfoides y mieloides. Cada una de estas células progenitoras se diferencia en las líneas definitivas de las células sanguíneas. La eritropoyesis comprende el desarrollo en varias etapas de células precursoras de los glóbulos rojos. Las más precoces se definen por características conductales y no morfológicas. Durante las últimas etapas de la diferenciación, las células precursoras de los eritrocitos pierden gradualmente su maquinaria de producción de RNA y acumulan cantidades crecientes de hemoglobina en su citoplasma; al mismo tiempo el núcleo se toma más condensado y termina por desaparecer. La hemoglobina también experimenta transformaciones isomórficas durante el desarrollo embrionario.
*La formación más precoz de la sangre y los vasos sanguíneos extraembrionarios asociados con ella tienen su origen en los islotes sanguíneos en la pared mesodérmica del saco vitelino. Gran parte de la vascularización del cuerpo del embrión tiene su origen en estructuras intraembrionarias. Los precursores de las células endoteliales (angioblastos) surgen de la mayoría de los tejidos mesodérmicos del cuerpo excepto del mesodermo de la notocorda y del precordal. Los vasos sanguíneos embrionarios se forman mediante tres mecanismos principales: 1) coalescencia in situ, 2) migración de angioblastos hacia los órganos y 3) brotes a partir de los vasos existentes. El crecimiento de vasos sanguíneos hacia el interior de los primordios de algunos órganos se produce bajo el estímulo de factores angiogénicos.
*Los primeros tres pares de arcos aórticos forman las arterias que irrigan la cabeza. El cuarto par de arcos se desarrolla de forma asimétrica, ya que el arco izquierdo forma parte del cayado de la aorta del adulto. El quinto par de arcos nunca se forma. El sexto par de arcos aparece en forma de un plexo capilar que une con el cuarto arco. La parte distal del sexto arco izquierdo da lugar al ductus arterioso, un cortocircuito que permite que la sangre pase de largo sin entrar en los pulmones inmaduros y llegue directamente a la aorta. Muchas de las grandes arterias del adulto se originan a partir de tres pares de ramas aórticas: la intersegmentaria dorsal, la segmentaria lateral y la segmentaria ventral. Las arterias coronarias surgen de plexos capilares asociados con el epicardio. Estos plexos se conectan en segunda instancia con la aorta.
*El sistema venoso tiene su origen en redes capilares muy complejas que al comienzo se conectan con componentes del sistema de las venas cardinales. Las venas cardinales anterior y posterior drenan la cabeza y el tronco, respectivamente. Luego, desembocan en pares de venas cardinales comunes y por último en el seno venoso del corazón. Las venas subcardinales pares se asocian con el mesonefros en desarrollo. Las venas umbilicales extraembrionarias pares y las vitelinas atraviesan el hígado en desarrollo y desembocan directamente en el seno venoso. Las venas pulmonares se originan como estructuras separadas que desembocan en la aurícula izquierda. El sistema linfático hace su aparición en forma de seis sacos linfáticos primarios. Éstos luego se conectan mediante conductos linfáticos. Los linfáticos de casi todo el organismo confluyen en el conducto torácico, que desemboca en el sistema venoso a nivel de la base de la vena yugular interna izquierda.
• El corazón surge del mesodermo esplácnico a partir de un primordio en forma de herradura. En su origen, los tubos endocárdicos bilaterales se fusionan en la línea media. El tubo cardíaco fusionado forma a continuación un asa en forma de S y pronto pueden identificarse las regiones especificas del corazón. Comenzando por el tracto de entrada, estas regiones son el seno venoso, las aurículas, los ventrículos y el tracto de salida (bulbo del corazón). El tracto de salida más tarde se divide en el cono arterioso y el tronco arterioso.
• Los cojinetes endocárdicos auriculares son engrosamientos situados entre las aurículas y los ventrículos. El miocardio subyacente induce a las células del revestimiento endocárdico del cojinete a abandonar la capa endocárdica y a transformarse en células mesenquimatosas que invaden la gelatina cardiaca. Estos sucesos constituyen la base para la formación de las válvulas aurículo-ventriculares.
• La división interna del corazón comienza con la separación de las aurículas de los ventrículos y la formación de las válvulas tricúspide y mitral. Las aurículas derecha e izquierda se separan mediante el crecimiento del septum primum y del septum secundum, pero durante toda la vida embrionaria persiste un cortocircuito que va desde la aurícula derecha hacia la aurícula izquierda a través del foramen secundum y del foramen oval. El seno venoso y las venas cavas desembocan en la aurícula derecha, y las venas pulmonares lo hacen en la aurícula izquierda. Los ventrículos están separados por el tabique interventricular. Las crestas troncoconales de disposición espiral dividen el tracto de salida común en los troncos aórtico y pulmonar. Las válvulas semilunares impiden el reflujo de la sangre desde estos vasos hacia el corazón.
• Además de la inervación sensitiva, el corazón recibe inervación simpática y parasimpática. El sistema de conducción distribuye el estimulo contráctil a través de todo el corazón. El sistema de conducción deriva de células musculares cardiacas modificadas, el corazón comienza a latir al comienzo de la cuarta semana de la gestación. La maduración fisiológica del latido cardíaco sigue a la maduración del sistema del marcapasos y a la inervación del corazón.
• La circulación fetal trae sangre oxigenada de la placenta a través de la vena umbilical y la lleva a la aurícula derecha, de donde la mayor parte pasa a la aurícula izquierda. Otra sangre que entra a la aurícula derecha pasa al ventrículo derecho. La sangre que sale del ventrículo derecho entra en el tronco pulmonar, que envía algo de sangre a los pulmones y la mayor parte a la aorta a través del ductus arterioso. La sangre que llega a la aurícula izquierda pasa al ventrículo izquierdo y de allí a la aorta, desde donde irriga todo el cuerpo. La sangre poco oxigenada entra en las arterias umbilicales y es transportada a la placenta para su renovación.
• Las malformaciones más frecuentes del corazón son los defectos del tabique interauricular, que en la vida posnatal permiten el paso de sangre de la aurícula izquierda a la derecha. Los defectos del tabique interventricular, que también dan como resultado el cortocircuito de izquierda a derecha, son más graves. Los defectos que bloquean un canal por donde fluye la sangre (por ejemplo, la atresia tricuspídea) deben acompañarse de cortocircuitos secundarios para que sean compatibles con la vida. Un canal aurículo-ventricular persistente puede atribuirse a un defecto en la formación o en el posterior desarrollo de los cojinetes endocárdicos aurículo-ventriculares. La mayoría de las malformaciones del tracto de salida del corazón parecen estar relacionadas con la división inapropiada realizada por las crestas troncoconales. La base de estas anomalías se encuentra a menudo en defectos de la cresta neural.
• Las malformaciones de las arterias principales suelen provenir de la aparición o la inapropiada desaparición de componentes específicos del sistema de los arcos aórticos. Algunas, como el cayado aórtico doble o el cayado aórtico derecho, pueden entorpecer la deglución o la respiración a causa de la compresión que ocasionan. El ductus arterioso persistente tiene su origen en la falta de cierre apropiado del ductus después del nacimiento. La coartación de la aorta debe ser compensada ya sea con un ductus arterioso persistente o con la apertura de conductos vasculares colaterales que permitan a la sangre superar el lugar de la coartación.
• A causa de su complejo forma de originarse, las venas suelen presentar muchas variaciones, pero estas malformaciones a menudo son asintomáticas. El retomo pulmonar anómalo que trae sangre oxigenada a la aurícula derecha, ha de acompañarse de un cortocircuito de derecha a izquierda para ser compatible con la vida. Las malformaciones del sistema linfático pueden causar tumoraciones locales como el higroma quístico, que da como resultado la aparición de una tumefacción en forma de collar en el cuello.
CORRELACIÓN CLÍNICA 16_1a
Con una incidencia de casi uno por cada cien nacidos vivos, las malformaciones cardíacas representan el tipo más frecuente de anomalías congénitas. A causa del estrecho equilibrio fisiológico de la circulación, la mayoría de las malformaciones son sintomáticas. Desde un punto de vista clínico. las malformaciones cardíacas suelen clasificarse como las que se asocian con cianosis (defectos cianosantes) en la vida posnatal y las que no lo hacen (defectos no cianosantes).
La cianosis se produce cuando la sangre contiene más de 5 mg/dl de hemoglobina no saturada. La cianosis se reconoce fácilmente por el color azulado o purpúreo de la piel en áreas de densa circulación capilar superficial. Se asocia con policitemia, un aumento en la en la sangre debido la disminución global de la saturación de oxígeno. A largo plazo la cianosis se concentración de eritrocitos asocia con una deformidad de dedos en palillos de tambor y retraso del crecimiento. En los casos más graves de cianosis, el niño suele adoptar una posición en cuclillas para facilitar la reoxigenación de la sangre.
