10
ESTABLECIMIENTO DEL SISTEMA NERVIOSO
CONFORMACIÓN INICIAL DEL SISTEMA NERVIOSO
HISTOGÉNESIS EN EL SISTEMA NERVIOSO
Proliferación en el tubo neural
Linajes celulares en la histogénesis del sistema nervioso central
Organización fundamental del tubo neural en desarrollo en un corte transversal
FORMACIÓN Y SEGMENTACIÓN DEL PATRÓN CRANEOCAUDAL
Establecimiento del patrón de las regiones del rombencéfalo y la médula espinal
Establecimiento del patrón en la región del mesencéfalo
Establecimiento del patrón en la región del prosencéfalo
Organización estructural de un nervio periférico
Patrones y mecanismos de los procesos de formación de las neuritas
Relaciones entre las neuritas y sus dianas durante el desarrollo de un nervio periférico.
Factores que controlan el número y el tipo de conexiones entre las neuritas y los órganos diana en el sistema nervioso periférico.
La muerte celular (apoptosis)
Sistema nervioso simpático
Sistema nervioso parasimpático
Diferenciación de las neuronas autónomas
Megacolon agangliónico congénito (enfermedad de Hirschsprung)
CAMBIOS ESTRUCTURALES MÁS TARDÍOS EN EL SISTEMA NERVIOSO CENTRAL
Histogénesis del sistema nervioso central
VENTRÍCULO, MENINGES Y FORMACIÓN DEL LÍQUIDO CEFALORRAQUÍDEO
PARES CRANEALES
DESARROLLO DE LA FUNCIÓN NEURAL
CORRELACION CLÍNICA 10_1
Malformaciones congénitas del sistema nervioso
DEFECTOS DE CIERRE DEL TUBO NEURAL
OTROS DEFECTOS DE CIERRE
PREGUNTAS DE REPASO
En la formación del sistema nervioso están implicados muchos procesos fundamentales. Algunos de éstos predominan en ciertas etapas de la embriogénesis; otros se presentan sólo en una fase limitada y en localizaciones restringidas. Los principales son:
ESTABLECIMIENTO DEL SISTEMA NERVIOSO
Como se describió en el capítulo 4, la inducción primaria del sistema nervioso provoca la formación de una placa neural de ectodermo engrosado que cubre la notocorda. Los estudios moleculares llevados a cabo sobre todo en anfibios han arrojado nueva luz sobre los mecanismos de la inducción neural. Gran parte del ectodermo dorsal de los embriones en fase de gastrulación produce una proteína de señal, la proteína 4 morfogenética ósea (BMP_4), que parece inhibir al ectodermo dorsal para que no forme tejido neural. En vez de enviar señales inductivas positivas al ectodermo suprayacente, los inductores neurales, nogina y cordina (y la folistatina en los anfibios), bloquean la influencia inhibitoria del BMP_4, permitiendo así que el ectodermo dorsal forme tejido neural (la placa neural (figura 4_10)). Bajo la influencia de la nogina y la cordina solos, la placa neural inducida desarrolla características de prosencéfalo, mientras que el ectodermo neural inducido en presencia de factor 8 de crecimiento de los fibroblastos (FGF_8) forma la placa neural caudal (médula espinal). El ectodermo neural inducido en presencia tanto de nogina como de FGF_8 desarrolla características de prosencéfalo y de rombencéfalo. Poco después de la inducción neural, la placa neural comienza a plegarse para formar el tubo neural (figura 5_11).
Durante estas etapas muy incipientes del establecimiento del sistema nervioso, se presenta un impresionante cambio en la distribución de las moléculas de adhesión celular (CAM) sobre las superficies de las células ectodérmicas. Las CAM sobresalen de la membrana plasmática. Es típico de las células de características similares se adhieran por medio de iones de Ca++, que fijan las CAM de una célula a las de otra (figura 4_15). En los embriones de pollo, las células del epiblasto inicial expresan tanto N_CAM como L_CAM (E-cadherina) antes de que tenga lugar la inducción primaria. Conforme la placa neural toma forma después del primer suceso de inducción, los profundos cambios que se han producido en el ectodermo se reflejan no sólo en las formas de las células sino también en su expresión de las CAM. Las células neuroepiteliales pierden su L_CAM y expresan sólo N_CAM, en tanto que las células ectodérmicas no inducidas pierden la N_CAM pero retiene la L_CAM.
El tubo neural, que es la manifestación estructural de las primeras etapas del establecimiento del sistema nervioso, es una estructura prominente. En los seres humanos, domina el polo cefálico del embrión (figura 5_10). Este capítulo describe la forma en que el tubo neural se desarrolla en los componentes principales morfológicos y funcionales del sistema nervioso.
CONFORMACIÓN INICIAL DEL SISTEMA NERVIOSO
El cierre del tubo neural se produce primero en la región en la que aparecen los primeros somitas y se extiende en ambas direcciones, craneal y caudal (figura 5_10). Las regiones no fusionadas del tubo neural se conocen como los neuroporos anteriores y posteriores. Incluso antes del cierre de los neuroporos (24 días de gestación para el neuroporo craneal y 26 para el caudal), pueden distinguirse algunas subdivisiones fundamentales del sistema nervioso incipiente. Son reconocibles la futura médula espinal y el encéfalo, dentro del cual es posible distinguir el cerebro anterior (prosencéfalo), el cerebro medio (mesencéfalo) y el cerebro posterior (rombencéfalo) (figura10_1 A).
Una fase importante en la conformación del sistema nervioso es el plegamiento global del polo cefálico del embrión en forma de C. Este plegamientos se asocia con la aparición, al final de la tercera semana, de una prominente curvatura cefálica del encéfalo en el nivel del mesencéfalo (figura 10_1 A). Pronto el encéfalo casi se dobla sobre sí mismo por la curvatura cefálica. Al comienza de la quinta semana aparece la curvatura cervical en la unión entre el rombencéfalo y la médula espinal.
Hacia la quinta semana las tres partes iniciales del encéfalo se han subdividido adicionalmente en cinco (figuras 10_1 B y 10_2). El prosencéfalo da lugar al telencéfalo (cerebro terminal), que tiene unas prominentes evaginaciones laterales las cuales terminarán por convertirse en los hemisferios cerebrales, y un diencéfalo en situación más caudal. El diencéfalo en situación más caudal. El diencéfalo se reconoce con facilidad a causa de las prominentes vesículas ópticas bilaterales que se evaginan desde sus paredes laterales. El mesencéfalo, que está doblado en ángulo agudo en la curvatura cefálica, permanece indiviso y tubular en su estructura general. El techo del rombencéfalo se adelgaza bastante y aparecen los primeros indicios de la subdivisión del mismo en metencéfalo y mielencéfalo más caudal. Estas cinco subdivisiones del encéfalo primitivo representan una organización fundamental que persiste hasta la edad adulta. Muchos otros componentes estructurales y funcionales añaden capas de complejidad al encéfalo en las siguientes semanas de la vida embrionaria.
HISTOGÉNESIS EN EL SISTEMA NERVIOSO
Proliferación en el tubo neural
Poco después de la inducción, la placa neural que está engrosándose y el tubo neural primario se organizan en un epitelio seudoestratificado (figura10_3). En este tipo de epitelio los núcleos parecen estar localizados en varias capas separadas dentro de las células neuroepiteliales alongadas. Los núcleos emprenden grandes desplazamientos de posición en el citoplasma de las células neuroepiteliales.
Las células neuroepiteliales se caracterizan por un alto grado de actividad mitótica, y la posición de los núcleos en el interior del tubo neural y su etapa del ciclo mitótico guardan una estrecha relación (figura 10_4). La síntesis de DNA tiene lugar en los núcleos localizados cerca de la membrana limitante externa (la lámina basal que rodea al tubo neural). Conforme estos núcleos se preparan para entrar en mitosis, migran dentro del citoplasma hacia la luz del tubo neural, donde comienzan la división mitótica.
Los núcleos de las dos células hijas resultantes migran hacia arriba en el citoplasma hacia la membrana limitante externa. En este punto tiene dos opciones: 1) comenzar de nuevo la síntesis del DNA y regresar al ciclo mitótico, o 2) dejar para siempre el ciclo mitótico y terminar ocupando posiciones por debajo de la membrana limitante externa como neuroblastos. Éstos, que son células precursoras de las neuronas, comienzan a producir procesos celulares que terminan por convertirse en axones y dendritas.
Linajes celulares en la histogénesis del sistema nervioso central
Los orígenes de la mayoría de las células que se encuentran en el sistema nervioso central maduro pueden seguirse desde las células madre pluripotenciales situadas en el neuroepitelio primitivo (figura 10_5). Estas células pasan por numerosas divisiones mitóticas antes de madurar y convertirse en células progenitoras bipotenciales que dan lugar ya sea a células progenitoras neuronales o a gliales. Esta bifurcación del desarrollo se acompaña de un cambio en la expresión de los genes.
Las células madre pluripotenciales expresan una proteína filamentosa intermedia denominada nidina. La nidina se regula curso abajo conforme los descendientes de las células progenitoras bipotenciales se separan en células progenitoras neuronales, que expresan proteínas de neurofilamentos, y células progenitoras gliales, que expresan proteína ácida glial fibrilar.
Las células progenitoras neuronales dan lugar a una serie de neuroblastos. Los primeros neuroblastos bipolares poseen dos finos procesos citoplasmáticos delgados que contactan tanto con la membrana limitante externa como con la limitante del tubo neural. Por retracción del proceso interno, el neuroblasto bipolar pierde contacto con la limitante interna en el proceso de transformarse en un neuroblasto unipolar. Los neuroblastos unipolares acumulan grandes masas de retículo endoplásmico rugoso (sustancia de Nissl) en su citoplasma y después comienzan a emitir varios procesos citoplásmicos. En este punto, se les conoce como neuroblastos multipolares. Sus principales actividades en el desarrollo son emitir procesos axonales y dentríticos y establecer conexiones con otras neuronas u órganos terminales.
El otro linaje principal que se desprende de las células progenitoras bipotenciales en la línea glial. Las células progenitoras gliales continúan realizando mitosis y su progenie se divide en varias líneas. Una, la célula progenitora O_2 A (figura 10_5), es la precursora de dos líneas de células gliales que termina formando los oligodendrocitos y los astrocitos tipo 2. Otro linaje glial da lugar a los astrocitos tipo 1. El origen de los oligodendrocitos es objeto de debate desde hace mucho tiempo, pero los estudios recientes han demostrado que en los seres humanos estas células se derivan de células progenitoras localizadas en la zona de ventricular central (figura 10_6) a lo largo de la placa del suelo. Desde ahí se diseminan a todo el encéfalo y a la medula espinal y terminan formando las vainas de mielina que cubren los procesos neuronales en la sustancia blanca. La formación de los precursores de los oligodendrocitos depende de una señal inductiva procedente de la notocorda (posiblemente el sonic hedgehog). Si se transplanta la notocorda a lo largo del tubo neural dorsal, los precursores de los oligodendrocitos se diferencian allí, lo cual demuestra que las células que tienen el potencial de formar oligodendrocitos residen en esa región, pero que normalmente se desarrollan por falta de una señal inductora apropiada.
El tercer linaje glial tiene una historia más compleja. Las células progenitoras radiales dan lugar a las células de la glía radial, que actúan como cables de guía en el encéfalo para la migración de las neuronas jóvenes (figura 10_21). Cuando las neuronas están migrando a lo largo de las células de la glía radial durante la mitad del embarazo, inhiben la proliferación de dichas células. Después de la migración neural, las células de la glía radial, ahora libres de la influencia inhibidora de las neuronas, reanudan el ciclo mitótico. Su progenie puede transformarse en varios tipos celulares; al parecer algunos pueden cruzar líneas de linaje y convertirse en astrocitos tipo 1 (figura 10_5). Otra progenie se diferencia en diversos tipos de células especializadas de la glía o incluso en células ependimarias. Según algunos autores, las células neuroepiteliales restantes representan otra fuente de células ependimarias.
No todas las células del sistema nervioso central se originan en el neuroepitelio. Las células de microglía, que tienen funciones fagocíticas después de producirse un daño cerebral, son células inmigrantes derivadas del mesodermo. Las células de microglía no se encuentran en el encéfalo en desarrollo sino después de que penetran en él los vasos sanguíneos.
Organización fundamental del tubo neural en desarrollo en un corte transversal
La médula espinal en desarrollo es un prototipo útil para el estudio de los rasgas estructurales y funcionales generales del sistema nervioso central ya que conserva su organización fundamental durante la mayor parte de su desarrollo. Con el comienzo de la división celular en el tubo neural, el neuroepitelio se engruesa y parece estratificarse. La capa de células más próxima a la luz (canal central) del tubo neural sigue siendo epitelial y se llama zona ventricular (la zona ependimaria de al antigua bibliografía. Esta zona, que aún contiene células mitóticas, termina por convertirse en el epéndimo, un epitelio columnar que tapiza el sistema ventricular y el canal central del sistema nervioso central (figura 10_6).
Más allá de la zona ventricular se encuentra la zona intermedia (antes capa del manto), que contiene los cuerpos celulares de los neuroblastos posmitóticos en proceso de diferenciación. Conforme los neuroblastos continúan produciendo procesos axonales y dendríticos, los procesos conforman una zona marginal periférica que contiene procesos neuronales pero no cuerpos celulares.
A medida que la médula espinal madura, la zona ventricular se convierte en la sustancia gris, en la cual están localizados los cuerpos de las neuronas. La zona marginal recibe el nombre de sustancia blanca a causa del color que le dan los numerosos tractos de fibras nerviosas mielinizadas que pasan por esa capa (figura 10_6). Durante el desarrollo las poblaciones progenitoras en fase de proliferación en la zona ventricular terminan agotándose y las células remanentes se diferencian en el epitelio y la capa ependimaria.
Una vez que las capas básicas de la médula espinal se han establecido, es posible reconocer varios rasgos topográficos importantes en los cortes transversales de la médula. Un surco limitante en el interior del canal central divide la médula en una placa alar dorsal y una placa basal ventral a cada lado del canal central. Las placas alares derecha e izquierda están unidas en la región dorsal al canal central por una delgada placa del techo, y las dos láminas basales están conectadas en la región ventral por una placa del suelo.
La placa basal representa el componente motor de la médula espinal. Los axones que se originan de las neuronas localizadas en el asta ventral de la sustancia gris salen de la médula espinal convertidos en las raíces motoras ventrales de los nervios espinales (figura 10_13).
La sustancia gris de la placa alar, llamada asta dorsal, ésta asociada con funciones sensitivas. Los axones sensitivos procedentes de los ganglios raquídeos (derivados de la cresta neural entran en la médula espinal en forma de raíces dorsales y hacen sinapsis con las neuronas del cuerno dorsal. una pequeña proyección de sustancia gris situada entre las astas dorsal y ventral en los niveles T1 a L2 de la médula contiene los cuerpos celulares de las neuronas autonómicas. Esta proyección se denomina asta lateral o en ocasiones columna gris intermediolateral (figura 10_6).
La placa del suelo es mucho más que una conexión anatómica entre las placas basales derecha e izquierda. Las células de la futura placa del suelo son las primeras que se diferencian en la placa neural después de la inducción primaria del sistema nervioso. El trabajo experimental ha demostrado una influencia inductiva específica de la notocorda sobre las células neuroepiteliales que la cubren. Si se injerta una notocorda adicional a lo largo de la superficie lateral del tubo neural, las células neuroepiteliales situadas más cerca de ésta adquieren las propiedades de las células del suelo (figura 10_7). A la inversa, si se extirpa un segmento de notocorda normal, las células neuroepiteliales que la cubren no adquieren las propiedades de las células de la placa del suelo.
A través de su acción sobre la placa del suelo, la notocorda también ejerce un profundo efecto sobre la organización de las raíces dorsales y ventrales que entran y salen de la médula espinal. Si falta la notocorda, el tubo neural se cierra, aunque no se reconocen como tales las raíces dorsales y ventrales. En su lugar aparecen numerosas fibras nerviosas ectópicas (figura 10_8). Si se incide la futura placa del suelo, el lado del tubo neural sobre el cual se localiza la notocorda desarrolla raíces dorsales y ventrales normales, mientras que el lado que carece de estas estructuras da lugar a raíces ectópicas (figura 10_7 C).
Algunos estudios recientes proporcionan una base molecular para entender la configuración del patrón del corte transversal en el interior de la placa y el tubo neural primitivos (figura 10_9). En la placa neural temprana los factores de transcripción que contienen homebox, Pax_3, Pax_7, Msx_1 y Msx_2 se expresan en toda la placa neural. Incluso antes de que la placa neural, la notocorda, que en esta etapa está adherida a la placa neural en la línea media, libera sonic hedgehog. Las señales locales del shh estimulan a las células de la placa neural situadas directamente sobre la notocorda para que se transformen en placa del suelo. Una de las primeras etapas en esta transformación es la represión de la expresión de Pax_3 y Pax_7, lo cual permite que las células del neuroectodermo localizadas cerca de la línea media de la placa neural asuman un destino ventral (es decir, la placa del suelo, placa basal). Las células mismas de la placa del suelo se convierten en lugares de producción de sonic hedgehog. El shh producido tanto por la notocorda como por la placa del suelo se difunde entonces lateralmente a través de la placa neural e induce a las células de ambos lados de la placa del suelo a convertirse en motoneuronas.
Además de inducir las motoneuronas, la placa del suelo desempeña otros papeles en el sistema nervioso en desarrollo. Varios grupos de procesos neuronales cruzan de una lado del sistema nervioso central al otro a través de la placa del suelo en forma de axones comisulares. Estos axones, que provienen de cuerpos celulares neuronales ubicados en la mitad dorsal del tubo neural, son atraídos hacia la placa del suelo por moléculas específicas que se producen en esa región (por ejemplo, la netrina_1). En animales mutantes que crecen de netrina_1, los axones comisulares están desorganizados y no cruzan hacia el otro lado a través de la placa del suelo. La placa del suelo no sólo atrae ciertos tipos de axones, sino que también rechaza a otros. Un claro ejemplo es el nervio patético (IV par craneal), cuyos axones no cruzan al otro lado desde sus cuerpos celulares de origen a causa de la presencia de algunas moléculas que lo rechazan y que aún no están aclaradas en la placa del suelo del mesencéfalo. El desarrollo de la organización en el corte transversal del tubo neural comprende no sólo una influencia ventralizante de la notocorda, sino también una influencia opuesta dorsalizante del ectodermo epidérmico adyacente al tubo neural en desarrollo.
En las regiones laterales de la placa neural (futura región del tubo neural), la BMP-4 y la BMP_7 expresadas por el ectodermo no neural en la unión entre el ectodermo y la placa neural en la unión entre el ectodermo y la placa neural lateral, ejercen un efecto inductor dorsalizante sobre las células neuroectodérmicas al elevar los niveles de Pax_3, Pax_7, Msx_1 y Msx_2. Esto redunda en la formación de la placa del techo y la placa alar. Además, la misma influencia inductiva estimula la expresión de un factor de transcripción, el slug, en las futuras células de la cresta neural antes de que hayan dejado el tubo neural. Después del cierre del tubo neural, la influencia de la BMP induce la formación de interneuronas sensitivas en la mitad dorsal (placa alar) de la médula espinal.
FORMACIÓN Y SEGMENTACIÓN DEL PATRÓN CRANEOCAUDAL
Por medio de la inducción neural, el sistema nervioso central incipiente se organiza en regiones amplias que desarrollarán características craneales, medias y caudales (pág. 70). Pronto, esto va seguido de la aparición de las subdivisiones morfológicas mencionadas en la figura 10_2. En un nivel aún mas fino, los segmentos llamados rombómeros aparecen en la región del romboencéfalo (figura 5_12), y en el prosencéfalo aparece una serie menos diferenciada de subdivisiones llamadas prosómeros.
