Capítulo 5.
ESTABLECIMIENTO DEL PLAN CORPORAL BÁSICO DEL EMBRIÓN
BASE MOLECULAR DE LA ORGANIZACIÓN DEL CUERPO DE LOS VERTEBRADOS
Procesos moleculares fundamentales en el desarrollo.
Factores de trascripción.
Moléculas de activación (señal).
Moléculas receptoras.
Transducción de las señales
Moléculas y morfología
DESARROLLO DE LA CAPA GERMINAL ECTODÉRMICA
Neurulación: formación del tubo neural
Segmentación del tubo neural.
Cresta Neural
Plácodas sensoriales e inducciones secundarias en la región craneal
DESARROLLO DE LA CAPA GERMINAL MESODERMICA
Desarrollo básico de la capa mesodérmica
Mesodermo paraxial
Formación de los somitas individuales.
Organización del somita y del plan básico de la segmentación del cuerpo.
Mesodermo intermedio
Mesodermo de la placa (lámina) lateral.
Formación del celoma.
Mesodermo extraembrionario y tallo de conexión (corporal).
Primeras etapas en la formación del sistema circulatorio.
Corazón y grandes vasos.
Sangre y vasos sanguíneos
DESARROLLO DE LA CAPA GERMINAL ENDODÉRMICA
ESTRUCTURA BÁSICA DEL EMBRIÓN DE CUATRO SEMANAS
Apariencia macroscópica
Sistema Circulatorio
Después de terminada la gastrulación, el embrión propiamente dicho es de un disco plano trilaminar que contiene las capas germinales ectodermo, mesodermo y endodermo. Su eje cefalocaudal esta definido por la localización de la estría primitiva. A causa del patrón de migración celular a través de la estría primitiva y de la regresión de la estría hacia el extremo caudal del embrión, se establece un notable gradiente de diferenciación cráneo-caudal. Este gradiente es marcado inicialmente por la formación de la notocorda y mas adelante por la aparición de la placa neural, que proviene de la inducción primaria del ectodermo dorsal por la notocorda.
Durante la tercera semana, a pesar de la apariencia morfológica relativamente carente de rasgos del embrión pos-gastrulación, hay evidencias de que durante este periodo, e incluso antes, el plan básico del cuerpo se está estableciendo mediante la expresión de un patrón de genes específicos. Una de las primeras manifestaciones morfológicas de este patrón es la segmentación regular que se hace evidente a lo largo del eje craneocaudal del embrión. Este plan segmentario, que es una característica dominante de todos los embriones jóvenes, se hace menos evidente conforme avanza el desarrollo. No obstante, incluso en el adulto la organización regular de las vértebras, las costillas y los nervios raquídeos persiste como un recuerdo de pasado filogenético y ontogenético segmentado del ser humano. Sólo recientemente los embriólogos han comenzando a entender los controles celulares y moleculares que subyacen en el proceso de la segmentación.
Otro cambio crucial para la comprensión de la organización fundamental del plan corporal es el plegamiento lateral del embrión joven, que así pasa de ser una estructura conformada por tres capas celulares en esencia planas, apiladas y similares a un hojaldre (las capas germinales embrionarias primarias) a una estructura cilíndrica, con el ectodermo en el exterior, el endodermo en el interior y el mesodermo entre los dos anteriores. La base celular para el plegamiento lateral todavía es mejor descrita que comprendida.
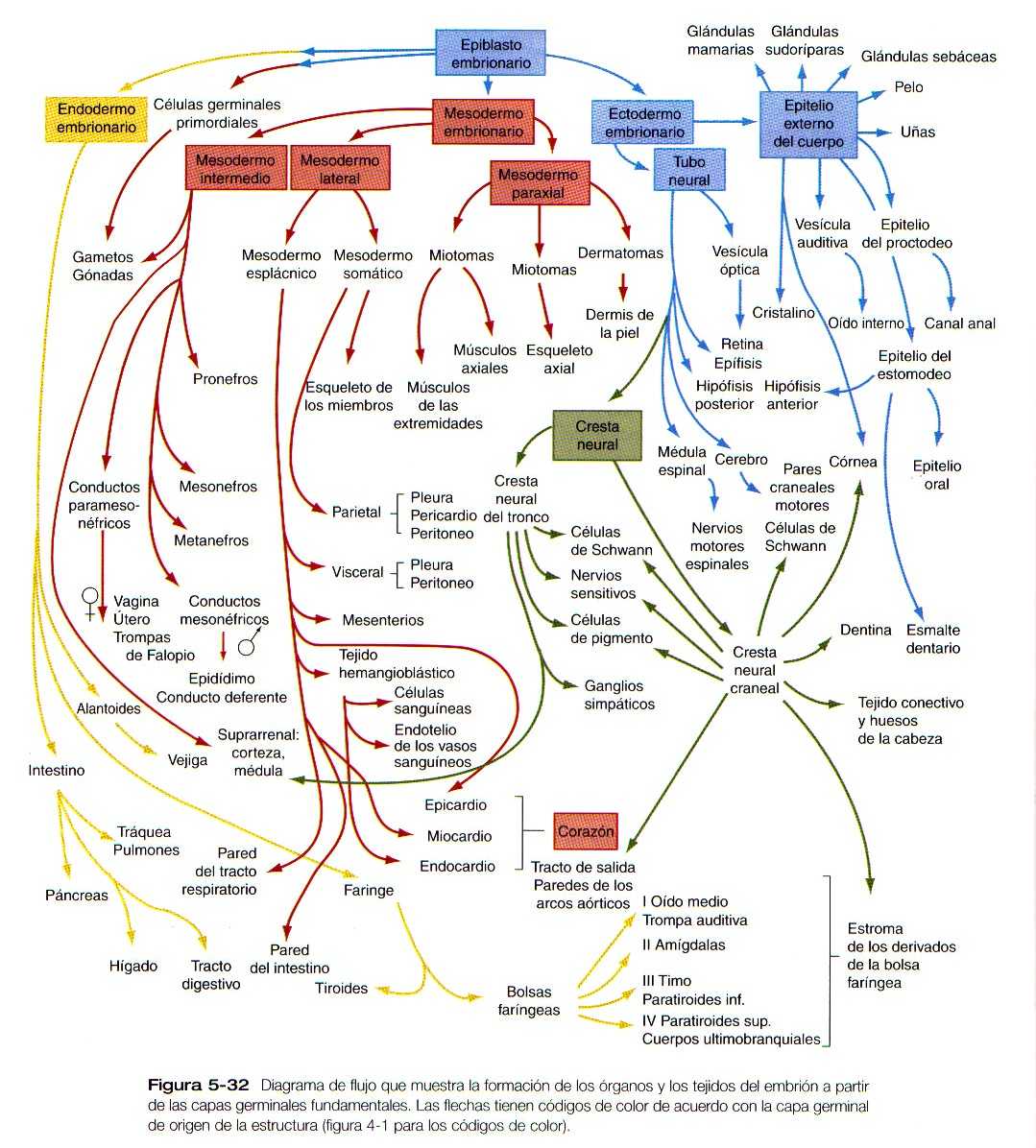
Este capitulo se centra en el establecimiento del plan global del cuerpo. Además, describe la aparición de los primordios de los principales sistemas orgánicos a partir de las capas germinales primarias indiferenciadas. (Figura 5_32).
BASE MOLECULAR DE LA ORGANIZACIÓN DEL CUERPO DE LOS VERTEBRADOS
Desde 1990 la aplicación de nuevas técnicas en biológica molecular ha revolucionado la comprensión de los mecanismos tanto del desarrollo embrionario normal como del anormal. No es posible tener una comprensión actual del desarrollo embrionario sin integrar los aspectos moleculares y morfológicos fundamentales de la embriología. Esta sección sirve como introducción general sobre las familias más importantes de moléculas que se sabe que dirigen el desarrollo embrionario. Dado que casi cada semana se está informando acerca de nuevos genes activos en el desarrollo, el enfoque usado en este texto es presentar ejemplos importantes que ilustren los correlatos moleculares o el control de los sistemas en desarrollo, en vez de pretender dar una explicación totalizante en modo alguno.
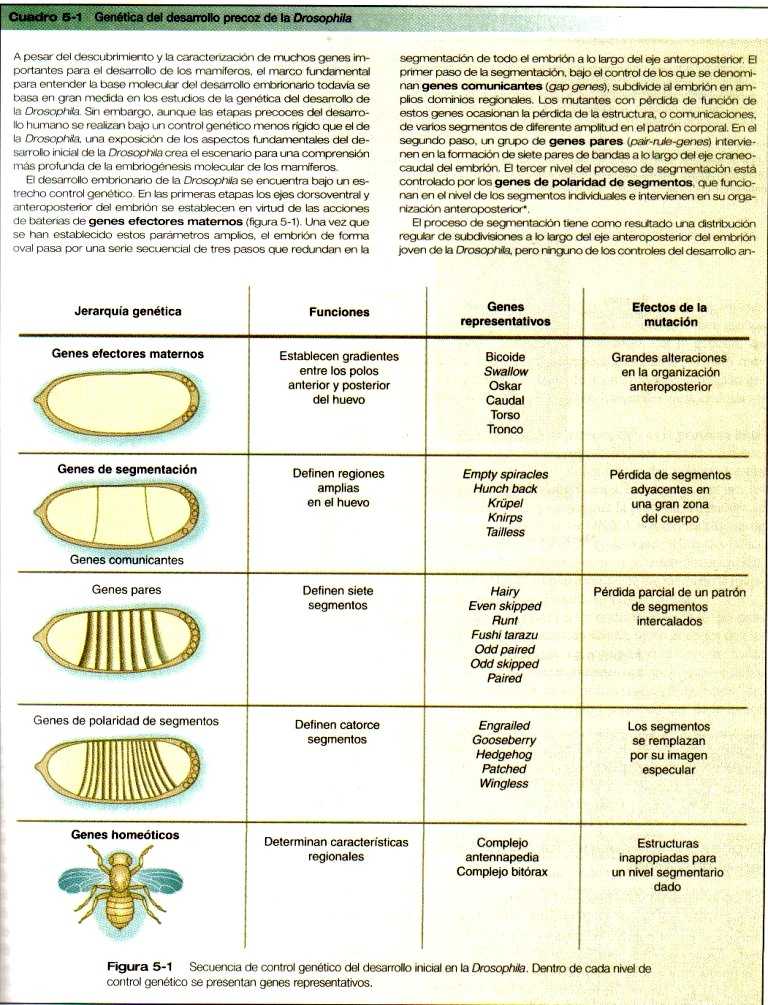
Uno de los hallazgos más importantes del último decenio es el mantenimiento y la conservación de los genes que guían el desarrollo. Los estudios de secuenciación han demostrado muy pocos cambios en las bases de los nucleótidos de muchos genes reguladores desde el punto de vista del desarrollo, que están representados en especies que van desde los gusanos hasta la Drosophila, y hasta los seres humanos. A causa de este conservacionismo filogenético, ha sido posible identificar en los mamíferos zonas homólogas de los genes de los que se sabe, por los estudios genéticos, que tienen importantes funciones en el desarrollo de las especies. (Cuadro 5_1) También es cada vez más claro que el mismo gen puede tener diferentes funciones en distintas fases del desarrollo y en órganos distintos. Tanto antes como después del nacimiento, pueden expresarse genes específicos en procesos normales y anormales. Uno de los temas actuales en la investigación del cáncer es el papel que tienen formas mutantes de genes importantes desde el punto de vista del desarrollo (por ejemplo, los protoncogenes), tienen en la conversión de las células normales en células tumorales.
Procesos moleculares fundamentales en el desarrollo.
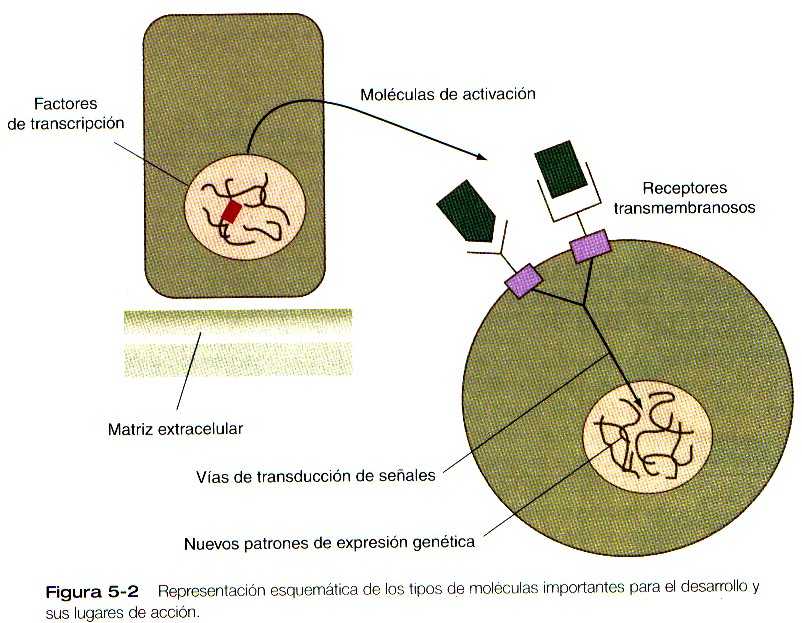
Desde el punto de vista funcional, muchas de las moléculas importantes que guían el desarrollo embrionario pueden agruparse en un número relativamente pequeño de categorías; algunas de ellas permanecen dentro de las células que las producen y actúan como factores de transcripción. (Figura 5_2). Los factores de transcripción son proteínas poseedoras de dominios que se fijan al ácido desoxirribonucleico (DNA) de las regiones promotoras o intensificadoras de genes específicos. También poseen un dominio que interactúa con la polimerasa II del ácido ribonucleico (RNA) u otros factores de transcripción y, en consecuencia, regulan la cantidad de ácido ribonucleico mensajero que el gen produce. Otras moléculas actúan como moléculas de activación (señales) intercelulares. Tales moléculas salen de las células que las producen y ejercen sus efectos en otras células, que pueden ser vecinas o estar localizadas a gran distancia de las células productoras de las moléculas de activación. Muchas de estas moléculas son miembros de grandes familias de proteínas relacionadas entre si y llamadas factores de crecimiento. Para ejercer su efecto, las moléculas de activación suelen unirse a moléculas receptoras que a menudo son proteínas de transmembranas que sobresalen a través de la membrana plasmática de las células a las que están ligadas.
Cuando estas moléculas receptoras forman complejos con las moléculas de activación, desencadenan una cascada de sucesos en la vía (corriente) de la transducción de señales que transmite la señal al núcleo de la célula diana. Esta señal influye sobre la naturaleza de los productos genéticos producidos por esta célula y a menudo en el futuro desarrollo de la célula. En la última década, se han identificado muchas nuevas moléculas importantes para el desarrollo y se han descrito sus patrones de expresión durante el desarrollo. Se están haciendo grandes esfuerzos tendentes a aclarar las cascadas moleculares que controlan los procesos del desarrollo (por ejemplo, los acontecimientos moleculares que estimulan la producción de una molécula importante para el desarrollo y el mecanismo por el cual esa molécula a su vez ejerce un efecto desencadenante sobre otros procesos sintéticos).