Después del nacimiento, la cianosis se asocia con la presencia de un cortocircuito de izquierda a derecha en la cual la sangre venosa se mezcla con la sangre sistémica. Algunos defectos cardiacos son no cianosantes durante muchos años pero después se toman cianosantes. Estos defectos se caracterizan al comienzo por un cortocircuito de izquierda a derecha en el cual la sangre sistémica oxigenada refluye hacia la aurícula o el ventrículo derechos. El resultado neto es un aumento de la carga de bombeo del ventrículo derecho, que a la postre lleva a una hipertrofia ventricular derecha. Después de un período prolongado, el aumento de flujo a través de los pulmones provoca una reacción hipertensiva en la vasculatura pulmonar, que aumenta de forma efectiva la presión en el ventrículo y la aurícula derechos. Cuando la presión sanguínea en el lado derecho del corazón excede a la de la cámara izquierda correspondiente, el cortocircuito se revierte y la sangre poco oxigenada pasa a la circulación sistémica, con lo que se produce la cianosis. A partir de este momento el estado del paciente aquejado por la lesión cardiaca suele deteriorarse con mucha rapidez,
Malformaciones cardiacas 16-1b (continuación: 2)
CORTOCIRCUITOS DE CÁMARA A CÁMARA
Defectos del tabique interauricular
Varios tipos de defectos anatómicos en el tabique interauricular pueden dar como resultado un cortocircuito persistente entre las dos aurículas. Las variedades más comunes son causadas por la excesiva reabsorción del tejido que rodea al foramen secundum o por un crecimiento hipoplásico del septum secundum (figura 16-24 A). Una variedad menos frecuente es un defecto septal bajo, cuya causa suele ser la falta de unión entre el borde libre del septum prímum y los cojinetes endocárdicos (figura 16-24 B). Si el defecto es el resultado de una deficiencia del tejido de los cojinetes endocárdicos, los defectos asociados de las válvulas aurículo-ventriculares pueden complicar la lesión. La falta de tabícación de la aurícula da como resultado una aurícula común, un grave defecto que suele asociarse con otros defectos cardiacos. Los defectos del tabique interauricular se encuentran entre las malformaciones cardiacas más frecuentes. Estos defectos se están asociando cada vez más con el cromosoma 21. Los individuos con síndrome de Down (trisomía del cromosoma 21) tienen una alta incidencia de defectos de ambos tabiques. auricular y ventricular.
Los individuos que tienen mutaciones autosómicas dominantes del gen Nkx2-5 (Pág. 416} tienen una alta incidencia de anomalías del septum secundum que dan como resultado comunicaciones interauriculares. Con los defectos del tabique auricular se asocia también una alta incidencia de bloqueo auriculo-ventricular. que puede ocasionar la muerte súbita a los Individuos afectados cuyo corazón no se ayude con un marcapasos. Antes del descubrimiento de esta mutación se sospechaba que muchos de los casos de bloqueo aurículo ventricular se debían a la interrupción del haz aurículo-ventricular causada por el procedimiento de reparación. Durante los primeros días de la cirugía cardiaca, antes de que se determinara con precisión la anatomía del haz aurículo-ventricular, el bloqueo de rama inducido por la cirugía era un problema frecuente en la reparación de los defectos bajos del tabique auricular.
Los defectos del tabique auricular no complicados suelen ser compatibles con varios años de vida asintomática. Incluso durante el período libre de síntomas, la sangre de la aurícula izquierda, que se encuentra a una presión un poco más alta que la de la aurícula derecha, pasa hacia ésta última. Esta sangre adicional causa la hipertrofia de la aurícula derecha y el resultado es un aumento del flujo de sangre hacia los pulmones. En un período de muchos años. puede desarrollarse hipertensión pulmonar. Esto aumenta la presión sanguínea del ventrículo derecho y a la postre la de la aurícula derecha. Con sólo unos pocos milímetros de mayor presión de la aurícula derecha se invierte el flujo sanguíneo en el cortocircuito interauricular y ocurre la cianosis.
Una alteración más grave es el cierre prematuro del foramen oval. En esta situación todo el aporte de sangre que entra en la aurícula derecha pasa al ventrículo derecho, lo que causa una gran hipertrofia de todo el lado derecho del corazón.
El lado izquierdo queda muy hipoplásico debido a la poca sangre que bombean las cámaras izquierdas. Aunque este defecto suele ser compatible con la vida intrauterina, los bebés casi siempre fallecen poco después de nacer porque el corazón izquierdo hipoplásico no puede manejar la carga circulatoria normal.
La base usual para el canal aurículo-ventricular persistente es el subdesarrollo de los cojinetes endocárdicos que da como resultado una falta de división del canal aurículo-ventricular inicial en dos canales, derecho e izquierdo. Pueden existir varias causas específicas para los defectos del canal auriculo-ventricular, las cuales van desde la expresión inapropiada del Msx-1 hasta defectos en la producción o en la recepción de componentes de la matriz extracelular con actividad inductiva (por ejemplo, subunidades de los adherones) por parte de las células endocárdicas (figura 16-16).
Un canal auriculo-ventricular persistente suele asociarse con graves defectos de los tabiques auricular e interventricular (figura 16-25). Este grave defecto lleva a una alteración del crecimiento y a una expectativa de vida considerablemente reducida. Resulta interesante anotar que, pese a la posibilidad para la mezcla de las sangres venosa y arterial, la dirección predominante del cortocircuito es de izquierda a derecha, y algunos pacientes tienen poca cianosis.
Atresia tricuspídea
En la atresia triscuspídea, cuya etiología aún no se conoce bien, el orificio valvular normal entre la aurícula y el ventrículo derechos está completamente ocluido (figura 16-26, B). Un defecto como éste por sí solo causa la muerte puesto que la sangre no puede llegar a los pulmones para oxigenarse. Sin embargo, los niños pueden sobrevivir con esta malformación, y esto demuestra un punto importante en la embriología cardiaca. A menudo una lesión primaria se acompaña de una o más lesiones secundarias (por lo general comunicaciones) que permiten la supervivencia, aunque casi siempre con un nivel funcional bastante deficiente.
En esta situación, los cortocircuitos secundarios deben lograr dos objetivos. Primero, que persista una comunicación interauricular que debe derivar la sangre que no pasa por la válvula tricúspide atrésica hacia la aurícula izquierda. La sangre de la aurícula izquierda entonces fluye hacia el ventrículo izquierdo. Segundo, uno o más cortocircuitos secundarios deben permitir que la sangre gane acceso a los pulmones para que pueda oxigenarse. La sangre del ventrículo izquierdo puede pasar al ventrículo derecho y al sistema de las arterias pulmonares si existe un defecto en el tabique interventricular. Otra posibilidad es que la sangre del ventrículo izquierdo pase hacia la circulación sistémica donde puede obtener acceso a los pulmones si pasa hacia las arterias pulmonares desde la aorta a través del ductus arterioso persistente. Desde los pulmones la sangre oxigenada regresa a la aurícula izquierda, quizás para ser reciclada a través de los pulmones de nuevo antes de entrar en la circulación sistémica.
También puede presentarse atresia mitral, pero ésta es mucho menos frecuente que la atresia tricuspídea. También en ésta tienen que estar presentes algunos defectos secundarios compensatorios para permitir la supervivencia. Los bebés afectados por estas lesiones suelen sobrevivir tan sólo unos pocos meses o años.
Los defectos del tabique interventricular, que no son tan frecuentes como los defectos del tabique interauricular, suelen afectar a la porción membranosa del tabique donde convergen varios tejidos embrionarios (figura 16-27). Puesto que la presión de la sangre en el ventrículo izquierdo es más alta que la del derecho, esta lesión se asocia inicialmente con un cortocircuito no cianosante de izquierda a derecha del flujo de la sangré (figura 16-28). No obstante, el aumento del flujo sanguíneo hacia el ventrículo derecho causa la hipertrofia de éste último y puede llevar a la hipertensión pulmonar, que en último término causa la inversión del cortocircuito. La fisiopatología es similar a la de los defectos auriculares.
MALFORMACIONES DEL TRACTO DE SALIDA
El tracto de salida del corazón (región troncoconal) está sujeto a diversas malformaciones. Los estudios experimentales han demostrado una gran contribución de las células de la cresta neural a esta región. Los experimentos de extirpación e injerto han demostrado la aportación específica de células de la cresta neural cardiaca en el desarrollo normal del tracto de salida del corazón (figura 16-29). Si se extirpa la cresta neural cardiaca, las células ectodérmicas de la plácoda nodosa colonizan el tracto de salida, pero no se produce la septación normal del mismo, lo que lleva a un tronco arterioso persistente. Si se injerta cresta neural del mesencéfalo o del tronco en lugar de la cresta neural cardiaca o si se injerta cresta neural heterotópica en posición lateral a la cresta neural cardiaca, el resultado siempre es el tronco arterioso persistente. El último experimento muestra que la cresta neural extraña bloquea la migración o la función normales de la cresta neural cardiaca.
Aunque no todas las malformaciones de esta área pueden atribuirse a un desarrollo defectuoso de la cresta neural. Hay evidencias circunstanciales que sugieren que este puede ser un factor importante, varios defectos del tracto de salida se asocian con traslocaciones o deleciones del cromosoma 22, y en la mayoría de estos casos esta implicada la cresta neural.
Las lesiones del tracto de salida pueden crearse de forma experimental al bloquearse la función de genes específicos, sobre los genes localizados en el cromosoma 22 y en los genes que afectan a las propiedades de las células de la cresta neural craneal. por ejemplo, se observan anomalías del tracto de salida en ratones que tienen deficiencia de neurotropina-3(NT-3) un miembro de la familia del factor de crecimiento neural.