Establecimiento del patrón de las regiones del rombencéfalo y la médula espinal
Los rombómeros, que se presentaron en el capítulo 5, son el reflejo morfológico de un patrón de expresión muy ordenado en forma segmentada de varios genes de desarrollo importante (figura 10_10). Los genes Krox_20 y Hox parecen ser esencial en la determinación de la naturaleza segmentaría del rombencéfalo: Si el Krox_20 en los ratones es bloqueado, no se forman los rombómeros 3 y 5. El Krox_20 actúa comúnmente en el flujo de expresión de los genes Hox y ejerce control regulador local sobre ellos. El Krox_20 también corre arriba en el flujo de expresión de los genes para los receptores que restringen que las células de los rombómeros pares y los impares se mezclen entre sí. De una forma que aún no se comprende del todo, el ácido retinoico parece ejercer un control aún más amplio de la expresión de los genes Hox. Como en la segmentación de las vértebras (págs. 165_166), el exceso de ácido retinoico causa un desplazamiento rostral en la expresión de los genes Hox y un desplazamiento caudal a nivel de los rombómeros de manera que un rombómero dado se transforma en el siguiente más caudal.
Por el mismo motivo, el resultado de una deficiencia de ácido retinoico es la formación de un rombencéfalo pequeño sin rombómeros caudales.
La correspondencia entre los rombómeros del rombencéfalo en desarrollo y otras estructuras de la región del cráneo y de los arcos branquiales es notable (capítulo 13). Los pares craneales, que tienen un patrón muy ordenado en virtud del cual van a inervar estructuras derivadas de los arcos branquiales y otras estructuras de la cabeza, tienen u origen igualmente muy ordenado con respecto a los rombómeros (figura 10_11). Por ejemplo, el par craneal V inerva estructuras derivadas del primer arco branquial. Los pares craneales VII y IX inervan las estructuras del segundo y el tercer arcos, respectivamente. En embriones de aves, las especies más ampliamente estudiadas, los cuerpos celulares de los componentes motores de los pares craneales V, VII y IX al comienzo se encuentran exclusivamente en los rombómeros 2, 4 y 6. Los estudios con inyección de colorantes han demostrado que la progenie de un solo neuroblasto se queda dentro del rombómero que contiene la célula inyectada, lo cual sugiere que los compartimientos celulares. Los axones que contribuyen a un para craneal se extienden hacia los lados en el interior del rombómero y convergen en un lugar común de salida en el punto medio craneocaudal del rombómero. En una etapa algo más tardía del desarrollo, las motoneuronas que se originan en el siguiente rombómero más caudal (3, 5,7) extienden sus axones hacia los lados. No obstante, antes de que los axones alcancen el borde del rombómero, se cruzan en los rombómeros 2, 4 ó 6 y convergen sobre el lugar de salida de los axones motores en el rombómero de número par. Los estudios comparativos indican diferencias de especie en la relación entre los rombómeros y los orígenes de los pares craneales. Las relaciones entre loa neurómeros y los pares craneales se muestran en la tabla 10_1.
Los cuerpos celulares (en el interior del sistema nervioso central, la colección de cuerpos celulares de un par craneal se denomina núcleo) de los pares craneales que inervan los arcos branquiales surgen a lo largo del eje craneocaudal. Los núcleos motores de otros pares craneales que inervan estructuras somáticas (por ejemplo, los músculos extraoculares o la lengua) surgen en una columna craneocaudal diferente a lo largo del rombencéfalo y no ocupan rombómeros adyacentes (figura 10_11).
Las evidencias directas e indirectas indican que las propiedades de las paredes de los rombómeros impiden que los axones se extravíen hacia los rombómeros vecinos. Una propiedad celular, que también es característica de las regiones de somitas que restringen el movimiento de las células de la cresta neural, es la capacidad de las células de la pared del rombómero de unirse a lectinas específicas. En aparente contradicción con la compartimentalización que se acaba de describir, los procesos que crecen a partir de los neuroblastos sensitivos y de los nervios de un tracto llamado fascículo longitudinal medial son libres para cruzar las fronteras romboméricas. Los vasos sanguíneos entran primero en el rombencéfalo en la región de la placa del suelo poco después de la salida de los axones motores y se diseminan dentro de las uniones romboméricas. Se desconoce la forma en que las ramas vasculares reconocen las fronteras de los rombómeros.
En contraste con el rombencéfalo, el patrón de los nervios que surgen de la médula espinal no parece ser determinado por la compartimentalización craneocaudal que opera en la médula. En vez de ello, el carácter segmentario de los nervios raquídeos lo determina el mesodermo de los somitas a lo largo del tubo neural. Las motoneuronas que crecen a partir de la médula espinal y las células migratorias de la cresta neural pueden penetrar con facilidad el mesodermo rostral del somita, pero parece que la mitad caudal del mismo las rechaza. El resultado es un patrón regular de crecimiento de los nervios raquídeos por cada segmento corporal. Si se rota el tubo neural inicial sobre su eje craneocaudal el resultado no es un patrón anómalo de los nervios raquídeos; esto fortalece aún más el punto de vista de que el patrón de éstos no se genera dentro del tubo neural mismo. A pesar de muchos estudios realizados sobre las diferencias moleculares que hay entre el mesodermo anterior y el posterior de las somitas, todavía no se ha dilucidado la base molecular exacta de la repulsión o la atracción de los nervios raquídeos por las regiones específicas de los somitas.
Establecimiento del patrón en la región del mesencéfalo
Uno de los mecanismos fundamentales del establecimiento del patrón en la región del mesencéfalo es un centro emisor de señales moleculares localizado en la frontera entre el mesencéfalo y metencéfalo (figura 5_13). La principal molécula de señales parece ser el FGF_8, que se expresa en un estrecho anillo situado en el borde rostral del primer rombómero, una subdivisión del metencéfalo. En coordinación con el Wnt_1,el FGF_8 induce la expresión de los genes engrailed, En_1 y En_2, que se expresan en concentraciones decrecientes respecto del centro de señales de FGF_8 (figura 5_13). Este entro de señales induce y polariza la región dorsal del mesencéfalo y el cerebelo.
Los injertos del centro de señales, llamado región ístmica, o las cápsulas que liberan FGF_8 sólo dentro de regiones más craneales del prosencéfalo del embrión de ave inducen un segundo techo (mesencéfalo dorsal o colículos de los mamíferos). De manera parecida, lo injertos ístmicos en regiones del rombencéfalo pueden inducir estructuras cerebelares supernumerarias.
Una conclusión derivada de los estudios moleculares recientes es que, al menos en algunas especies, la frontera entre los futuros mesencéfalo y rombencéfalo no corresponde a la estrechez anatómica existente entre el mesencéfalo y el rombencéfalo. En vez de ello, está localizada en situación craneal a esa constricción en un plano marcado por el límite caudal de la expresión del gen con homebox, Otx_2.
Establecimiento del patrón en la región del prosencéfalo
Aunque algunos investigadores han subdividido el prosencéfalo inicial en segmentos llamados prosómeros (figura 10_12), existe muy poca información definitiva sobre el control del establecimiento del patrón del prosencéfalo. Los estudios realizados con los rombómeros, las células pueden migrar a través de las fronteras entre los prosómeros. En especial en la región del futuro telencéfalo, no parece haber gran compartimentalización. Se ha sugerido que una estrecha banda de expresión de sonic hedgehog en la frontera entre los futuros tálamos dorsal y ventral es un centro de señales del prosencéfalo, pero se necesita más investigación para verificar esa hipótesis. Aunque la notocorda no existe subyacente al prosencéfalo embrionario y tampoco posee una placa del suelo, el sonic hedgehog se expresa en una franja craneocaudal situada a lo largo del telencéfalo y el diencéfalo. Como en la médula espinal incipiente, el sonic hedgehog induce la formación de las motoneuronas ventrales en esas regiones del prosencéfalo.
SISTEMA NERVIOSO PERIFÉRICO
Organización estructural de un nervio periférico
La formación del nervio periférico comienza con el crecimiento de los axones desde los neuroblastos motores localizados en la placa basal (la futura asta ventral de la sustancia gris) de la médula espinal (figura 10_13). Cerca de la parte dorsal de la médula espinal, unos procesos delgados también comienzan a crecer a partir de los neuroblastos derivados de la cresta neural que se han aglomerado para formar ganglios raquídeos.
Las dendritas, que conducen los impulsos hacia el cuerpo celular de la neurona, crecen desde las neuronas sensitivas hacia la periferia. Los axones, que conducen los impulsos hacia fuera del cuerpo celular, penetran en la región dorsolateral de la médula espinal y terminan en el asta dorsal (la sustancia gris de la placa alar). En el interior de la sustancia gris, una interneuronas cortas conectan las terminaciones de los axones sensitivos con las motoneuronas. Estas tres neuronas conectadas (motrices, sensitivas e interneuronas) constituyen un arco reflejo simple a través del cual un estímulo sensitivo puede convertirse en una repuesta motriz sencilla. Las fibras nerviosas autónomas también están asociadas con los nervios raquídeos típicos.
En un nervio periférico los procesos neuronales pueden ser mieleinizados o no mielinizados. A nivel celular, la mielina es una vaina espiral de múltiples capas principalmente de material fosfolípidico formado por las células de Schwann (derivadas de la cresta neural) que se envuelven muchas veces alrededor de una fibra nerviosa a la manera de u pastel relleno de mermelada (figura 10_14). Este envoltorio sirve de aislante que determina en gran medida el carácter del impulso eléctrico (potencial de acción) que viaja a lo largo del proceso neuronal. Las fibras nerviosas no mielinizadas están empotradas en el citoplasma de las células de Schwann, pero carecen de las disposiciones espirales características de los procesos mielinizados (figura 10_14).
Las células de Schwann que rodean a los axones mielinizados y no mielinizados son diferentes no sólo en su morfología sino también, nada sorprendente, en sus patrones de expresión genética. A través de las acciones de una familia de proteínas similares a los factores de crecimiento (neurregulinas), el axón asociado con u precursor de una célula de Schwann promueve la diferenciación de la célula de Schwann y ayuda a determinar si ésta va a producir mielina o formara una célula de Schwann sin mielina. Los precursores de la célula de Schwann que no están asociados con axones no reciben el soporte de la neurregulinas y sufren la muerte celular programada, un mecanismo que preserva una proporción apropiada entre células de Schwann y axones.
En el sistema nervioso central el color de la sustancia blanca es el resultado de su elevado contenido de fibras nerviosas mielinizadas, mientras que la sustancia gris contiene sin mielina. Las células de Schwann no están presentes dentro del sistema nervioso central; allí la mielinización está a cargo de los oligodendrocitos. Aunque una célula de Schwann en una fibra nerviosa periférica mielinizada puede envolverse alrededor de sólo un axón o una dendrita, una sola célula de oligodendroglia puede mielinizar varias fibras en el sistema nervioso central.
Patrones y mecanismos de los procesos de formación de las neuritas
El crecimiento de las neuritas (axones o dendritas) implica muchos factores tanto intrínsecos como extrínsecos. Si bien es similar en muchos aspectos, el desarrollo de los axones y el de las dendritas muestra algunas diferencias fundamentales.
Una neurita que está alongándose activamente lleva en el extremo un cono de crecimiento (figura 10_15). Los conos de crecimiento se caracterizan por la presencia de una región expandida de citoplasma con numerosas proyecciones a manera de espículas denominadas filopodios Los estudios in vitro e in vivo de los nervios vivientes han demostrado que la morfología de un cono de crecimiento activo se encuentra en un estado de flujo constante, en el cual los filopodios se extienden y retraen en forma regular como si probasen el ambiente local. Los conos de crecimiento contienen numerosos organelos citoplásmicos, pero gran parte de la forma y la función de los filopodios depende de las grandes cantidades de mrcrofilamentos de actina que llenan estos procesos. En presencia de agentes que rompen los filamentos de actina, los filopodios se retraen y los conos de crecimiento dejan de funcionar de forma adecuada normal.
El hecho de que los conos de crecimiento progresen, se queden estáticos o cambien de dirección depende en gran medida de sus interacciones con el ambiente local. Si éste es favorable, un filopodio permanece extendido y se adhiere al sustrato que lo rodea, en tanto que otros filopodios del mismo cono de crecimiento se retraen. Dependiendo de la localización de los filopodios adheridos, el cono de crecimiento puede dirigir la neurita a la cual esté adherido hacia delante o hacer que cambie la dirección de su crecimiento. Este crecimiento parece ser orientado por cuatro tipos amplio de influencias ambientales: quimoatracción, atracción de contacto, quimorrepulsión y repulsión o rechazo de contacto. Al parecer, los procesos nerviosos en crecimiento se encuentran en ambientes diferentes cada par de cientos de micras y que algunos ambientes les envían señales para que continúen extendiéndose hacia delante, mientras que otros actúan enviando señales de “detenerse” o “girar”.
Los conos de crecimiento pueden responder a gradientes de concentración de sustancias difusibles (por ejemplo, el factor de crecimiento neural) o a débiles campos eléctricos locales. Una de las principales familias de moléculas quimio atrayentes se llama netrinas. Las opuestas (repelentes) de las netrinas son miembros de una familia de proteínas secretadas llamadas semaforinas.
Los conos de crecimiento también pueden responder a señales físicas o químicas procedentes del microambiente que los rodea. Por ejemplo, la mitad caudal del somita recházale crecimiento y la progresión de axones motores y de células de la cresta neural hacia esa zona. El rechazo se manifiesta por el colapso del cono de crecimiento y la retracción de los filopodios. Por otra parte, las glucoproteínas de la matriz extracelular, como la fibronectina y especialmente la laminina, promueve fuertemente la adhesión y el crecimiento de las neuritas. Las Proteínas integrantes de las membranas (integrinas) de las neuritas se fijan específicamente a la secuencia arginina-glicina-asparagina de las glucoproteínas y promueve la adhesión al sustrato que contiene estas moléculas.
Otras moléculas, como la N_cadherina, la N_CAM y la L1, intervienen en la adhesión intercelular en diversas etapas de la migración celular o el alargamiento de las neuritas. La N_cadherina, que usa Ca++ como agente iónico para enlazar las moléculas similares, interviene firmemente en la unión intercelular de células del neuroepitelio. También interviene en la adhesión de neuritas paralelas en crecimiento. En un nervio periférico, un axón pionero suele preceder a los demás en el crecimiento hacia su diana. Otros axones lo siguen y forman fascículos (haces) de axones, Las proteínas de adhesión intercelular como la L1, que ayudan a enlazar fibras nerviosas paralelas, facilitan la formación de fascículos. Si se administran anticuerpos contra la proteína L1 a un área de crecimiento de neuritas, este proceso se interrumpe. La N_CAM está presente en la superficie de la mayoría de los procesos nerviosos y fibras musculares embrionarias, e interviene en la iniciación de los contactos neuromusculares. Los anticuerpos contra N_CAM impide el desarrollo de las uniones neuromusculares en los embriones. Las neuritas en crecimiento interactúan con muchas otras moléculas, y el grado completo en que lo hacen apenas está empezando a aclararse.
Aunque pudiera pensarse que el cono de crecimiento es el que dirige el crecimiento de las neuritas, otros factores son importantes para el alargamiento de los axones. El transporte axonal es esencial para el crecimiento y el mantenimiento de los axones y las dendritas. En este proceso intracelular, los materiales producidos por el cuerpo celular de la neurona son transportados hacia los extremos de estas neuritas, que pueden tener varios metros de longitud en los seres humanos.
La columna vertebral citoesquelética del axón es un conjunto ordenado de microtúbulos y neurofilamentos. Los microtúbulos son polímeros largos y tubulares compuestos por subunidades de tubulina. Conforme un axón crece desde su cuerpo celular, las subunidades de tubulina se transportan por el axón y se polimerizan en el extremo distal del microtúbulo. El ensamblaje de neurofilamentos se organiza y se polariza de una manera similar: El lugar de estas adiciones citoesqueléticas está cerca de la base del cono de crecimiento, lo cual significa que el axón se alarga porque en su extremo distal algo se añade y no porque algo empuja desde las proximidades del cuerpo celular neuronal. Un acompañante característico del crecimiento axonal es la producción de grandes cantidades de proteínas asociadas con el crecimiento (GAP). Una de las más prominentes entre éstas es la GAP_43, que sirve de sustrato a la proteincinasa C, y está concentrada en el cono de crecimiento.
Los axones y las dendritas en crecimiento difieren entre sí. a diferencia de los axones, las dendritas contienen microtúbulos con polaridad dispuesta en ambas direcciones (figura 10_16). Otra gran diferencia es la ausencia de GAP_43 en las dendritas en crecimiento. Uno de los primeros signos de polaridad en una neurona en desarrollo es la concentración de GAP_43 en el axón en crecimiento y su desaparición de los procesos dendríticos.
Relaciones entre las neuritas y sus dianas durante el desarrollo de un nervio periférico.
Las neuritas en desarrollo continúan alargándose hasta que han establecido contacto con un órgano terminal apropiado. En el caso de las motoneuronas, ese órgano terminal es una fibra muscular en desarrollo. Las dendritas de las neuronas sensitivas se relacionan con varios tipos de dianas. El extremo dela neurita primero debe reconocer su diana apropiada, y entonces debe establecer una conexión funcional con ella.
En el caso de las motoneuronas, hay pruebas de que unas señales muy específicas guían a los nervios y loa axones individuales hacia sus músculos diana. Los estudios de rastreo y transporte han demostrado que los nervios motores en crecimiento que van a las extremidades inervan los músculos de éstas en un orden bien definido y que después de unas pequeñas alteraciones, buscarán los músculos correctos (figura 9_26). Algunos datos recientes sugieren que incluso en el nivel de las neuronas, los axones “rápidos” son atraídos hacia los precursores de fibras musculares rápidas y que los axones “lentos” van hacia los de fibras musculares lentas. Existen muchos ejemplos similares de especificidad de diana en las dendritas del sistema nervioso periférico y tanto de las dendritas como de los axones del sistema nervioso central.
Cuando un axón motor y una fibra muscular se encuentran, una serie compleja de cambios se producen tanto en el nervio como en la fibra muscular marcando la formación de una sinapsis funcional, en este caso llamada unión neuromuscular (figura 10_17). Los primeros cambios consisten en: 1) el cese del crecimiento del axón, 2) la preparación de la terminación nerviosa para la liberación final de las moléculas neurotransmisoras apropiadas y 3) las modificaciones de las fibras musculares en el lugar del contacto con el nervio para que el estímulo neural se pueda recibir y expresar como un estimulo contráctil. En la estabilización de las propiedades morfológicas y funcionales de esta sinapsis muy especializada intervienen componentes tanto neurales como musculares de la unión neuromuscular.
Uno de los primeros signos de la especialización en una unión neuromuscular incipiente es la formación de las vesículas sinápticas, causada por influencia de la fibra muscular. Estas vesículas almacenan y después liberan la sustancia neurotransmisora acetilcolina a partir del terminal nervioso (figura 10_17). Antes de que la fibra muscular en desarrollo entre en contacto con la motoneurona, los receptores de acetilcolina (de tipo no_unitivo) se diseminan a todo lo largo de la fibra muscular. Después del contacto inicial con el nervio, los mionúcleos situados en la vecindad de la unión neuromuscular producen receptores de acetilcolina específicos de la unión que reside en los pliegues posteriores a la unión que y son inducidos por el nervio en la membrana de la fibra muscular, y los receptores dispersos que no son de la unión desaparecen. Entre la terminal nerviosa y el aparato postsináptico de la fibra muscular se extiende una lámina basal que contiene moléculas que estabilizan a los receptores de acetilcolina en la unión neuromuscular y también acetilcolinesterasa, una enzima producida por la fibra muscular.