Factores de transcripción.
Muchas familias de moléculas actúan como factores de transcripción. Algunos de ellos son generales y se encuentran en prácticamente todas las células de un organismo. Otros factores de transcripción son específicos para ciertos tipos de células y etapas del desarrollo. Por lo general, los factores de transcripción específicos (Tabla 5_1) son muy importantes en la iniciación de los patrones de expresión genética que redundan en cambios esenciales para el desarrollo.
Una clase de factores de transcripción es la proteína básica hélice-asa-hélice (helix-loop-helix) que contiene una corta sección de aminoácidos en los cuales dos hélices a están separadas por un asa de aminoácidos. Esta región básica adyacente, permite que la proteína reguladora se fije a secuencias específicas de DNA. Las regiones básicas de estas proteínas fijan el DNA, y el dominio hélice-asa-hélice interviene en la homodimerización o la heterodimerización. Esta configuración es común a varios de los factores de transcripción que regulan la biogénesis. (Figura 8_27).
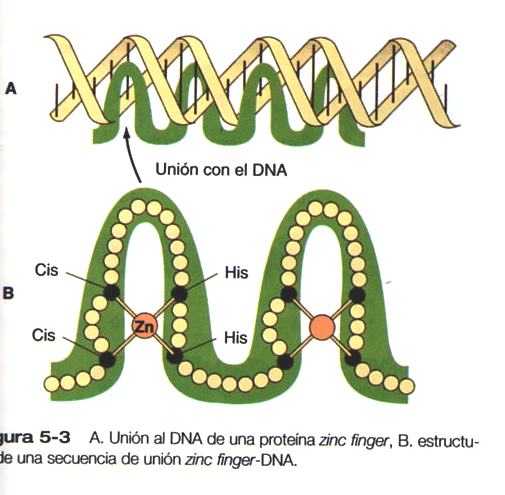
Otra familia de factores de transcripción es la de las proteínas zinc finger. En estas proteínas, unas unidades de cisteina e histidina localizadas con regularidad se unen mediante iones de cinc que hacen que la cadena polipeptídica se doble para formar estructuras digitiformes (Figura 5_3). Estos “dedos” pueden insertarse en regiones especificas de la hélice del DNA.
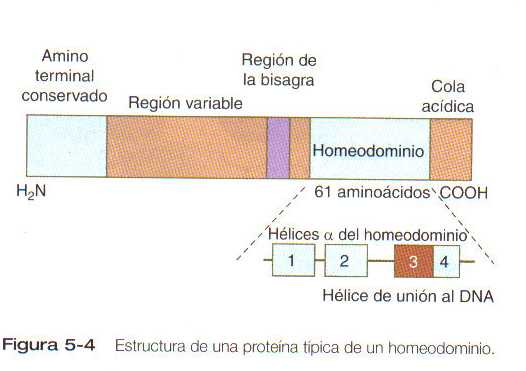
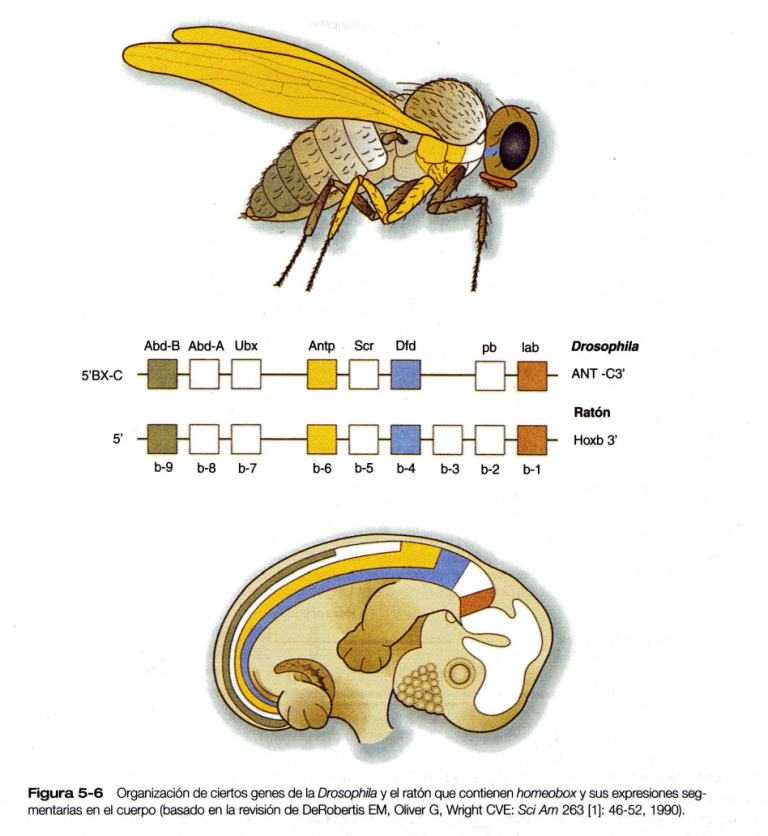
Uno de los tipos más importante de factores de transcripción está representado por las proteínas de homeodominio. Estas, contienen un homeodominio muy conservado de 61 aminoácidos, que es un tipo de región hélice-asa-hélice. (Figura 5_4). Los 183 nucleótidos que codifican al homeodominio reciben el nombre colectivo de homeobox. Las regiones de homeobox fueron descubiertas en los genes homeóticos de los complejos antennapedia y bitórax de la Drosophila, (Figura 5_1) de aquí su nombre. Esta designación puede ser confusa para los estudiantes porque, desde su descripción inicial, se han encontrado homeobox en otros genes relacionados de forma mas distante fuera del grupo de genes homeóticos.
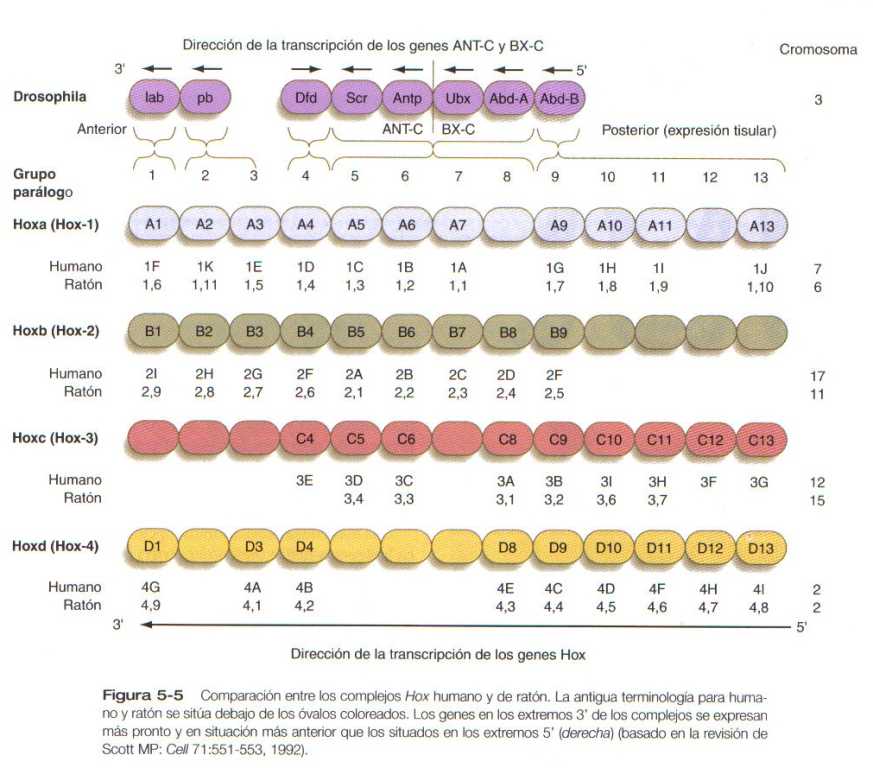
El complejo antennapedia bitórax de la Drosophila consta de ocho genes que contienen homeobox localizados en dos regiones en un cromosoma. Los ratones y los seres humanos poseen al menos 38 genes homeobox homólogos (que en los vertebrados se llaman genes Hox), que se encuentran en cuatro regiones en cuatro cromosomas diferentes. (Figura 5_5). Los genes Hox de los cuatro cromosomas de los mamíferos se organizan en 13 grupos parálogos.
Los genes Hox de los vertebrados están fuertemente implicados en la segmentación craneocaudal del cuerpo, y su expresión espaciotemporal procede de acuerdo con algunas reglas de notoria regularidad. Los genes se activan y se expresan conforme a una secuencia estricta en la dirección 3 a 5, que corresponde a sus posiciones en los cromosomas. En consecuencia, tanto en la Drosophila y como en los mamíferos, los genes 3 se expresan primero y en situación mas anterior que los genes 5 (Figura 5_6). Las mutaciones de los genes Hox dan como resultado transformaciones morfológicas anómalas de las estructuras segmentarías normas en las cuales suele expresarse un gen específico. En general, las mutaciones de perdida de la función redundan en transformaciones de posterior en anterior (es decir, que las células de un segmento dado forman el equivalente estructural del próximo segmento más anterior), y las mutaciones de ganancia de función dan como resultado transformaciones estructurales de anterior en posterior.
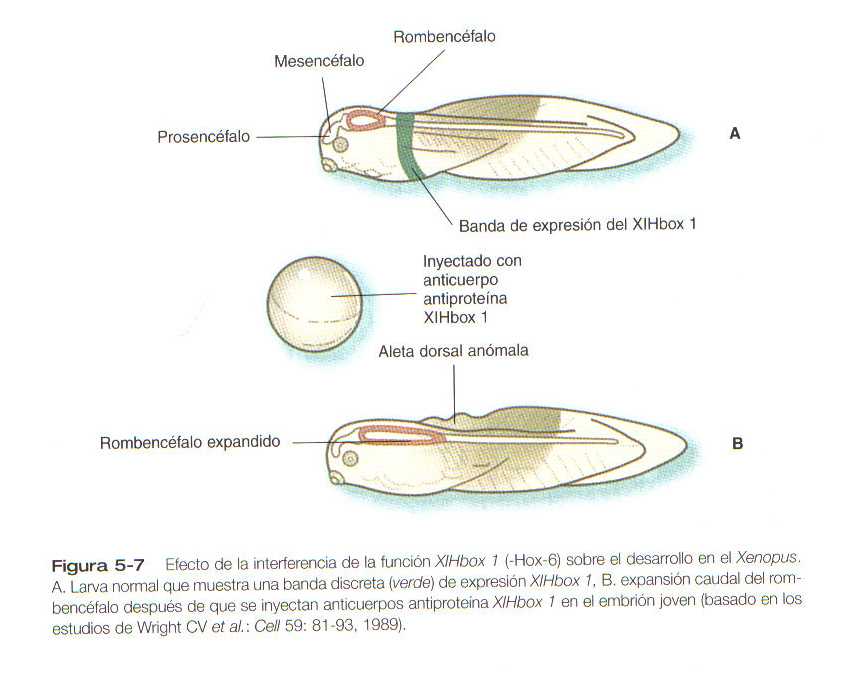
La figura 5_7 ilustra un experimento en el cual la inyección de un anticuerpo en un gen homeobox de un embrión inicial de sapo tuvo como resultado la transformación de la médula espinal anterior en un rombencéfalo expandido.
Los sucesos moleculares que tiene lugar tanto corriente arriba como corriente abajo en flujo de expansión de los genes Hox aun no se conocen del todo pero el factor de crecimiento de los fibroblastos (FGF) puede causar una acción selectiva de los genes posteriores que contienen homeobox, mientras que el factor B de transformación del crecimiento (TGF-B) puede activar de forma selectiva los genes Hox. El acido retinoico, un poderoso agente teratógeno en los seres humanos, puede causar una posteriorización de la expresión de los genes Hox con las consecuencias aberraciones morfológicas.
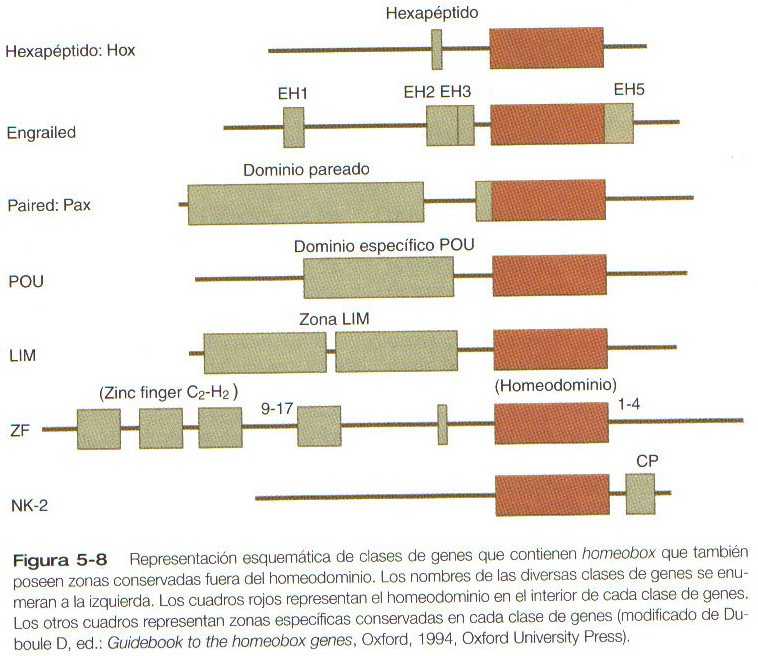
Otras familias de genes contienen no sólo un homeobox sino también otras secuencias conservadas (Figura 5_8). Algunas, como los genes Engrailed y Lim, constan tan sólo de unos pocos miembros por grupo, pero otros, como los genes POU y paired (Pax), constituyen grandes familias, algunos de cuyos miembros se expresan en muchas estructuras en desarrollo.
La familia de los genes POU obtiene su nombre de los primeros genes identificados, es decir, Pit-1, un gen que se expresa sólo en la pituitaria. Oct-1 y Oct-2, y Unc-86, un gen expresado en un nematodo. Los genes de familia POU contienen, además de un homeobox, una región que codifica 75 aminoácidos, que también se une al DNA a través de una estructura de hélice-asa-hélice (helix-loop-helix).
