ORGANOS DE LOS SENTIDOS
Primeros acontecimientos en el desarrollo del ojo
Formación del cristalino
Formación de la cornea
Retina y otros derivados de la copa óptica
Retina neural
Iris y cuerpo ciliar
Cuerpo vítreo y sistema de la arteria hialoidea
Capas coroidea y esclerótica
Párpados y glándulas lacrimales
Desarrollo del oído interno
Desarrollo del oído medio
Desarrollo del oído externo
CASO CLÍNICO
PREGUNTAS DE REPASO
Los órganos de los sentidos surgen en gran parte de los engrosamientos celulares que constituyen las plácodas ectodérmicas. Estas se forman en su mayor parte como respuesta a inducciones secundarias del sistema nervioso central y pueden subdividirse en 2 grupos. El primero da lugar a una variedad de estructuras de órganos de los sentidos como el oído interno, el cristalino y el epitelio sensorial olfatorio. El segundo, que está en estrecha asociación con los arcos bronquiales, da lugar a neuronas sensoriales que se combinan con neuronas derivadas de la cresta neural para formar los ganglios sensitivos de los pares craneales.
Este capitulo se centra en el desarrollo del ojo y el oído, los órganos de los sentidos más complejos e importantes de los seres humanos. Los órganos del gusto y del olfato se estudian en el capítulo 13, ya que su desarrollo está íntimamente asociado con el de la cara y la faringe. Los componentes de los pares craneales se analizan en el capítulo 10.
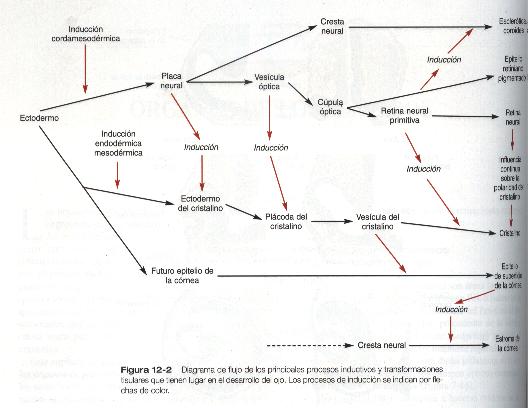
El ojo es una estructura muy compleja cuyos componentes son derivados de varias fuentes, entre los que se incluyen la pared del diencéfalo, el ectodermo superficial suprayacente, y el mesénquima migratorio de la cresta neural craneal. Durante el desarrollo ocular se producen dos acontecimientos básicos: uno es una serie continua de señales inductivas que dan como resultado el esbozo inicial de los componentes principales del ojo; el otro, la diferenciación coordinada de muchos de estos componentes.
Para que se produzca la visión normal, muchas de las complejas estructuras intraoculares deben relacionarse de forma apropiada con otras estructuras vecinas. Por ejemplo, la córnea y el cristalino deben hacerse transparentes y alinearse de forma apropiada para crear una vía adecuada que permita que la luz alcance la retina. La retina a su vez debe estar configurada ordenadamente para que pueda recibir las imágenes visuales y transmitir los patrones de las mismas a los lugares apropiados del encéfalo a través de prolongaciones neurales que se extienden desde la retina hasta el nervio óptico.
Primeros acontecimientos en el desarrollo del ojo
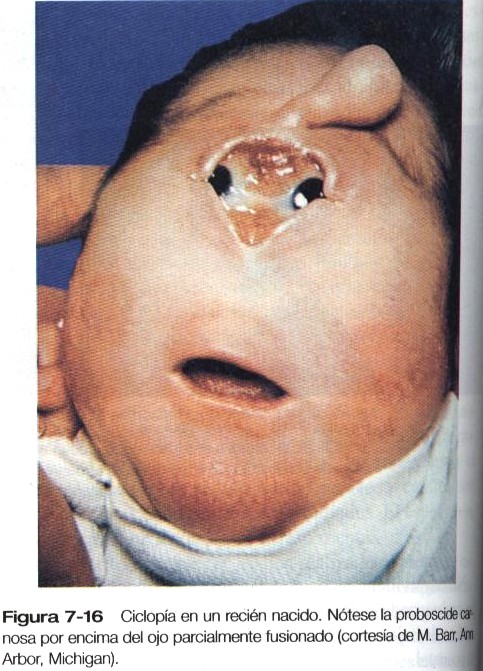
Los esbozos de los campos oculares son áreas localizadas a ambos lados del diencéfalo que expresan Pax-6 (véase en la próxima sección la explicación detallada del Pax-6 en el desarrollo del ojo). Una inducción procedente de la lámina procordal (probablemente el sonic hedgehog) reprime el Pax-6 en la línea media ventral y mantiene separados los dos campos ópticos; si no se presenta dicha influencia, el tejido prosencefálico es deficiente, los campos ópticos convergen y el resultado es una ciclopía (figura 7-16).
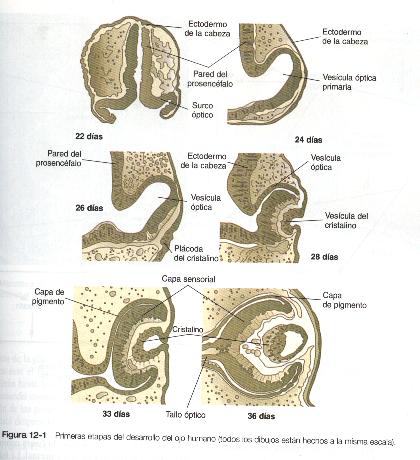
El desarrollo del ojo comienza a hacerse evidente hacia los 22 días de gestación, cuando las paredes laterales del diencéfalo empiezan a evaginarse y forman los surcos ópticos (figura 12-1). Al cabo de unos pocos días estos surcos se agrandan y forman las vesículas ópticas, que están muy cerca del ectodermo superficial suprayacente. La aposición de la pared externa de la vesícula óptica con el ectodermo superficial es esencial para la transmisión de un importante mensaje inductivo que estimula a las células del ectodermo superficial a engrosarse y formar el cristalino (figura 12-2).
La interacción entre la vesícula óptica y el ectodermo que la cubre fue uno de los primeros procesos inductivos que se descubrieron. Al comienzo se caracterizó gracias a los experimentos de extirpación y trasplante llevados a cabo en embriones de anfibios. Cuando las vesículas ópticas se extirpan precozmente, el ectodermo superficial se diferencia en células ectodérmicas ordinarias y no se forman las fibras del cristalino. A la inversa, cuando se combinan las vesículas ópticas con ciertos tipos de ectodermos diferentes de el del ojo, este ectodermo es inducido a formar fibras del cristalino. La investigación posterior en embriones de anfibios ha demostrado que una serie previa de inducciones procedentes de la placa neural y del mesodermo subyacente condicionan al ectodermo para lograr su inducción final en cristalino por parte de la vesícula óptica. En los mamíferos el impedir la aposición de las vesículas ópticas y el ectodermo suprayacente, que entorpece la inducción del cristalino y es un mecanismo importante en la génesis de la microftalmía grave (ojos pequeños) o la anoftalmía (ausencia de ojos) que se observan en los mutantes de small eye y fidget.