Factores que controlan el número y el tipo de conexiones entre las neuritas y los órganos diana en el sistema nervioso periférico.
En muchas etapas de la formación de un nervio periférico, las interacciones entre las neuritas en crecimiento y las estructuras diana influyen sobre el número y la calidad tanto de las fibras nerviosas como de las estructuras diana. La existencia de tales mecanismos se demostró a comienzos del siglo XX mediante el injerto de yemas de las extremidades en flancos. Los nervios motores y los ganglios sensitivos que inervaban las extremidades injertadas eran sustancialmente más grandes que los nervios raquídeos contralaterales, que sólo inervaban estructuras de la pared del cuerpo. El examen de la médula espinal en el nivel del transplante mostró astas ventrales de sustancia gris más grandes y que contenían más motoneuronas de lo normal para los niveles de la médula espinal que sólo inervaban las regiones de los flancos.
Experimentos adicionales de este tipo esclarecen las relaciones anatómicas normales, que muestran volúmenes relativamente más grandes de sustancia gris y nervios más grandes en niveles a partir de los cuales se inervan las extremidades normales.
Los experimentos de deleción, en los cuales se quita la yema de una extremidad antes de la evaginación de la neurita, o la ausencia congénita de extremidades, tienen como resultado la reducción del número de neuronas periféricas y del volumen de la sustancia gris en las regiones afectadas.
La muerte celular (apoptosis) de las neuronas desempeña un papel importante en el desarrollo. Por ejemplo, cuando se inerva por primera vez un músculo, éste recibe un número mucho más grande de neuronas que las que existen en el adulto normal. En un momento crítico del desarrollo mueren cantidades masivas de neuronas. Parece haber razones para este fenómeno al parecer paradójico, entre las que se incluyen:
Todas estas razones de la muerte celular neuronal pueden ser parte de una estrategia biológica general que reduce las conexiones iniciales superfluas para asegurar que se hagan suficientes conexiones correctas. La otra estrategia de desarrollo, que parece usarse mucho menos, es el control del crecimiento y la conexión de las neuritas con sus órganos terminales apropiados de forma tan ajustada que haya poco espacio para el error desde el comienzo. A causa de la naturaleza global del desarrollo de los mamíferos, un control tan fino del desarrollo despojaría al embrión de la flexibilidad que necesita para compensar las variaciones inducidas por su formación genética o por el ambiente en otros aspectos del desarrollo.
Los mecanismos por los cuales las estructuras inervadas de las dianas evitan la muerte de sus neuronas, apenas están comenzando a dilucidarse. Una hipótesis general es que las células diana liberan factores tróficos químicos que las neuritas captan, por lo general mediante la unión a receptores específicos. El factor trófico entonces mantiene el crecimiento de la neurita. Un ejemplo clásico de un factor trófico es el factor de crecimiento neural, que mantiene el crecimiento e impide la muerte de las neuronas sensitivas. Hay otras moléculas bien caracterizadas que son buenas candidatas para ser factores tróficos.
SISTEMA NERVIOSO AUTÓNOMO
El sistema nervioso autónomo es el componente del sistema nervioso periférico que asume muchas de las funciones involuntarias del organismo, como la actividad glandular y la motilidad del sistema digestivo, la frecuencia cardíaca, el tono vascular y la actividad de las glándulas sudoríparas. Se divide en dos partes principales, los sistemas nerviosos simpático y parasimpático. Los componentes del sistema nervioso simpático surgen de los niveles toracolumbares de la columna vertebral (T1 a L2), mientras que los del sistema nervioso parasimpático tienen un origen dual ampliamente separado de las regiones craneal y sacra. Ambos componentes del sistema nerviosos autónomo constan de dos tipos de neuronas: las preganglionares y las posganglionarias. Las segundas se derivan de la cresta neural (capítulo 11).
Sistema nervioso simpático
Las neuronas preganglionares del sistema nervioso simpático surgen del asta intermedia (columna eferente visceral) de la sustancia gris de la médula espinal. En niveles desde T1 hasta L2 sus axones mielinizados procedentes de la médula discurren en paralelo por las raíces ventrales con los axones motores que inervan la musculatura esquelética (figura 10_18). Poco después de que las raíces dorsal y ventral del nervio raquídeo se unan, los axones simpáticos preganglionares, que se derivan del neuroepitelio del tubo neural, abandonan el nervio raquídeo por el ramo comunicante blanco. A continuación entran en uno de una serie de ganglios simpáticos donde hacen sinapsis con las neuronas posganglionares derivadas de la cresta neural.
Los ganglios simpáticos, en su gran mayoría organizados en forma de dos cadenas que corren en situación ventrolateral con relación a los cuerpos vertebrales, están compuestos por células de la cresta neural que migran desde el tubo neural en proceso de cierre a lo largo de una vía especial (figura 11_4). Una vez que los neuroblastos simpáticos migratorios han alcanzado el lugar en que se forma la cadena ganglionar simpática, se extienden tanto en dirección craneal como caudal hasta que la longitud de las cadenas se aproxima a la que se observa en el adulto. Algunos de los neuroblastos simpáticos migran en dirección ventral más allá del nivel ganglionar para formar los ganglios colaterales (por ejemplo, los ganglios celíaco y mesentérico), que ocupan posiciones algo variables en el interior de las cavidades corporales. La médula suprarrenal puede considerarse en un sentido amplio como un ganglio simpático muy modificado.
Las neuronas simpáticas preganglionares en crecimiento pueden terminar ya sea en el interior de la cadena ganglionar o pasar de largo para terminar en ganglios simpáticos más distantes donde hacen sinapsis con los cuerpos celulares de los neuroblastos simpáticos posganglionares de segundo orden (figura 10_18). Los axones de algunos neuroblastos posganglionares, que no son mielinizados, dejan la cadena ganglionar como un grupo paralelo y vuelven a entrar al nervio raquídeo más cercano a través del ramo comunicante gris. Una vez allí, estos axones siguen creciendo hasta que alcanzan dianas periféricas apropiadas, como las glándulas sudoríparas, los músculos piloerectores y las paredes de los vasos sanguíneos. Los axones de otras neuronas simpáticas posganglionares dejan sus ganglios respectivos en forma de plexos de fibras nerviosas y crecen hacia otras dianas viscerales.
Sistema nervioso parasimpático
Aunque también muestra una organización preganglionar y posganglionar, el sistema nervioso parasimpático tiene una distribución bastante diferente de la del sistema simpático. Las neuronas preganglionares parasimpáticas se originan, al igual que las del sistema nervioso simpático, en la columna eferente visceral del sistema nervioso central. No obstante, los niveles de origen de estos neuroblastos están en el mesencéfalo y en el rombencéfalo (específicamente asociados con los pares craneales III, VII, IX y X) y en los segmentos sacros segundo a cuarto de la médula espinal en desarrollo. Los axones procedentes de estos neuroblastos preganglionares recorren grandes distancias antes de encontrarse con las neuronas posganglionares derivadas de la cresta neural. La característica de estas neuronas es que están englobadas en pequeños ganglios o plexos dispersos en las paredes de los órganos a los cuales inervan.
Los precursores de las neuronas posganglionares crestales a menudo emprenden grandes migraciones (por ejemplo, desde el rombencéfalo hasta localizaciones finales en la pared del intestino). Las propiedades migratorias de los precursores crestales de las neuronas parasimpáticos son impresionantes, pero esta población de células también experimenta un gran incremento hasta que el número final de neuronas entéricas se aproxima al número de neuronas existentes en la médula espinal. Cada vez hay más pruebas de que unos factores localizados en le pared del intestino estimulan la mitosis de las células de la cresta neural que migran hasta allí. Una asombrosa demostración de los poderes estimuladores del intestino es la capacidad que tienen segmentos de pared del intestino injertadas a lo largo de tubo neural para causar una gran expansión de la región del tubo neural que se encuentra más próxima al injerto (figura 10_19).
Diferenciación de las neuronas autónomas
En la diferenciación de las neuronas autónomas intervienen al menos dos pasos principales. El primero es la determinación de ciertas células migratorias de la cresta neural en neuronas autónomas en vez de hacerlo en otros posibles derivados crestales.
Las células de la cresta neural tienen la opción de convertirse en componentes del sistema simpático o del parasimpático o del parasimpático. Esto se demostró en aves mediante injertos heterotópicos de células crestales. Por ejemplo, cuando la cresta neural cefálica, que normalmente formaría neuronas parasimpáticos, se injertó entre somitas 18 y 24, las células injertadas migraron y se establecieron en la médula suprarrenal como células cromafines, que normalmente son derivados del sistema nervioso simpático. Por el contrario, las células de la cresta neural del tronco injertadas en la región cefálica a menudo migraron hacia la pared del intestino y se convirtieron en neuronas parasimpáticas. posganglionares.
Un segundo gran paso en la diferenciación de las neuronas autónomas implica la elección del neurotransmisor que la neurona habrá de usar. Es característico que las neuronas parasimpáticas posganglionares sean colinérgicas (es decir, que usen acetilcolina como neurotransmisor), mientras que las neuronas simpáticas sean adrenérgicas (noradrenérgicas), y usen norepinefrina como neurotransmisor.
Conforme legan a su destino final, las neuronas autonómicas son noradrenérgicas. Entonces entran en una fase durante la cual seleccionan la sustancia neurotransmisora que se caracterizará su estado maduro. Hay evidencias experimentales considerables que sugieren que la elección del neurotransmisor procede con independencia de otros sucesos concomitantes, como la elongación de los axones y la inervación de órganos diana específicos.
En etapas muy tardías de su desarrollo, las neuronas autonómicas conservan su flexibilidad en la selección de un neurotransmisor. Por ejemplo, las neuronas simpáticas de las ratas recién nacidas normalmente son adrenérgicas, y si crecen en condiciones estándar de cultivo in vitro, producen grandes cantidades de norepinefrina e insignificantes de acetilcolina. Si las mismas neuronas se cultivan en un medio condicionado por la presencia de células miocárdicas, éstas experimentan una conversión funcional y a cambio producen grandes cantidades de acetilcolina (figura 10_20).
Un ejemplo de la transmisión natural del fenotipo de neurotransmisor de noradrenérgico a colinérgico se produce en la inervación simpática de las glándulas sudoríparas de la rata. Las transiciones de los neurotransmisores dependen de señales derivadas de la diana. Una de ellas es el factor de diferenciación colinérgica, una proteína básica glucosilada de 45kDa. Esta molécula, que está presente en el medio condicionado por cardiomiocitos, es uno de varios factores químicos ambientales que pueden ejercer una fuerte influencia en las fases tardías de la diferenciación de las neuronas autonómicas.
Megacolon agangliónico congénito (enfermedad de Hirschsprung)
Si un recién nacido muestra síntomas de estreñimiento completo sin que haya alguna obstrucción física demostrable, la causa suele ser la ausencia de ganglios parasimpáticos en el colon sigmoide y en el recto. Esta afección, que recibe el nombre de megacolon agangliónico congénito o enfermedad de Hirschsprung, por lo general se atribuye a la falta de colonización de la pared del colon inferior por precursores neuronales parasimpáticos derivados de la cresta neural, presumiblemente de origen sacro a causa de su distribución. En casos raros, grandes extensiones del colon pueden carecer de ganglios.
CAMBIOS ESTRUCTURALES MÁS TARDÍOS EN EL SISTEMA NERVIOSO CENTRAL
Histogénesis del sistema nervioso central
Una gran diferencia entre el encéfalo y la médula espinal es la organización de las sustancias blanca y gris. En la médula espinal la sustancia gris ocupa una situación central, y la blanca la rodea (figura 10_6). En muchas partes del cerebro, esta organización está invertida, con una zona de sustancia blanca y capas de sustancias gris situadas superficial a esta zona.
Uno de los procesos fundamentales en la histogénesis del encéfalo es la migración celular. Desde sus lugares de origen en las proximidades de los ventrículos cerebrales, los neuroblastos migran hacia la periferia siguiendo patrones preestablecidos. Estos patrones suelen dar como resultado la conformación en múltiples capas de la sustancia gris. Los protagonistas del fenómeno migratorio son las células de la glía radial, que se extiende hacia la periferia en largos procesos en forma radial a partir de los cuerpos celulares situados cerca de la luz de los ventrículos (figura 10_21). Las neuronas posmióticas jóvenes, que suelen ser células bipolares simples, se enroscan alrededor de las células de la glía radial y las usan como guías en su migración desde sus lugares de origen a la periferia.
En áreas de la corteza cerebral caracterizadas por múltiples capas de sustancia gris, las grandes neuronas que pueblan la capa más interna, migran primero. El resto de las capas de sustancia gris está formado por neuronas más pequeñas que migran a través de la primera capa y de otras capas que se han formado antes para establecer una nueva capa gris en la periferia. Con este patrón de histogénesis, el estrato más externo de neuronas es el último que se ha formado, y la capa más interna la que se formó primero. En un ratón mutante llamado weaver, se presentan unos defectos conductuales específicos relacionados con la función anómala del cerebelo. La base morfológica de este mutante es una anomalía de las células de la glía radial del cerebelo y una migración anormal consecuente de las células que normalmente forman el estrato granular de la corteza cerebral. Otro mutante, llamado reeler se caracteriza por un comportamiento anormal y por la ausencia de una corteza cortical normal. Recientemente se ha demostrado que una proteína, llamada reelina está ausente en el mutante reeler. Su función celular exacta no se ha determinado hasta ahora, pero puede servir como señal de “pare” para la migración neuronal radial o como señal de inserción para las neuronas migratorias.
Cada vez es más evidente que la corteza cerebral, en apariencia homogénea, es en realidad una matriz de unidades columnares radiales discretas que constan de células de la glía radial y de neuroblastos que migran a lo largo de ellas. Puede haber hasta 2000 millones de unidades columnares radiales en la corteza del cerebro humano. Las unidades radiales comienzan como unidades proliferativas, en las cuales la mayoría de las neuronas corticales se generan entre los días 40 y 125.
v Los cambios posteriores en el sistema nervioso central son tan amplios que un tratamiento exhaustivo de un solo aspecto, como la morfología, está mucho más allá del alcance de este libro. En su lugar, esta sección resalta aspectos fundamentales de la organización del sistema nervioso central y resume los cambios principales en la organización del encéfalo y la médula espinal.
Como sucede con muchos aspectos de la diferenciación neural, el número de unidades radiales parece ser sensible a su propio aporte neural. Por ejemplo, en el caso de la anoftalmía congénita (ausencia de ojos), la llegada de fibras nerviosas y neuroblastos visuales el área visual de la corteza occipital está reducida. El resultado son anomalías tanto macro como microscópicas de la corteza visual, relacionadas principalmente con la reducción del número de unidades radiales en esa región.
Médula espinal
En la médula espinal, los estímulos procedentes de muchos nervios periféricos sensitivos se realizan en forma de arcos reflejos locales o se canalizan hacia el encéfalo por tractos de axones. Así mismo, los mensajes motores que se originan en el encéfalo se distribuyen a localizaciones periféricas apropiadas a través de los tractos motores y de las raíces ventrales individuales de los nervios raquídeos. Si bien varios aspectos de la organización de la médula espinal ya se discutieron en este capítulo, algunos se revisarán brevemente por el valor que tienen para la comprensión de la organización básica del encéfalo.
La médula espinal en sus primeras etapas se divide en las regiones de las placas alar y basal, que son las precursoras de las regiones sensitivas y motoras de la médula (figura 10_6). La médula madura tiene una organización similar, pero esas regiones se subdividen aún más en componentes somáticos y viscerales. En el encéfalo se añade además otro nivel de entrada y salida de estímulos que tiene componentes “especiales”. Estos se resumen en el cuadro 10_1.
Un cambio macroscópico de la médula espinal y que reviste importancia clínica es el acortamiento relativo de la médula en relación con la columna vertebral (figura 10_22).
En el primer trimestre la médula se extiende a toda la longitud del cuerpo, y los nervios raquídeos pasan a través de los espacios invertebrales directamente situados en su lugar de origen. En los meses que siguen, el crecimiento de la parte caudal del cuerpo sobrepasa al de la columna vertebral y la médula espinal, pero el crecimiento de la médula se retrasa bastante con respecto al de la columna vertebral. Esta disparidad no es muy evidente en las regiones craneal y torácica, pero en el nacimiento la médula espina termina en el nivel de L3. Al llegar a la edad adulta termina en el nivel de L2.
La consecuencia de esta disparidad del crecimiento es la considerable elongación de las raíces nerviosas dorsales y ventrales lumbares y sacras para adaptarse al aumento de la distancia entre su punto de origen y el espacio invertebral correspondiente. Esto les da la apariencia colectiva de una cola de caballo (de aquí su nombre de cauda equina). Un filum terminale delgado, similar a un filamento, que se extiende desde el extremo caudal de la médula espinal hasta la base de la columna vertebral marca el desplazamiento original de la médula. Esta disposición es muy útil para el clínico porque el espacio que queda por debajo de la terminación de la médula es un lugar seguro para extraer líquido cefalorraquídeo para su análisis.
El mielencéfalo, la subdivisión más caudal del rombencéfalo (figura 10_2), se convierte en el bulbo raquídeo del encéfalo adulto (figuras 10_1 B y 10_23). En muchos aspectos es una estructura de transición situada entre el encéfalo y la médula espinal, y las similitudes existentes entre su organización funcional y la de la médula espinal son fáciles de reconocer (figura 10_24). Gran parte del bulbo sirve como vía de paso para los tractos que conectan el encéfalo con las zonas aferentes y eferentes de la médula espinal, pero también contiene centros encargados de la regulación de funciones vitales como el latido cardíaco y la respiración.
La organización fundamental de las placas alar y basal con un surco limitante intermedio se mantiene casi sin cambios en el mielencéfalo. El principal cambio topográfico desde la médula espinal es una marcada expansión de la placa del techo para formar el característico techo delgado que cubre al canal central que se ha expandido, y que en el mielencéfalo recibe el nombre de cuatro ventrículo (figura 10_35) (los detalles de los ventrículos y de las cubiertas del encéfalo y la médula se analizan más adelante en este capítulo).
Las columnas de núcleos (agregados de cuerpos celulares neuronales situados en el encéfalo) viscerales especiales aferentes (que van hacia el cerebro) y eferentes (que salen del cerebro) aparecen en el mielencéfalo como representaciones de las estructuras derivadas de los arcos branquiales.
Metencéfalo
El metencéfalo, la subdivisión más craneal del rombencéfalo, consta de dos partes principales: la protuberancia (o puente), que continúa con el bulbo, y el cerebelo, el componente más moderno del encéfalo desde el punto de vista filogenético y el de aparición más tardía desde el ontogenético (figura 10_23). La formación de estas estructuras depende del funcionamiento del gen engrailed_1 en la región mesencéfalo/rombencéfalo primitiva.
Como uno de sus nombres lo indica, la protuberancia sirve como un puente por el cual pasan los tractos de fibras nerviosas que comunican los centros superiores del encéfalo y la médula espinal. Su organización fundamental sigue siendo parecida a la del mielencéfalo, con tres grupos de núcleos aferentes y eferentes (figura 10_25). Además de éstos, en la sustancia blanca ventral están presentes otros núcleos protuberanciales especiales que se originan en los neuroblastos derivados de la placa alar. La parte caudal de la protuberancia también tiene una placa del techo ensanchada similar a la del mielencéfalo.