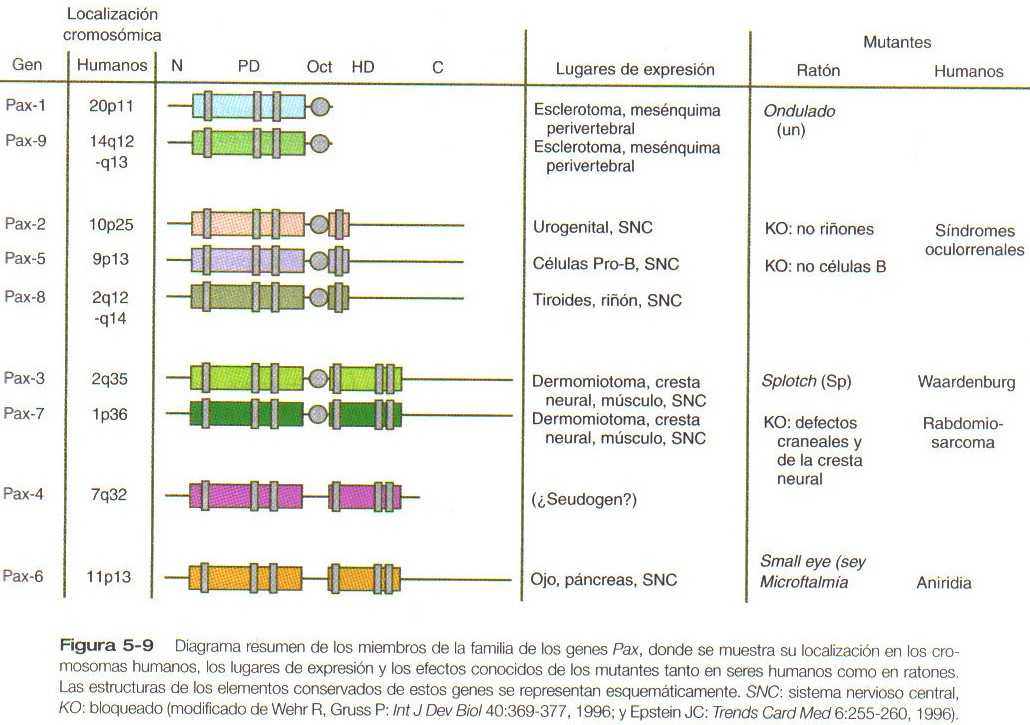
La familia de los genes Pax, que consta de nueve miembros conocidos, es un grupo importante de genes que intervienen en muchos aspectos del desarrollo de los mamíferos (Figura 5_9). Todas las proteínas pax contienen un dominio par de 128 aminoácidos, que se fija al DNA. Diversos miembros del grupo también contienen dominios parciales o completos o de homeobox y una secuencia conservada de un octapéptido.
Moléculas de activación (señal).
Las moléculas de activación o señal. Muchas de las cuales son factores peptídicos de crecimiento, son mediadoras en la mayoría de las interacciones, como inducciones que se dan entre grupos de células en los embriones. Muchas de las moléculas de activación que son muy importantes en el desarrollo embrionario pertenecen a dos grandes familias: las familias TGF-B y FCF. Los miembros de otra familia de moléculas de activación descubierta hace poco tiempo, las proteínas hedgehog, median algunas de las interacciones inductivas más potentes descubiertas hasta la fecha. El factor de crecimiento neural, que estimula el crecimiento de las neuronas sensoriales y simpáticas, fue el primer factor de crecimiento que se investigo intensamente, trabajo que comenzó en el decenio de 1950.
La familia TGF-B consta de un gran número de moléculas (hasta 30 genes) que desempeñan una amplia variedad de funciones durante la embriogénesis y la vida posnatal (Tabla 5_2). Los miembros de la familia FGF (FGF-1 a FGF-9) de forma similar cumplen diversas funciones en la embriogénesis, que van desde la estimulación de la proliferación de las células mesenquimatosas hasta la inducción de la elongación de la yema de las extremidades hasta la estimulación del crecimiento de los capilares a la proliferación y la supervivencia de ciertas neuronas.
Una de las familias más influyentes de las moléculas de activación en el embrión es la de las proteínas hedgehog desempeña un papel crucial al ser producida en diversos centros de organización del embrión (Tabla 5_3). La proteína sonic hedgehog es especial porque sufre autodivision proteo lítica, de la cual resulta un péptido de 19kDa con N-Terminal y un péptido de 27 kDa con C Terminal. El péptido N-Terminal, que establece una unión covalente con el colesterol durante el procesamiento proteolítico, es la porción de la molécula dividida que parece incorporar la mayoría de las propiedades importantes de activacion de la molécula, hasta la fecha, se han descrito en los mamíferos tres formas de hedgehog (sonic, india y desert) derivados de tres genes diferentes. Después de unirse a una molécula receptora en la célula diana, las señal del sonic hedgehog estimula a la célula diana a producir nuevos productos génicos o a realizar nuevas vías de diferenciación.
Moléculas receptoras.
Para que las moléculas intercelulares de señal ejerzan un efecto en las células diana, tiene que interactuar normalmente con los receptores de dichas células. La mayoría de los receptores están localizados en la superficie celular, pero algunos en especial los de las moléculas liposolubles como los esteroides, los retinoides y las hormonas tiroideas son intracelulares.
Los receptores de la superficie celular suelen ser proteínas transmenbranosas con dominios extracelulares, transmembranosos y cito plásticos (Figura 5_2). El dominio extracelular contiene un lugar de unión para el ligando, que suele ser una hormona, una citosina o un factor de crecimiento. Cuando el ligando se une al receptor, efectúa un cambio de conformación en el dominio citoplásmico de la molécula receptora. Los receptores de superficie celular son de los siguientes tipos principales: los que tienen actividad intrínseca de proteincinasa y los que usan al sistema de segundo mensajero para activar las proteincinasas citoplásmicas. Un ejemplo del primer tipo es la familia de los receptores para los FGF, en los cuales el dominio citoplásmico posee actividad de tirosincinasa. Los receptores de los factores de crecimiento de la superfamilia de los TFG-B también son de este tipo, pero en ellos el dominio citoplásmico contiene actividad de serina/treonina cinasa. En los receptores de superficie celular del segundo tipo, la actividad de la protincinasa esta separada de la molécula misma del receptor. Este tipo de receptor también es activado por la unión con el ligando (por ejemplo, neurotransmisores, hormonas peptídicas, factores de crecimiento), pero se requiere una serie de pasos intermedios para activar las proteincinasas citoplásmicas.
Transducción de las señales
La transducción de las señales es el proceso por el cual la señal enviada por el primer mensajero (es decir, el factor de crecimiento u otra molécula de activación) se expresa como una respuesta intracelular. La señal de transducción comienza cuando el primer mensajero, O-ligando, se une a un receptor y cambia su conformación. En el caso de los receptores que no poseen actividad intrínseca de las proteincinasas, la unión del ligado al receptor estimula una reacción en cadena o cascada, que conduce a la producción de un segundo mensajero, que activa las proteincinasas citoplásmicas. Una cascada típica (Figura 5_2) consta de una serie de pasos por los cuales el receptor activado, actuando a través de proteínas G (proteínas que ligan trifosfato de guanosina y disfosfato de guanosina), estimula una enzima efectora (por ejemplo, la adenilciclasa) para convertir las moléculas precursoras en segundos mensajeros (dos segundos mensajeros importantes son el monofosfato cíclico de adenosina y el trofosfato de inosina mas diacilglicerol). A continuación, el segundo mensajero activa las proteincinasas citoplásmicas, que fosforilan (añaden grupos fosfato) las proteínas diana, ya sea activándolas o desactivándolas. Después de otros pasos adicionales que suelen implicar a otras cinasas y la translocacion de las moléculas activadas hacia el núcleo, se modifica la transcripción del DNA. De esta manera la cascada de la transducción lleva a una respuesta celular que en el desarrollo embrionario podría ser la transformación de un tipo celular durante la diferenciación o la producción de un producto especifico por parte de la célula diana.
Moléculas y morfología
Se ha vuelto casi un axioma en los estudios del desarrollo que la morfología se construye sobre un patrón molecular. El patrón puede ser de amplio alcance, como en el caso de la expresión del gen Hox en la segmentación del eje corporal, o puede tener un dominio muy restringido, como la inducción de la córnea por el cristalino. Muchos aspectos de la morfogénesis son aún tan mal conocidos que todavía esta por descubrirse la base molecular. En el resto de este texto se presentan ejemplos del control molecular de la diferenciación celular o de la morfogénesis, en especial cuando ofrecen claves sobre los mecanismos que pueden explicar un desarrollo anómalo.
DESARROLLO DE LA CAPA GERMINAL ECTODÉRMICA
Neurulación: formación del tubo neural
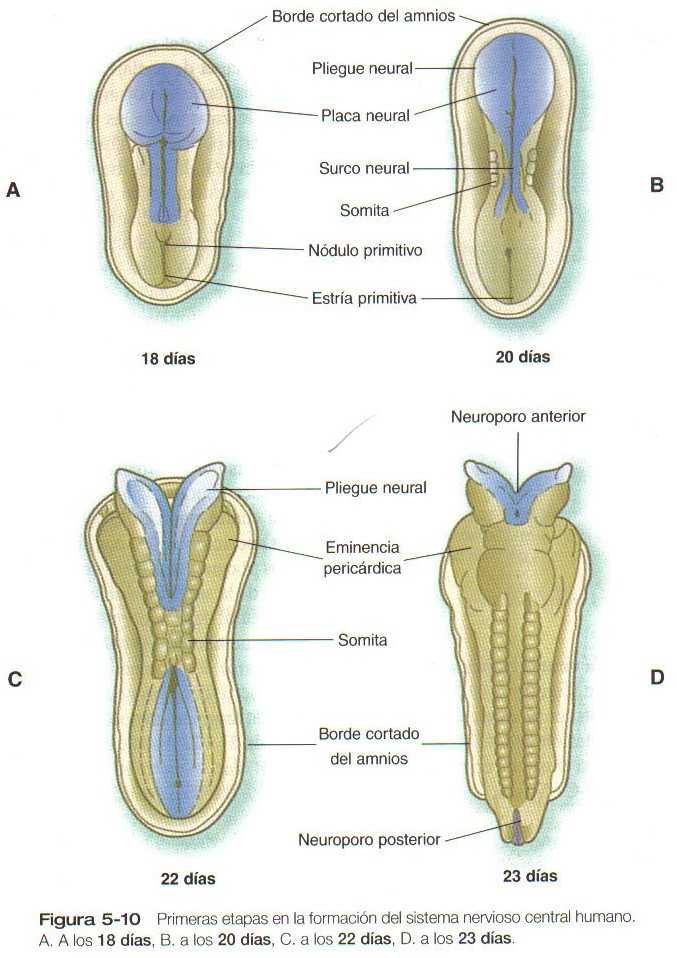
La principal respuesta morfológica inicial del ectodermo embrionario a la inducción neural es un incremento en la altura de las células que están destinadas a convertirse en componentes del sistema nervioso. Estas células transformadas se hacen visibles como una placa neural engrosada en la superficie dorsal del embrión joven (Figura 5_10 A Y 5_11 A). Invisible, pero también importante, es la restricción de la expresión de las moléculas de adhesión celular (CAM) de la N-CAM y la L-CAM/E-cadherina existente en el ectodermo antes de la inducción a sólo la N-CAM y la N-cadherina en la placa neural.
La primera de las cuatro etapas principales en formación del tubo neural es la transformación del ectodermo embrionario indiferenciado en una placa neural engrosada. La principal actividad de la segunda etapa es la remodelación de los contornos globales de la placa neural de forma que se hace más estrecha y larga. El moldeamiento de la placa neural se logra en gran medida por cambios regionales específicos en la forma de las células neuroepiteliales (por ejemplo, un aumento en altura de la célula a expensas del tamaño de la zona basal de la misma) y mediante la reorganización de las relaciones y uniones entre las células.
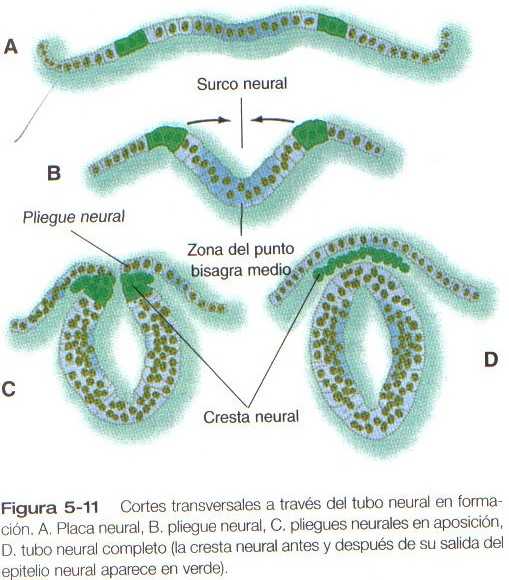
La tercera etapa principal en el proceso de la neurulación es el plegamiento lateral de la placa neural, cuyo resultado es la elevación de cada lado de la placa neural, cuyo resultado es la elevación de cada lado de la placa neural a lo largo de un surco neural localizado en la línea media (Figura 5_10B y 5_11B). Se han propuesto muchas explicaciones para el plegamiento lateral de la placa neural y el cierre posterior del tubo neural. La mayoría ha invocado un mecanismo único o dominante, pero actualmente es cada vez mas evidente que el plegamiento lateral es el resultado de varios mecanismos específicos de la región, tanto extrínsecos como intrinseco0s a la placa neural.
La línea medioventral de la placa neural, en ocasiones llamada zona del punto bisagra medio, parece actuar como una zona del punto bisagra medio, parece actuar como una zona de anclaje alrededor de la cual se elevan los dos lados formando un ángulo agudo respecto de la horizontal. En la zona media, es posible explicar el plegamiento en gran parte por cambios inducidos por la notocorda en la forma de las células neureopiteliales de la placa neural. Estas células se estrechan en el vértice y se ensanchan en la base (Figura 5_11 C ) mediante una combinación de una posición basal del núcleo (que causa una expansión lateral de la célula en esa área) y una contracción similar a un esfínter provocada por un anillo de microfilamentos que contienen actina en el citoplasma apical. Durante todo el plegamiento lateral de la placa neural en la región de la medula espinal, gran parte del área parietal de la placa neural permanece plana (Figura 5_11b). La elevación de los pliegues neurales parece lograrse en gran medida mediante factores extrínsecos al epitelio neural, en especial fuerzas de impulso generadas por la expansión de la superficie epitelial lateral a la placa neural.
La cuarta etapa en la formación del tubo neural consiste en la aposición de las dos superficies apicales más laterales de los pliegues neurales, su fusión (mediada por glicoconjugados de la superficie celular), y la separación completa del tubo neural del ectodermo suprayacente (Figura 5_11C y D). Al mismo tiempo, las células de la cresta neural comienzan a separarse del tubo neural.
El cierre del tubo neural comienza casi a mitad de camino en la longitud craneocaudal del sistema nervioso del embrión de 21 a 22 días de edad (Figura 5_10C). En los siguientes dos días, el cierre se extiende tanto en dirección cefálica como en dirección caudal, de forma que su superficie se parece al cierre de una cremallera de doble sentido. Las partes cefálica y caudal del tubo neural que no se cierran reciben el nombre de neuroporos anterior (craneal) y posterior (caudal). Los neuroporos, también terminan por cerrarse, de manera que la organización de todo el futuro sistema nervioso central recuerda a un cilindro hueco irregular sellado en ambos extremos. En ocasiones uno o ambos neuroporos permanecen abiertos, de lo cual resultan graves malformaciones congénitas.
En la región caudal al neuroporo posterior, el resto del tubo neural (más prominente en los animales de cola larga) se forma mediante el proceso de neurulación secundaria. En los mamíferos, este proceso parece comenzar con la formación de una condensación de las células mesenquimatosa se forma un canal directamente por cavitación (formación de un espacio en una masa de células). Este canal central hace continuidad con el que se formo durante la neurulación primaria en virtud del plegamiento lateral de la placa neural y el cierre del neuroporo posterior. A causa del escaso desarrollo de la yema de la cola, la neurulación secundaria no es un proceso importante en los seres humanos.