El par gen box Pax-6 desempeña una importante función durante todo el desarrollo inicial del ojo y en varias fases del desarrollo de la retina y el cristalino. El Pax-6 se expresa inicialmente en el cristalino, en las plácodas nasales y en gran parte del diencéfalo. En la Drosophila, al Pax-6 se le ha denominado el gen maestro del desarrollo del ojo; es decir, que puede activar la cascada de unos 2.500 genes que guían el desarrollo de este órgano. La importancia del Pax-6 se demuestra en la formación de ojos ectópicos en las antenas y en las patas de la Drosophila cuando el gen no se expresa de modo apropiado. En ausencia de expresión de Pax-6 (mutante eyeless) los ojos no se forman. En el mutante small eye, el equivalente del eyeless en los mamíferos, la vesícula óptica inicial se forma pero, como ya se ha dicho, el desarrollo del ojo no avanza porque el ectodermo superficial no puede responder a la señal inductiva emitida por la vesícula óptica. La reciente identificación en los seres humanos de dos genes (Eya (ojos ausentes) y Six (sine oculis) que son activados por el Pax-6 en la Drosophila sugiere que a pesar de las grandes diferencias en la estructura y en el desarrollo del ojo de los vertebrados y de los insectos, el aparato genético básico se ha conservado durante la filogenia. En los ratones, el Eya – 1 y el Eya – 2 se expresan en las plácodas del cristalino y al parecer son necesarios para la inducción de las plácodas y su diferenciación precoz, pero en ausencia de la función del Pax-6, no se expresan y el ojo no se desarrolla.
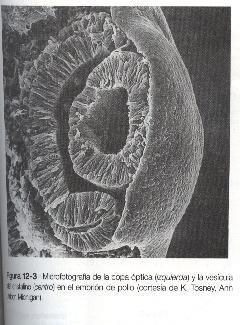
Conforme avanza el proceso de inducción del cristalino, la cara externa de la vesícula óptica comienza a aplanarse y termina por adoptar una forma cóncava. El resultado es la transformación de la vesícula óptica en copa óptica (figura 12-1). Entre tanto, el ectodermo inducido del cristalino se engruesa e invagina para formar la vesícula del cristalino, que se desprende del epitelio superficial del cual se ha originado (figuras 12-1 y 12-3). Entonces la vesícula del cristalino se convierte en el agente primario de una nueva inducción mediante la acción que ejerce sobre el ectodermo de superficie que lo cubre y hace que allí comience el desarrollo de la córnea (figura 12-2).
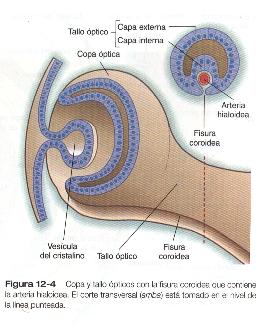
La formación de la cúpula óptica es un proceso asimétrico que tiene lugar en el borde ventral de la vesícula óptica y no en su centro. El resultado es la formación de una hendidura llamada fisura coroidea, que se continúa con el surco del tallo óptico (figura 12-4). Durante gran parte del desarrollo ocular inicial, la fisura coroidea y el surco óptico forman un canal por el cual pasa la arteria hialoidea hacia la cámara posterior del ojo. Al comienzo, el tallo óptico representa un cuello estrecho que conecta la cúpula óptica con el diencéfalo, pero conforme el desarrollo avanza, es invadido por las prolongaciones neuronales que proceden de las células ganglionares de la retina. Después de que estas prolongaciones han encontrado su camino hasta las regiones adecuadas del cerebro, el tallo óptico pasa a llamarse nervio óptico.
Más adelante en el desarrollo, la fisura coroidea se cierra y no queda nada de ella en el iris normal. La falta de de cierre de la fisura coroidea da como resultado la anomalía denominada coloboma (Pág. 282). Otro par gen box, el Pax-2, se expresa en las células no neuronales del tallo óptico y en el nervio óptico inicial mientras que está siendo invadido por los axones. Las células que expresan Pax-2 proporcionan claves de dirección a los axones retinianos en crecimiento que entran en el tracto óptico contralateral por el nervio óptico y el quiasma óptico. El coloboma y los trastornos visuales se observan con frecuencia en individuos portadores de genes Pax-2 mutantes. En los ratones mutantes Pax-2, los axones retinianos no cruzan la línea media a través del quiasma óptico, sino que permanecen en el tracto óptico ipsilateral.
Formación del cristalino
Desde las primeras etapas, la formación del cristalino depende de las instrucciones impartidas por el Pax-6. El ectodermo superficial de la cabeza requiere la acción del Pax-6 para la inducción de la plácoda del cristalino, aunque de ninguna manera es este el uncí gen implicado. La influencia del Pax-6 continúa hasta las etapas avanzadas del desarrollo del cristalino, cuando controla la actividad de los genes para la producción de las proteínas cristalinas.
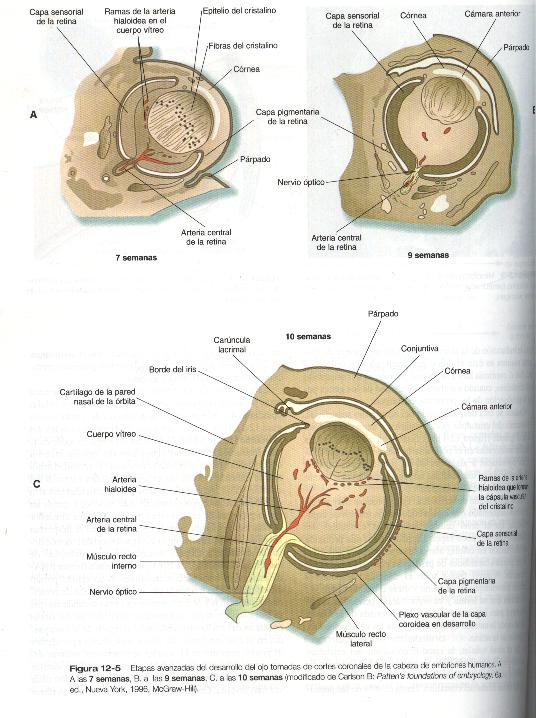
Cuando se esta desprendiendo del ectodermo superficial, la vesícula del cristalino es casi esférica y tiene una cavidad central grande (figura 12-1). Al final de la sexta semana las células localizadas en el polo interno de la vesícula del cristalino comienzan a alargarse en un primer paso hacia su transformación en unas células largas y transparentes llamadas fibras del cristalino (figura 12-5).
La diferenciación del cristalino es un proceso muy preciso y bien orquestado que comprende varios niveles de organización. En el nivel celular, las células epiteliales del cristalino que son poco especializadas sufren una profunda transformación en células alargadas transparentes que contienen grandes cantidades de proteínas cristalinas especializadas. A nivel tisular todo el cristalino responde a señales provenientes de la retina y otras estructuras del ojo, de manera que su forma y su organización general se adaptan mejor para la transmisión no distorsionada de los rayos de luz desde la entrada en la córnea hasta las células fotorreceptoras de la retina.
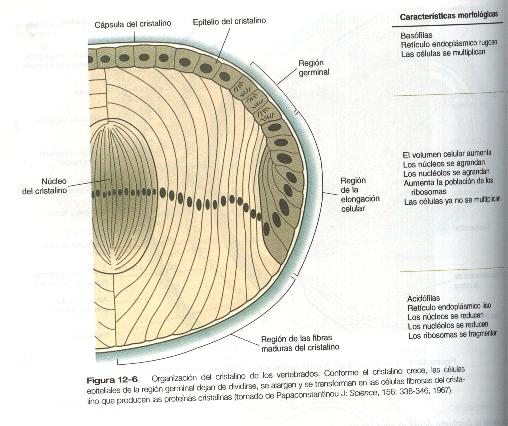
En el nivel celular, la citodiferenciación del cristalino consiste en la transformación de células epiteliales cristalinas con actividad mitótica en las células posmitóticas alargadas de las fibras del cristalino. Hasta el 90% de las proteínas solubles de estas células posmitóticas son cristalinas .El cristalino de los mamíferos contiene tres proteínas cristalinas principales: alfa, beta y gamma.