El cerebelo es complejo tanto en su estructura como en sus funciones, pero desde el punto de vista filogenético surgió como una especialización del sistema vestibular y se relacionaba con el equilibrio. Otras funciones, como la orquestación de la coordinación motora general y la participación en los reflejos auditivos y visuales, se le asignaron más tarde.
Inicialmente el cerebelo se sitúa en los labios rómbicos del embrión de cinco a seis semanas de edad (figura 10_26). Los labios rómbicos están localizados en el borde craneal del techo adelgazado, dentro del cual se proyectan parcialmente. Hasta el final del tercer mes la expansión de los labios rómbicos es interior, pero después el rápido aumento del volumen del cerebelo hace que se dirijan hacia fuera (figura 10_27).
Conforme el volumen del cerebelo en desarrollo se expande, los dos labios rómbicos laterales se unen en la línea media, dando al primordio del cerebelo un aspecto campaniforme. El cerebelo entonces entra en un período de rápido desarrollo y expansión externa. En su interior el cerebelo experimenta un complejo proceso de histogénesis (en general, el proceso subyacente a la organización celular del cerebelo puede apreciarse si se revisa la sección de la histogénesis de la corteza cerebral [figura 10_21], que continúa hasta bastante después del nacimiento. Muchas fibras que surgen del vasto número de neuronas generadas en el cerebelo salen de éste a través de un par de grandes pedúnculos cerebelosos superiores, los cuales se expanden hacia el interior del mesencéfalo.
Mesencéfalo
El mesencéfalo, o cerebro medio, es desde un punto de vista estructural una parte relativamente sencilla del encéfalo en la cual se conservan en esencia las relaciones fundamentales entre las placas basal y alar (figura 10_28). En la región de las placas alares, los neuroblastos migran hacia el techo (tectum), donde forman dos pares de abultamientos que reciben el nombre genérico de tubérculos cuadrigéminos. El par caudal, que constituye los llamados colículos inferiores, tiene una estructura relativamente sencilla y desde el punto de vista funcional forma parte del sistema auditivo. Los colículos superiores tienen una arquitectura estratificada más compleja a través de los patrones migratorios de los neuroblastos que les dan origen. Los colículos superiores forman parte integral del sistema visual, y sirven como una importante estación de relevo sináptico entre el nervio óptico y las áreas visuales de la corteza cerebral (occipital). Las conexiones entre los colículos superiores e inferiores ayudan a coordinar los reflejos visuales y auditivos.
Las placas basales mesencefálicas primitivas se convierten en el tegmento, la región menos definida desde el punto de vista estructural. En él están localizados los núcleos eferentes somáticos de los pares craneales III y IV, que inervan la mayor parte de los músculos extrínsecos del ojo. Un pequeño núcleo eferente visceral, el de Edinger_Westphal, es responsable de la inervación del músculo esfínter pupilar del ojo. Hay otros dos pares de núcleos voluminosos de sustancia gris, el núcleo rojo y la sustancia negra, cuyo origen aún se ignora.
La tercera región principal del mesencéfalo consiste en unos abultamientos ventrolaterales prominentes de sustancia blanca llamados pedúnculos cerebrales. Algunos de los principales fascículos descendentes pasan a través de estas estructuras desde los hemisferios cerebrales hacia la médula espinal.
Diencéfalo
Después del mesencéfalo en dirección craneal, la organización del encéfalo en desarrollo se modifica tanto que es difícil relacionar su morfología definitiva con le plan fundamental de la placa alar/placa basal. De hecho, es una idea ampliamente aceptada que las estructuras del prosencéfalo (Diencéfalo y telencéfalo) son derivados muy modificados de la placa alar y de la placa del techo sin una representación importante de la placa basal.
El desarrollo inicial del diencéfalo se caracteriza por la aparición de dos pares de engrosamientos prominentes en las paredes laterales del tercer ventrículo, los cuales se sitúan en la pared del canal central que en esta región esta muy desarrollado (figura 10_23). El par de masas más grandes corresponde al tálamo en desarrollo, en el cual los fascículos nerviosos procedentes de los centros cerebrales superiores hacen sinapsis con los de otras regiones del cerebro y el tronco cerebral. Entre muchos núcleos talámicos se encuentran los que reciben información de los sistemas auditivo visual y los que envían a las regiones correspondientes de la corteza cerebral. En el desarrollo ulterior los engrosamientos talámicos pueden llegar a fusionarse en la línea media a través del tercer ventrículo. Esta conexión se llama masa intermedia.
Ventral al tálamo, los engrosamientos del hipotálamo incipiente están separados del tálamo por el surco hipotalámico. Como ya se mencionó, el hipotálamo recibe información de muchas áreas del sistema nervioso central. Éste también actúa como el principal centro regulador, que controla muchas funciones homeostáticas básicas como el sueño, el control de la temperatura, el hambre , el equilibrio hidroelectrolítico, las emociones y los ritmos de las secreciones glandulares (por ejemplo, las de la hipófisis).
Varias de sus funciones son neurosecretoras, y por ello el hipotálamo sirve como zona de conexión entre la integración neural de la información sensitiva y el ambiente humoral del organismo.
En los embriones jóvenes (de unas siete a ocho semanas de edad), un par de abultamientos menos prominentes situados dorsales al tálamo forman el primordio del epitálamo (figura 10_23), un grupo de núcleos de desarrollo relativamente deficiente relacionados con las funciones de la masticación y la deglución. La parte más caudal de la placa del techo diencefálica forma una pequeña evaginación que se convierte en la epífisis (cuerpo pineal), una glándula antigua desde el punto de vista filogenético que suele comportarse como receptor luminoso. Bajo la influencia de los ciclos de luz y oscuridad, la glándula pineal segrega melatonina (principalmente por la noche), una hormona que inhibe la función del control hormonal del eje hipófisis-gónadas.
La hipófisis (glándula pituitaria) se desarrolla a partir de dos primordios ectodérmicos que en un comienzo están separados y que más tarde se unen. Uno de los primordios, llamado proceso infundibular, se forma por una proyección ventral del suelo diencefálico. El otro primordio, la bolsa de Rathke, una evaginación situada en la línea media derivada del ectodermo del estomodeo ya no crece hacia el suelo del diencéfalo en la cuarta semana. Un suceso inductivo procedente del diencéfalo suprayacente primero estimula la formación de un primordio de la bolsa de Rathke en el ectodermo del estomodeo. A través de la acción del gen Rpx (gen “bolsa de Rathke con homeobox”) y de los genes portadores de homeobox del tipo Lim (Lhx_3 y Lhx_4), el primordio de la bolsa de Rathke termina por definirse (figura 10_29). Las evidencias experimentales procedentes de varias especies animales indican que en el embrión inicial las células de la bolsa de Rathke se originan en la cresta anterior de la placa neural.
El proceso infundibular está estrechamente relacionado con el hipotálamo (figura 1_14), y ciertas neuronas neurosecretoras hipotálamicas envían sus prolongaciones hacia el interior del proceso infundibular, que termina por convertirse en el lóbulo posterior o neurohipófisis.
Durante todo el desarrollo, la estructura histológica del infundíbulo conserva un carácter neural.
A medida que avanza el desarrollo, la bolsa de Rathke se alarga hacia el infundíbulo (figura 10_29). Mientras que su extremo ciego envuelve parcialmente al infundíbulo como de una copa de dos capas, el tallo de la bolsa de Rathke comienza a regresar. La pared externa de la copa se engruesa y adquiere un aspecto glandular en el curso de su diferenciación en la parte distal (lóbulo anterior) de la hipófisis. La capa de la copa, que está íntimamente adherida al lóbulo posterior, se convierte en la parte intermedia. Permanece separada del lóbulo anterior por una hendidura o luz residual, que representa el vestigio de la luz original de la bolsa de Rathke.
Al avanzar el embarazo, la hipófisis experimenta una fase de diferenciación celular. Más adelante en el periodo fetal, los tipos celulares específicos comienzan a producir pequeñas cantidades de hormonas. Las cascadas moleculares subyacentes en la diferenciación de los tipos celulares específicos de la hipófisis están comenzando a descubrirse (cuadro 10_2).
Aunque la bolsa de Rathke casi siempre comienza a perder sus conexiones con el epitelio estomodeo hacia el final del segundo mes, algunas porciones de tejido pueden persistir en ocasiones a lo largo del trayecto de la elongación del tallo. Si el tejido es normal, se denomina hipófisis faríngea. No obstante, a veces el tejido residual se convierte en una neoplasia y forma tumores productores de hormonas llamados craneofaringiomas.
Las cúpulas ópticas son grandes evaginaciones de la red del diencéfalo que aparecen durante la embriogénesis inicial (ellas y los nervios ópticos (II par craneal) se discuten el capítulo 12).
Telencéfalo
El desarrollo del telencéfalo está dominado por la tremenda expansión de las vesículas telencefálicas bilaterales que terminarán por convertirse en los hemisferios cerebrales (figura 10_23). Las paredes de las vesículas telencefálicas rodean a los ventrículos laterales expandidos, que surgen como evaginaciones procedentes del tercer ventrículo que está en la línea media del diencéfalo (figura 10_35). Aunque los hemisferios cerebrales aparecen primero como estructuras laterales, la dinámica de sus crecimiento hace que se aproximen a la línea media por encima del techo del diencéfalo y el mesencéfalo (figura 10_30). Los dos hemisferios cerebrales jamás se fusionan en la línea media dorsal porque están separados por un delgado tabique de tejido conectivo (parte de la duramadre) conocido como la hoz del cerebro. Por debajo de este tabique los dos hemisferios cerebrales están unidos por el techo del epéndimo del tercer ventrículo.
Aunque los hemisferios cerebrales experimentan una gran expansión durante los primeros meses del embarazo, sus superficies externas permanecen lisas hasta la semana 14. Conforme siguen creciendo, los hemisferios cerebrales comienzan a mostrar surcos y pliegue en varias zonas. El plegamiento y la expansión más importante lo realizarán los futuros lóbulos temporales, que se proyectan lateral y rostralmente desde la parte caudal de los hemisferios cerebrales. Entre el cuarto y noveno meses de la gestación, la expansión de los lóbulos temporales, frontales y parietales cubre por completo las áreas de la corteza cerebral conocidas como la ínsula (figura 10_31). Mientras que estos grandes cambios precursores de los principales detalles superficiales de la corteza cerebral definitiva. Varios de los surcos y las fisuras principales comienzan a aparecer ya desde el sexto mes. Hacia el octavo mes los surcos y las circunvoluciones que caracterizan al cerebro maduro toman forma.
En el interior, la base de cada vesícula telencefálica se engruesa para dar lugar al cuerpo estriado (figura 10_32) que tiene forma de coma. El cuerpo estriado, dorsal al tálamo adopta la forma de una C conforme avanza el desarrollo. Con la histodiferenciación de la corteza cerebral, muchos fascículos nerviosos convergen hacia la región del cuerpo estriado, al que se subdivide en dos núcleos principales: lenticular y caudado.
Estas estructuras, que son componentes de la compleja agrupación de núcleos conocidos como los ganglios basales, intervienen en el control inconsciente del tono muscular y en movimientos corporales complejos.
Aparte de las vesículas telencefálicas, el otro componente principal del telencéfalo inicial es la lámina terminal, que forma su pared rostral media (figura 10_33 y 10_35). Al comienzo los dos hemisferios cerebrales se forman por separado, pero hacia el final del primer trimestre de la gestación, unos haces de fibras nerviosas comienzan a cruzar de un hemisferio cerebral al otro. Muchas de estas conexiones se establecen a través de la lámina Terminal.
El primer grupo de conexiones que aparece en la lámina Terminal se convierte en la comisura anterior (figura 10_23) B), que conecta las áreas olfatorias de ambos lados del cerebro. La segunda conexión es la comisura del hipocampo (fórnix). La tercera es el cuerpo calloso, la conexión más importante entre las mitades derecha e izquierda del cerebro. Al comienzo forma un pequeño haz en la lámina Terminal pero experimenta una gran expansión y termina formando una amplia banda que conecta un gran parte de la base de los hemisferios cerebrales (figura 10_33).
Otras comisuras que no se relacionan con la lámina Terminal son las comisuras habenulares anterior y posterior (figura 10_30), que están localizadas cerca de la base de la glándula pineal, y el quiasma óptico, la región del diencéfalo donde parte de las fibras del nervio óptico cruzan hacia el otro lado del cerebro.
Los neuroanatomistas subdividen el encéfalo en varios componentes funcionales con base en el desarrollo filogenético de la región. El componente más antiguo y primitivo se denomina rinencéfalo (también archicortéx y paleocórtex). Como su nombre indica, interviene en la olfacción. Desde el punto de vista morfológico el llamado neocórtex es el elemento dominante de los hemisferios cerebrales. En el desarrollo inicial, gran parte del telencéfalo está ocupado por áreas rinencefálicas (figura 10_34), pero con la expansión de los hemisferios cerebrales, el neocórtex se convierte en el componente que ocupa la mayor parte del cerebro.
Los nervios olfatorios (I par craneal), que surgen de plácodas ectodérmicas bilaterales situadas en le cabeza, envían fibras hacia atrás, a los bulbos olfatorios, que son evaginaciones procedentes del rinencéfalo. Una subpoblación de células procedentes de la plácoda olfatoria migra a lo largo del nervio olfatorio hacia el cerebro, y termina estableciéndose en el hipotálamo, donde se convierte en las células que segregan la hormona luteinizante.
VENTRÍCULO, MENINGES Y FORMACIÓN DEL LÍQUIDO CEFALORRAQUÍDEO
El sistema ventricular del cerebro representa una expansión del canal central del tubo neural. Conforme ciertas partes del cerebro adquieren su forma, el canal central se expande y se convierte en cavidades bien definidas llamadas ventrículos unidos mediante conductos estrechos (figura 10_35). Los ventrículos están tapizados por el epitelio ependimario y llenos de un líquido cefalorraquídeo claro. Este líquido se forma en áreas especializadas denominadas plexos coroideos, localizados en regiones específicas del techo de los ventrículos tercero, cuarto y laterales. Los plexos coroideos son estructuras muy vascularizadas que se proyectan hacia el interior de los ventrículos (figura 10_30) y segregan el líquido cefalorraquídeo.
El líquido cefalorraquídeo tiene una circulación característica. Apenas formando, fluye desde los ventrículos laterales hacia el tercero y por último hacia el cuarto ventrículo. Gran parte de él escapa luego a través de tres pequeños orificios situados en el techo del cuarto ventrículo y entra al espacio subaracnoideo situado entre dos capas meníngeas. Una parte importante del líquido deja el cráneo y baña la médula espinal en forma de una capa protectora.
Si se presenta un desequilibrio entre la producción y la reabsorción del líquido cefalorraquídeo o si su circulación se bloquea, el líquido puede acumularse en el sistema ventricular del cerebro y causa incremento de la presión hidrostática que puede producir un gran aumento de tamaño del sistema ventricular.
Esto a su vez ocasiona un adelgazamiento de las paredes del cerebro y un marcado incremento de la circunferencia cefálica; este padecimiento se conoce como hidrocefalia (figura 10_36). El bloqueo del líquido puede tener su origen en una estenosis (estrechamiento) congénita de las partes más estrechadas del sistema ventricular, o puede ser el resultado de ciertas infecciones virales que afectan al feto.
Una malformación específica que conduce a la hidrocefalia es la malformación de Arnold-Chiari, en la cual una evaginación del cerebelo en forma de lengüeta se hernia a través del foramen e impide mecánicamente la salida del líquido cefalorraquídeo del cráneo. Este padecimiento suele estar asociado con alguna variedad de defecto de cierre de la médula espinal o de la columna vertebral (figura 10_40). En una época se consideraba que la malformación de Arnold-Chiari provenía de una tracción de la médula espinal que tiraba del cerebelo a través del foramen magno, pero una hipótesis más moderna sostiene que el volumen reducido de una fosa posterior, más pequeña de lo normal, causa un desplazamiento caudal del rombencéfalo.
Al comienzo del período fetal aparecen dos capas de mesénquima alrededor del cerebro y la médula espinal. La capa gruesa externa, que es de origen mesodérmico, forma la resistente duramadre así como los huesos membranosos del cráneo. Una capa interna delgada originada en la cresta neural se divide más tarde en la piamadre, una capa delgada en estrecha aposición con el tejido neural, y la capa media, la aracnoides. Los espacios que se crean en el interior de la capa formada por la pía y la aracnoides se llenan de líquido cefalorraquídeo.
PARES CRANEALES
Aunque se basan en el mismo plan fundamental que los nervios raquídeos, los pares craneales (figura 10_37) han perdido su organización segmentaria regular y se han especializado (tabla10_2). Una de las principales diferencias es la tendencia de muchos de los pares craneales a ser sensitivos (basados en las raíces dorsales) o motores (basados en las raíces ventrales) y no mixtos, como es el caso de los nervios raquídeos.
Los pares craneales pueden subdividirse en varias categorías con base en su función y su origen embriológico. Los pares craneales I y II (olfatorio y óptico) suelen considerarse como evaginaciones cerebrales y no como verdaderos nervios. Los pares craneales III, IV, VI y XII son nervios motores puros que parecen haber evolucionado a partir de raíces ventrales primitivas. Los pares V, VII, IX y X son nervios mixtos con componentes sensitivos y motores, y cada uno inerva los derivados de un arco branquial diferente (tabla 10_2 y figuras 10_38 y 13_21).
Los componentes sensitivos de los nervios que inervan los arcos branquiales (V, VII, IX y X) y el nervio auditivo (VIII) tienen un origen múltiple tanto en la cresta neural como en las plácodas ectodérmicas, localizadas a lo largo del encéfalo en desarrollo (figura 5_14). Estos nervios tienen ganglios sensitivos complejos, a menudo múltiples. Algunas neuronas de los ganglios tienen su origen en la cresta neural, y otras neuronas provienen del ectodermo de las plácodas (las plácodas ectodérmicas se estudian en la pág. 90).
DESARROLLO DE LA FUNCIÓN NEURAL
Durante las primeras cinco semanas del desarrollo embrionario no hay evidencias de funciones neurales. La actividad refleja primitiva puede provocarse por primera vez hacia la sexta semana, cuando al tocar la piel que rodea la boca con un fino pelillo se produce una flexión contralateral del cuello. En las siguientes seis a ocho semanas, la región de la piel que es sensible a la estimulación táctil se extiende desde la cara a las palmas de las manos y la región superior del tórax; hacia las doce semanas toda la superficie del cuerpo, excepto la espalda y la coronilla, es sensible. Conforme estas áreas de sensibilidad se amplían, la naturaleza de los reflejos provocados, incluye desde movimientos generalizados hasta respuestas específicas de partes más localizadas del cuerpo.
Existe una secuencia craneocaudal de aparición de movimientos reflejos.
Desde el final del cuarto mes el feto empieza con un patrón de períodos de actividad seguidos por otros de inactividad. Muchas mujeres perciben por primera vez los movimientos fetales por esta época. Entre el cuarto y el quinto meses, el feto desarrollo la capacidad de asir con fuerza una varilla de vidrio. Aunque son posibles algunos débiles movimientos protorrespiratorios, no son sostenidos. El reflejo de succión aparece durante el sexto mes. Algunos cambios detectables en los patrones de las ondas cerebrales se observan alrededor de las 28 semanas; también en este momento algunos bebés prematuros pueden mantener la respiración y sobrevivir dentro de una incubadora.
Gran parte del desarrollo del comportamiento de los fetos después del sexto mes se ha deducido por la observación del comportamiento de los bebés prematuros. Los cambios de conducta que se observan en el último trimestre son más sutiles y suelen reflejar no sólo que los circuitos neuronales se han establecido y terminado, sino también su maduración estructural y funcional.