Segmentación del tubo neural.
Poco después de que el tubo neural ha tomado forma, es posible distinguir entre la región del futuro encéfalo y el resto de la médula espinal. La región a partir de la cual se va a formar el encéfalo sufre una serie de subdivisiones que constituyen la base para la organización macroscópica fundamental del encéfalo adulto. El primer grupo de subdivisiones tiene como resultado un encéfalo que consta de tres partes: uno anterior (pros encéfalo), un encéfalo medio (mesencéfalo) y uno posterior (rombencéfalo).
Diversos mecanismos del desarrollo trabajan al unísono para dividir el futuro encéfalo en las subdivisiones estructurales y funcionales necesarias. Muy pronto en el desarrollo, quizás durante o la inducción de la placa neural poco después de ella, dos amplias regiones del sistema nervioso en desarrollo se bloquean como resultado de las influencias de tejidos subyacentes. Mediante la acción de moléculas de señal (por ejemplo, la proteína Lim-1 y una proteína secretada y recientemente identificada llamada cerberus), la placa mesodérmica pro cordal especifica el patrón inicial del desarrollo para el prosencéfalo dentro de la placa neural a medida que comienza a formarse la parte rostral de la misma y de la cabeza. Posteriormente, se defienden el rombencéfalo y la médula espinal mediante las señales emanadas de la notocorda subyacente y del mesodermo paraxial adyacente.
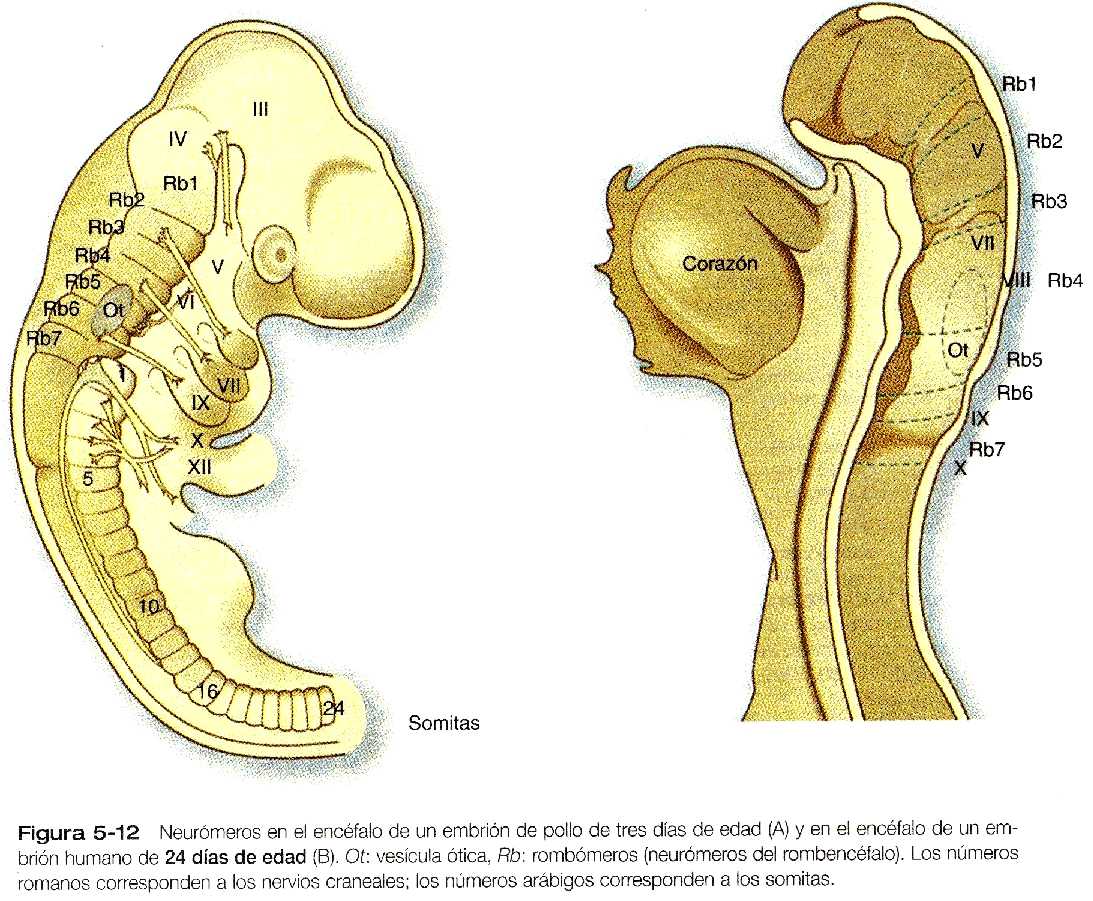
Hay otro nivel de segmentación que se sobrepone a la organización morfológica macroscópica tradicional del encéfalo en desarrollo, y cuya importancia apenas comienza a entenderse. Desde hace mas de 150 años, algunos investigadores han descrito unos segmentos regulares, que solo son visibles en forma transitoria y que se llaman neurómeros, (Figura 5_12) en las regiones del tubo neural de las cuales surge el rombencéfalo, pero solo hace poco tiempo se ha apreciado su importancia. En el embrión humano, los neurómeros (a menudo llamados rombómeros) pueden observarse en el rombencéfalo, desde comienzos de la cuarta semana hasta finales de la quinta (Figura 5_12B). Centros de proliferación celular individual y localizada en el rombencéfalo dan lugar a los rombómeros. Estos se organizan como parejas pares e impares y, una vez que se establecen, actúan como los compartimientos aislados en los embriones de los insectos. A causa de propiedades especificas de la superficie, las células de los rombómeros adyacentes no se entremezclan a través de las fronteras existentes entre los segmentos pares e impares; no obstante, las células marcadas de dos rombómeros pares o de dos impares colocadas unas al lado de otras sí se entremezclan. Durante su breve existencia, los rombómeros sirven de base para la organización fundamental del rombencéfalo. En el adulto, la organización segmentada de los rombómeros se manifiesta en el origen rombomérico específico de muchos de los pares craneales y las partes de la formación reticular en el interior del tronco del encéfalo.
El desarrollo de los rombómeros se asocia con patrones muy regulares de expresión de una amplia variedad de genes, que trabajan en forma armónica todavía hoy poco conocida, para especificar la naturaleza de cada rombómero. Los genes Hox, en especial, parecen ser fundamentales en la determinación de la naturaleza segmentaria de los rombómeros individuales. Otro gen importante en el proceso de segmentación del rombencéfalo es el factor de transcripción zinc finger, el Krox-20, que actúa corriente arriba de los genes Hox.
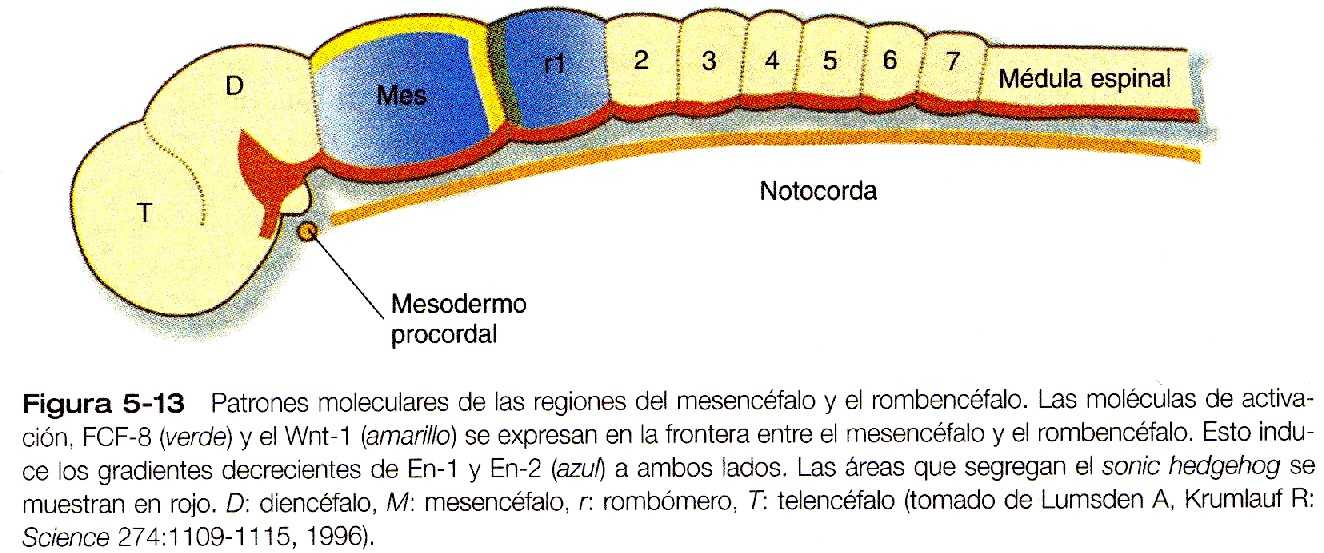
Otra frontera segmentaria en el encéfalo en desarrollo se produce en la unión entre el mesencéfalo y el rombencéfalo (Figura 5_13). Esta frontera es el lugar de la producción de dos poderosas moléculas de señal, la FGF-8 y la Wnt-1 (un gen homólogo del wingless, un gen de polaridad segmentaria de la Drosophila (Figura 5_1). Estas señales regulan un gradiente de expresión de dos genes Engrailed (también genes de polaridad segmentaria), factores de transcripción que son fundamentales en la organización del desarrollo tanto del mesencéfalo como del cerebelo.
La organización fundamental y los controles del desarrollo del prosencéfalo (telencéfalo y diencéfalo) se siguen debatiendo y algunos expertos han concluido que esta región se organiza sobre una base de segmentación transversa, mientras que otros se inclinan hacia una organización más longitudinal. Como ya se dijo, la aparición del prosencéfalo junto con la parte rostral de la cabeza depende de señales moleculares que emanan de la lámina procordal. Por medio del análisis de los patrones de expresión genética, algunos investigadores han dividido el diencéfalo en formación en cuatro neurómeros, pero ha sido más difícil demostrar subdivisiones en el promordio del telencéfalo.
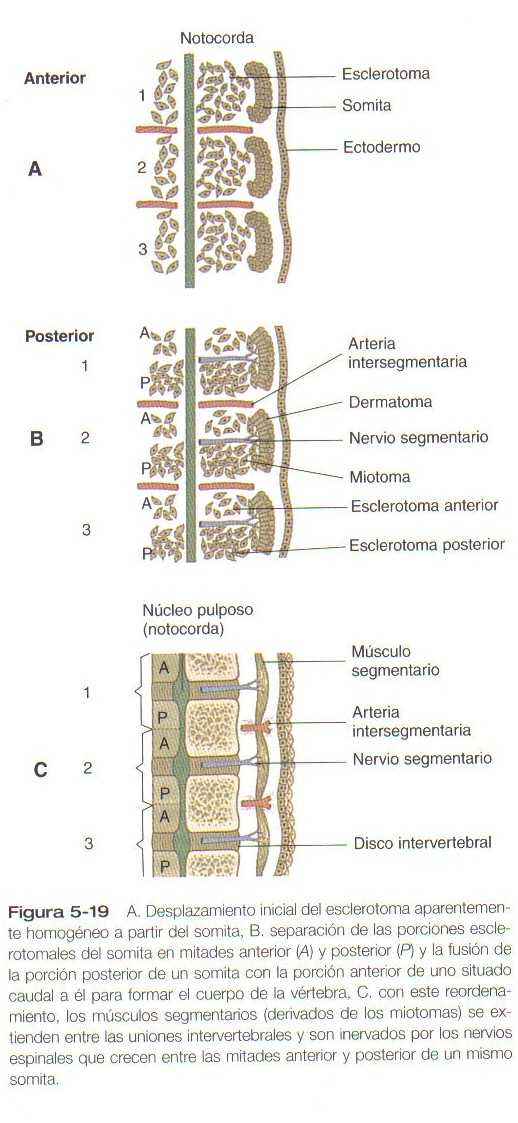
Aunque los neurómeros no se observan en la región del tubo neural que da lugar a la médula espinal, la organización regular de las raíces nerviosas motrices y sensitivas es prueba de una organización segmentaria fundamental también en esta región del cuerpo. En contraste con el encéfalo, parece que la segmentación de la médula espinal es impuesta por señales que proceden del mesodermo paraxial y no de señales moleculares intrínsecas del tubo neural (En la figura 5_19 se ilustra la relacion entre los nervios raquideos y los segmentos mesodérmicos del cuerpo [los somitas y sus derivados]).
Cresta Neural
Cuando el tubo neural ha acabado de cerrarse y se está separando del ectodermo cutáneo, una población de células denominada cresta neural deja la parte dorsal del tubo neural y comienza a extenderse por todo el cuerpo del embrión (Figura 5_11). La cresta neural produce una gran cantidad de estructuras en el embrión, y su importancia es tal que en ocasiones recibe el nombre de cuarta capa germinal del cuerpo.