La formación de las fibras del cristalino que contienen estas proteínas comienza con el alargamiento de las células epiteliales del polo posterior de la vesícula del cristalino (figura 12-1). Estas células producen las fibras del núcleo del cristalino (figura 12-6). El resto de las fibras procede de la transformación de células cuboidales del epitelio anterior del cristalino. Durante la vida embrionaria la actividad mitótica se extiende a todas las células epiteliales externas del cristalino. Por la época del nacimiento, la actividad mitótica se detiene en la región central de este epitelio, y queda un anillo germinal de células con actividad mitótica alrededor de ella. Las células hijas de la región germinal se desplazan hacia la parte ecuatorial de la elongación celular, donde dejan de dividirse y adoptan las características citológicas de las células productoras de RNA y comienzan a formar RNA – mensajeros de las proteínas cristalinas. Estas células pronto experimentan una tremenda elongación, se llenan de proteínas cristalinas y se transforman en fibras secundarias que se disponen en capas concéntricas alrededor de las fibras primarias del núcleo del cristalino. La región de la línea media en la cual confluyen las fibras secundarias procedentes de puntos opuestos en el ecuador se denominan suturas anterior y posterior ( figura 12-5 D). Con esta organización, las fibras del cristalino dispuestas hacia la periferia son cada vez más jóvenes. Conforme el cristalino crece, nuevas fibras secundarias se desplazan desde el ecuador hacia la corteza externa.
Las proteínas cristalinas muestran un patrón y una secuencia de aparición muy característicos; las proteínas alfa cristalinianas aparecen en primer lugar en las células epiteliales que aún no se han diferenciado desde el punto de vista morfológico. La síntesis de las proteínas beta cristalinianas se ve cuando las fibras comienzan a alargarse, en tanto que la expresión de las de tipo gamma se ve restringida hasta la etapa finadle la diferenciación de las células. Cada grupo de proteínas cristalinianas está compuesto por varios miembros; sus patrones de activación y de acumulación son diferentes (algunos de los miembros de una familia se activan de forma coordinada). Los patrones de expresión de las proteínas cristalinianas varían considerablemente de una especie a otra. Se supone que estas proteínas facilitan la transparencia óptica del cristalino para permitir la transmisión de suficiente luz, pero todavía quedan muchos detalles por conocer.
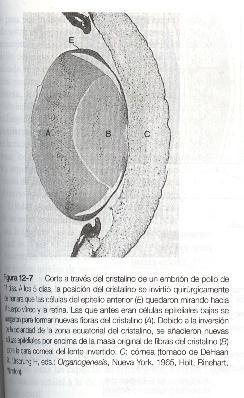
Durante gran parte de su vida, el cristalino se encuentra bajo la influencia de la retina. Después de la inducción del cristalino, las secreciones de la retina, de las cuales el factor de crecimiento de los fibroblastos (FGF) es un componente primordial, se acumulan en el humor vítreo por detrás del lente y estimulan la formación de las fibras del cristalino. Un ejemplo evidente de la influencia constante de la retina en la morfología del cristalino se observa cuando se rota a un cristalino en desarrollo de manera que su polo anterior quede mirando hacia la retina. Muy pronto, y se supone que bajo la influencia de las secreciones retinianas, las células epiteliales bajas del antiguo polo anterior comienzan a alargarse y a formar un juego adicional de fibras del cristalino (figura 12-7). En el lado corneal del cristalino rotado se forma un nuevo epitelio. Estas adaptaciones estructurales son evidencias asombrosas de la existencia de un mecanismo que asegura la alineación correcta entre el cristalino y el resto del sistema visual durante todo el desarrollo.
Formación de la cornea
La formación de la cornea es el resultado del último gran suceso inductivo que tiene lugar durante la formación del ojo (figura 12-2), en el cual la vesícula del cristalino actúa sobre el ectodermo que cubre su superficie. El resultado de esta inducción es la transformación de un ectodermo de superficie típico, que consta de una capa basal de células cuboideas y un peridermo superficial, con una estructura transparente de varias capas con una matriz extracelular compleja y aporte de células procedentes de varias fuentes.
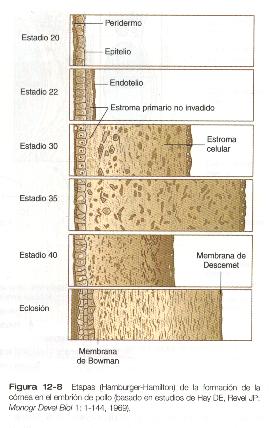
La influencia inductiva del cristalino estimula un cambio en las células ectodérmicas basales. Aumenta su altura, en gran parte como resultado de la elaboración de organelos secretorios (un ejemplo, el aparato de Golgi) en sus extremos basales. Conforme estos cambios van completándose, las células comienzan a segregar colágenos de los tipos I, II Y IX para formar el estroma primario de la córnea (figura 12-8).
Desde el estroma primario las células de la cresta neural situadas alrededor de la cúpula óptica migran en dirección central entre éste y la cápsula del cristalino. Si bien la morfología de estas células durante esta migración es mesenquimatosa, una vez que han terminado su viaje se transforman en el epitelio cuboide llamado endotelio corneal. En este punto la cornea primitiva consta de: 1) un epitelio externo. 2) un estroma primario aún celular y 3) un endotelio interno.
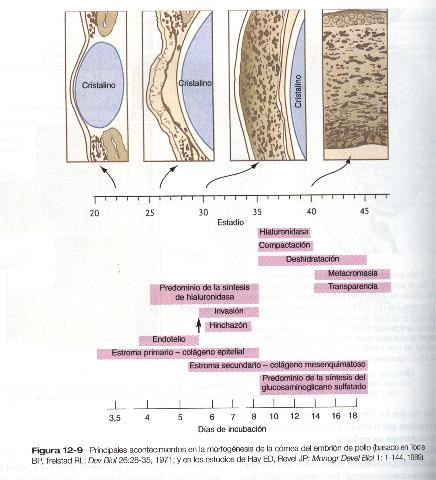
Después de que el endotelio de la córnea ha formado una capa continua, sus células sintetizan grandes cantidades de ácido hialurónico y lo segregan hacia el estroma primario. Debido a sus pronunciadas características hidrofílicas, el ácido hialurónico hace que el estroma primario se hinche de gran manera. Esto crea el sustrato apropiado para la segunda oleada migratoria de las células hacia la córnea en desarrollo (figura 12-9). Estas células, también derivadas de la cresta neural, son de naturaleza fibroblástica; migran y proliferan en los espacios ricos en ácido hialurónico localizados entre las capas de colágeno en el estroma corneal primario. La fase migratoria de las células del estroma corneal primario cesa cuando éstas comienzan a producir grandes cantidades de hialuronidasa, que degrada gran parte del ácido hialurónico que existe en el estroma primario. En otras partes del embrión (por ejemplo, la yema de la extremidad), también hay una estrecha correlación entre las cantidades elevadas del ácido hialurónico y la migración celular y una detención de la migración cuando este se elimina. Con la eliminación del ácido hialurónico, disminuye el grosor de la córnea. Una vez que los fibroblastos migratorios se han asentado, se considera que el estroma corneal primario se ha transformado en estroma secundario.