El desarrollo de los circuitos funcionales puede ilustrarse en la médula espinal. Es posible identificar varias etapas de maduración estructural y funcional (figura 10_44). La primera es una etapa refleja, que se caracteriza por la diferenciación inicial (incluye el crecimiento de los axones y las dendritas) de las neuronas de acuerdo con una secuencia bien definida que comienza con las motoras y termina con las sensitivas y que por último incluye las interneuronas que conectan a las primeras con las segundas (figuras 10_13 y 10_44 A). La segunda etapa consiste en el cierre del circuito primario, lo cual permite la expresión de reflejos segmentarios locales. Mientras que el circuito local se está estableciendo, otros axones están creciendo a través de fascículos descendentes en la médula o están cruzando desde el otro lado de la médula.
Cuando estos axones hacen contacto con los componentes del reflejo simple que se estableció en la segunda etapa, ha quedado establecida la base anatómica para los reflejos intersegmentarios y cruzados de la médula. Más adelante en el períodos fetal, estos circuitos más complejos se completan y los oligodendrocitos mielinizan a los fascículos.
La maduración funcional de los fascículos individuales, según lo indica su mielinización, tiene lugar durante un período amplio y no termina antes de que comience la edad adulta. En especial en la vida posnatal inicial, la maduración de los tractos funcionales en el sistema nervioso puede seguirse con el examen neurológico clínico.
La mielinización comienza en el sistema nervioso periférico, en el cual las raíces motoras se mielinizan antes que las sensitivas (lo que sucede a lo largo de los meses segundo hasta el quinto). La mielinización comienza en la médula espinal aproximadamente a las once semanas y avanza siguiendo un gradiente craneocaudal. Durante el tercer trimestre, este proceso comienza en el cerebro, pero allí, a diferencia de lo que sucede en el sistema nervioso periférico, la mielinización se aprecia primero en los fascículos sensitivos (por ejemplo, el sistema visual). La mielinización de los fascículos de asociación situados en la corteza cerebral se produce después del nacimiento. En los fascículos corticoespinales, la principal conexión directa entre la corteza cerebral y los nervios motores que emanan de la médula espinal hacia las 40 semanas, la mielinización se extiende en sentido caudal sólo hasta el nivel del bulbo raquídeo. La mielinización continúa después del parto, y su curso puede apreciarse en virtud de la movilidad creciente de los bebés durante su primer año de vida.
CORRELACION CLÍNICA 10_1
Malformaciones congénitas del sistema nervioso
En un sistema orgánico tan importante y complejo como el sistema nervioso, no es sorprendente que el cerebro y la médula espinal estén sujetos a una amplia variedad de malformaciones congénitas. Éstas van desde anomalías estructurales graves originadas por el cierre incompleto del tubo neural hasta alteraciones funcionales causadas por factores desconocidos que actúan en las fases tardías del embarazo.