Plácodas sensoriales e inducciones secundarias en la región craneal
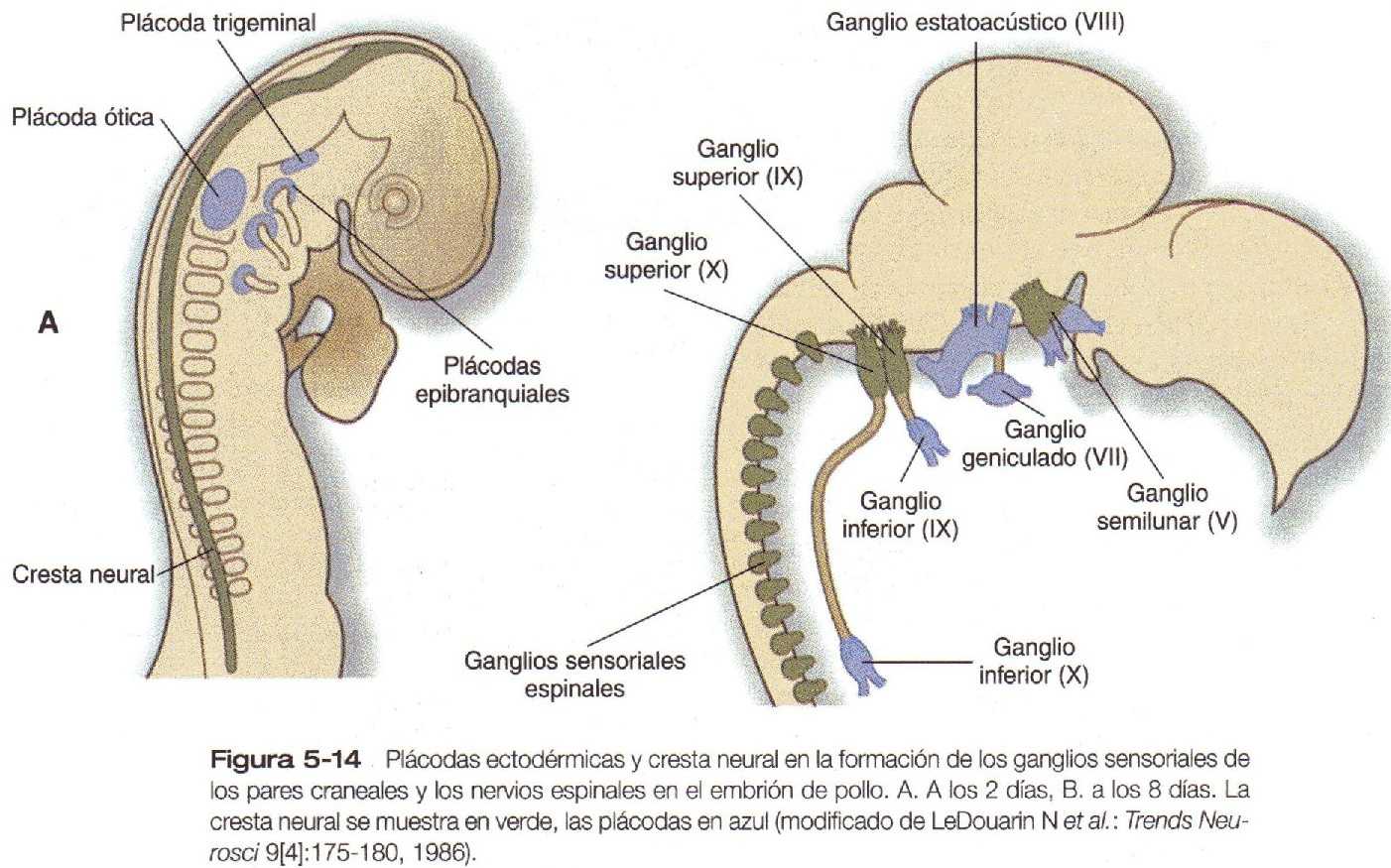
Conforme la región craneal comienza a tomar forma, aparecen varias series de plácodas ectodérmicas (engrosamientos ectodérmicos) a los lados del tubo neural. La mayoría, si no todas, resultan de procesos inductivos secundarios entre otros tejidos (en la mayoría de los casos el tubo neural o la cresta neural) y el ectodermo suprayacente. Entre las primeras plácodas prominentes están las plácodas cristalinas, que terminan formando el cristalino del ojo y las plácodas óticas, que formarán el oído interno. En las regiones más rostrales de la cabeza, las plácodas ectodérmicas dan lugar al epitelio sensorial olfatorio de la nariz, y una proyección similar desde el techo del estomodeo (boca primitiva) da lugar al lóbulo anterior de la hipófisis (adenohipófisis). En la región del rombencéfalo, varios pares de plácodas que se desarrollan en armonía con la cresta neural local participan en la formación de los ganglios sensitivos de los pares craneales (Figura 5_14).
DESARROLLO DE LA CAPA GERMINAL MESODERMICA
Desarrollo básico de la capa mesodérmica
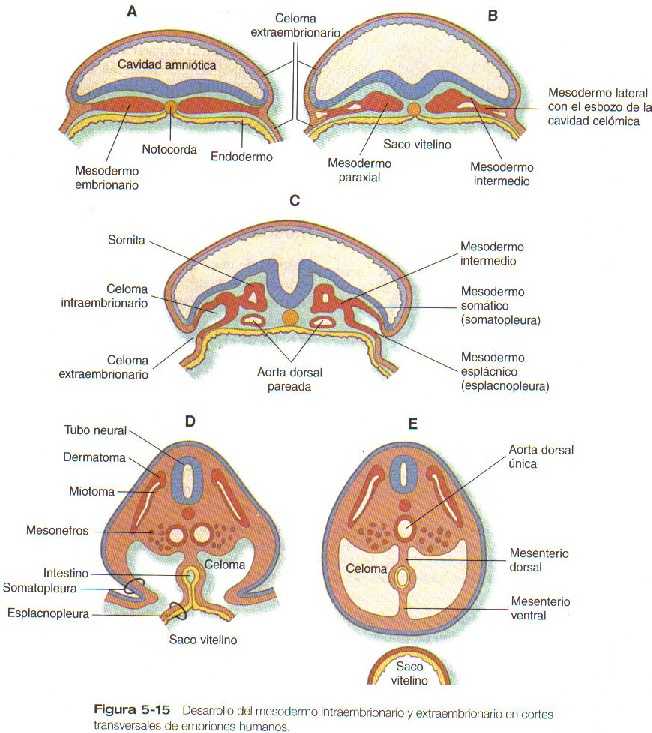
Después de pasar por la línea primitiva, las células del mesodermo se extienden hacia los lados entre el ectodermo y el endodermo como una capa continua de células mesenquimatosas. A continuación en las secciones transversales de los embriones pueden reconocerse tres regiones en el mesodermo (Figura 5_15 B). La más próxima al tubo neural es una columna engrosada de células mesenquimatosas conocida como mesodermo paraxial o lámina segmentaria. Este tejido pronto se organiza en somitas. La región contigua al mesodermo paraxial es una región compacta de mesodermo intermedio o lámina intermedia, que dará lugar al sistema urogenital. Más lateralmente está el mesodermo lateral o lámina lateral que se divide en dos capas y forma el recubrimiento uterino de la pared del cuerpo y los miembros, así como el recubrimiento externo de la pared del tracto digestivo.
Mesodermo paraxial
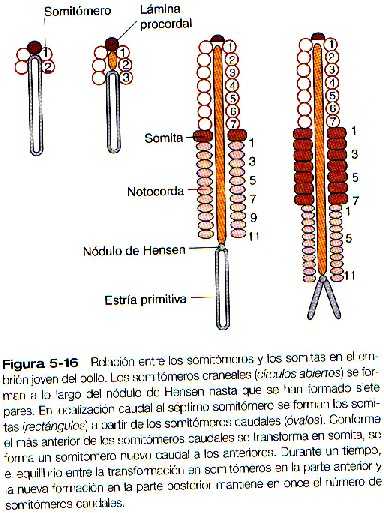
Conforme el nódulo primitivo y la estría primitiva involucionan hacia el extremo caudal del embrión, dejan en dirección craneal a él a la notocorda y a la placa neural. Lateralmente a la placa neural, el mesodermo paraxial parece ser un cordón homogéneo de células mesenquimatosas estrechamente adosadas. No obstante, si se examinan microfotografías electrónicas de barrido de este mesodermo con tecinas estereoscópicas, puede discernirse una serie de segmentaciones regulares y pares de segmentos. Estas segmentaciones, llamadas somitómeros, han sido muy estudiadas en embriones de aves, pero también se encuentran en los mamíferos. Nuevos pares de somitómeros se forman a lo largo del nódulo primitivo a medida que éste regresa hacia al extremo caudal del embrión (Figura 5_16). Solo cuando se han formado unos 20 pares de somitómeros y el nódulo primitivo ha involucionado mucho en dirección caudal, se desarrolla el primer par de somitas (masas de mesodermo paraxial en forma de ladrillos) caudales al séptimo par de somitómeros.
Después de que el primer par de somitas se ha establecido unos 20 días después de la fecundación, se desarrolla una relación regular entre la regresión de la estría primitiva y la formación de somitas y somitómeros adicionales. Los primeros siete pares de somitómeros en la región craneal no se separan ni segmentan más; el primer par de somitas se forma a expensas del octavo par de somitómeros. En los diferentes tipos de embriones estudiados hasta la fecha hay una relación constante entre el par más caudal de somitas definitivos y el número de somitómeros (por lo general de diez a once) que se pueden demostrar por detrás de ellas. En intervalos de unas pocas horas, el par de somitómeros localizado en posición caudal a los últimos somitas formados se transforma en un nuevo par de somitas, y un nuevo par de somitómeros aparece en el extremo caudal del mesodermo paraxial cerca del nódulo primitivo (Figura 5_16). Una vez que la regresión del nódulo primitivo ha terminado, no se forman más somitómeros, pero si siguen formándose mas somitas hasta que desaparece el último de los somitómeros caudales.
A pesar de decenios de investigación, todavía siguen sin comprenderse los mecanismos básicos de la formación inicial de los omitas, aunque parece que la segmentación es una propiedad intrínseca del mesodermo paraxial. Sin embargo, últimamente se ha aprendido mucho sobre la biología de los somitas individuales una vez que están formados. La forma en que los somitas y los somitómeros dejan de formarse se entiende un poco mejor que como se producen, pero una posibilidad es que la muerte genéticamente determina (apoptosis) de las células de la cola desordena el mesodermo paraxial o los últimos somitómeros en formación y actúa como señal de detención para formar nuevos somitas.
Formación de los somitas individuales.
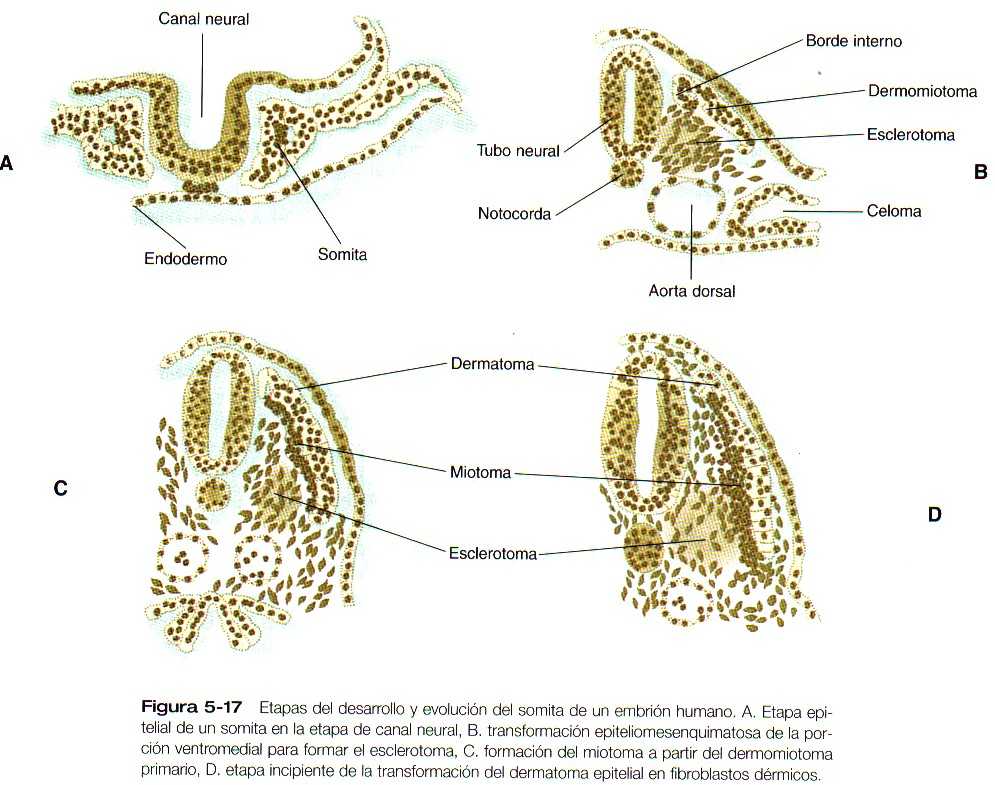
La formulación de un somita individual implica la transformación de bloques segmentarios de células con morfología mesenquimatosa en una esfera de células epiteliales en el mesodermo paraxial (Figura 5_17 A). Aunque la segmentación del mesodermo paraxial parece estar bajo un control intrínseco, la epitelización de los primeros somitas depende de una acción inductora del ectodermo suprayacente todavía desconocida. Esta señal estimula la expresión de un gen llamado paraxis, un factor básico de transcripción hélice-asa-hélice (hélix-loop-hélix). Posiblemente a causa de la mediación del paraxis, la transformación de mesénquima en epitelio va precedida de un aumento en las propiedades de adhesión celular de las células presomíticas. Las células del somita epitelial se organizan de manera que sus superficies apicales rodean una pequeña luz central (que contiene un pequeño número de células centrales indiferenciadas) y sus superficies basales exteriores están rodeadas por una lámina basal (que contiene una laminina, fibronectina, y otros componentes de la matriz extracelular).
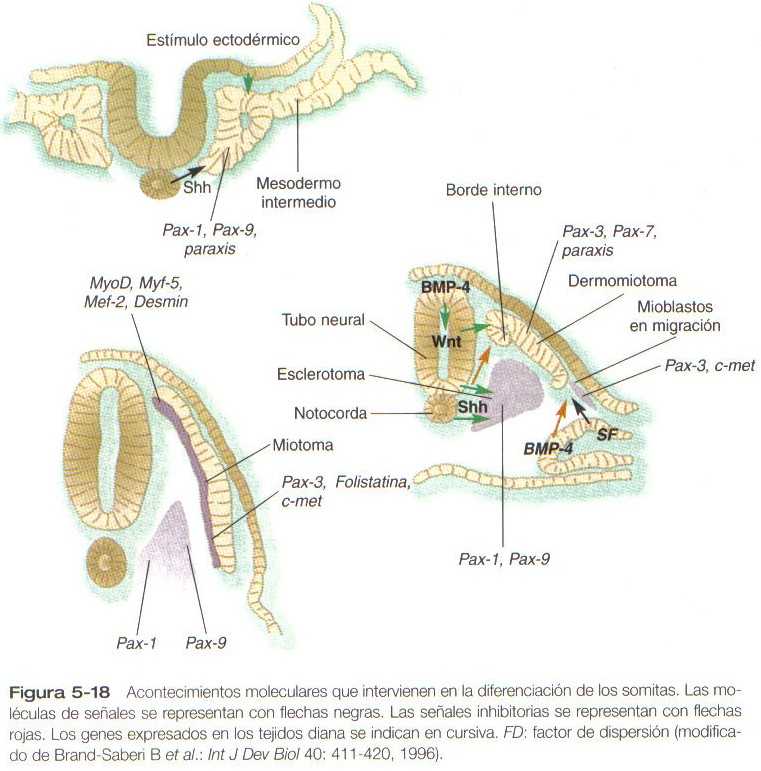
Poco después de la formación del somita epitelial, las células de su pared ventromedial sufren un estimulo inductivo en forma de una molécula de activación, el sonic hedgehog, que se origina en la notocorda y en la pared ventral del tubo neural. La respuesta es la expresión de Pax-1 y Pax-9 en la mitad ventral del somita, que ahora se llama esclerotoma (Figura 5_18). Esto conduce a un aumento de las mitosis, la pérdida de las moléculas de adhesión celular(N-cadherina), la disolución de la lámina basal en esa región y la transformación de células epiteliales (y células centrales) de esa región, nuevamente en células con una morfología mesenquimatosa (estas células se llaman mesénquima secundario). Estas células mesenquimatosas secundarias migran o son desplazadas hacia la línea media a partir del somita (Figura 5_17 B) y comienzan a producir proteoglicanos tipo condroitín sulfato y otras moléculas características de la matriz cartilaginosa a medida que se disponen alrededor de la notocorda.