Tronco arterioso persistente
Malformaciones cardíacas (continuación 3)
Transposición de los grandes vasos
En raras ocasiones las crestas truncoconales no tienen las disposición para la división del tracto de salida en dos canales. El resultado son los dos arcos circulatorios por completo independientes, de los cuales el ventrículo derecho se continua con la aorta y el ventrículo izquierdo se continua en la arteria pulmonar(figura 16-31 B). Si la afección no se corrigiese. el arco circulatorio izquierdo continuaría bombeando sangre altamente oxigenada a través del lado izquierdo del corazón y los pulmones, en tanto que el lado derecho del corazón bombearía sangre venosa por la aorta hacia los canales circulatorios sistémicos y de vuelta hacia la aurícula derecha. Esta lesión, que es la causa mas frecuente de cianosis en los recién nacidos, es compatible con la vida solo si se acompaña de continuaciones interauriculares e interventriculares y ductus arterioso persistente, incluso con estas compensaciones anatómicas, la calidad de la sangre que irriga al organismo es baja.
Estenosis pulmonar y aórtica
Si la tabicación del tracto de salida por parte de las crestas troncoconales es asimétrica, puede suceder que o bien la aorta o bien la arteria pulmonar sean anormalmente estrecha, lo que provoca una estenosis aórtica y pulmonar (figuras 16-32 y 16-33). La gravedad de los síntomas esta en relación con el grado de la estenosis. En los casos mas extremos, la estenosis es tan pronunciada que la luz del vaso esta prácticamente obliterada.
Esta alteración recibe el nombre de atresia aortica o pulmonar. En ratones mutantes que tienen bloqueado el gen para la conexina 43, encargado de codificar una proteína que es componente del canal de las uniones comunicantes (gap-junctions), se ha observado una lesión que recuerda a la estenosis pulmonar. Aún no se sabe por qué esta lesión genética afecta sobre todo a la salida pulmonar del corazón.
Una de las lesiones más conocidas de este tipo es la tetralogía de Fallot, que se caracteriza por: 1) estenosis pulmonar, 2) un defecto del tabique interventricular membranoso, 3) una aorta grande (aorta a caballo), cuyo orificio se extiende hacia el ventrículo derecho, y 4) hipertrofia ventricular derecha. El defecto básico en la tetralogía de Fallot es una fusión asimétrica de las crestas troncoconales y una alineación defectuosa de las válvulas aórtica y pulmonar. A causa de la estenosis pulmonar y del orificio aórtico más amplio de lo normal, algo de sangre ventricular derecha poco oxigenada sale a través de la aorta aumentada de tamaño, lo cual provoca cianosis. La tetralogía de Fallot es la lesión cardiaca cianosante más frecuente.
CORRELACIÓN CLÍNICA 16-2
Malformaciones de los vasos sanguíneos
Debido a su modo de formación, en la que se favorece a un conducto vascular dentro de una densa red, los vasos sanguíneos (en especial las venas) están sujetos a numerosas variaciones con respecto de la normalidad. La mayor parte de las variantes anatómicas observadas en las salas de disección tienen poca importancia clínica. Los experimentos en animales sugieren que en la génesis de ciertas malformaciones de las grandes arterias pueden intervenir alteraciones de la cresta neural. Cuando se extirpa la cresta neural cardiaca en jóvenes embriones de ave, se presentan malformaciones que afectan a las arterias carótidas y al cayado de la aorta. Ciertas anomalías de los grandes vasos pueden ocasionar graves síntomas o tener importancia
durante la realización de una cirugía.
Otra malformación relativamente común no mortal del sistema vascular es la coartación de la aorta, que tiene dos variantes principales. Una consiste en un estrechamiento brusco de la aorta descendente después de la entrada del ductus arterioso (figura 16-38 B). La otra variante, llamada coartación preductal de la aorta, se produce antes del ductus (figura 16-38 A). La primera variedad (coartación posductal) es con mucho la más frecuente, ya que a ella corresponde el 95% de los casos. La embriogénesis de la coartación aún no está aclarada. Varias causas pueden llevar a la misma situación. En los pacientes con síndrome de Down o con síndrome de Turner, la incidencia de la coartación de la aorta es más elevada.
Una anomalía vascular frecuente es la falta de cierre del ductus arterioso después del nacimiento (figura 16-37). Al menos la mitad de los bebés que padecen esta afección están asintomáticos, pero con el paso de los años el fuerte flujo de sangre desde la circulación sistémica a alta presión (aórtica) hacia la circulación pulmonar sobrecarga la vascularización pulmonar y produce una hipertensión pulmonar y a la postre insuficiencia cardiaca.
La interrupción del cayado aórtico es una malformación vascular relativamente poco común que suele dar como resultado una interrupción distal a la salida de la arteria subclavia izquierda (figura 16-36). Para que sea compatible con la vida, esta lesión debe acompañarse de un ductus arterioso persistente y permeable que permita el flujo de sangre hacia la parte caudal del cuerpo. Esta lesión se ha producido en ratones a los que les falta el factor de trascripción de winged-helix MFH-1(mesenchyne fork head-1).
Malformaciones de las venas cavas
Duplicación del cayado de la aorta
En raras ocasiones, el segmento del arco aórtico dorsal derecho localizado entre la salida de la arteria subclavia derecha y el punto en que se une al arco aórtico izquierdo persiste en vez de degenerar. El resultado es un anillo vascular completo que rodea la traquea y el esófago (figuras 16-34 a y 16-35) . un cayado aórtico doble puede causar disnea (dificultad para respirar) en los bebes mientras que los alimentos, incluso si la afección es asintomática al comienzo de la vida, el posterior crecimiento suele estrechar el diámetro del anillo en relación con el tamaño de la traquea y del esófago, lo que puede producir síntomas mas tardíamente.
Cayado aórtico derecho
Un cayado aórtico hacia la derecha se produce por la persistencia del todo el arco aórtico embrionario y la desaparición del arco izquierdo desde el segmento caudal hasta la salida de la arteria subclavia izquierda (figura 16-34 B). Esta malformación es en esencia una imagen en espejo del desarrollo normal del cayado de la aorta, y puede aparecer como anomalía aislada o como parte de un situs inversus completo del individuo. los síntomas suelen ser leves o inexistentes a menos que una arteria subclavia izquierda aberrante comprima el esófago o la traquea.
esta alteración puede causar disnea y disfagia
brazo derecho (figura 16-34 C). Como sucede con el doble cayado aórtico
aórtico izquierdo y pasa por detrás del esófago y la traquea para alcanzar el
normalmente desaparece), la arteria subclavia derecha nace del cayado
C) derecha y el segmento mas distal del arco aórtico derecho (que
y si persiste el segmento que queda entre la salida de la arteria subclavia
salida de la séptima arteria intersegmentaria torácica derecha( figura 16-5 B
Arteria subclavia derecha que nace del
Interrupción del cayado aórtico izquierd
del cayado de la aorta
DUCTUS ARTERIOSO PERMEABLE. COARTACIÓN DE LA AORTA
En la coartación preductal de la aorta, que puede estar relacionada con una expresión inadecuada del MFH-1, el ductus arterioso suele permanecer permeable después del nacimiento. La sangre que irriga al tronco y las extremidades alcanza la aorta descendente a través del ductus. Esto puede llevar a una cianosis diferencial, en la cual la cabeza, la región superior del tronco y los brazos tienen color normal, pero la región inferior del tronco y las piernas están cianóticas a causa del flujo de sangre venosa dentro de la aorta a través del ductus arterioso permeable.
La vascularización tiene que compensar la coartación posductal en una forma diferente, ya que la localización del estrechamiento en este caso separa efectivamente el flujo arterial de la cabeza y los brazos respecto del flujo del tronco y las piernas. El cuerpo responde abriendo conductos de circulación colateral y conexiones a través de arterias que por lo común son relativamente pequeñas y que llevan la sangre desde la parte superior hacia la inferior del cuerpo (figura 16-38 C).Estos conductos son las arterias torácicas internas, las arterias relacionadas con el omoplato y la arteria espinal anterior. El flujo sanguíneo anormalmente grande que discurre por estas arterias se dirigen hacia ramas segmentarías (por ejemplo, las arterias intercostales) y llega a la aorta descendente después de la coartación. El aumento del flujo sanguíneo en las arterias intercostales causa unas muescas en las costillas que pueden verse con facilidad en las imágenes radiográficas. Pese a estas adaptaciones circulatorias compensatorias, la presión arterial en los pacientes que tienen una coartación posductal es mucho mas alta en los brazos que en las piernas.
Retorno pulmonar anómalo
Por la forma como cada una de las primitivas venas pulmonares se unen y por la absorción posterior de la parte distal del sistema venoso pulmonar en la pared de la aurícula izquierda , pueden presentarse conexiones anómalas de las venas pulmonares con el corazón (figura 16-39). Una de las mas comunes es que una o mas ramas de la vena pulmonar entren en la aurícula derecha en vez de hacerlo en la izquierda. En otros casos (retorno pulmonar anómalo total), todas las venas pulmonares desembocan en la aurícula derecha o en la vena cava superior, Este caso tiene que acompañarse de un cortocircuito asociado (por ejemplo, comunicación interauricular) para que la sangre oxigenada llegue a la circulación sistémica.