DEFECTOS DE CIERRE DEL TUBO NEURAL
La falta de cierre del tubo neural tiene lugar casi siempre en los neuroporos anterior y posterior, pero también es posible en otras localizaciones. En esta afección la médula espinal o el cerebro en el área afectada están extendidos y abiertos y la pared del canal central o el sistema ventricular constituye su superficie externa. Varios defectos de cierre pueden diagnosticarse mediante la detección de niveles elevados de alfafetoproteína en el líquido amniótico o mediante ecografía. Los defectos de cierre de la médula espinal, raquisquisis, y los defectos de cierre del cerebro se llaman craneosquisis. Ésta última es incompatible con la vida. La raquisquisis (figuras 10_39 y 10_40 A) se asocia con una amplia variedad de problemas graves, entre los que se incluyen infección crónica, déficit motores y sensitivos y trastornos de la función vesical. Estos defectos suelen acompañar a la anencefalia (figura 7_4), en la cual una deficiencia casi total de estructuras craneales.
OTROS DEFECTOS DE CIERRE
Un defecto en la formación de las estructuras óseas que cubren ya se la médula espinal o el cerebro pueden tener como resultado un amplio espectro de anomalías estructurales. En la columna vertebral el defecto más sencillo se llama espina bífida oculta (figuras 10_40 B), en la cual la médula ósea (arco neural) de una o más vértebras está incompleta.
En ocasiones el defecto pasa desapercibido durante muchos años. Los arcos neurales vertebrales son inducidos por la placa del techo del tubo neural, con la mediación del Msx_2. De este modo, es probable que la espina bífida oculta provenga de un defecto local en la inducción. El lugar del defecto en los arcos neurales de las vértebras suelen estar indicando por un mechón de pelo. Esta formación localizada de pelo puede provenir de la exposición de la piel en desarrollo a otras influencias inductivas del tubo neural o de sus cubiertas . normalmente los arcos actúan como la barrera contra esas influencias.
La siguiente categoría más grave de defectos es el meningocele, en el cual puede faltar la duramadre en el área del defecto y la aracnoides sobresale de forma evidente por debajo de la piel (figura 10_40). No obstante, la médula espinal ocupa su lugar y los síntomas neurológicos suelen ser menores. La afección más grave es el mielomeningocele, en le cual la médula espinal sobresale o está completamente herniada dentro de un abultado espacio subaracnoideo (figuras 10_40 D y 10_41. A causa de los problemas asociados con él desplazamiento de las raíces nerviosas, esta afección suele asociarse con trastornos neurológicos.