Bajo la influencia de los productos segregados por los genes Wnt producidos en la zona dorsal del tubo neural, que contrarrestan una influencia inhibitoria del sonic hedgehog, la mitad dorsal del somita epitelial se transforma en el dermomiotoma y expresa sus propios genes característicos (Pax-3, Pax-7, paraxis). Las células mesenquimatosas que surgen del borde dorsal interno del dermomiotoma forman una capa separada, el miotoma, mientras que el resto de las células epiteliales del somita se llaman dermatoma (Figura 5_17 C). Como su nombre lo indica, las células del miotoma producen músculo y las del dermatoma contribuyen a la formación de la dermis.
Organización del somita y del plan básico de la segmentación del cuerpo.
En corte transversal el somita epitelial puede subdividirse en cuatro cuadrantes, cada uno de los cuales da lugar a un grupo diferente de derivados (Tabla 5_4). Como se describió en la sección anterior, bajo la influencia del sonic hedgehog segregado por la notocorda, la mitad ventral del somita se transforma en el esclerotoma y contribuye a formar el cuerpo vertebral. Dos ratones mutantes ilustran la importancia de los componentes que interactúan en la diferenciación del comita. En el mutante Danforth de cola corta, la degeneración de la notocorda (inductora del esclerotoma) va seguida de una deficiente formación de los cuerpos vertebrales y de los discos intervertebrales. Una interrupción de la expresión del gen Pax-1 en el esclerotoma en el mutante ondulado (jorobado) también provoca defectos vertebrales.
Después de la inducción de la mitad dorsal del somita para formar el dermomiotoma inducido por el Wnt, se diferencia el borde interno de las células del dermomiotoma (Figura 5_17B). Estas células del borde interno se extienden por debajo del dermomiotoma para formar el miotoma, una capa diferente que se diferenciara en músculo, (figura 5_17 C). La porción interna del miotoma continua con la expresión de varios factores reguladores miogénicos a medida que las células se preparan para formar los músculos intrínsecos de la espalda (Tabla 5_4). Bajo la influencia de la proteína 4 morfogenética ósea (BMP-4), producida por el mesodermo lateral, se suprime la expresión de los factores reguladores miogénicos en las células del dermomiotoma lateral y estas continúan expresando el Pax-3 y también una molécula receptora, la c-met. Un factor de crecimiento, el factor de dispersión (también llamado factor de crecimiento hepático), segregado en la región de las yemas de los miembros, se fija al receptor c-met de las células del dermomiotoma lateral. Esto estimula a dichas células a migrar fuera del somita y dirigirse hacia la yema de la extremidad a la vez que continúan expresando el Pax-3, su marcador miotómico.
Conforme las células del esclerotoma se sitúan alrededor de la notocorda, las células de la mitad anterior de un somita se fusionan con las células de la mitad posterior del somita situado en posición más craneal. Con posterioridad, de esta fusión se formará parte de una vértebra. Este ordenamiento deja las vértebras óseas descolocadas con respecto a los músculos segmentarios del tronco derivados del miotoma; (Figura 5_19C) esto permite que al contraerse los músculos segmentarios muevan la columna vertebral en dirección lateral. La relación entre la mitad anterior de un somita y la mitad posterior del somita vecino recuerda los parasegmentos de la Drosophila (proceso similar al de las subdivisiones de los segmentos en dos partes), pero no se ha determinado aun si son similares también desde el punto de vista funcional en términos del control genético.
Mesodermo intermedio
En el embrión joven la estructura que conecta el mesodermo paraxial con el mesodermo de la placa lateral es un pequeño cordón de células llamado mesodermo intermedio (masa intermedia) que corre a lo largo de toda la longitud del tronco. Posiblemente debido a la falta de receptores, el mesodermo intermedio no parece estar bajo la influencia del BMP-4 adopta las características de la placa lateral. El mesodermo intermedio es el precursor del sistema urogenital; sus primeros signos de diferenciación se presentan en las regiones más craneales, donde aparecen fugazmente unos vestigios de la forma primordial del riñón, el pronefros. En la región lateral del mesodermo intermedio aparece un conducto pronéfrico a cada lado del embrión. El conducto pronéfrico es importante en la organización del desarrollo de gran parte del sistema urogenital adulto, que se forma sobre todo a partir de células de las porciones caudales del mesodermo intermedio.
Mesodermo de la placa (lámina) lateral.
Poco después de la gastrulación, el ectodermo situado por encima del mesodermo mas lateral produce BMP-4. Muy poco después, el propio mesodermo lateral comienza a producir BMP-4. Los estudios experimentales han demostrado que esta molécula tiene la capacidad de hacer que el mesodermo, ya sea paraxial o lateral parece depender de un equilibrio entre influencias medializadoras que emanan de las estructuras axiales (el tubo neural y la notocorda) e influencias lateralizadoras producidas inicialmente por el ectodermo lateral.
El mesodermo de la placa lateral se divide pronto en dos capas como resultado de la formación y la coalescencia de los espacios celómicos (cavidad corporal) en su interior. La capa dorsal (superficial) , que guarda una estrecha relación con el ectodermo, se llama mesodermo somático, y su combinación con el ectodermo se llama somatopleura (Figura 5_15 D). La capa ventral (profunda), denominada mesodermo esplácnico, se asocia íntimamente con el endodermo, y la combinación del endodermo y el mesodermo esplácnico se llama esplacnopleura. Las capas del mesodermo somático y esplácnico intraembrionario que revisten el saco amniótico y el saco vitelino.
Mientras que las capas del mesodermo somático y esplácnico están formándose, todo el cuerpo del embrión experimenta un proceso de plegamiento lateral cuyo resultado es la transformación hasta desde las tres capas germinales plana un cilindro, con un tubo de endodermo (intestino) en el centro, una capa de revestimiento externo tubular de ectodermo (epidermis), y una capa intermedia de mesodermo. Esta transformación se produce antes de la aparición de los miembros.
Formación del celoma.
Conforme al embrión experimenta el plegamiento lateral, las pequeñas vesículas celómicas que se formaron en el interior del mesodermo lateral se fusionan para formar la cavidad celómica (Figura 5_15). Al comienzo el celoma intraembrionario se continúa con el celoma extraembrionario, pero cuando se termina la fase de plegamiento en cada segmento del embrión, los dos espacio celómicos quedan separados. La última región del embrión que experimenta el plegamiento lateral es el área ocupada por el saco vitelino. En esta área, persisten pequeños canales que conectan los celomas intra y extraembrionario hasta que la pared ventral del cuerpo queda totalmente cerrada.
El embrión cilíndrico, el mesodermo somático constituye las paredes lateral y ventral del cuerpo, y el mesodermo esplácnico forma el mesenterio y la pared del tracto digestivo. El mesodermo somático de la placa lateral también forma el mesénquima de las yemas de los miembros, que comienzan a aparecer en la cuarta semana de la gestación (Figura 9_1).
Mesodermo extraembrionario y tallo de conexión (corporal).
Las delgadas capas del mesodermo extraembrionario que tapizan el amnios y el revestimiento ectodérmico del saco vitelino son continuas con los mesodermos somáticos y esplácnico, respectivamente (Figura 5_15 A y B). El extremo caudal del embrión se conecta con los tejidos trofoblásticos (futura placenta) a través del tallo de conexión. Conforme el embrión crece y se desarrolla un sistema circulatorio que se hace funcional, los vasos sanguíneos del embrión crecen a través del tallo de conexión para abastecer la placenta, y el tallo mismo se puede definir mejor como el futuro cordón umbilical. Al final, el mesodermo extraembrionario que tapiza la superficie interna del citotrofoblasto se convierte en el componente mesenquimatoso de la placenta.
Primeras etapas en la formación del sistema circulatorio.
A medida que el embrión crece durante la tercera semana, alcanza un tamaño que no permite que la simple difusión distribuya oxigeno y nutrientes a todas sus células o elimine con efectividad todos los productos de desecho. El desarrollo inicial del corazón y el sistema circulatorio es una adaptación embrionaria que permite el rápido crecimiento del embrión al proveerlo de un medio eficiente para la distribución de los nutrientes. El sistema circulatorio se enfrenta con el gran desafió de tener que crecer y remodelarse continuamente para seguir el crecimiento general del embrión a la vez que seguir el crecimiento general del embrión a la vez que seguir siendo funcional para cubrir las necesidades de las células embrionarias.
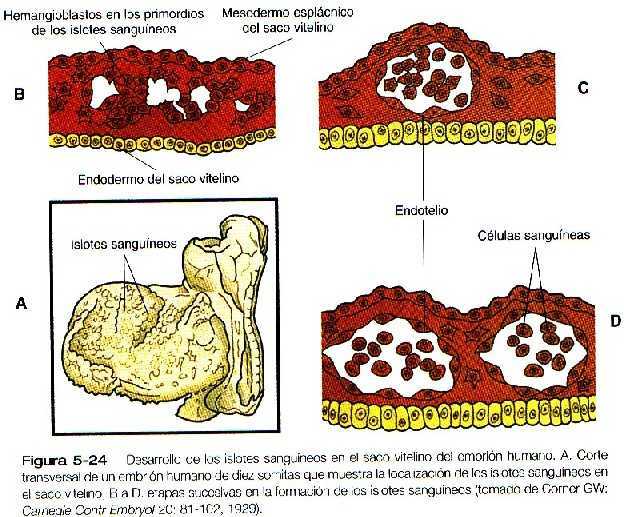
Las primeras fases del desarrollo del sistema circulatorio consisten en la migración del futuro mesodermo del que se va a originar el corazón a través de la estría primitiva, para formar las áreas del mesodermo esplácnico precardiaco. Las células de esta área expresan genes para dos grupos de factores de transcripción (Nkx-5 y la familia MEF2) que son importantes para el desarrollo inicial normal del corazón. Las células forma el corazón pasan por la parte mas caudal de la estría primitiva contribuyen al tracto de entrada. De estas áreas del mesodermo cardiogénico, el corazón y los grandes vasos se forman a partir de tubos pares bilaterales que se fusionan en la línea media por debajo del intestino anterior para producir un tubo único. La sangre tiene un origen diferente, pues las primeras células sanguíneas surgen de islotes sanguíneos en el revestimiento mesodérmico del saco vitelino. La sangre definitiva del adulto surge de células primordiales (stem-cells) que se originan en el mesodermo esplácnico intraembrionario.
Corazón y grandes vasos.
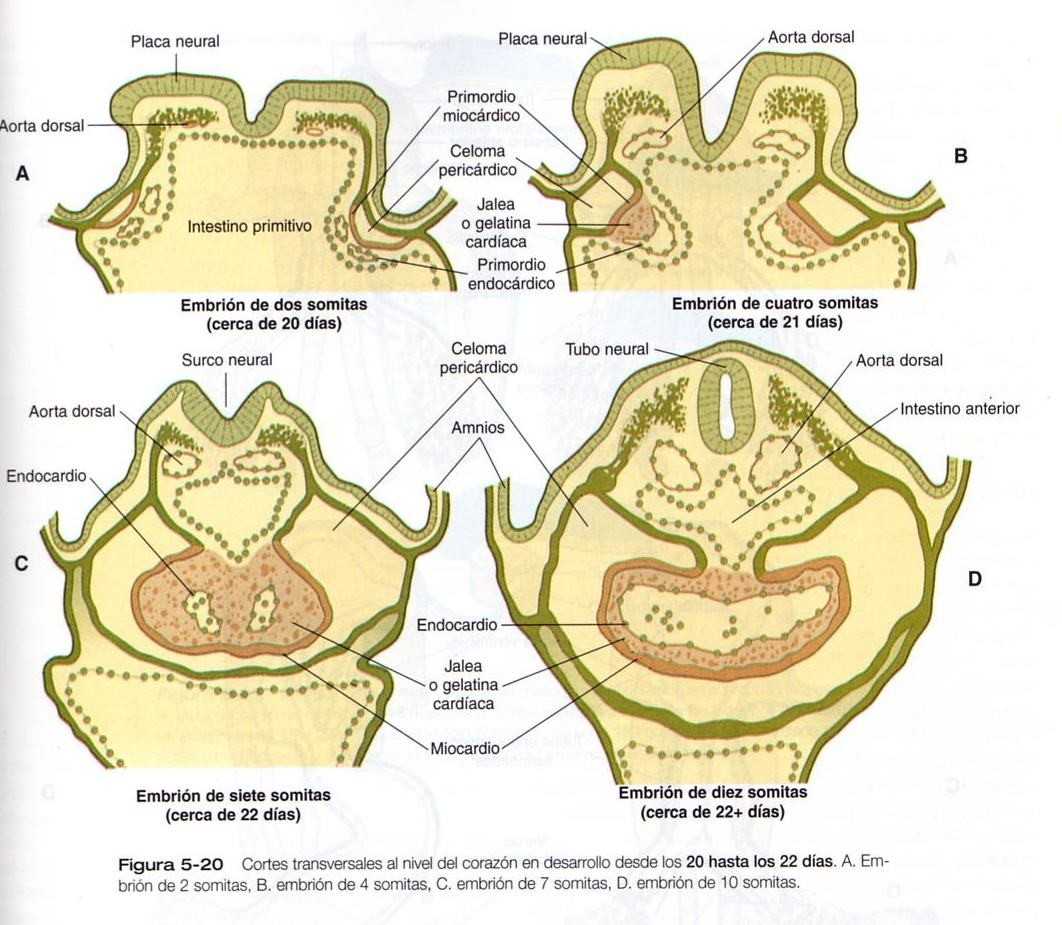
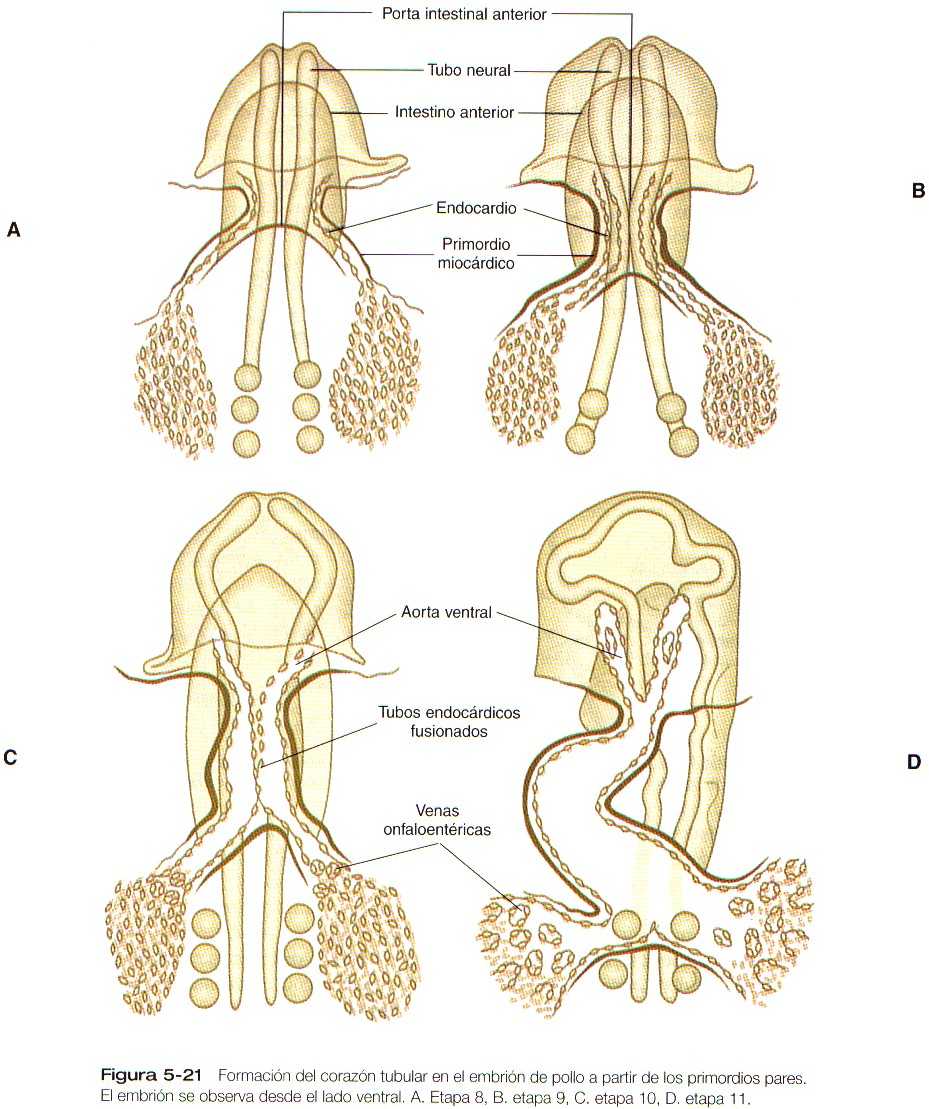
El corazón se deriva del mesodermo esplácnico a partir de un primordio tubular bilateral localizado en posición ventrolateral a la faringe primitiva (Figura 5_20). Los estudios experimentales en vertebrados inferiores han rastreado el mesodermo precardiaco hasta las etapas más precoces del desarrollo, en las que la región formadora del corazón constituye un área en forma de herradura del mesodermo que se extiende hacia atrás a ambos lados del intestino anterior (Figura 5_21). Se ha postulado la existencia de una influencia inductiva proveniente del endodermo vecino para estimular la formación inicial del corazón.