Malformaciones del sistema linfático
Aunque las variaciones anatómicas menores de los conductos linfáticos son frecuentes, las anomalías sintomáticas no lo son. Estas suelen presentarse como tumefacciones causadas por la dilatación de los grandes vasos linfáticos. Con diferencia, la principal anomalía linfática que se ve en los fetos es el higroma quistico, que se manifiesta como una gran tumefacción, en ocasiones incluso semejante a un collar, en la región del cuello (figura 7-1 A). Aunque la base embriológica del hidroma es incierta, la causa mas probable es el excesivo desarrollo y el crecimiento local del tejido linfático que posiblemente se originen como yemas comprimidas de los sacos linfáticos yugulares.
PREGUNTAS DE REPASO
1. Los eritrocitos nucleados que se encuentran circulando en el embrión se originan en:
A. El saco vitelino
B. Los cúmulos paraórticos
C. El hígado
D. La médula ósea
E. Ninguno de los anteriores
2. En un feto de siete meses de edad, la sangre que drena el músculo temporal izquierdo entra en el corazón a través de;
A. La vena cardinal anterior izquierda
B. El seno coronario
C. La vena cardinal común izquierda
D. La vena cava superior
E. Ninguna de las anteriores
3. ¿Qué estructura de la zona de los cojinetes endocárdicos libera las partículas inductivas llamada adherones?
A. Endocardio
B. Gelatina cardiaca
C. Miocardio
D. Epicardio
E. Ninguna de las anteriores
4. ¿Cuál de las siguientes estructuras contribuye la cresta neural?
A. Tronco arterioso
B. Aorta ascendente
C. Tronco de la pulmonar
D. Todas las anteriores
5. ¿Cuál de las siguientes malformaciones cardiovasculares necesita de un ductus arterioso permeable para que el individuo pueda sobrevivir?
A. Defecto del tabique interauricular
B. Defecto del tabique interventricular
C. Doble cayado aórtico
D. Arteria subclavia derecha que nace del cayado aórtico
E. Ninguna de las anteriores
6. Cinco días después de nacer un bebe se torna cianótico durante una prolongada crisis de llanto. La causa mas probable de la cianosis es que ha entrado sangre venosa en la circulación sistémica a través de:
A. El tabique interauricular
B. El ductus arterioso
C. El conducto venoso
D. La vena umbilical
E. El tabique interventricular