Un espectro similar de anomalías se asocia con los defectos craneales (figuras 10_42 y 10_43). El meningocele suele asociarse con un defecto pequeño en el cráneo, en tanto que el tejido cerebral solo (meningoencefalocele) o tejido cerebral que contiene parte del sistema ventricular (meningohidroencefalocele) puede sobresalir a través en el cráneo. Dependiendo de la naturaleza del tejido afectado, estas malformaciones pueden asociarse con déficit neurológicos. Las circunstancia mecánicas también pueden llevar a hidrocefalia secundaria en algunos casos.
La microcefalia es una afección poco común que se caracteriza por el subdesarrollo tanto del cerebro como de la calota craneal (figura 8_9). Aunque puede provenir del cierre prematuro de las suturas del cráneo, en la mayoría de los casos su etiología no está clara.
Muchos de los defectos funcionales del sistema nervioso se han caracterizado de forma bastante deficiente, y su etiología no es clara.
Los estudios realizados en ratones con defectos del movimiento o del comportamiento con base genética y causados por anomalías de la migración celular o de la histogénesis en ciertas regiones del cerebro, sugieren la posibilidad de que exista un espectro similar en los defectos de los seres humanos. Un buen ejemplo de ello es la lisencefalia, una afección en la cual la superficie del cerebro es lisa, en vez de tener las circunvoluciones y los surcos que caracterizan a la corteza normal. Las circunstancia subyacente en este defecto macroscópico es una anomalía en el establecimiento de las capas de las neuronas corticales de una forma que recuerda la patología que se observa en el ratón reeler (págs.232 y 233). El retraso mental es común y puede atribuirse a muchas causas, tanto genéticas como ambientales. El momento en que se produce la lesión en el cerebro puede ser más tardío, durante la vida fetal.
De manera característica, las neuronas simpáticas son adrenérgicas y las neuronas parasimpáticos son colinérgicas. No obstante, la elección normal del neurotransmisor puede ser modificada por factores ambientales de manera que una neurona simpática, por ejemplo, puede segregar acetilcolina.
CASO CLÍNICO
Un bebé nace con una raquisquisis en la columna lumbar. Durante las semanas siguientes la cabeza del niño también comienza a crecer. Los estudios radiológicos muestran que el sistema ventricular del bebé está muy dilatado y que las paredes del cerebro se han adelgazado.
Suponiendo que el bebé sobrevive,
¿Cuáles serán algunos de los problemas clínicos con que se enfrentará más adelante en su vida?.
PREGUNTAS DE REPASO
1. ¿Qué molécula producida por la notocorda es fundamental para la inducción de la placa del suelo del tubo neural?
A. Hoxa _5
B. Ácido retinoico
C. Pax_3
D. Msx_1
E. Sonic hedgehog
2. Los cuerpos celulares de la motoneuronas de los nervios raquídeos tienen su origen en:
A. Placa basal
B. Zona marginal
C. Placa del suelo
D. Placa del techo
E. Placa alar