Los fibroblastos del estroma secundario contribuyen a la organización de la córnea mediante la secreción de fibras de colágeno hacia la matriz estromal. No obstante, tanto las células epiteliales como las endoteliales de la córnea continúan segregando grandes capas de matriz acelular. Estas secreciones proporcionan las capas que constituyen la córnea madura, a saber, desde afuera hacia dentro: 1) epitelio externo, 2) membrana de Bowman, 3) estroma secundario, 4) membrana de Decemet y 5) endotelio corneal (figura 12-8).
Los cambios finales del desarrollo de la córnea incluyen la formación de un trayecto transparente carente de distorsiones ópticas, a través del cual la luz entra en el ojo. Uno de los cambios principales es un gran aumento de la transparencia, es decir, una mayor capacidad de transmisión de la luz desde cerca del 40% hasta cerca del 100%. Esto se logra eliminando casi toda el agua del estroma secundario. La eliminación inicial del agua tiene lugar con la degradación de la mayor parte del ácido hialurónico que, como ya se dijo, es hidrofilico. La segunda fase está mediada por la tiroxina. Que es liberada en la sangre por la glándula tiroides en desarrollo. Esta hormona tiroidea actúa sobre el endotelio corneal haciéndole bombear sodio desde el estroma secundario hacia la cámara anterior. Las moléculas de agua siguen a los iones de sodio y así se logra una forma efectiva de deshidratación del estroma corneal. La función de la glándula tiroides en este proceso queda demostrada de dos modos: cuando se transplantan glándulas tiroides relativamente desarrolladas en las membranas extraembrionarias de embriones jóvenes de pollo y se permite que la hormona tiroidea llegue a la circulación embrionaria a través de los vasos sanguíneos que irrigan a las membranas (membrana corioalantoidea), tiene lugar la deshidratación prematura de la córnea. A la inversa, la aplicación de inhibidores tiroideos retrasa el aclaración de la córnea.
El otro suceso tardío en la córnea es un pronunciado cambio en su radio de curvatura con relación a la de todo el globo ocular. Este cambio morfogenetico, que comprende numerosos sucesos mecánicos (incluida la presión del líquido intraocular) permite que la córnea junto con el cristalino deje pasar los rayos luminosos hacia la retina. Si se desarrollan irregularidades en la curvatura de la córnea durante su morfogénesis final, el individuo sufre de astigmatismo, que causa distorsiones de la imagen visual.
Retina y otros derivados de la copa óptica
Mientras que se desarrollan el cristalino y la córnea, están sucediendo cambios profundos en la copa óptica (figura 12-1). La copa interna de la copa se engruesa y las células epiteliales comienzan un largo proceso de diferenciación en neuronas y en células fotorreceptoras de la retina neural. La capa externa de la copa óptica sigue siendo relativamente delgada y termina por convertirse en la capa pigmentaria de la retina (figura 12-5). Al mismo tiempo, los labios externos de la copa óptica experimentan una transformación en el iris del cuerpo ciliar, implicados en el control de la cantidad de luz que entra al ojo y en la curvatura del cristalino, respectivamente.
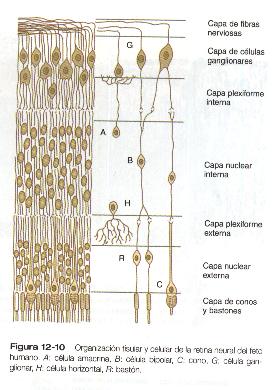
La retina neural es una estructura de múltiples capas y su desarrollo embrionario sólo puede apreciarse después de comprender su estructura adulta (figura 12-10). Cuando se ve un corte transverso de la retina al microscopio se observan franjas alternantes claras y densas que corresponden respectivamente a capas ricas en núcleos y en prolongaciones celulares. La vía sensorial de la retina neural es una cadena de tres neuronas que la cruzan en todo su espesor. El primer elemento de la cadena es la célula fotorreceptora, que puede ser un bastón o un cono. Un rayo de luz que entra al ojo pasa por todo el espesor de la retina neural hasta que incide sobre el segmento externo de un cono o un bastón (fotorreceptor) situados en la capa mas externa de la retina. El núcleo del cono y del bastón estimulados de la célula está localizado en la capa nuclear externa. Las células fotorreceptoras emiten una prolongación hacia la capa plexiforme externa, donde hacen sinapsis con una prolongación de una célula bipolar localizada en la capa nuclear interna. La otra prolongación de la neurona bipolar se introduce en la capa plexiforme interna y hace sinapsis con la tercera neurona de la cadena: la célula ganglionar. Los cuerpos de éstas últimas, localizados el la capa de células ganglionares, emiten largas prolongaciones que se desplazan por la capa mas interna de fibras nerviosas hacia su lugar de salida del ojo, el nervio óptico, a través del cual llegan al cerebro
Si todas las señales luminosas se procesaran sólo a través de las series sencillas de tres eslabones de neuronas de la retina, la agudeza visual sería mucho menor de lo que en realidad es. Para el momento en que un patrón visual se almacena en la corteza visual del cerebro ya han tenido lugar muchos niveles de integración. El primero está en la retina neural. En las uniones sinápticas tanto en las capas plexiformes interna como externa de la retina, están implicadas otras células como las horizontales y las amacrinas (figura 12-10) en la redistribución horizontal de una señal visual sencilla. Esto facilita la integración de los componentes de un patrón visual. Otro tipo importante de células de la retina es la célula glial de müller, que emite prolongaciones a casi todas las capas de la retina y parece desempeñar una función similar a la de los astrocitos en el sistema nervioso central.
Retina neural
Desde el epitelio columnar original de la capa sensorial interna de la copa óptica (figura 12-5), el primordio de la retina neural adopta la forma de un epitelio columnar engrosado y seudoestratificado con gran actividad mitótica organizado de manera similar al del tubo neural. Durante las primeras etapas del desarrollo de la retina, su polaridad se fija de acuerdo con la misma secuencia axial que se observa en las extremidades (capitulo 9). Primero se fija el eje nasotemporal (anteroposterior) y a continuación el eje dorsoventral. Por último, se establece la polaridad radial.
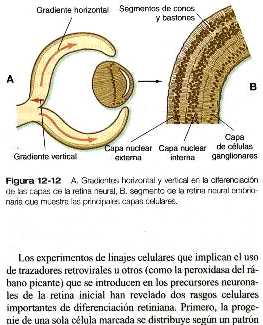
Conforme aumenta el número de células de la retina inicial, comienza la diferenciación de los tipos celulares. Hay dos gradientes principales de diferenciación en la retina: el primero sigue un trayecto aproximadamente lineal desde las capas internas hasta las externas de la retina; el segundo se desplaza en dirección horizontal desde el centro hasta la periferia.
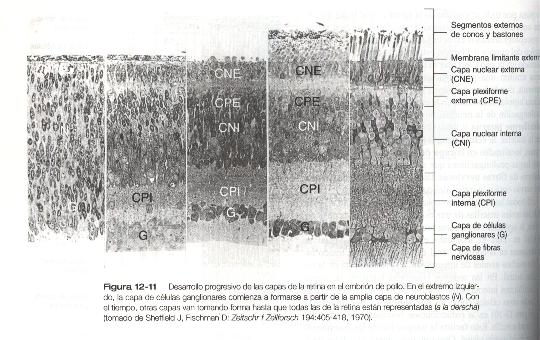
La diferenciación del primer gradiente comienza con la aparición de las células ganglionares y la definición precoz de la capa ganglionar (figura 12-11). A medida que las células ganglionares se diferencian, la actividad del gen Notch impide la diferenciación de las que las rodean. Una de las funciones principales de este gen es mantener las poblaciones celulares en un estado indiferenciado hasta que aparecen loas señales locales correspondientes para que sigan diferenciándose. Con la diferenciación posterior de las células horizontales y amacrinas, se forman las capas nucleares interna y externa. A medida que las células del interior de las capas nucleares emiten sus prolongaciones, las capas plexiformes interna y externa se definen mejor. Las neuronas bipolares, los conos y los bastones son los últimos en diferenciarse, y así termina el primer gradiente.