7¿De que arco aórtico procede la arteria carótida interna?
A. 1
B. 2
C. 3
D. 4
E. 5
8.Un niño de doce años dice a su medico que durante los últimos meses ha venido notando alguna dificultad para deglutir cuando come carne. El medico realiza un examen físico y solicita una radiografía de vías digestivas altas. Después de examinar las placas, el medico envía al niño para que le practiquen unos exámenes vasculares.¿Cuál es el razonamiento que respalda esta decisión?
9.Un individuo que tiene atresia de la válvula mitral no puede sobrevivir después del nacimiento si no tiene otros defectos del sistema cardiovascular que compensen el defecto primario, que en este caso es un bloqueo completo entre la aurícula y el ventrículo izquierdo. Defina al menos un grupo de defectos asociados que puedan compensar fisiológicamente la alteración causada por la atresia mitral.
10.¿cuál es la base embriológica de la duplicación de la vena cava inferior en una zona caudal a los riñones?
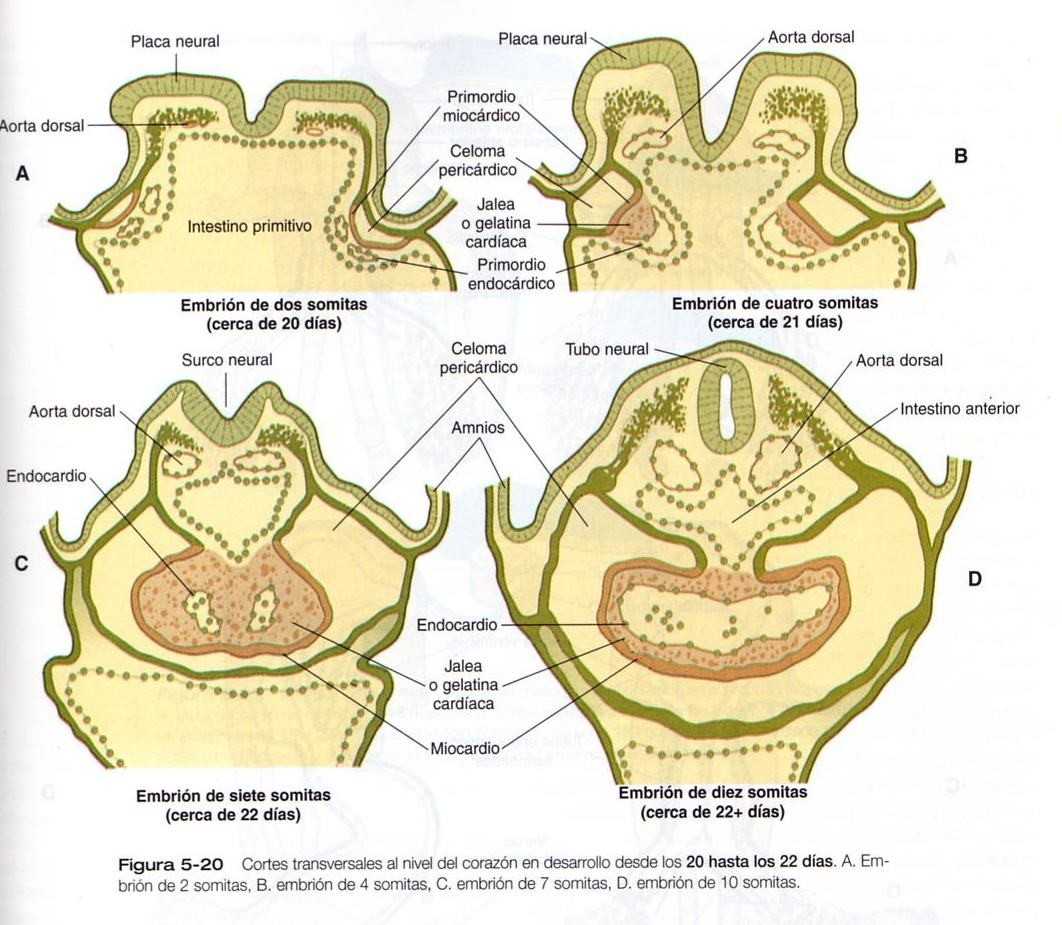{kind=link}
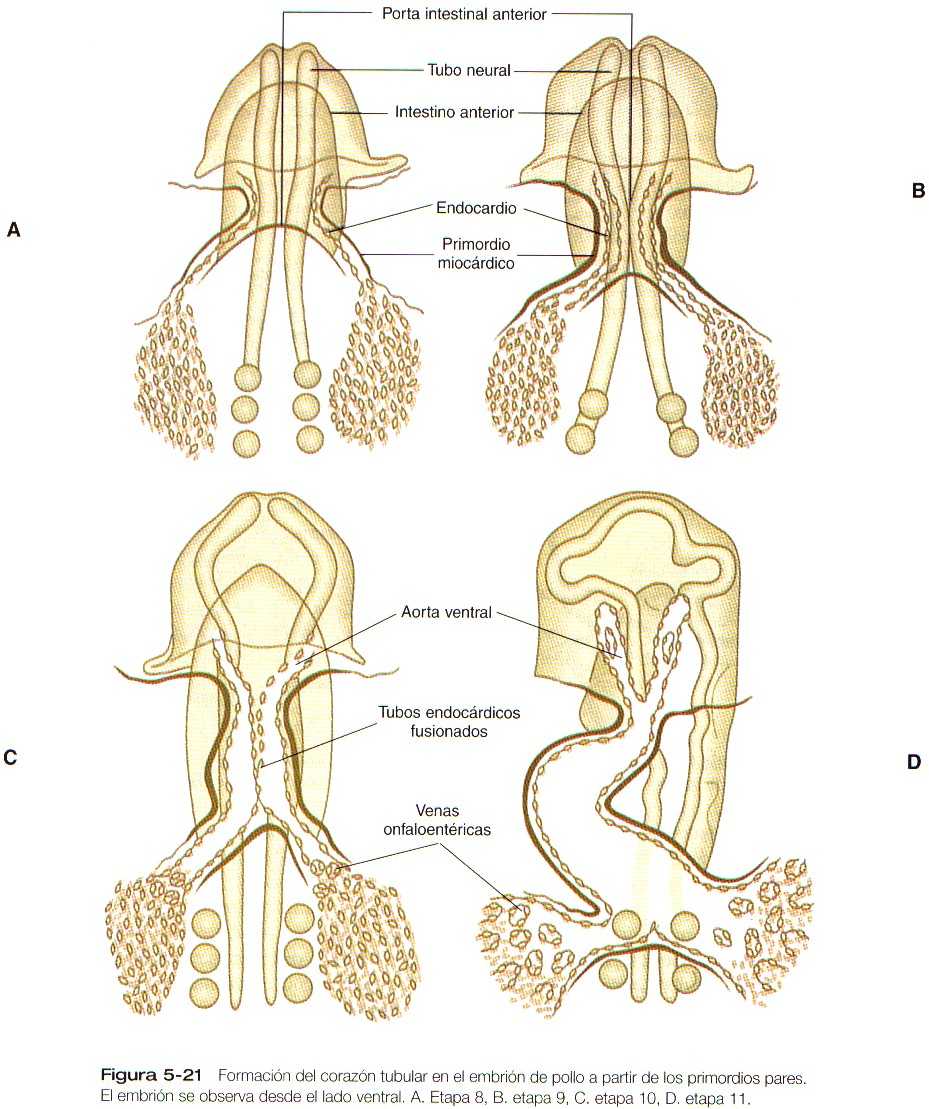{kind=link}
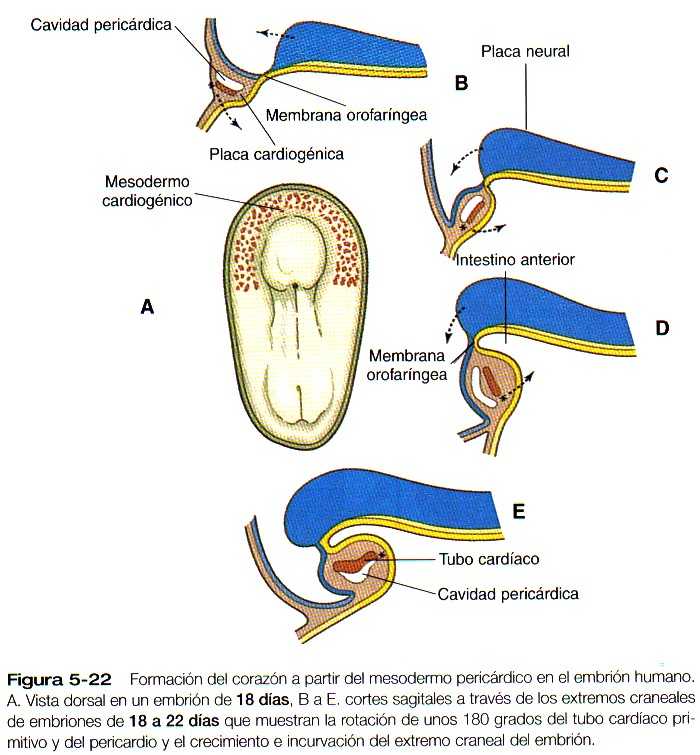{kind=link}