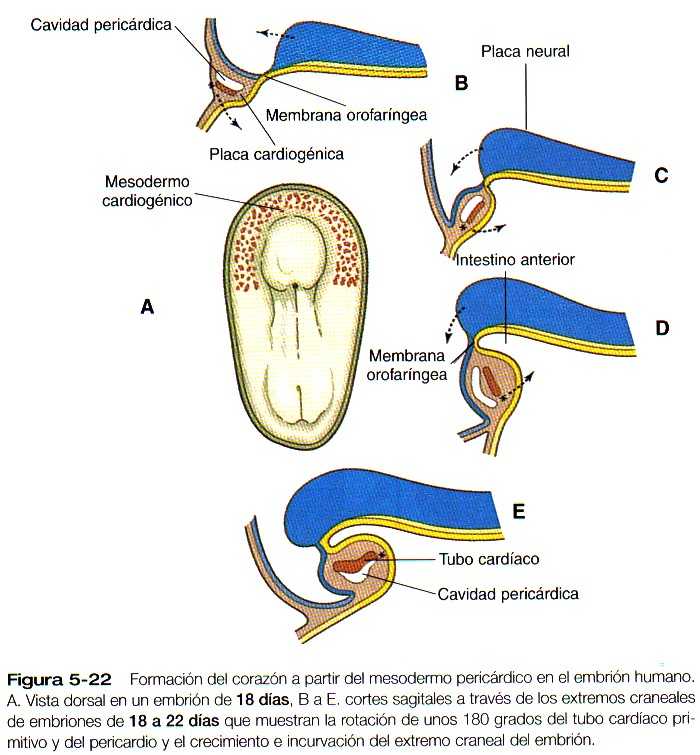
En embriones humanos el mesodermo precardiaco que puede identificarse de forma más precoz es una zona del mesodermo engrosada en forma de medialuna (creciente) y de localización rostral al disco embrionario en gastrulación al comienzo de la tercera semana (Figura 5_22). Conforme el mesodermo intermedio comienza a dividirse en las capas esplácnica y somática, puede reconocerse una placa cardiogénica en el mesodermo esplácnico rostral a la membrana orofaríngea (Figura 5_22B). En esta área el espacio entre las dos capas del mesodermo es el precursor de la cavidad pericárdica. La capa del mesodermo esplácnico de la región pericardiaca se engruesa para convertirse en el primordio miocárdico. Entre esta estructura y el endometrio del intestino primitivo aparecen unas vesículas mesodérmicas aisladas que pronto se fusionan para formar los primordios endocárdicos tubulares (Figura 5_20 A y B). Los primordios endocárdicos acaban de fusionarse y se convierten en la capa de revestimiento interno del corazón.
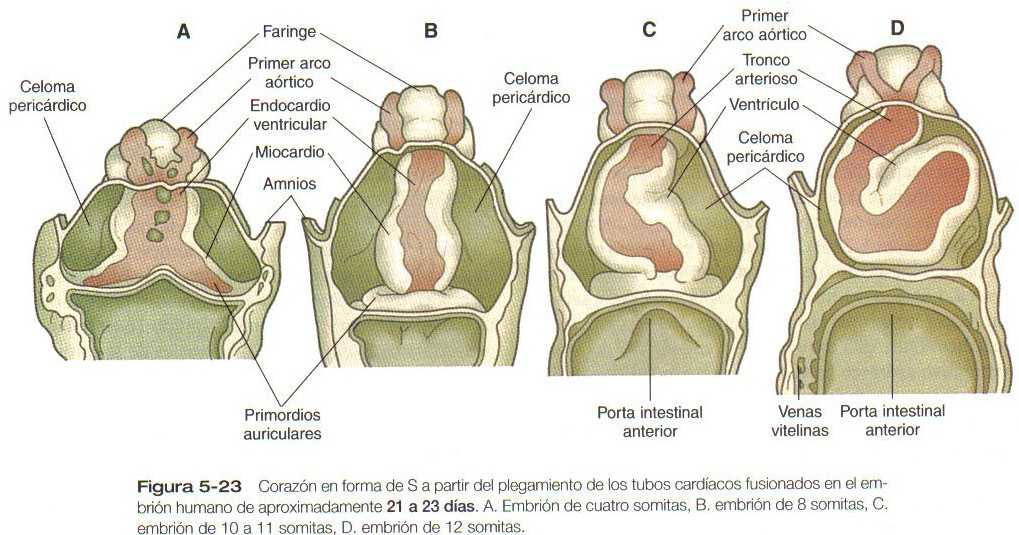
A medida que la cabeza del embrión adquiere su forma en virtud de los plegamientos lateral y ventral, los primordios cardiacos bilaterales se juntan en la línea media por delante del intestino y se fusionan para formar un primitivo corazón tubular único. Esta estructura consta de una capa de revestimiento interna endocárdica rodeada por una laxa capa de matriz extracelular especializada que desde hace tiempo ha recibido el nombre de Jalea (gelatina) cardíaca (Figura 5_20C), por fuera de ella se encuentra el miocardio, que formará la capa muscular del corazón. La capa de revestimiento externo del corazón, llamada epicardio, deriva del primordio proepicárdico que está localizado cerca del mesocardio dorsal. Las células que migran desde proepicardio cubren la superficie del corazón tubular. El corazón tubular está localizado en un espacio conocido como celoma pericárdico. Poco después de que se forma el corazón tubular único, comienza a plegarse y a formar un asa característica en forma de S que indica el inicio de su organogénesis para alcanzar la organización en la configuración del corazón adulto. (Figura 5_23).
El corazón primitivo no se forma aisladamente. En su extremo caudal los tubos endocárdicos no se fusionan sino que se extienden hacia la parte caudal del cuerpo formando el tracto venoso de entrada del corazón (Figura 5_23 A). De forma similar, el tubo endotelial que sale del corazón en su extremo craneal produce arcos vasculares que se incurvan alrededor de la faringe. Las células que migran de la cresta neural forman gran parte de las paredes de estos arcos. Hacia el día 21 o 22 después de la fecundación, la diferenciación de las células musculares cardíacas del miocardio ha avanzado lo suficiente para permitir que el corazón comience a latir.
Sangre y vasos sanguíneos
La formación de la sangre y los vasos sanguíneos comienza en la pared mesodérmica del saco vitelino así como en la pared del corion fuera del cuerpo del embrión. Estimuladas por una interacción inductiva por el endodermo del saco vitelino, muchos islotes de sangre, compuestas por células progenitoras (stem-cells) llamadas hemangioblastos, aparecen en el mesodermo esplácnico extraembrionario del saco vitelino (Figura 5_24). Como resultadote un mecanismo aún no del todo comprendido, las células centrales de los islotes sanguíneos se convierten en células formadoras de sangre (hemocitoblastos), en tanto que las del exterior adquieren las características de las células de revestimiento endotelial, que forman las paredes internas de los vasos sanguíneos. Conforme los islotes sanguíneos de la pared del saco vitelino se fusionan, se forman canales vasculares primarios que se extienden hacia el cuerpo del embrión. Se establecen conexiones con los tubos endoteliales procedentes del corazón embrionario y los grandes vasos, y así empieza a adquirir morfología el sistema circulatorio.
DESARROLLO DE LA CAPA GERMINAL ENDODÉRMICA
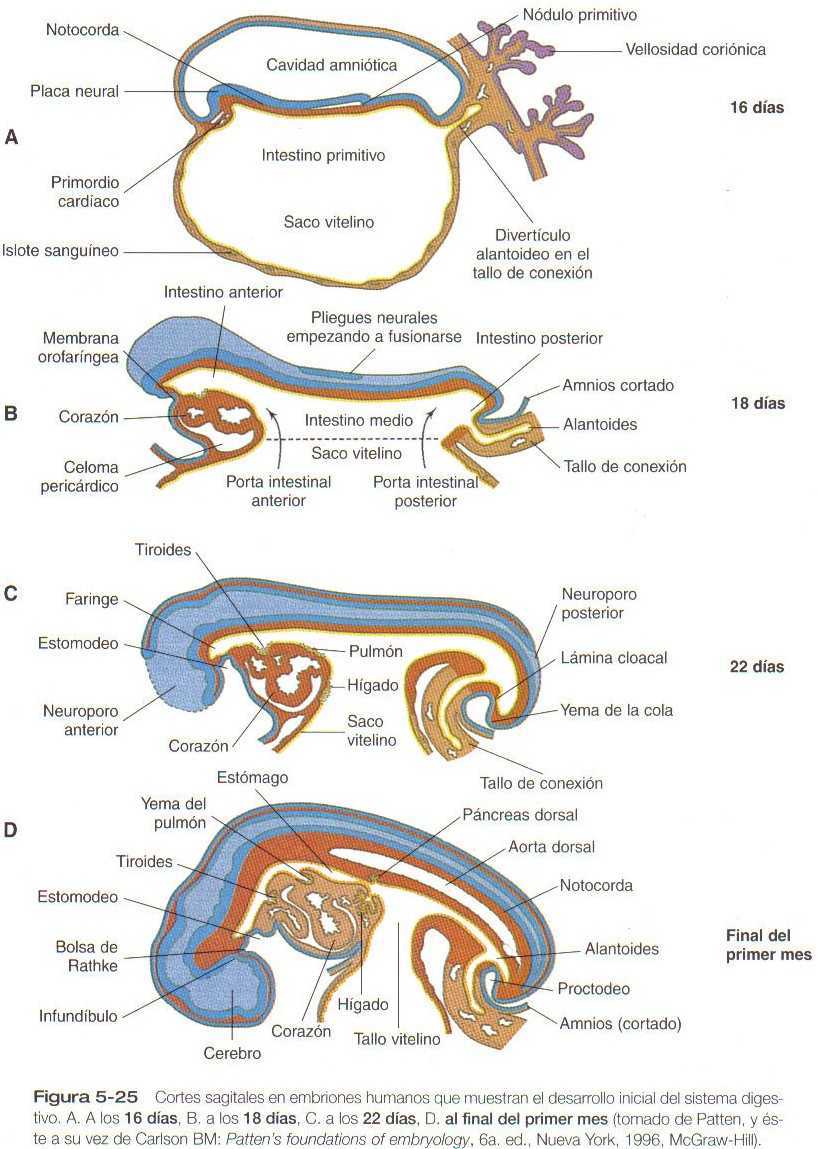
El desarrollo de la capa germinal endodérmica se continúa con la transformación de la capa endodérmica intraembrionaria plana en un tubo intestinal, como resultado del plegamiento lateral del cuerpo del embrión y la flexión ventral de los extremos craneal y caudal del embrión para formar una estructura parecida a una C (Figuras 5_15 y 5_25). Una de las importantes consecuencias morfológicas de este proceso de plegamiento es la diferenciación clara entre el saco vitelino y el tubo digestivo.
Al comienzo de la tercera semana, cuando las tres capas germinales del embrión están comenzando a formarse, el endodermo intraembrionario constituye el techo de un saco vitelino casi esférico (Figura 5_25). El crecimiento de ambos extremos de la placa neural, en especial el gran crecimiento de la futura región del encéfalo, da como resultado la formación del pliegue cefálico y del pliegue de la cola a lo largo del plano sagital del embrión. Este proceso, junto con el plegamiento lateral concomitante, da como resultado la formación y delimitación del intestino anterior y del intestino posterior. Este proceso también comienza a aislar el saco vitelino del intestino propiamente dicho. La secuencia de hechos en la formación del intestino tubular podría asimilarse a una bolsa que constriñese en la región ventral del embrión, aunque su mecanismo real se debe más al crecimiento global del embrión que a una constricción verdadera. La región de la bolsa imaginaria se convierte en el tallo de conexión, con el intestino embrionario arriba y el saco vitelino debajo. (Figuras 5_15D y 5_25D). La porción del intestino que todavía se comunica con el saco vitelino se denomina intestino medio, y los puntos de transición entre el intestino medio, cuya zona ventral está abierta, y las regiones intestinales anterior y posterior del intestino se llaman porta intestinal anterior y posterior (Figura 5_25B).
Los límites endodérmicos de la porta intestinal anterior y posterior también son lugares de la expresión de la molécula de señales sonic hedgehog. En la porta intestinal posterior la aparición del sonic hedgehog en el endodermo va seguida en corto tiempo por la expresión de otra molécula de señales, la BMP-4 (Tabla 5_2). Esto a su vez va seguido por la parición del gradiente de expresión mesodérmica de los grupos parálogos 9-13 de los genes Hox (Figura 5_5) donde aparece una ilustración de los grupos parálogos), de los caudales del Hox-d-9 se expresa en localización más craneal y el Hox-d-13 se expresa en localización más caudal, cerca de la cloaca. Esta distribución de la expresión de los genes Hox asociada con la formación del intestino posterior recuerda la que ya se describió para la formación precoz de la región del prosencéfalo. Es probable que este gradiente sea fundamental para guiar la diferenciación regional del intestino porque más adelante en el desarrollo el mesodermo asociado al intestino es capaz de inducir al endodermo indiferenciado a formar estructuras específicas relacionadas con el nivel craneocaudal del mesodermo.