El gradiente horizontal de diferenciación de la retina neural se basa en la distribución desde el centro hacia la periferia de la retina de la expresión del primer gradiente vertical (figura 12-12). La retina no puede crecer hacia dentro. Así, durante la fase de crecimiento en el ojo humano (o durante la vida en el caso de los animales que crecen continuamente, como los peces), las células precursoras inmadura de la retina situadas a lo largo del borde de la retina experimentan mitosis formando un anillo concéntrico en expansión constante en la zona periférica de la retina. Dentro del anillo de mitosis tiene lugar la diferenciación, de una manera muy similar a la del gradiente vertical.
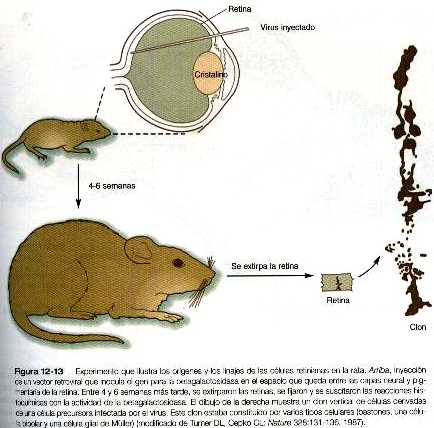
Los experimentos de linajes celulares que implican el uso de trazadores retrovirales u otros (como la peroxidasa del rábano picante) que se introducen en los precursores neuronales de la retina inicial han revelado dos rasgos celulares importantes de diferenciación retiniana. Primero, la progenie de una sola célula marcada se distribuye según un patrón radial notablemente recto que sigue el eje vertical de la diferenciación retiniana. Parece haber muy pocas mezcla lateral entre las columnas de las células de la retina (figura 12-13). El segundo rasgo de la diferenciación es que una sola célula precursora marcada puede dar lugar a más de un tipo de célula retiniana diferenciada.
Una etapa mas avanzada en la diferenciación retiniana es el crecimiento de los axones desde las células ganglionares a lo largo de la capa mas interna de la retina hacia el tallo óptico, crecen en su interior siguiendo señales emitidas por células que expresan Pax-2, y siguen su curso hacia los centros visuales del cerebro. Durante este crecimiento de los axones, éstos usan varias señales locales y de posición para establecer conexiones muy precisas con el cerebro. Gran parte de la investigación básica sobre el desarrollo de las conexiones entre la retina y el cerebro se ha hecho en embriones de peces y anfibios. En general, lo que hemos aprendido del desarrollo usual de estas especies se aplica al desarrollo del sistema visual humano.
Iris y cuerpo ciliar
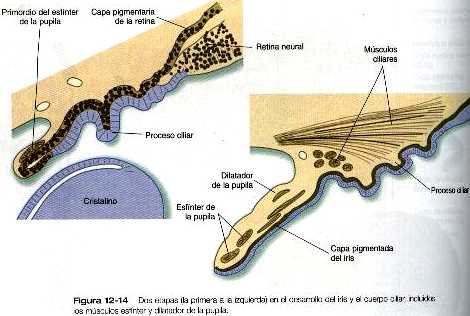
La diferenciación del iris y del cuerpo ciliar tiene lugar en el labio de la copa óptica donde se encuentran las retinas neural y pigmentaria. Más que sensoriales en su función, estas estructuras están implicadas en la modulación de la cantidad y las características de la luz que en último término incide sobre la retina.
El iris rodea parcialmente la parte exterior del cristalino y, mediante su contracción o relajación, controla la cantidad de luz que pasa por él. El iris contiene una capa epitelial interna no pigmentada y una capa externa pigmentada que se continúan con las capas neural y pigmentaria de la retina, respectivamente (figura 12_14). El estroma del iris, que es superficial a la capa pigmentada externa de esta estructura, se origina en la cresta neural y migra secundariamente hacia el iris. En este estroma se encuentran los primordios de los músculos esfínter y dilatador de la pupila. Éstos tienen un origen neuroectodérmico; parecen provenir de la capa epitelial anterior del iris.
Entre el iris y la retina neural se encuentra el cuerpo ciliar, una estructura muscular conectada al cristalino por grupos de fibras radiales que en su conjunto se llaman ligamento suspensorio del cristalino. Mediante las contracciones de la musculatura ciliar que actúan a través del ligamento suspensorio, el cuerpo ciliar modula la forma del cristalino para enfocar adecuadamente el estimulo visual de la retina. El desarrollo normal del cuerpo ciliar depende de una presión apropiada del líquido intraocular: si durante el desarrollo del ojo se elimina parte de este líquido, el resultado es un cuerpo ciliar defectuoso.
El color de los ojos es el resultado del nivel y la distribución de la pigmentación del iris. El color azulado del iris de la mayoría de los recién nacidos está causado por la pigmentación intrínseca de la capa pigmentada externa del iris. También aparecen células pigmentarias en el estroma del iris frente al epitelio pigmentario. Cuanto mayor sea la densidad del pigmento en esta área, mas marrón será el ojo. La pigmentación definitiva del ojo se desarrolla gradualmente durante los primeros seis a diez meses de vida postnatal.
Cuerpo vítreo y sistema de la arteria hialoidea
Al comienzo del desarrollo de la retina, un mesenquima laxo invade la cavidad de la copa óptica y forma una malla laxa de fibrillas junto con una sustancia gelatinosa que llena el espacio que queda entre la retina neural y el cristalino. Este material se llama cuerpo vítreo.
Durante gran parte del desarrollo embrionario el cuerpo vítreo es irrigado por la arteria hialoidea y sus ramas (figura 12_5). Esta arteria entra al ojo a través de la fisura coroidea del tallo óptico (figura 12_4), pasa por la retina y el cuerpo vítreo y termina ramificándose en la pared posterior del cristalino. Conforme avanza el desarrollo, las porciones de la arteria hialoidea que están en el cuerpo vítreo (así como sus ramas que irrigan el cristalino) involucionan por apoptosis de sus células endoteliales, lo que deja el conducto hialoideo. La parte mas proximal del sistema de la arteria hialoidea persiste y forma la arteria central de la retina y sus ramas.
Capas coroidea y esclerótica
Fuera de la copa óptica se dispone una capa de células mesenquimatosas, la mayoría de las cuales se origina en la cresta neural. Como respuesta a una acción inductiva del epitelio pigmentario de la retina, estas células se diferencian en estructuras que proporcionan el soporte vascular y mecánico al ojo. Las células mas internas de esta capa se transforman en una túnica muy vascularizada llamada capa coroidea o coroides (figura 12_5 c) y las mas externas forman una cubierta blanca de colágeno denso conocida como esclerótica. La esclerótica opaca forma la cubierta externa y rígida del ojo, y se continúa con la cornea. Los músculos extracelulares, que mueven el globo ocular, se insertan en la esclerótica.