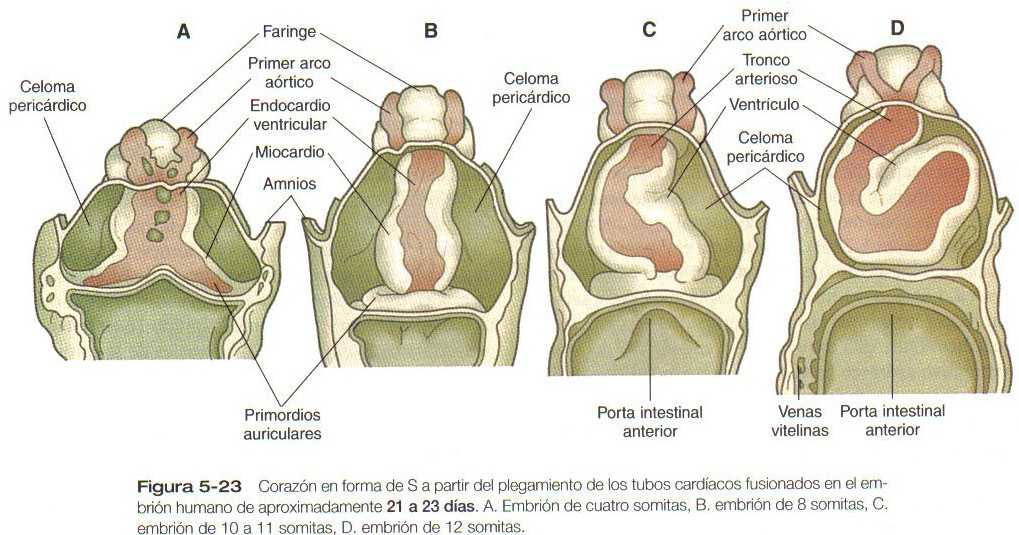{kind=link}
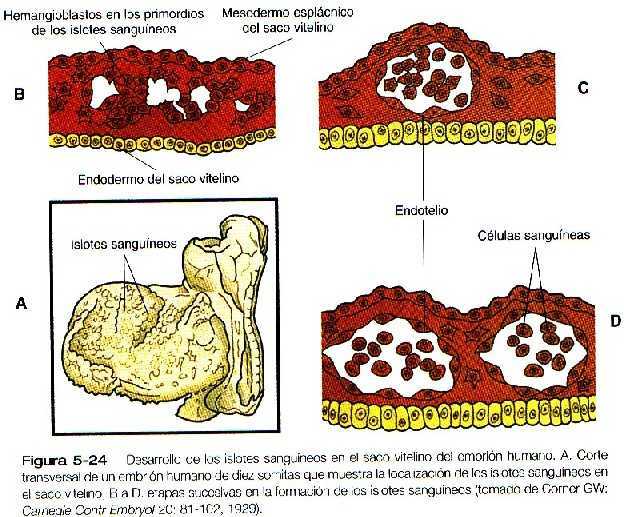{kind=link}
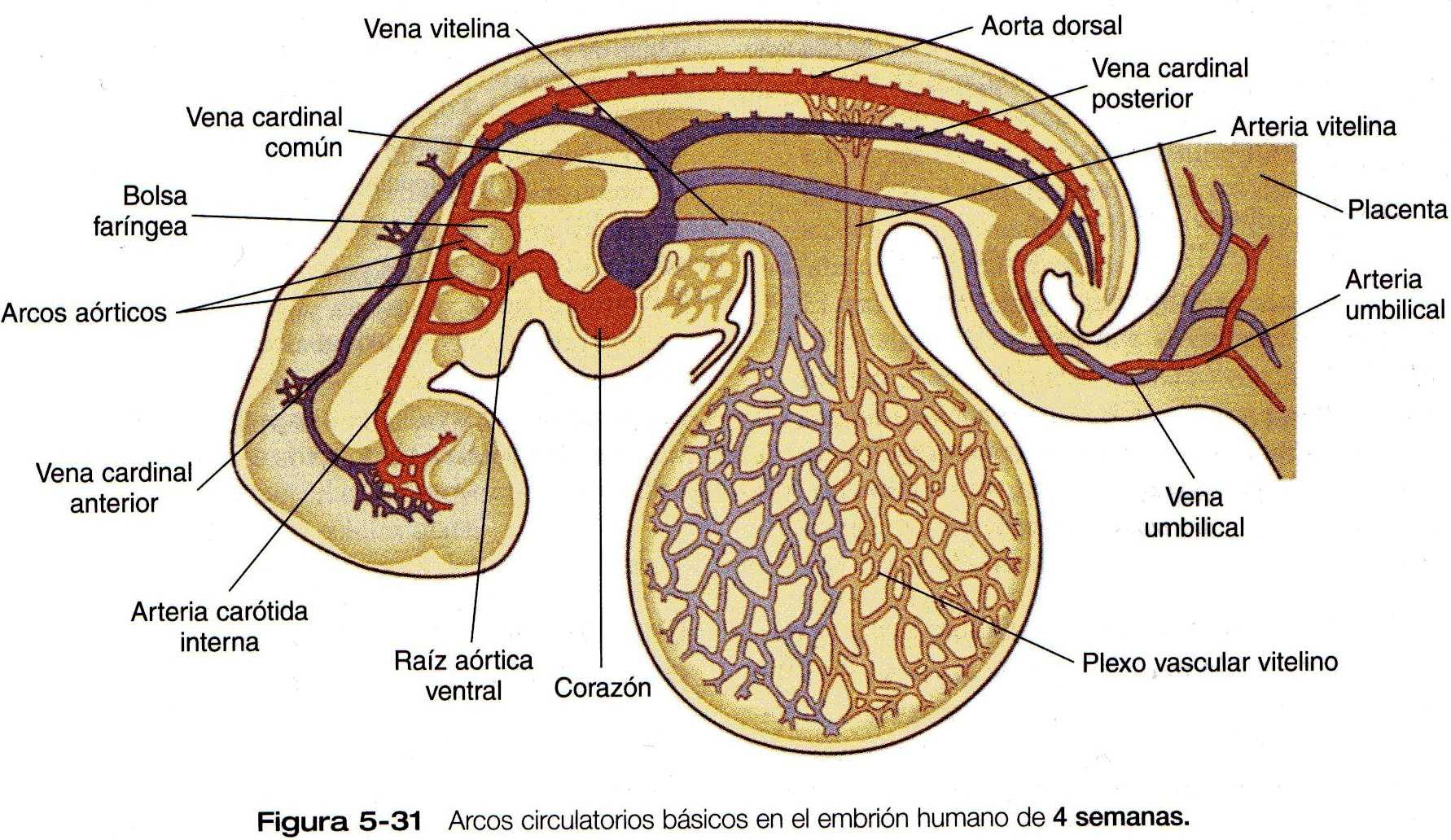{kind=link}
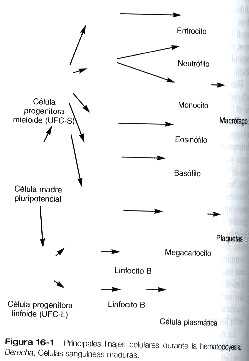{kind=link}
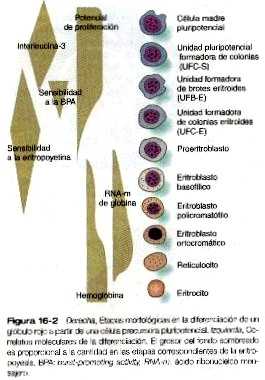{kind=link}
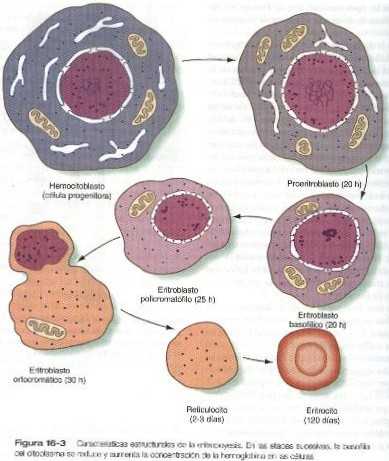{kind=link}
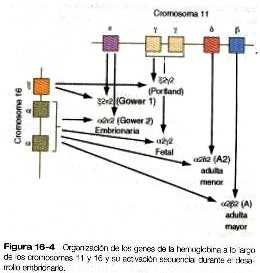{kind=link}
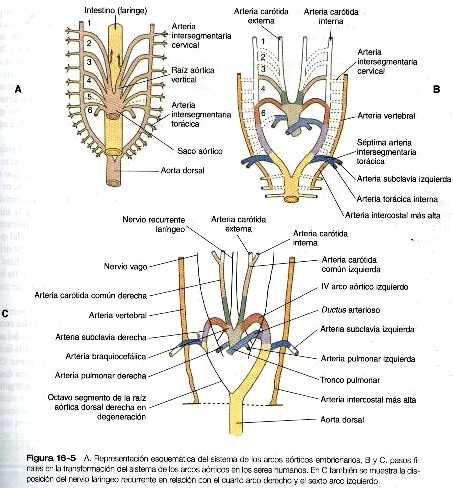{kind=link}
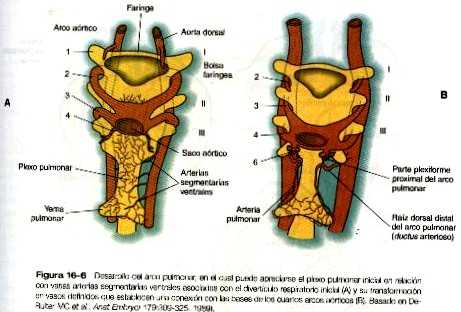{kind=link}
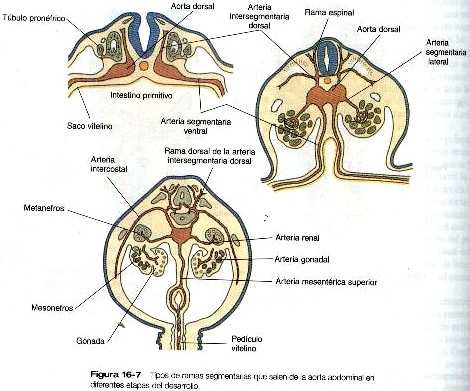{kind=link}
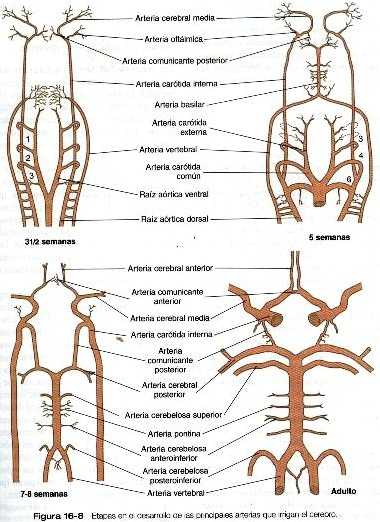{kind=link}
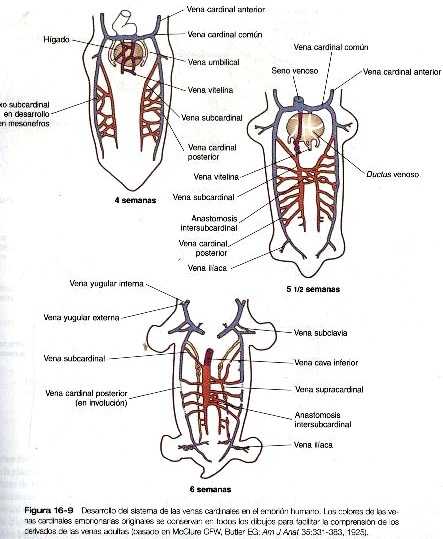{kind=link}
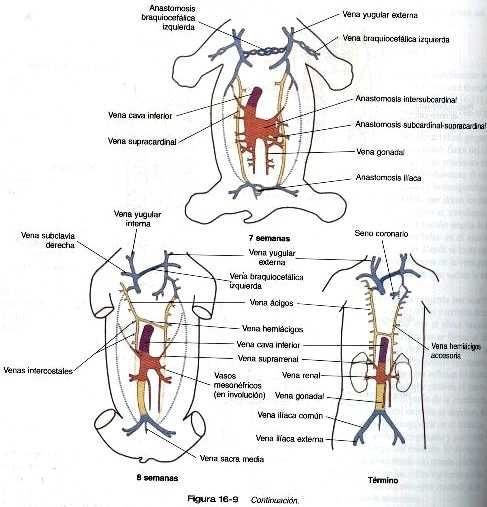{kind=link}
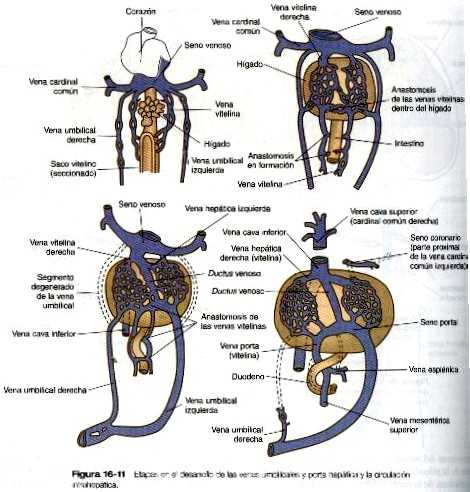{kind=link}
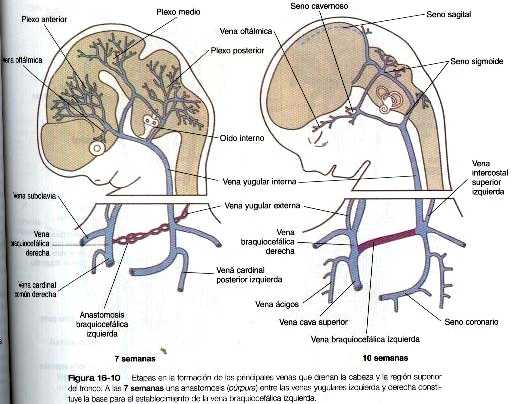{kind=link}
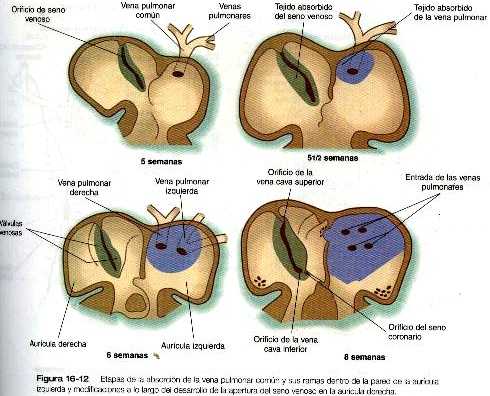{kind=link}
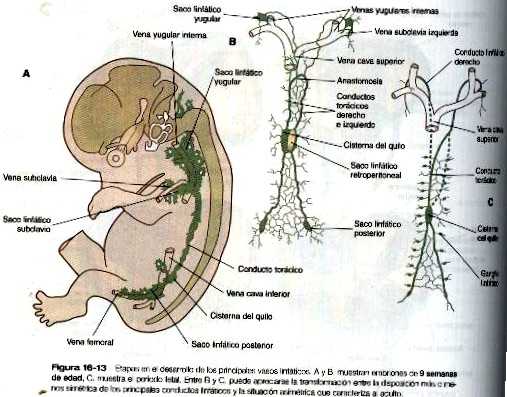{kind=link}
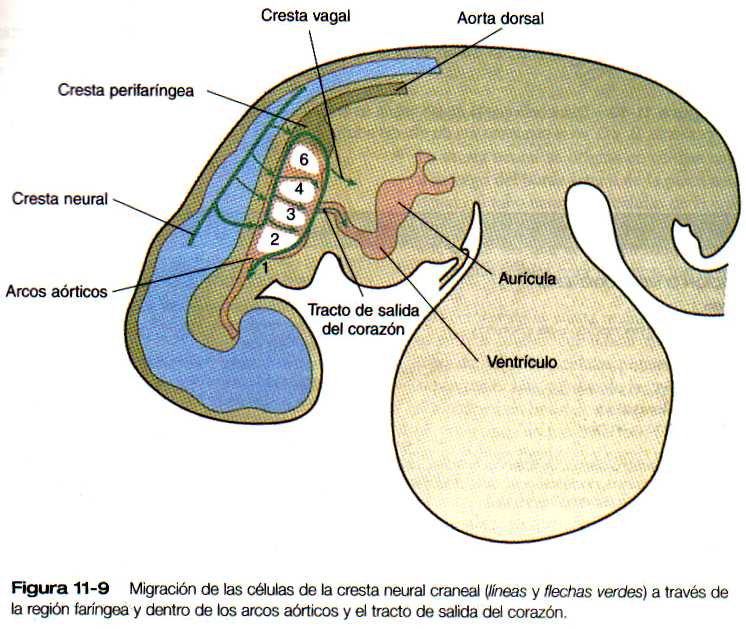{kind=link}
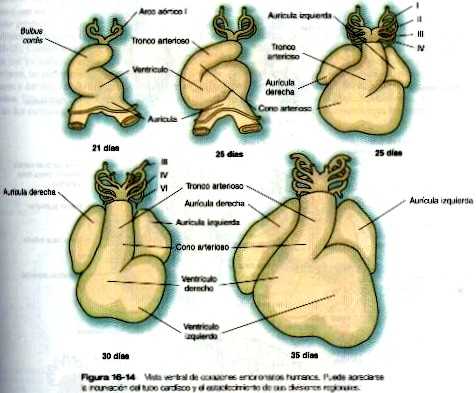{kind=link}
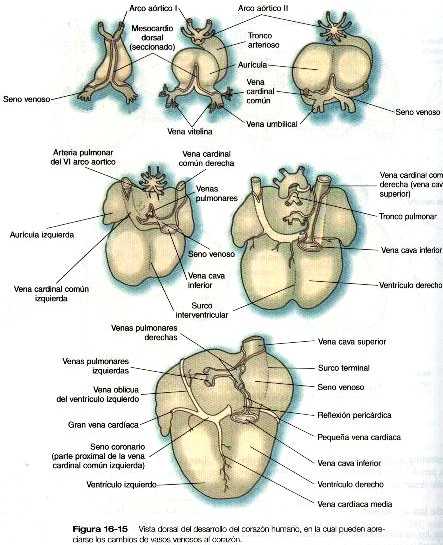{kind=link}
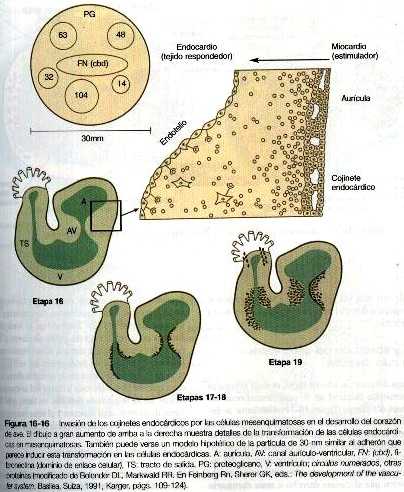{kind=link}
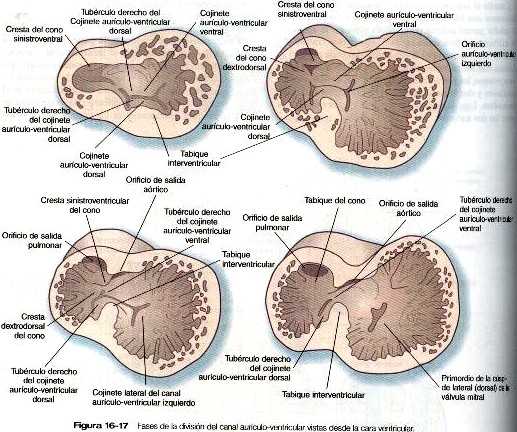{kind=link}