La extremidad anterior del intestino anterior permanece transitoriamente sellada por una doble capa de ectodermo y endodermo adosada llamada membrana orofaríngea (Figura 5_25B). Esta membrana separa la futura boca (estomodeo), que está tapizada por ectodermo, de la faringe, la parte del intestino anterior que está tapizada con endodermo. Sin la presencia de una capa intermedia de mesodermo, esta capa doble de dos hojas epiteliales es muy débil y rápidamente se rompe. El rápido crecimiento de la región cefálica, junto con la constricción de la región ventral, tiene un gran efecto topográfico sobre la región cardiaca en rápido crecimiento. En el embrión joven los primordios cardíacos están localizados por encima del intestino primitivo. Sin embargo, las fuerzas que dan lugar al intestino tubular hacen que los primordios cardíacos se plieguen 180 grados en dirección creaneocaudal al tiempo que los tubos cardíacos pares se desplazan uno hacia el otro y hacia la línea media ventral (Figura 5_22).
En la región del intestino posterior el crecimiento del cuerpo del embrión no es tan grande como en el extremo craneal; sin embargo, también se produce un plegamiento ventral aunque menos exagerado. Mientras se inicia la yema de la cola, una evaginación procedente del intestino posterior se extiende hacia el interior del mesodermo del tallo de conexión. Esta evaginación se llama alantoides (Figura 5_25B). En la mayoría de los mamíferos y las aves la alantoides representa una gran adaptación estructural para el intercambio de gases y la expulsión de desechos urinarios. Sin embargo, a causa de la efectividad de la placenta, la alantoides nunca se convierte en una estructura importante en el embrión humano. No obstante, debido a los vasos sanguíneos que se asocian con ella, sigue siendo una parte vital del vínculo entre el embrión y la madre.
Por debajo de la alantoides se encuentra otra capa doble de ectodermo y endodermo adosados llamada lámina cloacal o membrana proctodea (Figura 5_25C). Esta membrana, que posteriormente se rompe., cierra la cloaca, que en el embrión joven represente la salida común y única para los sistemas digestivo y urogenital. La depresión situada en la superficie por fuera de la membrana proctodea se llama proctodeo.
A medida que el intestino se hace cada vez más tubular, se producen una serie de interacciones inductivas locales entre el epitelio del tracto digestivo y el mesénquima circundante que dan lugar a la formación de la mayoría de las principales glándulas digestivas y endocrinas (por ejemplo, las salivales, la tiroides, el páncreas), el sistema respiratorio y el hígado. En la región del estomodeo, una inducción entre el prosencéfalo y el ectodermo del estomodeo, provoca la formación de la hipófisis anterior (adenohipófisis).
ESTRUCTURA BASICA DEL EMBRION DE CUATRO SEMANAS
Apariencia macroscópica
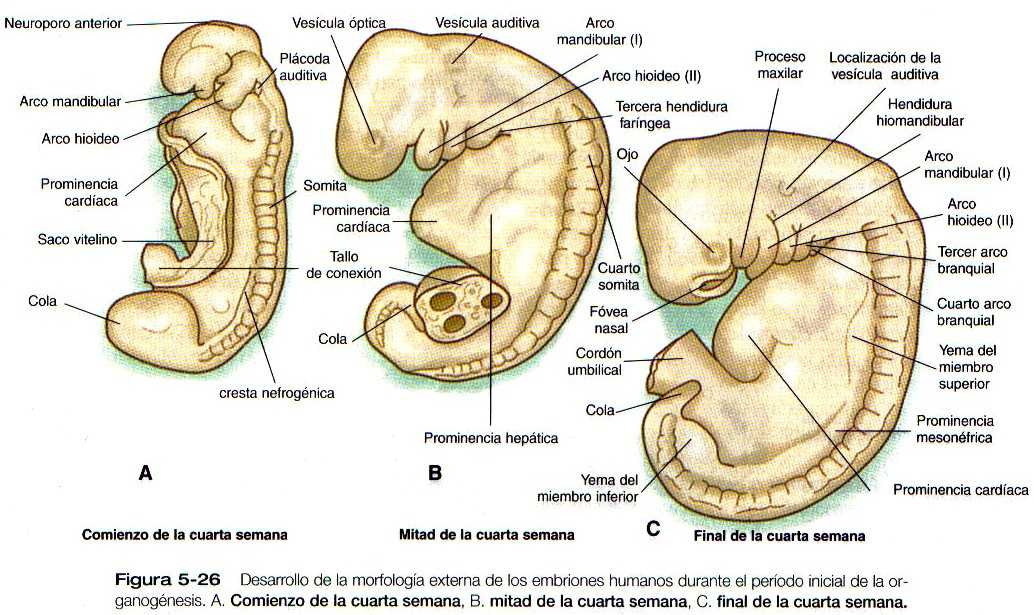
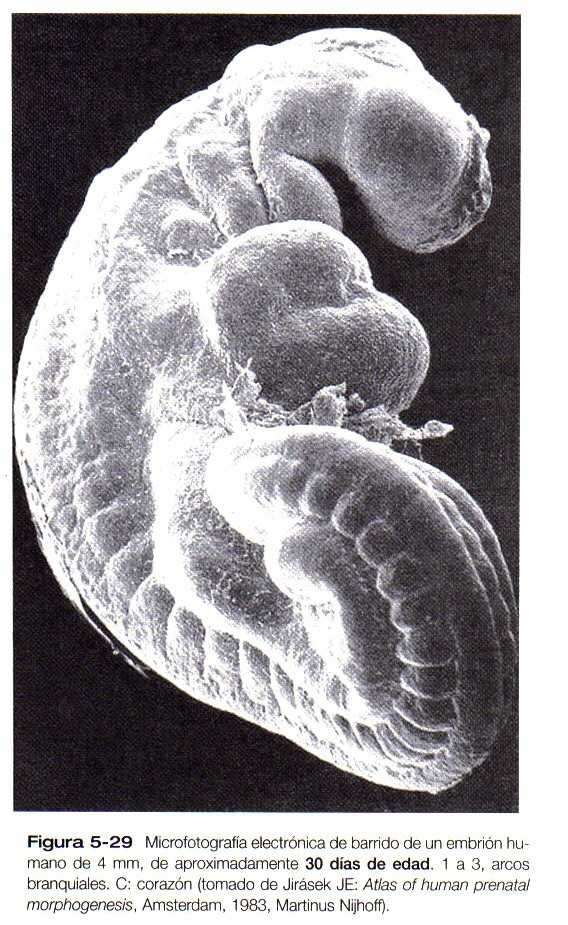
Hacia el fin de la cuarta semana del embarazo el embrión, que en este momento sólo mide 4mm de longitud, ha establecido los rudimentos de la mayor parte de los órganos a excepción de los miembros (que aún están ausentes) y el sistema urogenital (de lo que sólo se han desarrollado las primeras trazas de los riñones embrionarios). Visto desde afuera, el embrión tiene forma de C, y muestra una hilera prominente de somitas situados a lo largo y a ambos lados del tubo neural (Figura 5_26 y 5_27). Con excepción de los rudimentos de los ojos y los oídos y la membrana orofaríngea, que está comenzando a romperse (Figura 5_26B** y C y 5_29), la cabeza carece relativamente de rasgos. En la región cervical son prominentes los arcos branquiales. El tallo de conexión todavía ocupa una parte importante de la pared ventral del cuerpo y, por encima del tallo de conexión, el corazón y el hígado ocasionan abultamientos prominentes en el contorno de la pared ventral del cuerpo. Por detrás del tallo de conexión el cuerpo se une a la cola de forma algo espiral, que es evidente en los embriones de esta edad.
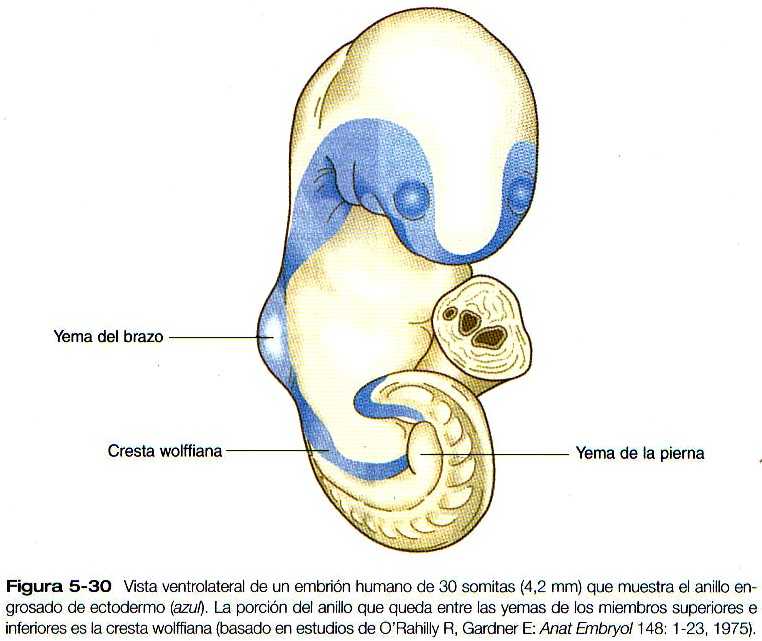
Otro aspecto importante pero poco entendido de los embriones de esta edad es la presencia de un anillo de ectodermo engrosado llamada cresta wolffiiana, que rodea lateralmente el cuerpo embrionario (Figura 5_30). Su función aún no se conoce del todo, pero en ella se forman los primordios de muchas estructures (por ejemplo, la nariz, el ojo, el oído interno, los arcos branquiales y los miembros) que requieren interacciones tisulares para su desarrollo inicial. Todavía no se ha determinado el papel que el ectodermo engrosado pueda jugar en la organogénesis inicial.
Sistema Circulatorio
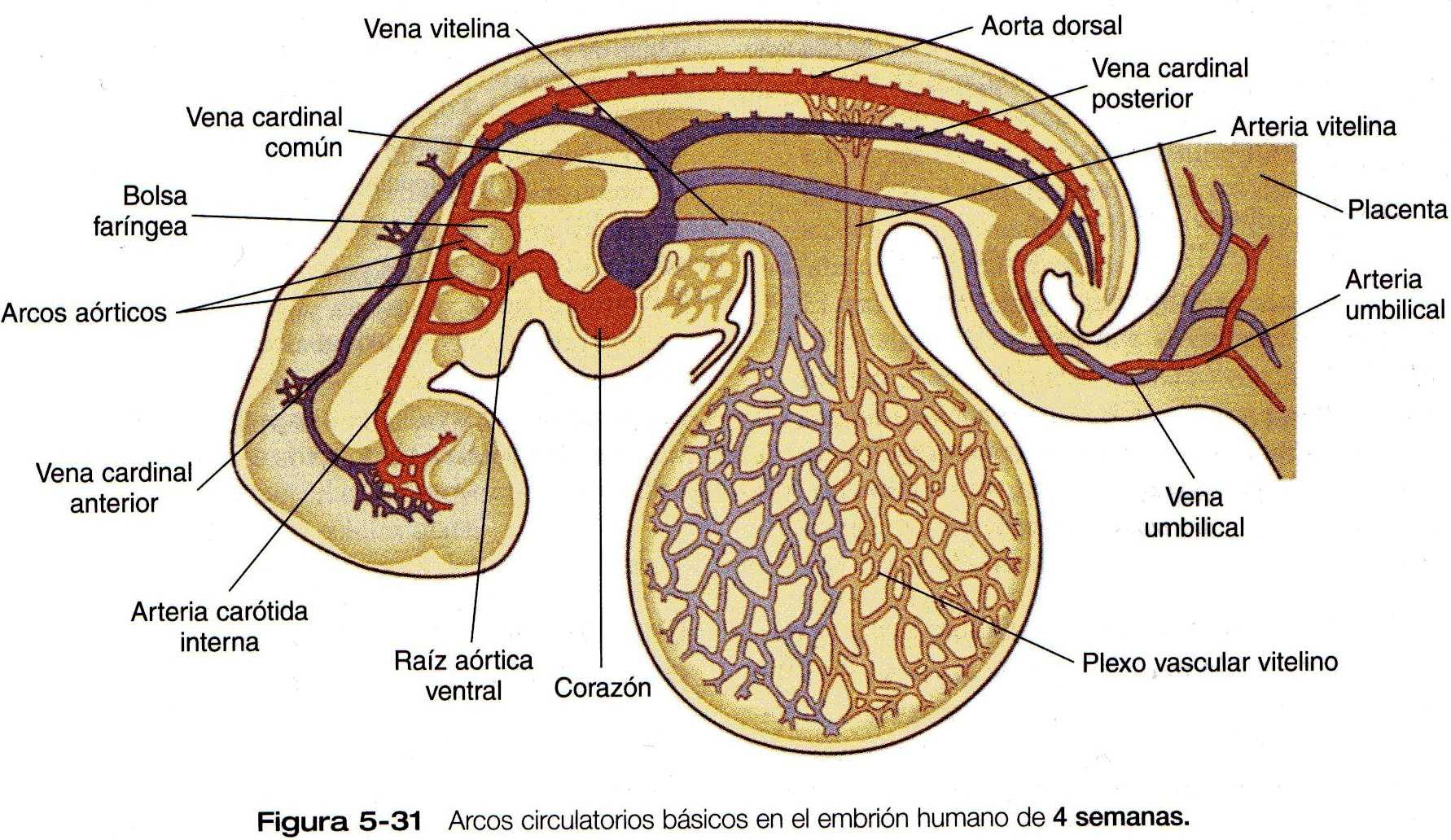
A las cuatro semanas de edad, el embrión tiene un corazón funcional de dos cámaras y un sistema vascular sanguíneo que consta de tres arcos circulatorios separados (Figura 5_31). El primero, el arco circulatorio intraembrionario, se organiza de forma similar a como lo hace el de los peces. Un tracto aórtico ventral de salida desde el corazón que se divide en una serie de arcos aórticos que pasan rodeando la faringe a través de los arcos branquiales y luego se reúnen en una aorta dorsal pareada en la parte dorsal que distribuye la sangre a todo el cuerpo. Un sistema de venas cardinales recoge la sangre y la regresa al corazón a través de un tracto común de entrada.
El segundo arco, que recibe el nombre de arco vitelino u onfaloentérico, es principalmente un asa circulatoria extraembrionaria que irriga el saco vitelino (Figura 5_31). El tercer arco circulatorio, también extraembrionario, consta de los vasos asociados con la alantoides. En los seres humanos, este tercer arco consta de los vasos umbilicales que transcurren a lo largo del tallo de conexión y se distribuyen en una complicada red en la placenta y en los tejidos coriónicos. Este grupo de bases represente la verdadera línea de conexión entre el embrión y la madre. Aunque las dos asas circulatorias extraembrionarias no persisten como tales después del nacimiento, las porciones intraembrionarias de estos arcos se mantendrán como vasos o ligamentos en el individuo adulto.
Derivados de las capas germinales del embrión
Hacia el final de la cuarta semana del desarrollo, ya se han establecido los primordios de casi todas las estructuras y los principales órganos del cuerpo muchos de ellos como resultado.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}