Párpados y glándulas lacrimales
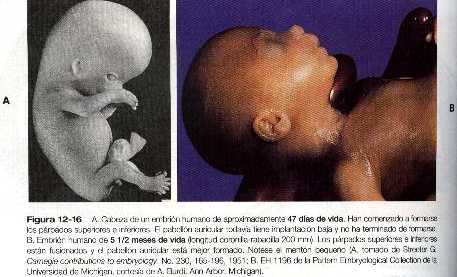
Los parpados comienzan a verse durante la séptima semana como pliegues de piel que se forman excéntricamente de la cornea (figuras 12_5 y 12_16 A). Una vez que su formación ha comenzado, crecen rápidamente sobre el ojo hasta que se encuentran y se fusionan hacia el final de la novena semana (figura 12_16 B). la fusión es transitoria y solo implica a las capas epiteliales de los párpados, lo que da como resultado una lamina epitelial común entre ellas. Antes de que los parpados vuelvan a abrirse, las pestañas y las pequeñas glándulas situadas a lo largo del borde palpebral comienzan a diferenciarse de la lámina epitelial común. Aunque al sexto mes pueden observarse signos de laxitud de la unión epitelial de los párpados, la reapertura de éstos sólo se produce bien entrado el séptimo mes de la gestación.
El espacio que queda entre la cara anterior del globo ocular y los párpados se conoce como saco conjuntival. En la fase de fusión de los párpados y a partir del ectodermo lateral superficial comienzan a crecer múltiples yemas epiteliales. Éstas formaran las glándulas lacrimales, las cuales producirán una secreción acuosa que baña la superficie externa de la córnea. Esta secreción pasa hacia la fosa nasal a través del conducto nasolacrimal (capítulo 13). Las glándulas lacrimales no están del todo desarrolladas al nacimiento, y los neonatos no tienen lágrimas cuando lloran; estos comienzan a producirse hacia las 6 semanas.
A pesar de los múltiples tipos de malformaciones del ojo y el sistema visual, la incidencia de los tipos mas frecuentes de estos defectos es bastante baja. En la correlación clínica 12_5 (falta) se dan ejemplos de algunas de las diferentes malformaciones oculares.
El oído es una estructura compleja compuesta por tres partes principales: el oído externo, el medio y el interno. El oído externo consta del pabellón (oreja), el meato auditivo externo (conducto auditivo externo) y las capas externas de la membrana timpánica (tímpano); su principal función es la de un aparato de captación de sonido. El oído medio actúa como un dispositivo de transmisión; esta función la cumple la cadena de los Tres huesillos del oído medio, que conectan la cara interna del tímpano con la ventana oval del oído interno. Otros componentes del oído medio son la cavidad (cavidad timpánica), la trompa auditiva (trompa de Eustaquio), la musculatura del oído medio y la cara interna de la membrana timpánica. El oído interno contiene el aparato sensitivo primario, que participa tanto en la audición como en el equilibrio. Estas funciones las cumplen respectivamente la cóclea (o caracol) y el aparato vestibular.
Desde el punto de vista embriológico, el oído tiene un origen doble. El oído interno surge a partir de una plácoda ectodérmica engrosada localizada en el rombencéfalo. Las estructuras del oído medio y el externo derivan del primer y del segundo arcos branquiales y de la primera hendidura branquial y de la bolsa faríngea que son estructuras intermedias.
Desarrollo del oído interno
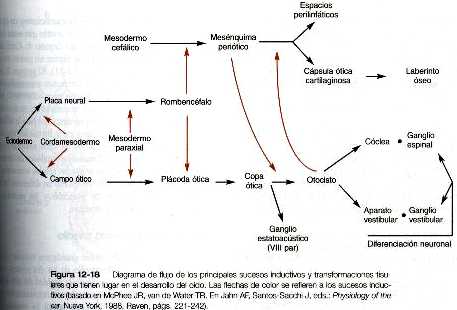
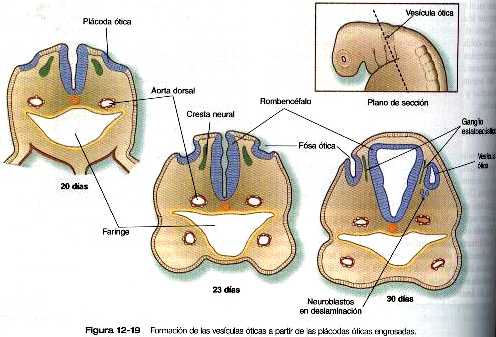
El desarrollo del oído comienza con la inducción inicial sobre el ectodermo superficial, primero a cargo de la notocorda (cordamesodermo) y después del mesodermo paraxial (figura 12_18). Estas dos inducciones preparan al ectodermo para una tercera, en la cual el rombencéfalo induce al ectodermo superficial adyacente para que se engruese y forme la plácoda ótica (figura 12_19).al final de la cuarta semana y posiblemente bajo la influencia del FGF- 3 segregado por el rombencéfalo adyacente, la plácoda ótica invagina, después se separa del ectodermo de la superficie para formar la vesícula ótica u otocisto. Ya desde las etapas más incipientes del desarrollo del otocisto, la expresión de ciertas moléculas presagia la formación de derivados morfológicos específicos del otocisto.
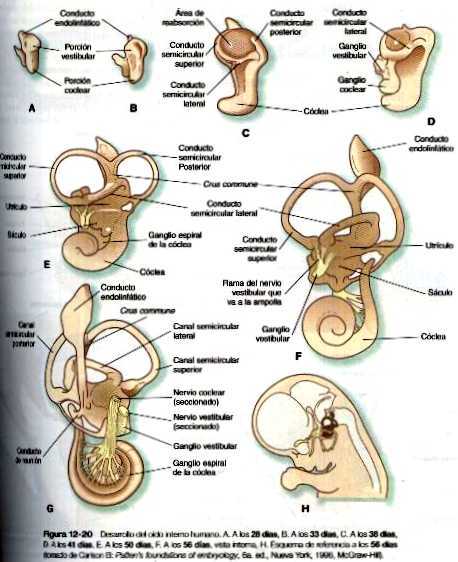
Pronto, la vesícula ótica comienza a alargarse y forma una región dorsal vestibular y una región ventral coclear (figura 12_20 A). El gen par homeobox, pax-2, tiene una gran participación en el desarrollo inicial de la vesícula ótica; si no se expresa, no se forman la cóclea ni el ganglio espiral. Bastante pronto surge el conducto endolinfático en forma de una proyección digitiforme corta que surge de la superficie dorsomedial del otocisto (figura 12_20 B). El FGF-3 secretado por los rombómeros 5 y 6 parece ser necesario para el desarrollo normal del conducto endolinfático. Cerca de la quinta semana, la aparición de dos crestas en la porción vestibular del otocisto presagia la formación de dos canales semicirculares (figura 12_20 C). Conforme estas crestas se expanden hacia los lados, un área de muerte celular programada situada en la cresta convierte las estructuras digitiformes densas en canales al producirse una zona de reabsorción (figura 12_20 C). Los precursores epiteliales de los canales semicirculares expresan los factores de trascripción de los genes portadores de los homeobox NKX5-1Y NKX5-2. Si estos genes se inactivan en los ratones, no se forman los canales semicirculares. Así, el desarrollo de las dos partes principales del oído interno está bajo controles genéticos separados: pax-2 para la porción auditiva (cóclea) y Nkx5 para la porción vestibular (canales semicirculares). La parte coclear del otocisto comienza a crecer en forma espiroidea; a las ocho semanas ha realizado una vuelta completa y dos vueltas a las diez semanas (figura 12_20 de la C a la F). la ultima media vuelta de la espiral del caracol (tiene un total de 21/2 vueltas) viene a completarse hacia la semana 25.