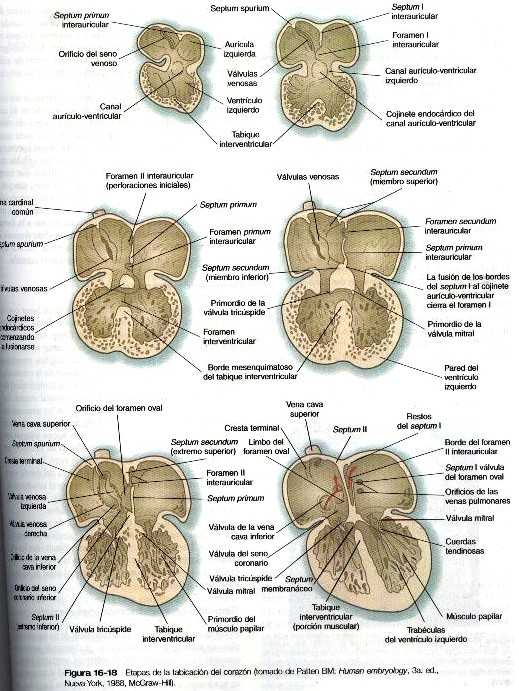{kind=link}
{kind=link}
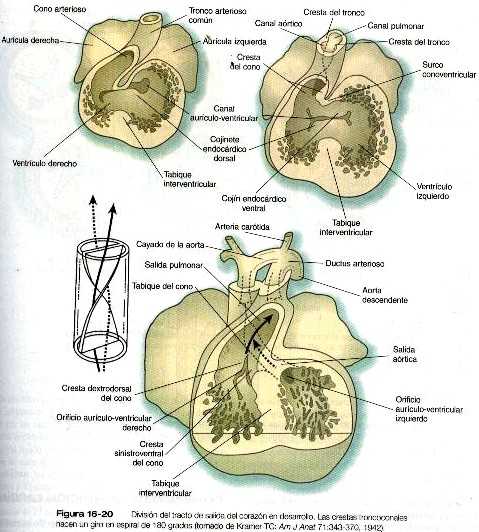{kind=link}
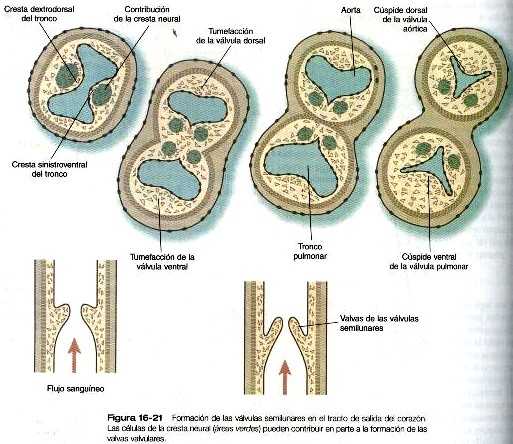{kind=link}
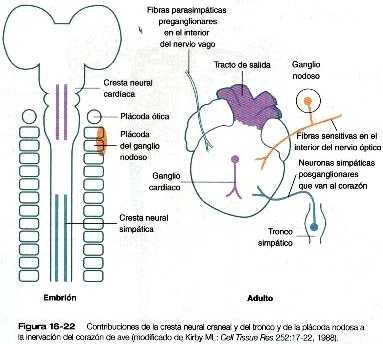{kind=link}
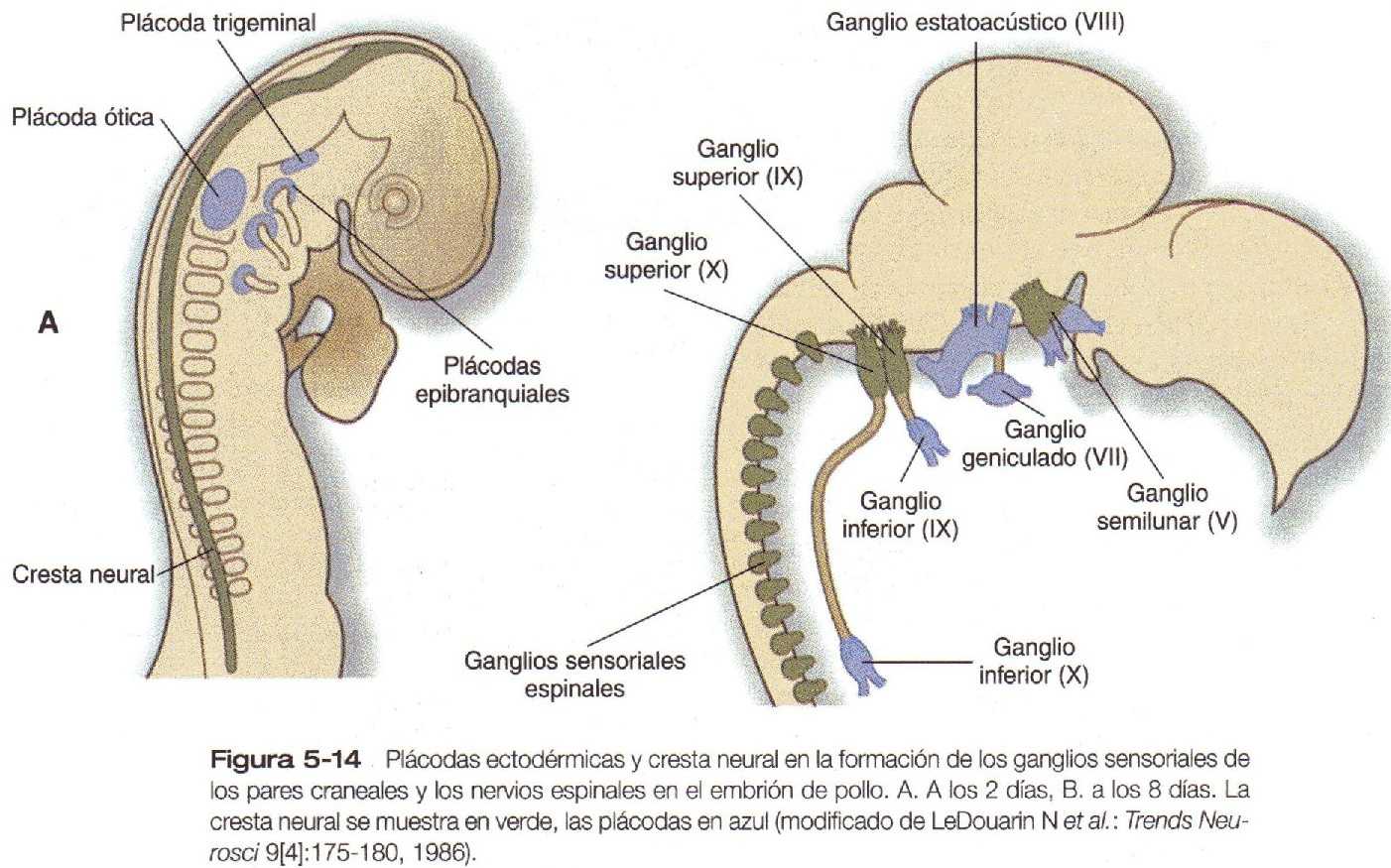{kind=link}
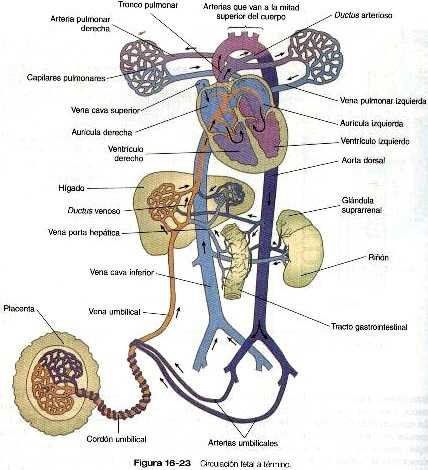{kind=link}
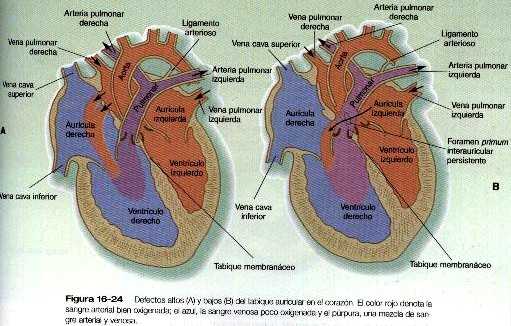{kind=link}
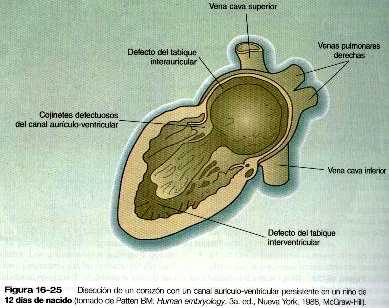{kind=link}
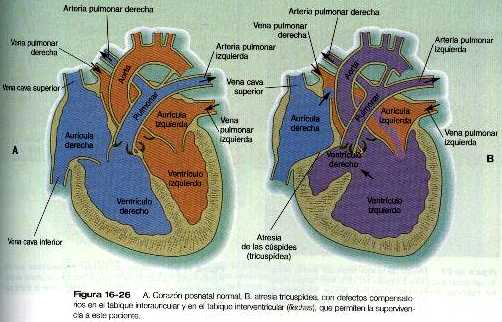{kind=link}
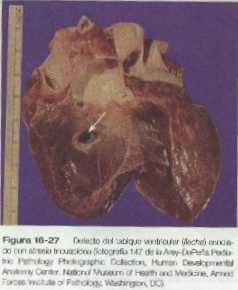{kind=link}
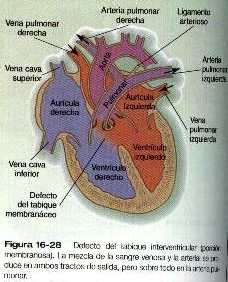{kind=link}
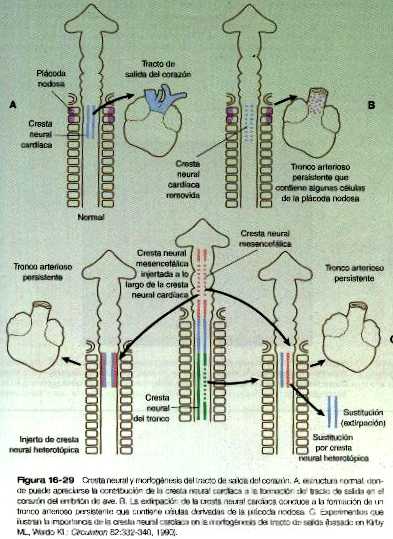{kind=link}
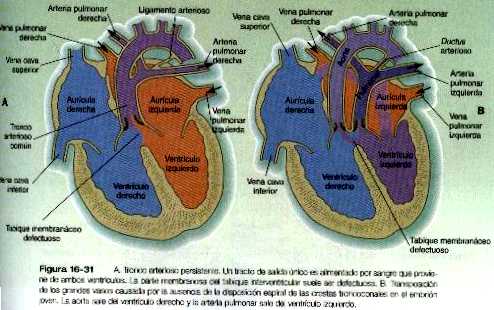{kind=link}
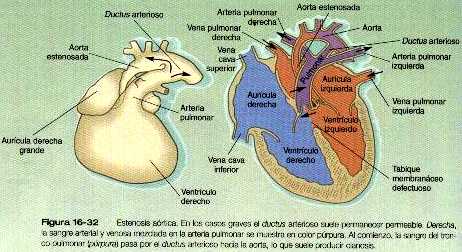{kind=link}
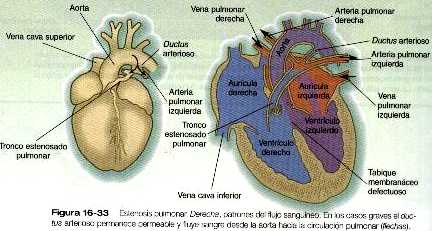{kind=link}
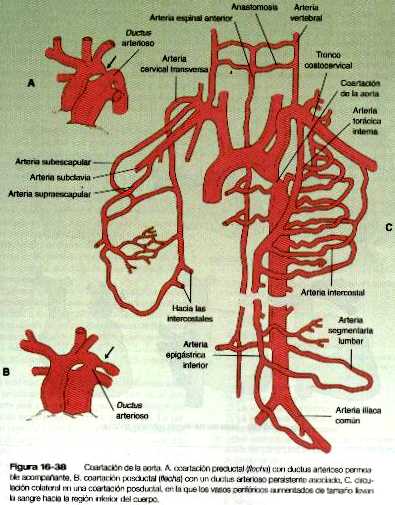{kind=link}
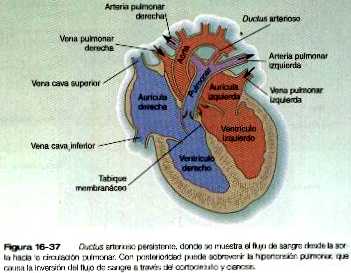{kind=link}
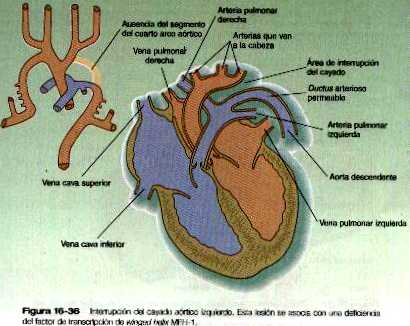{kind=link}
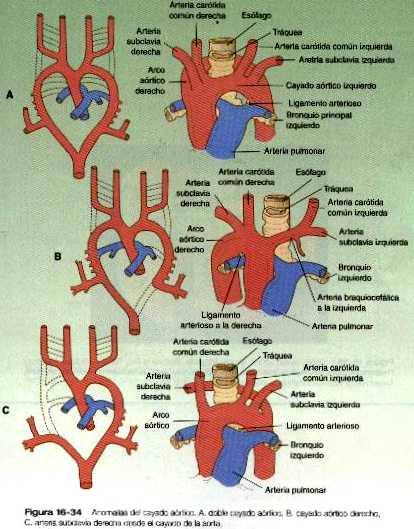{kind=link}
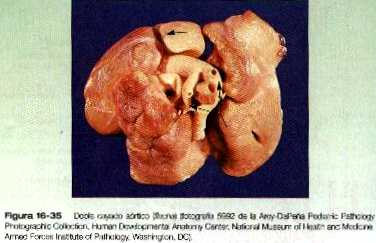{kind=link}
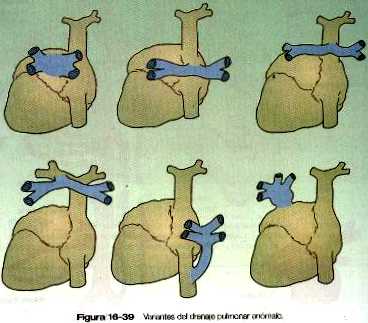{kind=link}
{kind=link}