El oído interno (laberinto membranoso) está rodeado y englobado en una cápsula de tejido esquelético que comienza como una condensación del mesénquima mesodérmico alrededor del otocisto en desarrollo a las seis semanas de gestación. El proceso de englobamiento del otocisto comienza con una inducción del mesenquima circundante por parte del epitelio del otocisto (figura 12_18). Esta inducción estimula a las células mesenquimatosas, sobre todo de origen mesodérmico, para que formen una matriz cartilaginosa (a partir de la octava semana). El cartílago capsular sirve después como un molde para la formación posterior del laberinto óseo verdadero. La conversión del laberinto cartilaginoso en óseo se produce entre las 16 y las 23 semanas de gestación.
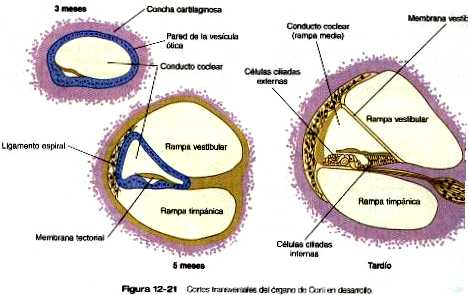
Las neuronas sensoriales que componen el VIII par craneal (en especial el ganglio estatoacústico) proceden de células que migran desde una porción de la pared interna del otocisto (figura 12_19). La parte coclear (ganglio espiral) del VIII par se origina en estrecha asociación con las células sensoriales (que en su conjunto reciben el nombre de órgano de Corti) que se desarrollan en la cóclea. Células de la cresta neural invaden el ganglio estatoacústico en desarrollo y forman las células satélites y de sostén que están en su interior. Las células sensoriales del órgano de Corti también derivan del epitelio del otocisto; tienen un patrón muy complejo de diferenciación (figura 12_21).al parecer, la generalización de precursores de los neuroblastos sensoriales del oído interno usa la vía del gen Notch para controlar la proporción de células epiteliales que se convierten en neuroblastos de una forma similar a la que se describió para la diferenciación de las células ganglionares en la formación inicial de la retina (Pág. 277). Como en otros sistemas sensoriales, unos controles muy finamente regulados del desarrollo aseguran aseguran el acople preciso entre las células sensoriales destinadas a recibir las ondas de sonido en diferentes frecuencias o la información de posición y las neuronas que transmiten señales al cerebro.
Desarrollo del oído medio
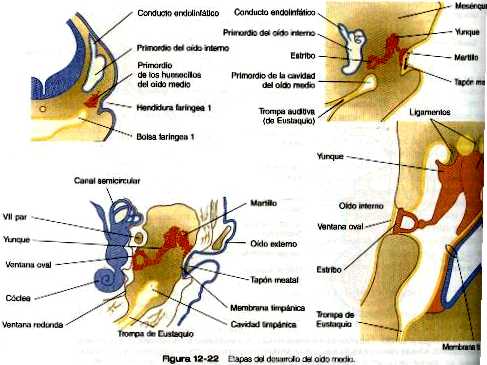
La formación del oído medio guarda una estrecha relación con los procesos del desarrollo de los dos primeros arcos branquiales (capitulo 13). Tanto la cavidad del oído medio como la trompa auditiva proceden del crecimiento de la primera bolsa faríngea. Llamada surco (o evaginación) tubotimpánico (figura 12_22). Este origen asegura que toda la cavidad del oído medio y la trompa auditiva estén tapizados por un epitelio derivado del endodermo.
Hacia el final del segundo mes del embarazo, el extremo ciego del surco tubotimpánico (bolsa faríngea 1) se aproxima a la porción mas interna de la primera hendidura faríngea. No obstante, estas dos estructuras aún están separadas por una masa de mesénquima. Más adelante, el epitelio endodérmico del surco tubotimpánico se adosa al revestimiento ectodérmico de la primera hendidura faríngea, aunque siempre estarán separados por una delgada capa del mesodermo. Esta estructura contiene tejidos de las tres capas germinales y se convierte en la membrana timpánica (tímpano). Durante la vida fetal, un hueso de forma anular denominado anillo timpánico rodea al tímpano. Algunos experimentos han demostrado que este hueso tiene una participación activa en la morfogénesis de la membrana timpánica.
En posición inmediatamente dorsal al extremo del surco tubotimpánico, a las seis semanas aparece una delgada condensación de mesénquima derivado de la cresta neural que gradualmente va adoptando la forma de los huesecillos del oído medio. Estos huesecillos, que descansan en un lecho de tejido conjuntivo embrionario muy laxo, se extienden desde la capa interna del tímpano hasta la ventana oval del oído interno. Auque la cavidad del oído medio está rodeado por el hueso temporal en desarrollo, el futuro antro del oído medio permanece lleno de mesénquima laxo casi hasta el final del embarazo. Durante el octavo y el noveno meses, gracias a la muerte celular programada y a otros procesos de reabsorción, la cavidad se va formando, lo que deja a los huesecillos del oído medio suspendidos en su interior. Al nacimiento puede haber restos del tejido conectivo del oído medio que limitan la libertad de movimiento de los huesecillos auditivos. La total movilidad se adquiere hacia los dos meses de vida extrauterina. La desaparición del tejido conectivo de la cavidad del oído medio coincide con la expansión del epitelio endodérmico del surco tubotimpánico, que al final tapiza toda la cavidad del oído medio.
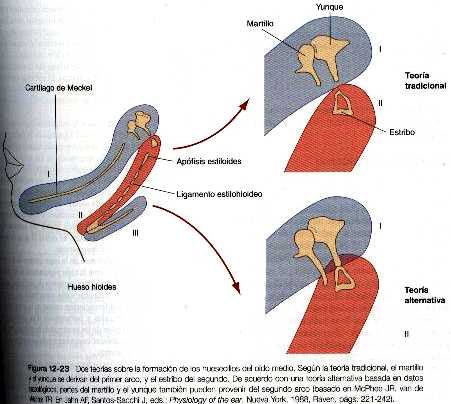
Los huesecillos auditivos tienen un origen doble. Según los datos de la anatomía comparada, el martillo y el yunque proceden del mesodermo del primer arco branquial, en tanto que el estribo se origina en el mesodermo del segundo arco (figura 12_23). No obstante, se ha puesto una segunda hipótesis basada en estudios realizados en sujetos con ciertos trastornos genéticos, según lo cual solo la cabeza del martillo y el cuerpo del yunque provienen del mesodermo del primer arco y el resto de estos dos huesos más el estribo se originan en el segundo arco.
Dos músculos ayudan a modular la transmisión de los estímulos auditivos a través del oído medio. El músculo tensor del tímpano, que está insertado en el martillo, se deriva del mesodermo del primer arco y, en consecuencia, está inervado por el nervio trigémino (V par). El músculo estapedio se inserta en el estribo, se origina en el segundo arco y está inervado por el nervio facial (VII par), que inerva a los derivados de ese arco.
Desarrollo del oído externo
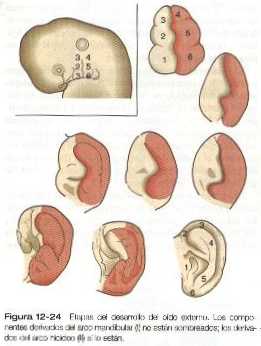
El oído externo (pabellón) se deriva de tejido mesenquimatoso de los dos primeros arcos branquiales que flanquean la primera hendidura faríngea (hiomandibular). Durante el segundo mes, tres masas nodulares de mesénquima (tubérculos auriculares) se forman a cada lado de la primera hendidura faríngea (figura 12_24). Los tubérculos auriculares crecen y terminan por fusionarse hasta formar un oído externo reconocible. Durante su formación, el pabellón se desplaza desde la base del cuello hasta su localización normal en el adulto a cada lado de la cabeza. Debido a su intima relación con los arcos branquiales y a su complejo origen, el oído externo es un buen indicador del desarrollo anómalo de la región faríngea. Otras anomalías del primero y el segundo arcos suelen acompañarse de malformaciones o mala posición de los pabellones auriculares.
El meato auditivo externo se forma durante el final del segundo mes mediante una investigación de la primera hendidura faríngea. Al comienzo del tercer mes el epitelio ectodérmico del meato en formación prolifera y constituye una masa sólida de células epiteliales que se llama tapón meatal (figura 12_22). Más adelante en el periodo fetal (a las 28 semanas) un conducto que se forma en el interior del tapón meatal se transforma en la prolongación del meato auditivo externo que va hasta el nivel del tímpano.
El oído externo y el conducto auditivo externo son muy sensibles a los fármacos. La exposición a agentes como estreptomicina, talidomida y salicilatos durante el primer trimestre puede causar agenesia o atresia de estas dos estructuras. Las malformaciones congénitas del oído se discuten en la correlación clínica 12_2.
· El ojo comienza como una evaginación (surco óptico) de la pared lateral del diencéfalo. El surco se agranda para formar las vesículas ópticas que inducen al ectodermo que las cubre a formar el primordio del cristalino. El tallo óptico, que conecta la copa óptica con el diencéfalo, tiene una hendidura que contiene la arteria hialoidea, la cual irriga al ojo en desarrollo. El gen homeabox pax-6 actúa como gen de control maestro en el desarrollo del ojo.
· Bajo la influencia del pax-6 se forma el cristalino a partir de un engrosamiento ectodérmico que se invagina para formar la vesícula del lente. Las células de la pared interna de esta vesícula se alargan y sintetizan las proteínas cristalinas específicas de esta estructura. En el cristalino en crecimiento el epitelio interno forma una masa esférica de fibras del cristalino que tienen forma alargada (células epiteliales). El epitelio anterior del cristalino está compuesto por células epiteliales cuboides. La polaridad global del cristalino se encuentra bajo la influencia de la retina.
· La cornea se forma gracias a la inducción hecha por el cristalino sobre el epitelio superficial. Después de la inducción, las células ectodérmicas basales segregan una matriz extracelular que sirve de sustrato para la migración de las células de la cresta neural que forman la capa endotelial de la córnea. Las células endoteliales de la córnea segregan grandes cantidades de ácido hialurónico hacia el interior de las cornea inicial. Esto permite la migración de una segunda oleada de células de cresta neural hacia la córnea. Estas células similares a fibroblastos segregan fibras de colágeno hacia la densa matriz del estroma corneal. Bajo la influencia de la tirosina, se extrae el agua de la matriz corneal y ésta se vuelve transparente.
· La retina neural se diferencia de la capa interna de la copa óptica. La capa externa forma la capa pigmentaria de la retina. La retina neural es una estructura compleja de múltiples capas en la cual 3 capas de neuronas están conectadas por prolongaciones celulares. La diferenciación celular en la retina neural sigue gradientes verticales y horizontales. Las prolongaciones celulares se desprenden de las neuronas de la retina y se van por el tallo óptico a establecer conexiones con los centros visuales localizados en el cerebro.
· El iris y el cuerpo ciliar se forman a partir del borde externo de la copa óptica. En el interior del iris se forman los músculos esfínter y dilatador de la pupila. Los niveles y la distribución del pigmento dentro del iris determinan el color de los ojos. Por fuera de la copa óptica. el mesénquima se transforma en una cubierta vascularizada llamada coroides y en la esclerótica, capa compuesta por colágeno resistente. Los párpados comienzan como pliegues de piel que se forman por encima de la cornea y luego se fusionan y cierran los ojos. Los parpados vuelven a abrirse al final del séptimo mes.
· Durante su desarrollo, los ojos son muy sensibles a varios teratógenos e infecciones intrauterinas. La exposición a estas noxas puede causar microftalmía o cataratas congénitas. La falta de cierre de la fisura coroidea da como resultado el coloboma.
· El oído interno surge por la inducción que causa el rombencéfalo en desarrollo sobre el ectodermo de superficie. Los pasos en su formación incluyen el engrosamiento del ectodermo (plácoda), la invaginación para formar una vesícula ótica y, más adelante, el crecimiento y la morfogénesis de las porciones auditiva (coclear) y vestibular (canales semicirculares). El desarrollo normal de la cóclea depende de la expresión apropiada del pax-2, en tanto que para la formación de los canales semicirculares se requiere Nkx5.
· El desarrollo del oído medio está asociado con la primera hendidura faringea y el primer arco branquial a ambos lados. En la cavidad del oído medio (antro timpánico) se forman los huesecillos del oído medio y los músculos que se relacionan con ellos.
· El oído externo surge de seis pequeños nódulos de mesénquima que se forman en el tejido del arco branquial que rodea a la primera hendidura faringea.
· La sordera congénita puede presentarse después de varias afecciones intrauterinas como la infección por el virus de la rubéola. Las anomalías estructurales del oído externo son comunes.
CASO CLÍNICO
Se pide a una pediatra que examine a un niño pequeño que acaba de llegar de un país en el cual el acceso a los servicios médicos es deficiente. El niño tiene orejas malformadas y de implantación baja, una mandibula hipoplástica y un profundo déficit auditivo. Los dientes también están mal alineados.
¿Cuál es el denominador común de este grupo de problemas?
La médica también solicitó un estudio de imágenes diagnósticas de los riñones y del tracto urinario del niño. ¿Para que lo hizo?
PREGUNTAS DE REPASO
1,¿De que tejido ocular las células derivadas de la cresta neural forman un constituyente importante?
A. Retina neural
B. Cristalino
C. Nervio óptico.
D. Cornea
E. Ninguno de los anteriores
2. La plácoda ótica surge gracias a mensajes inductivos emitidos por
A. El telencéfalo.
B. El rombencéfalo.
C. El infundíbulo.
D. El diencéfalo.
E. El mesencéfalo.
3¿Cuál de las siguientes moléculas intervienen en la dirección de los axones retinianos en su avance por el nervio óptico?
A. Pax-2.
B. FGF-3
C. Proteína 4 morfogenética ósea.
D. Pax-6.
E. Proteína 7 morfogenética ósea.
4. El ectodermo de la superficie recibe el mensaje inductivo de convertirse en epitelio corneal tras una oleada de inducciones que se originan en:
A. La copa óptica
B. El cordomesodermo.
C. La vesícula óptica.
D. La vesícula del cristalino
E. La retina neural.
5. El segundo arco branquial contribuye a la formación de:
A. La cóclea, el lóbulo de la oreja.
B. La trompa de Eustaquio, el yunque.
C. El estribo, el lóbulo de la oreja.
D. La trompa de Eustaquio, el estribo.
E. La vesícula ótica, el estribo.
6. Durante un examen medico convencional practicado a un niño se encontró que le faltaba un pequeño segmento en la parte inferior del iris. ¿Cuál es el diagnostico, cual es la base de esta afección y por qué podría esté niño ser tan sensible a la luz brillante?
7¿por qué las personas en ocasiones se les congestiona la nariz cuando lloran?
8¿Qué molécula de la matriz extracelular suele asociarse con las migraciones de las células mesenquimatosas y donde se produce un suceso de este tipo en el ojo en desarrollo?
9¿Por qué la audición del recién nacido con frecuencia no es tan aguda como lo es unos meses después?
10¿por qué es común que se asocien malformaciones o hipoplasia de la mandíbula con las anomalías de la forma o la implantación de las orejas?
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}