3. Un joven paciente que tiene un mechón de pelo en la región lumbar de la columna vertebral se somete a una cirugía para la corrección de un defecto congénito en esa región. Durante la cirugía, se encontró que la duramadre y la aracnoides que cubren la médula espinal estaban íntegras pero que faltaban los arcos neurales de varias vértebras. ¿Cuál es el diagnostico de este paciente?
A. Meningocele
B. Meningomielocele
C. Encefalocele
D. Espina bífida oculta
E. Raquisquisis
4. Los conos de crecimiento se adhieren con firmeza a un sustrato que contiene:
A. Acetilcolina
B. Laminina
C. Epinefrina
D. Norepinefrina
E. Sonic hedgehog
5. El defecto complete de cierre del tubo neural en la región de la médula espinal se llama:
A. Espina bífida oculta
B. Meningocele
C. Craneosquisis
D. Raquisquisis
E. Mielomeningocele
6. Los rombómeros son divisiones segmentarias del:
A. Prosencéfalo
B. Mesencéfalo
C. Rombencéfalo
D. Médula espinal
E. Ninguna de las anteriores
7. ¿Durante qué mes de la gestación las mujeres embarazadas suelen percibir por primera vez los movimientos fetales?
A. Segundo
B. Tercero
C. Cuarto
D. Sexto
E. Octavo
8. La bolsa de Rathke surge de:
A. Diencéfalo
B. Ectodermo del estomodeo
C. Mesencéfalo
D. Endodermo faríngeo
E. Infundíbulo
9. En los primeros días después del nacimiento, un bebé no expulsa materia fecal y el abdomen comienza a distenderse. El orificio anal es permeable. ¿Cuál es el diagnóstico más probable?
10. ¿ Cuál e la posible morfología de la médula espinal y los nervios branquiales de un bebé que nace con ausencia congénita de un brazo (amelia)?
{kind=link}
{kind=link}
{kind=link}
{kind=link}
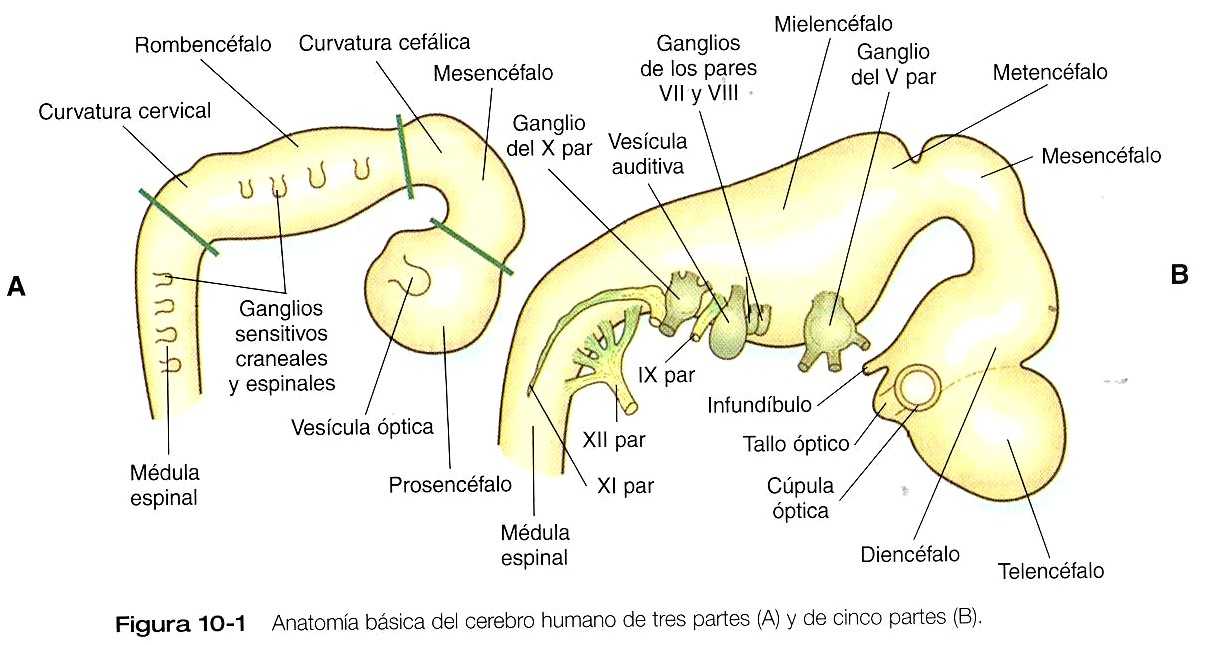{kind=link}
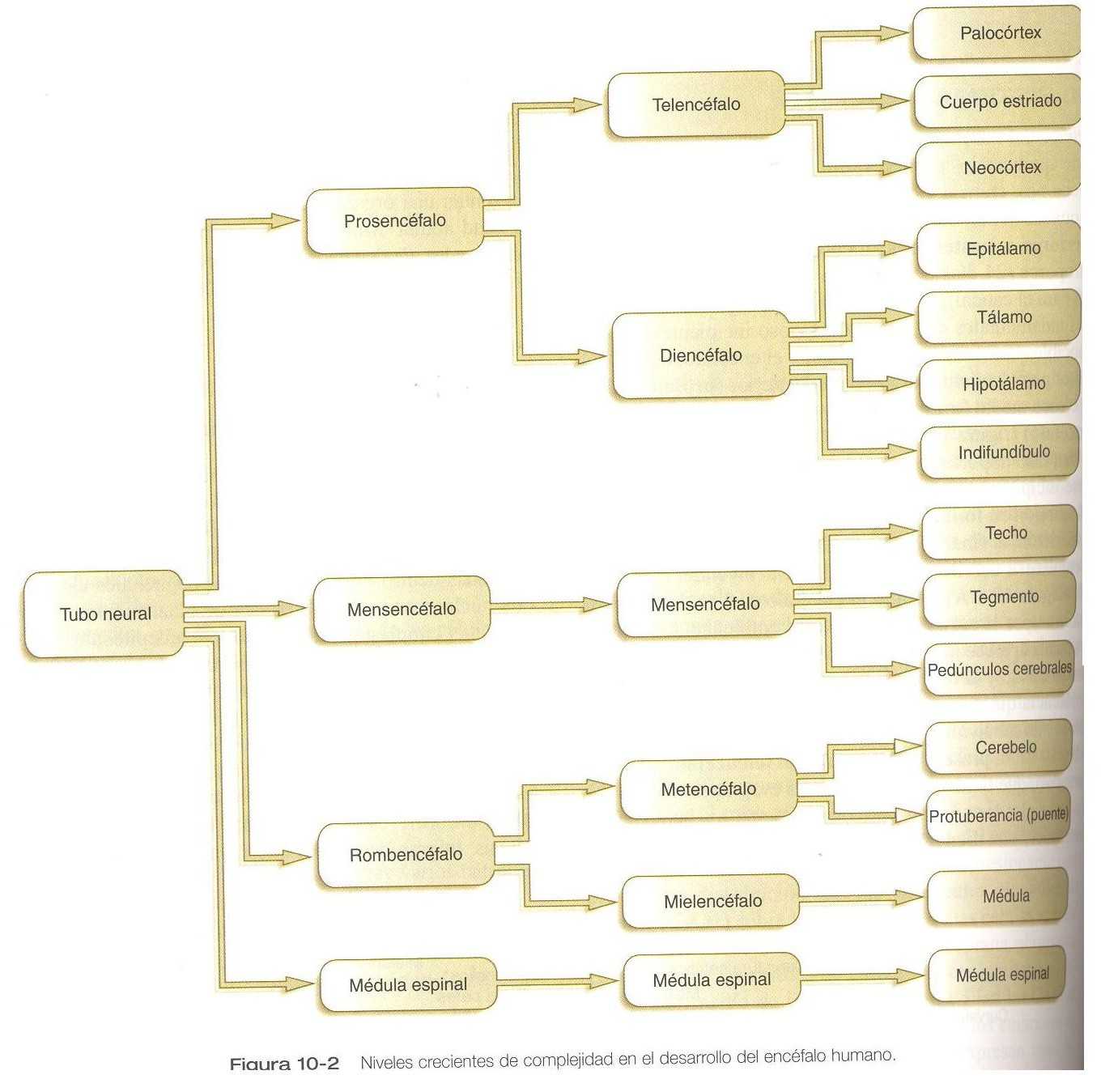{kind=link}
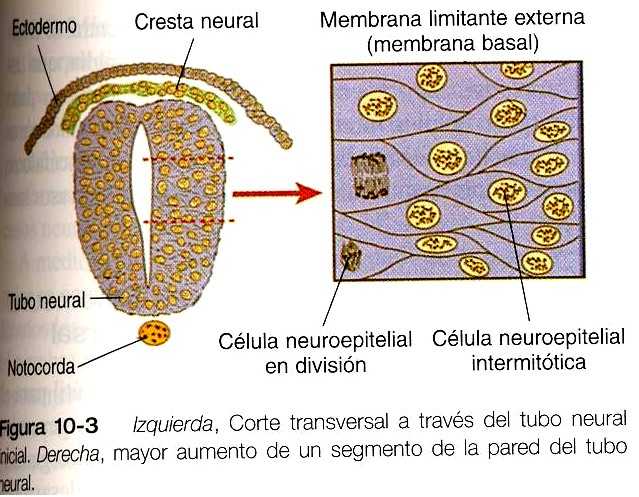{kind=link}
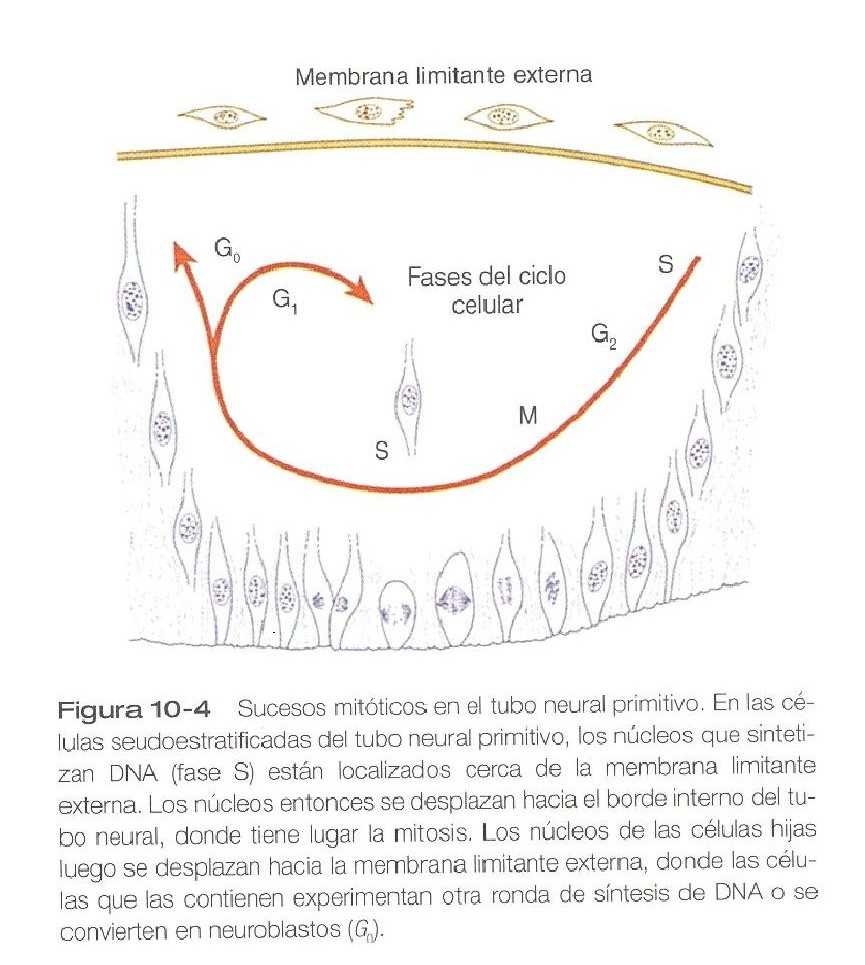{kind=link}
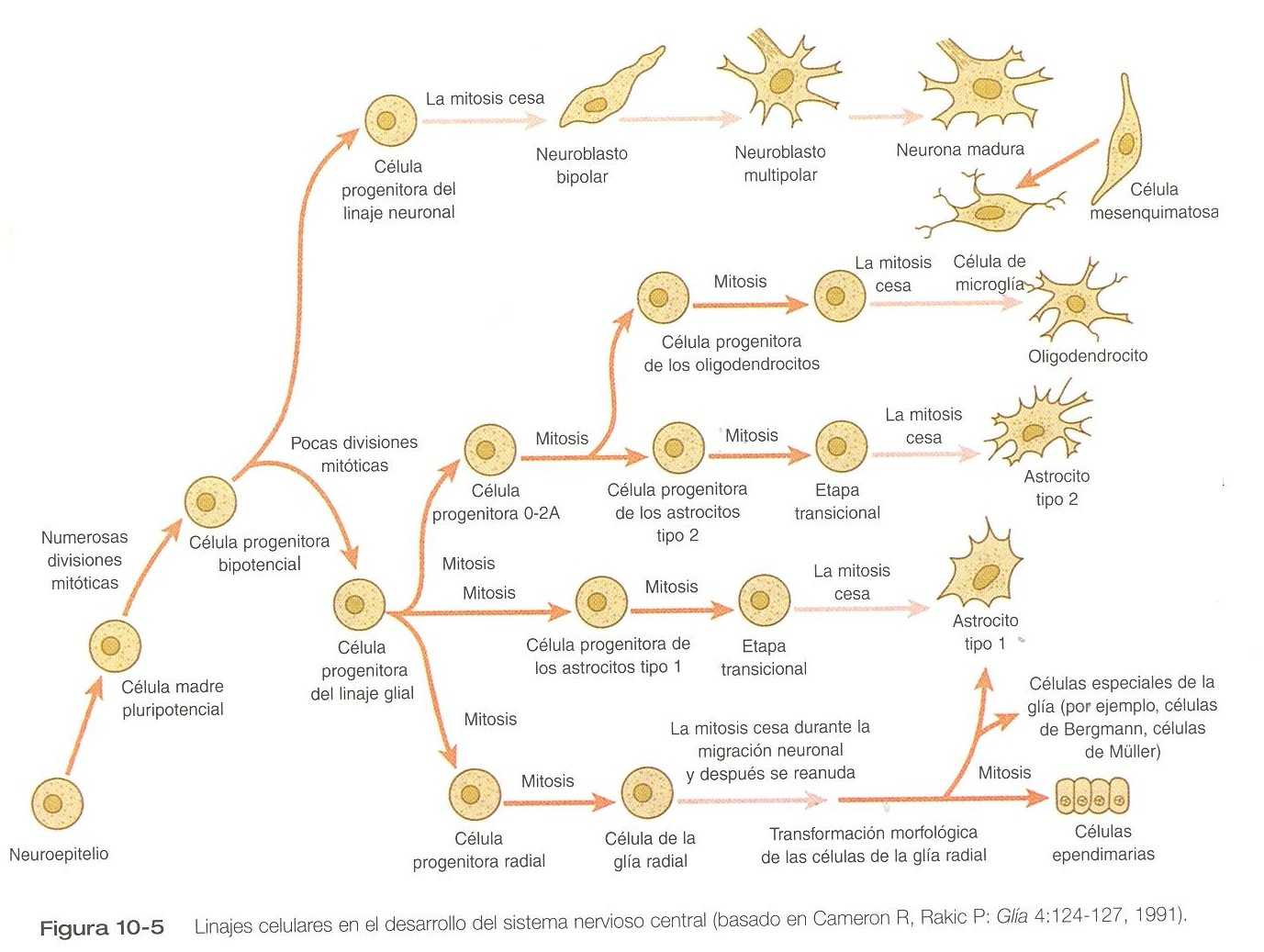{kind=link}
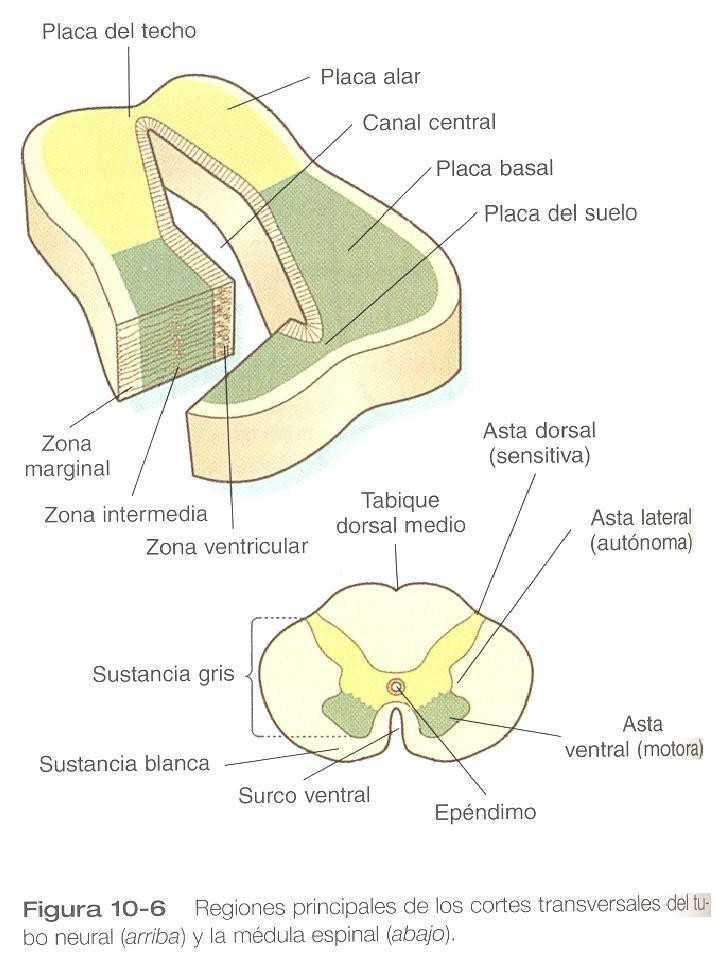{kind=link}
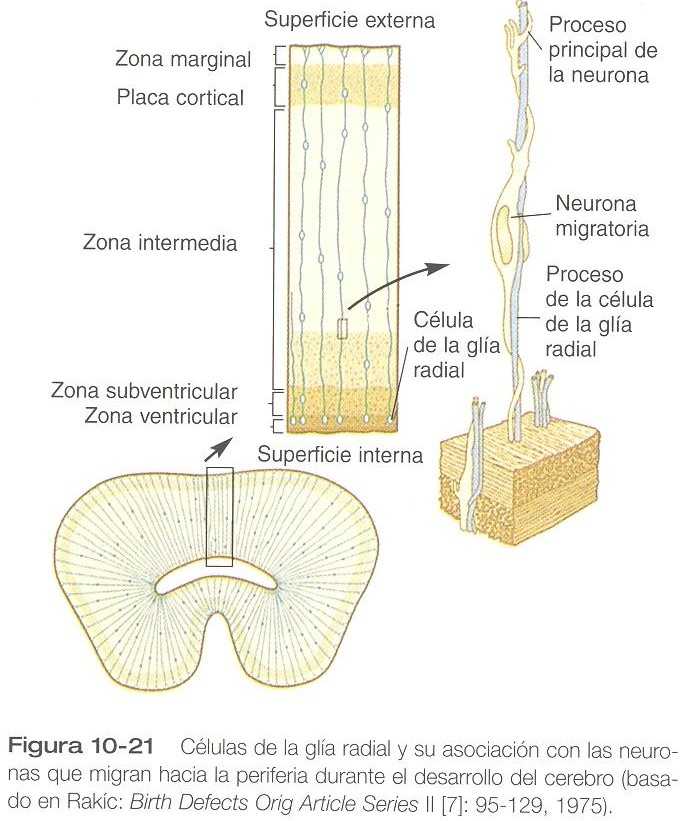{kind=link}
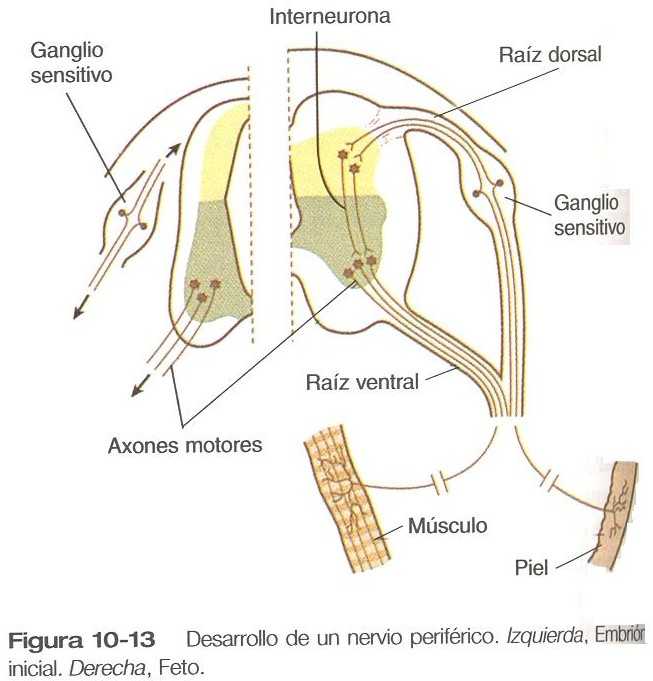{kind=link}
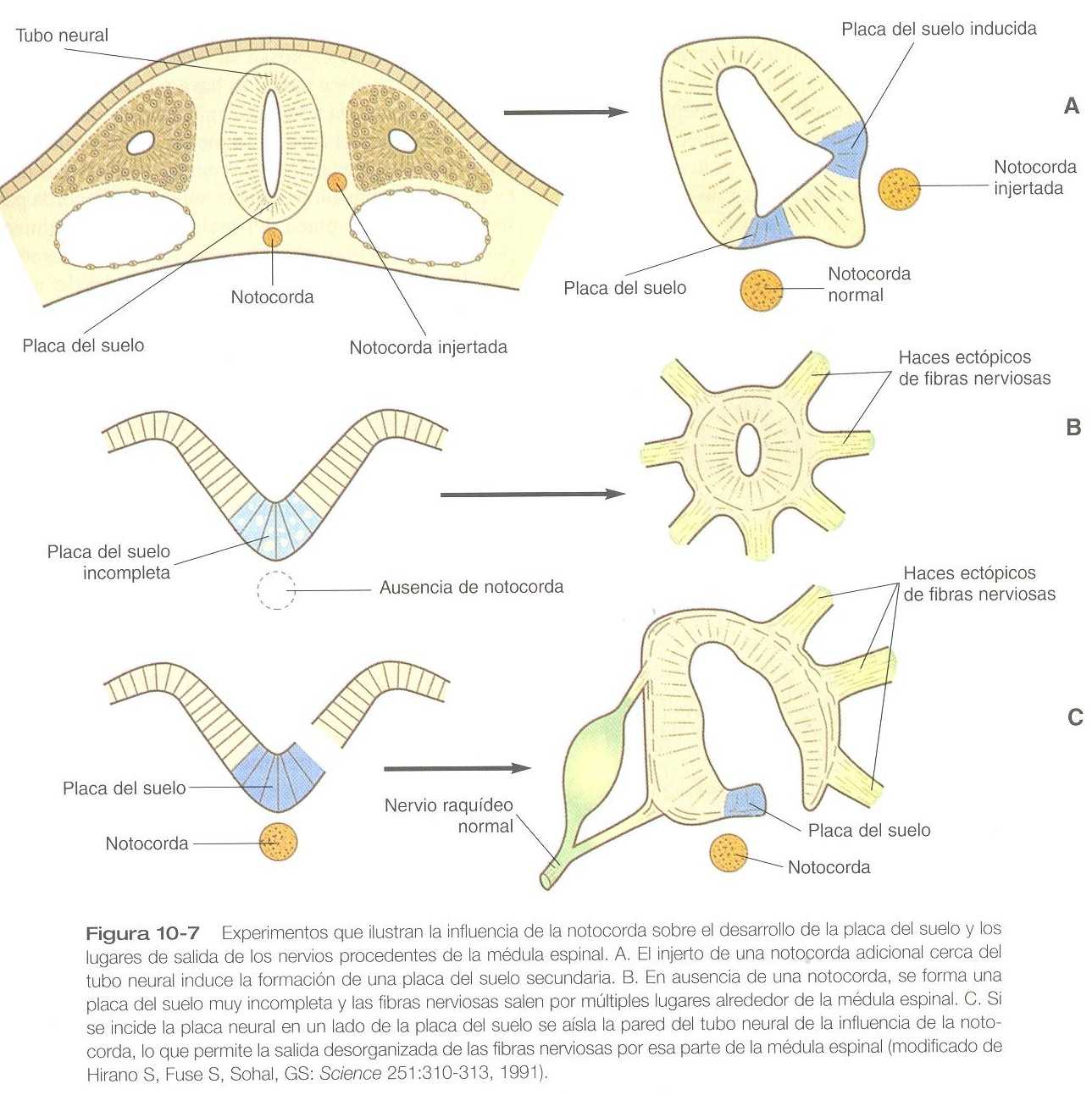{kind=link}
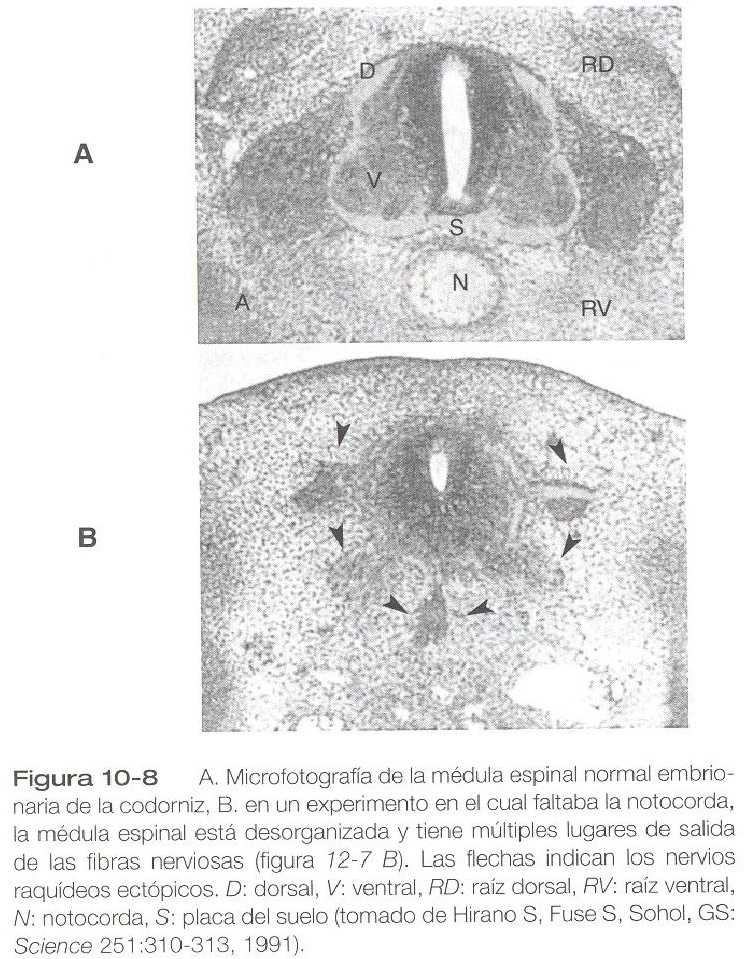{kind=link}
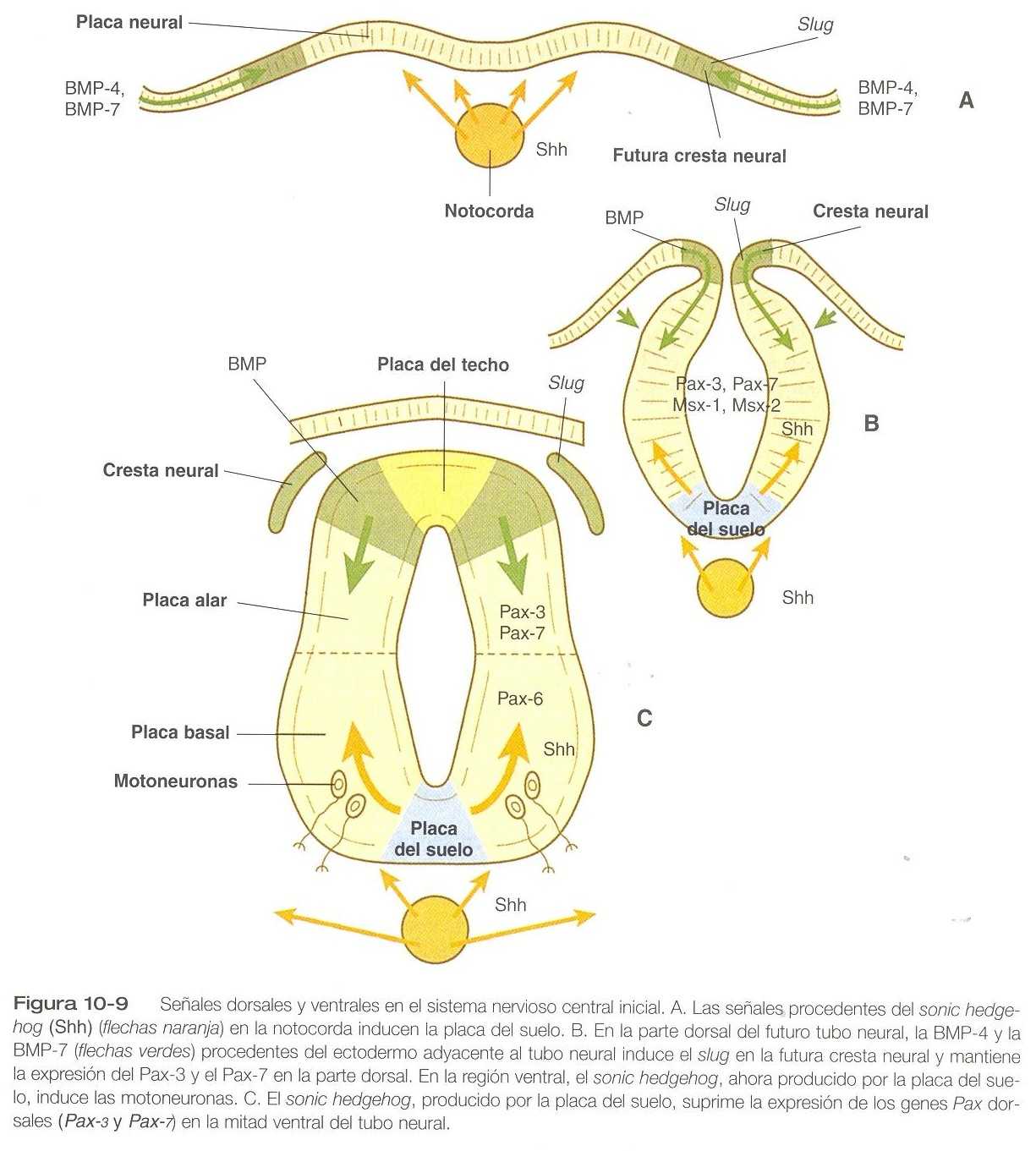{kind=link}
{kind=link}
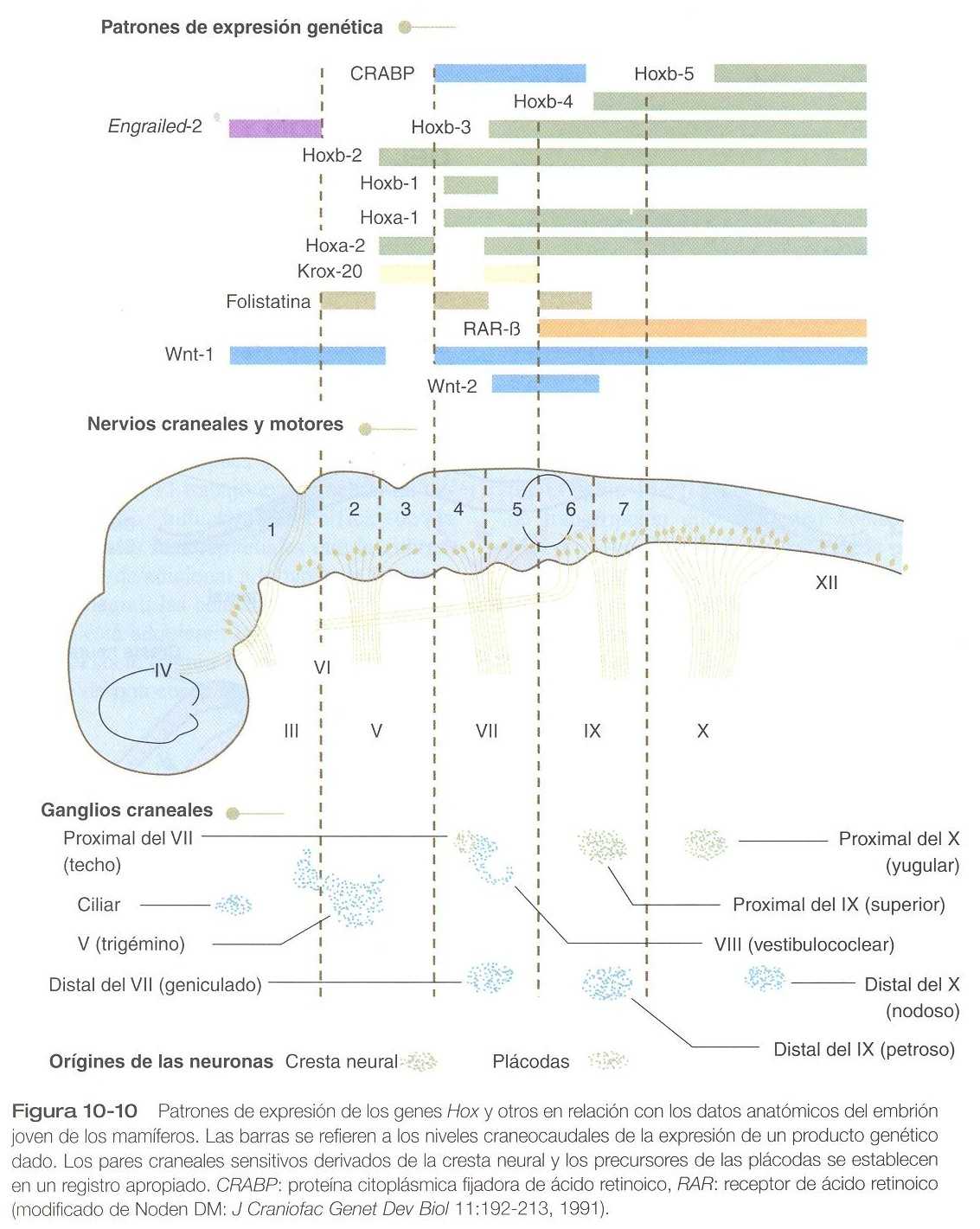{kind=link}
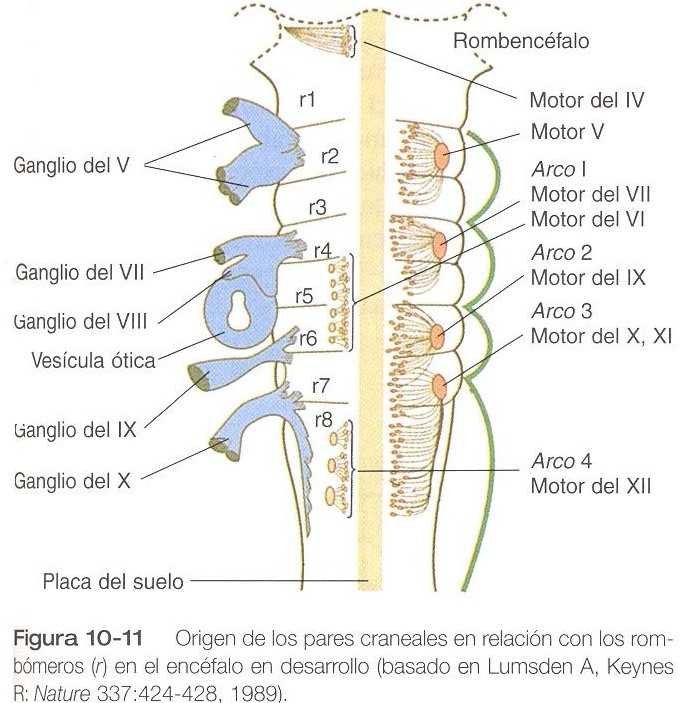{kind=link}
{kind=link}
{kind=link}
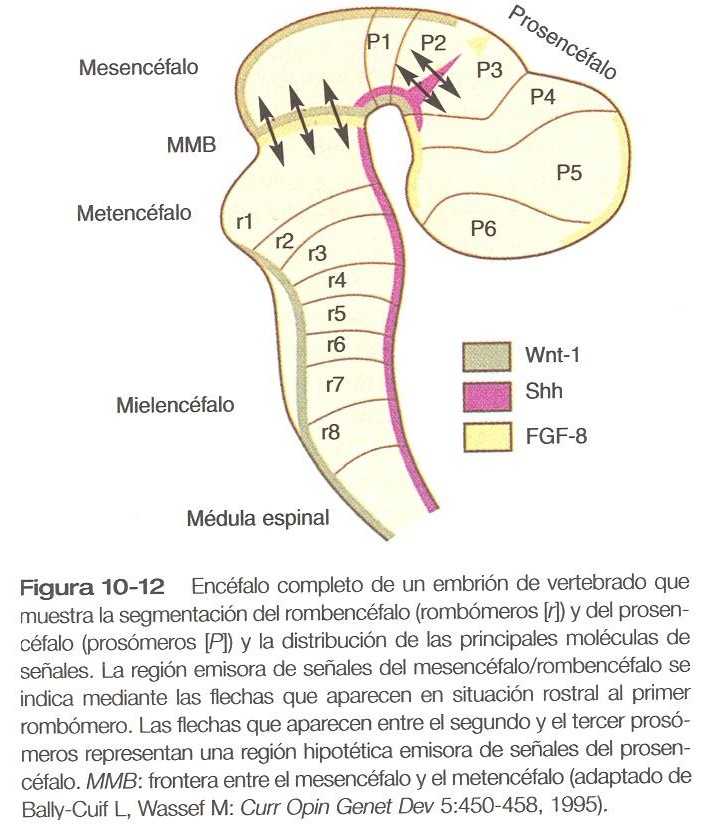{kind=link}
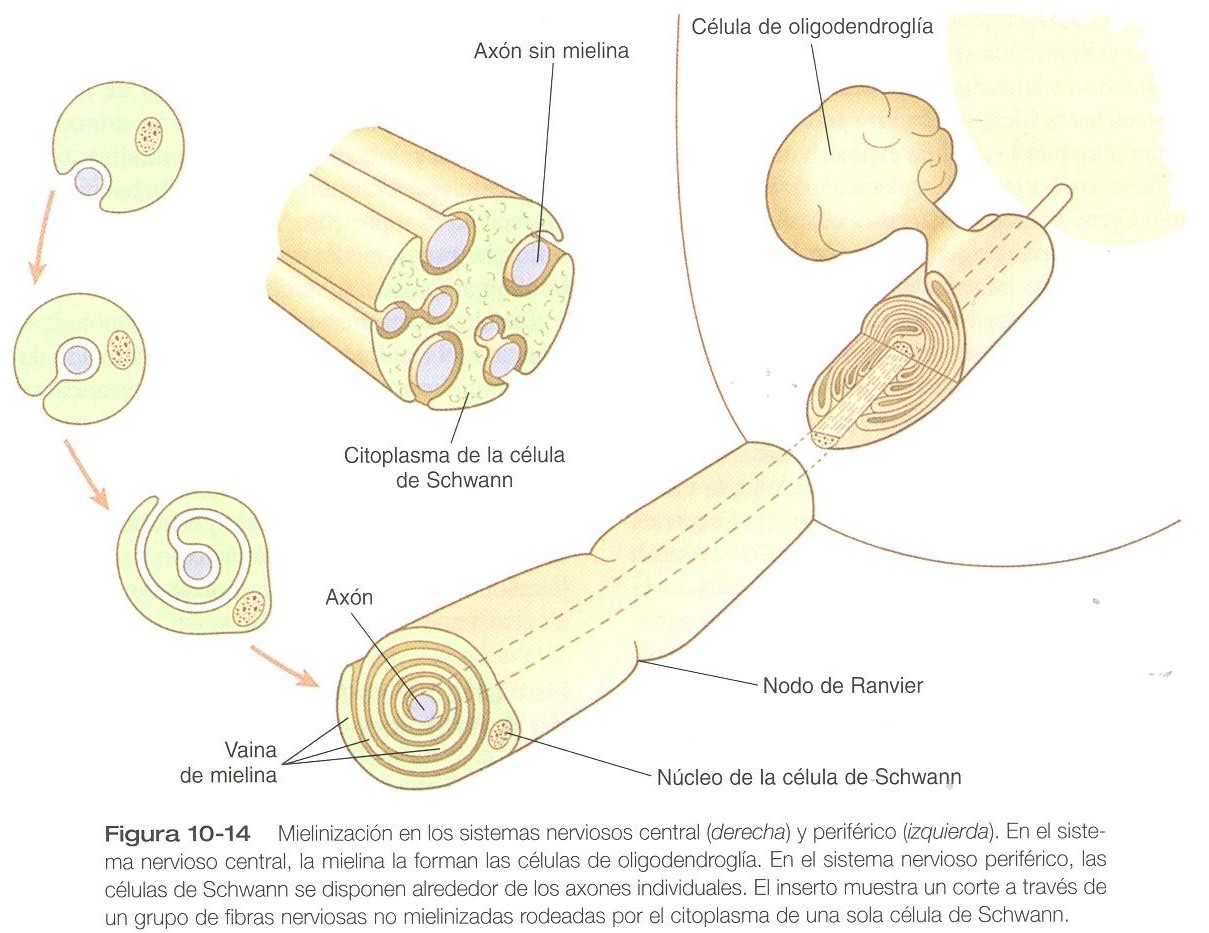{kind=link}
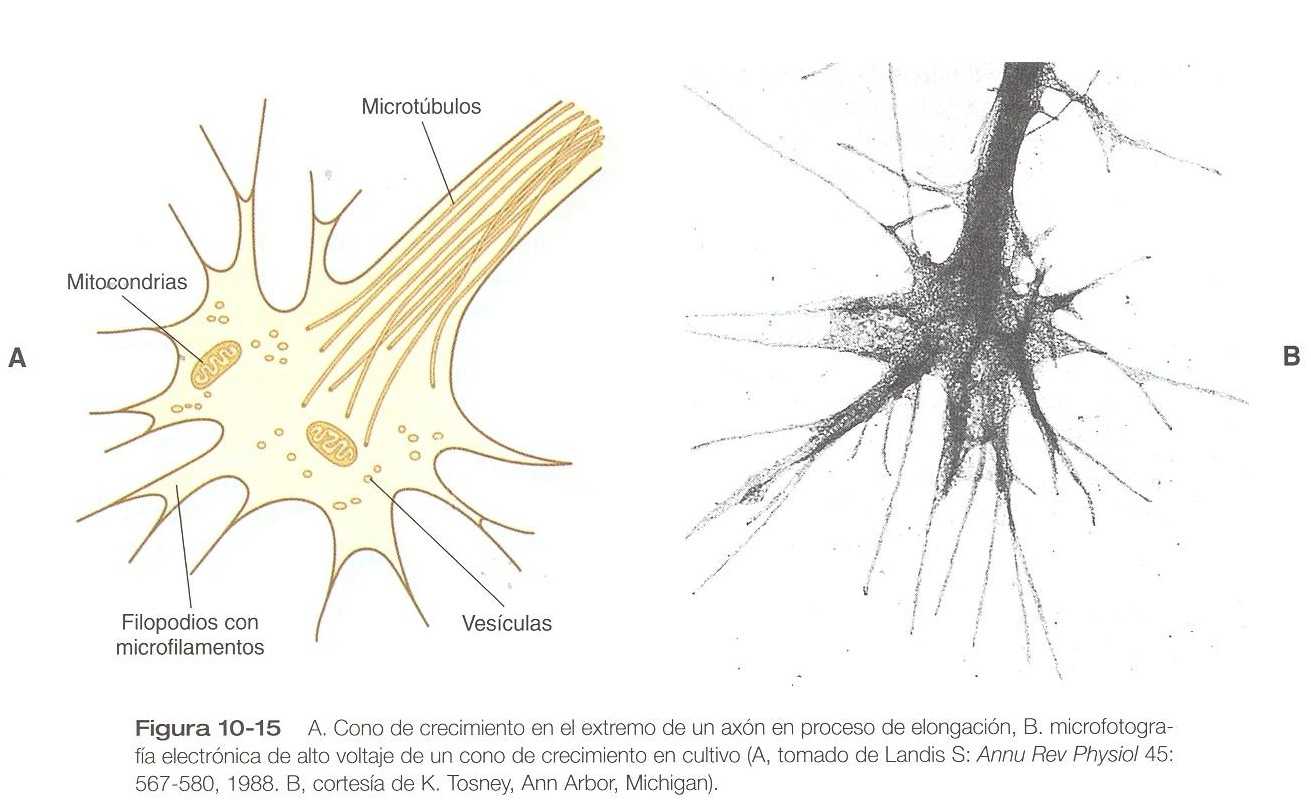{kind=link}
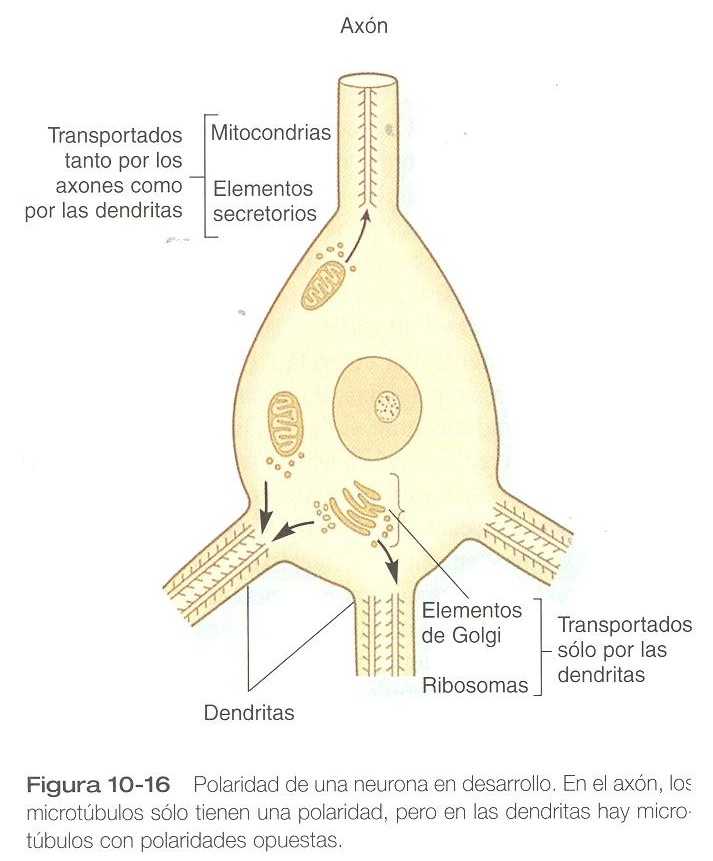{kind=link}
{kind=link}
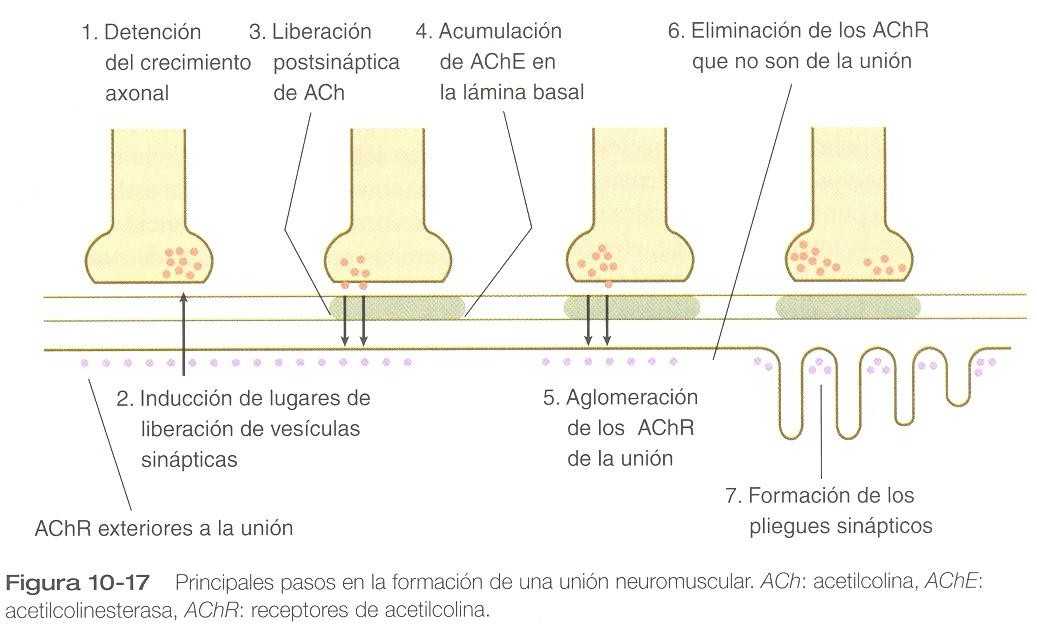{kind=link}
{kind=link}
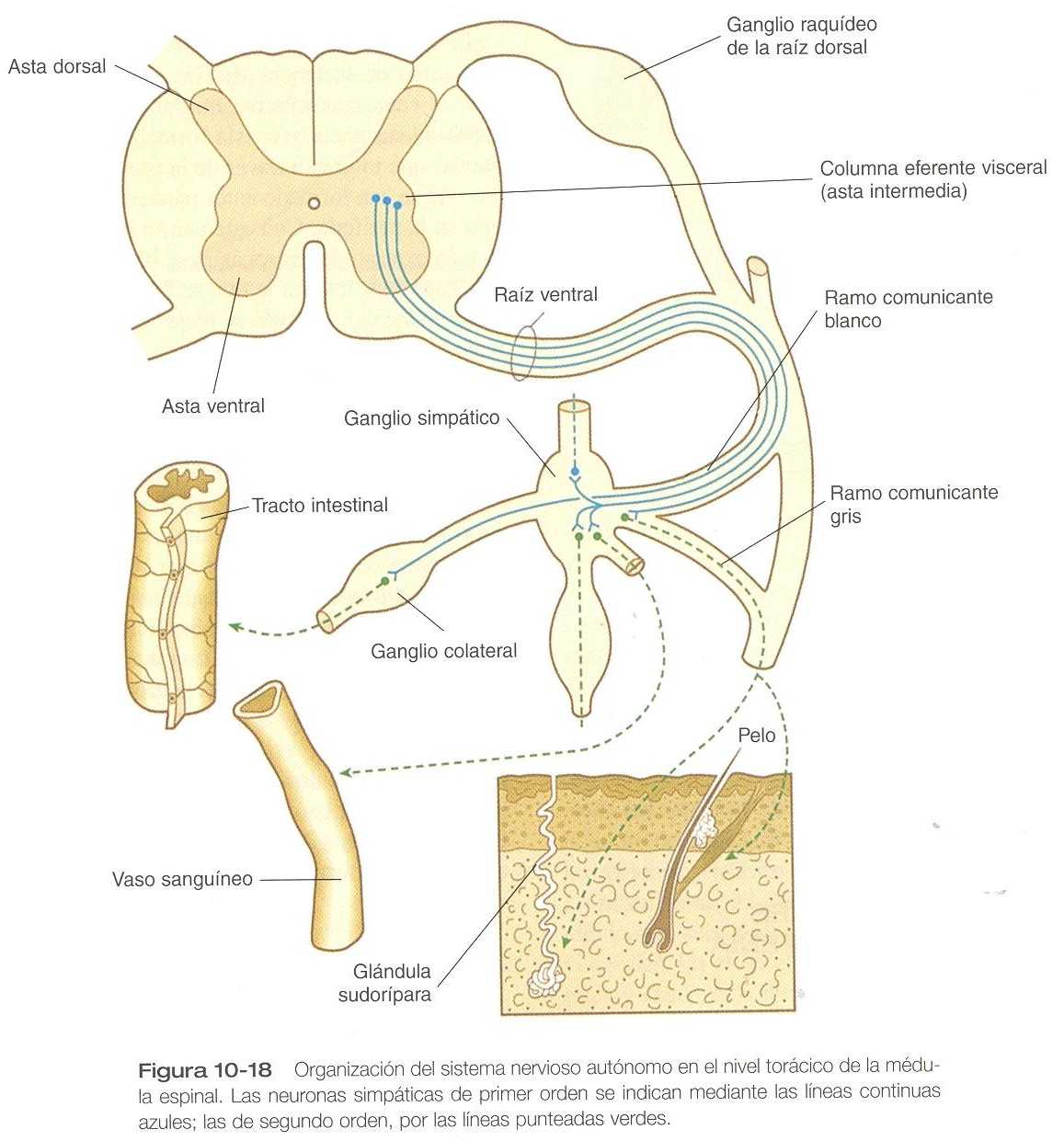{kind=link}
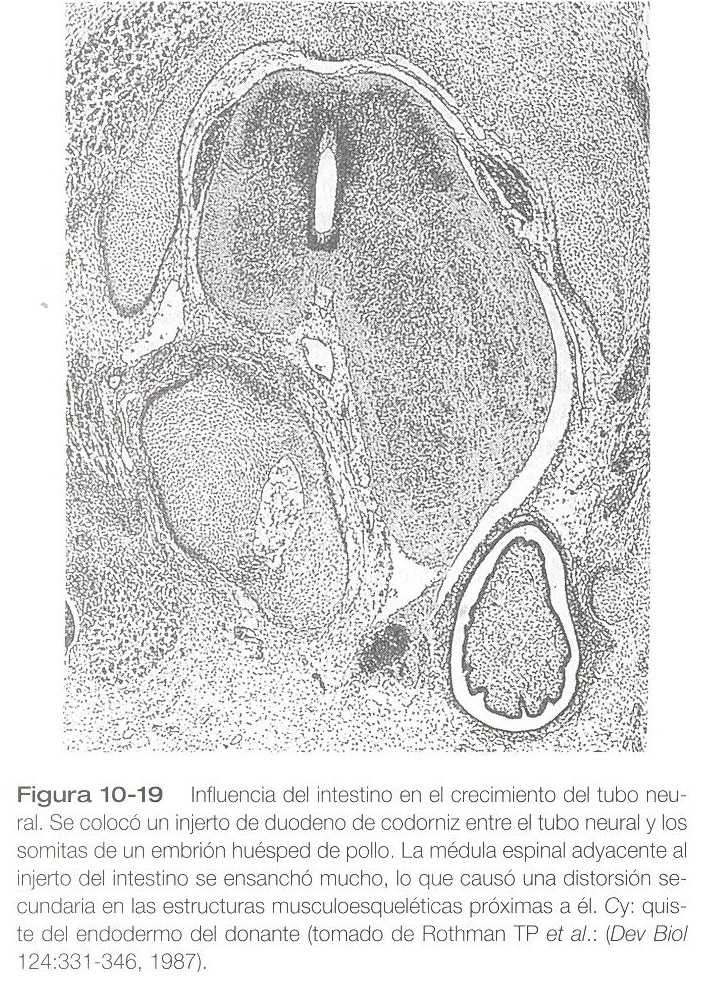{kind=link}
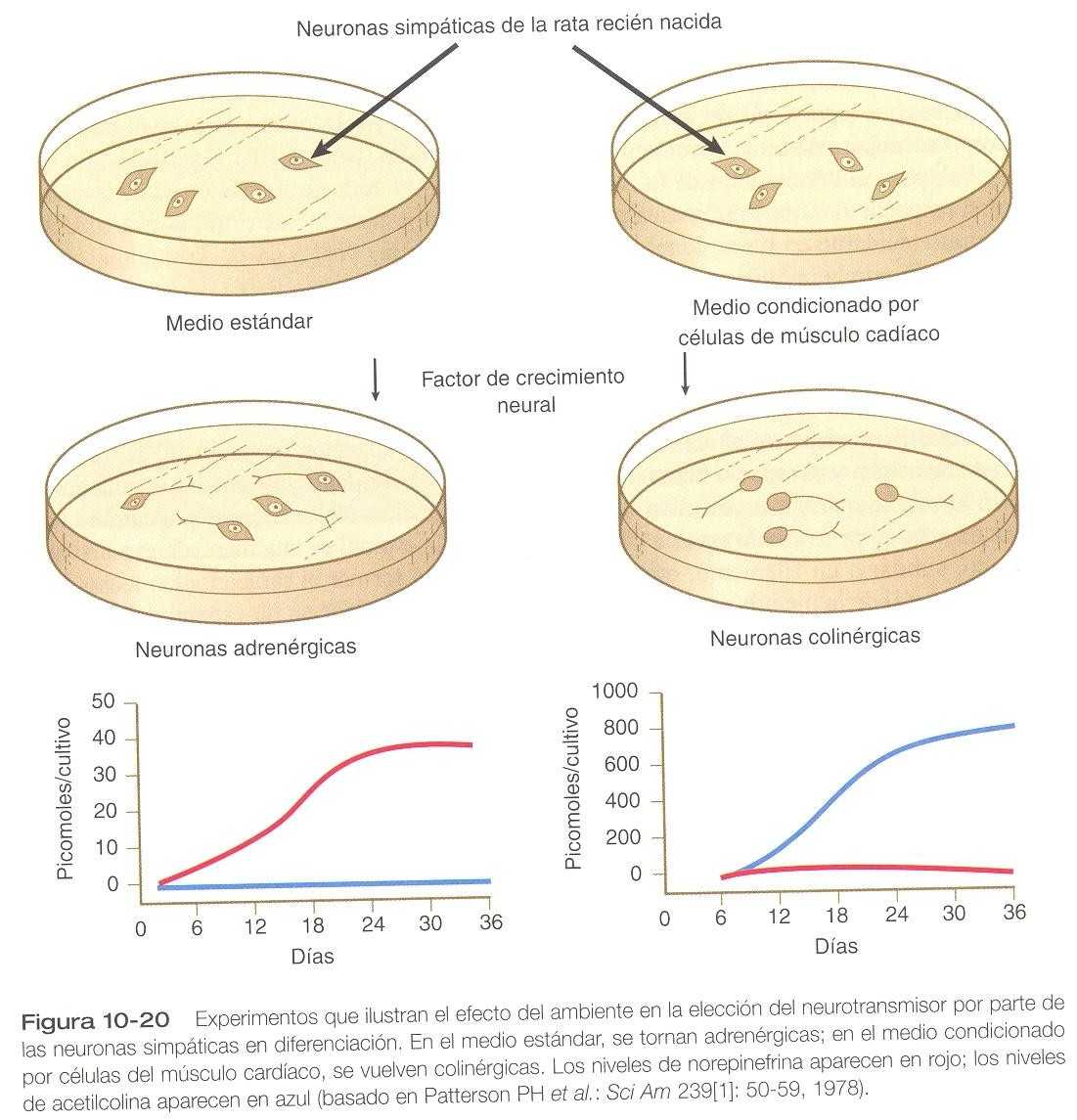{kind=link}
{kind=link}
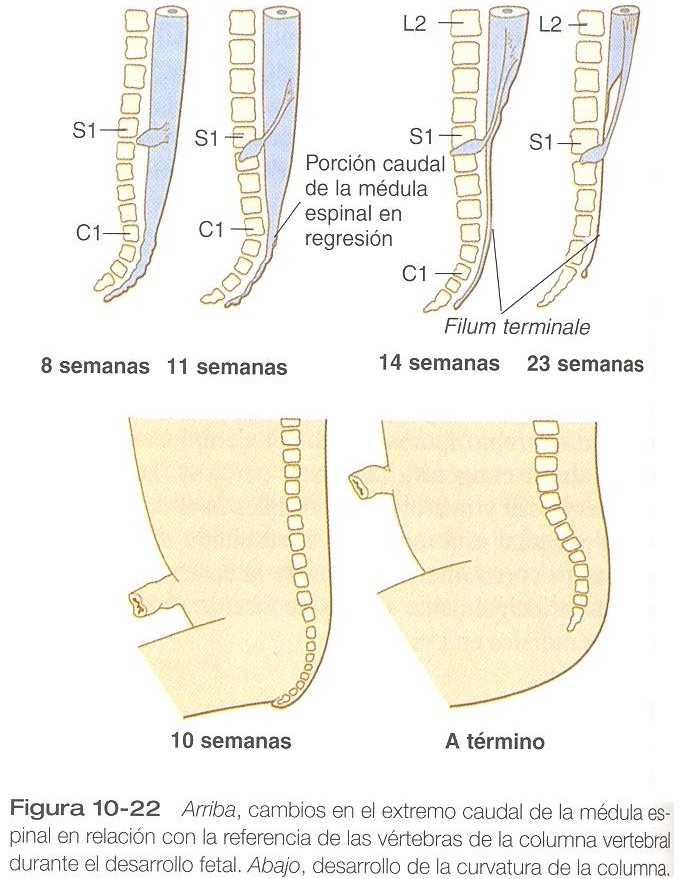{kind=link}
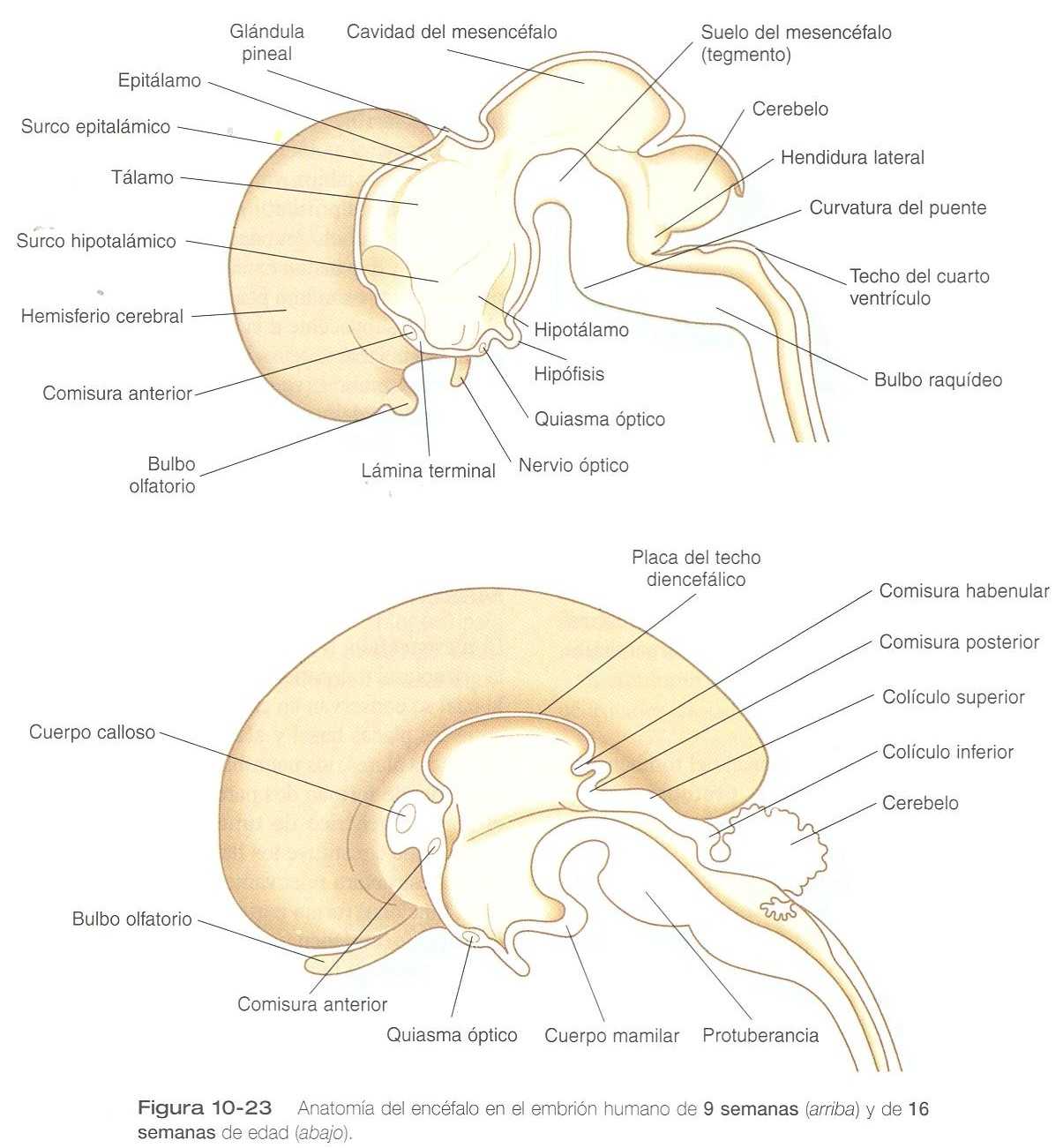{kind=link}
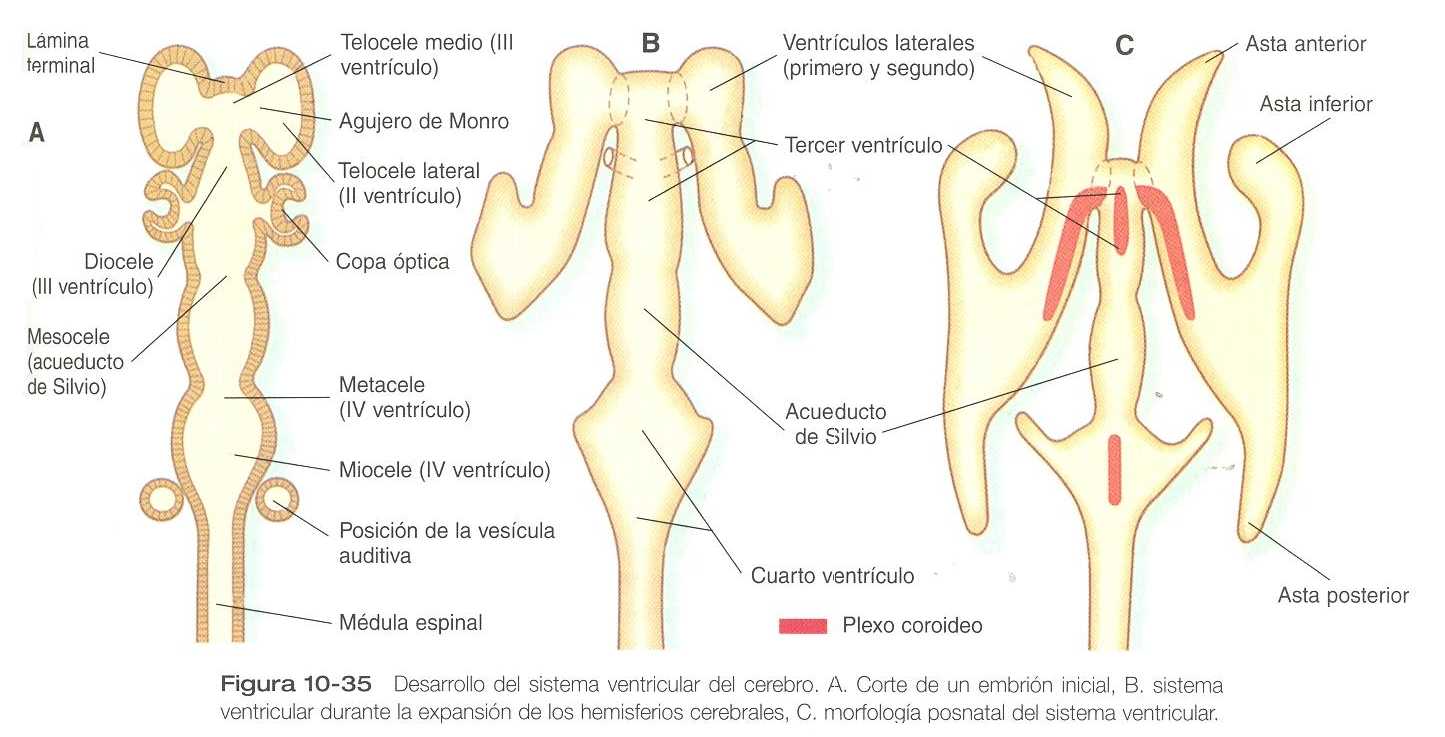{kind=link}
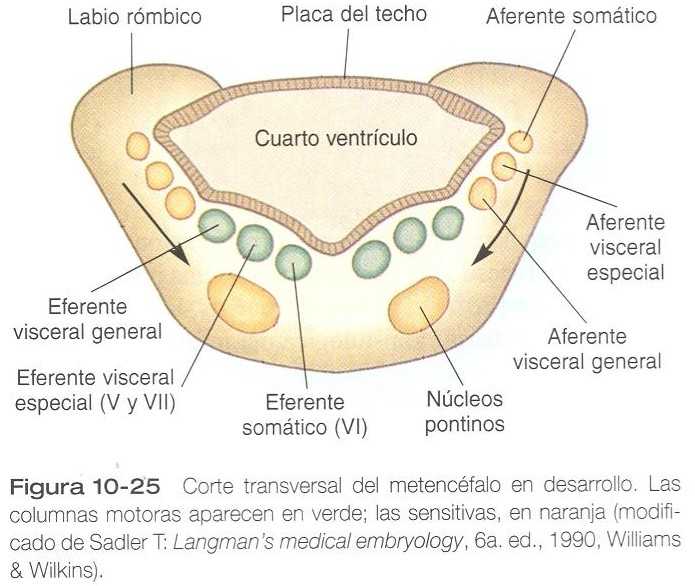{kind=link}
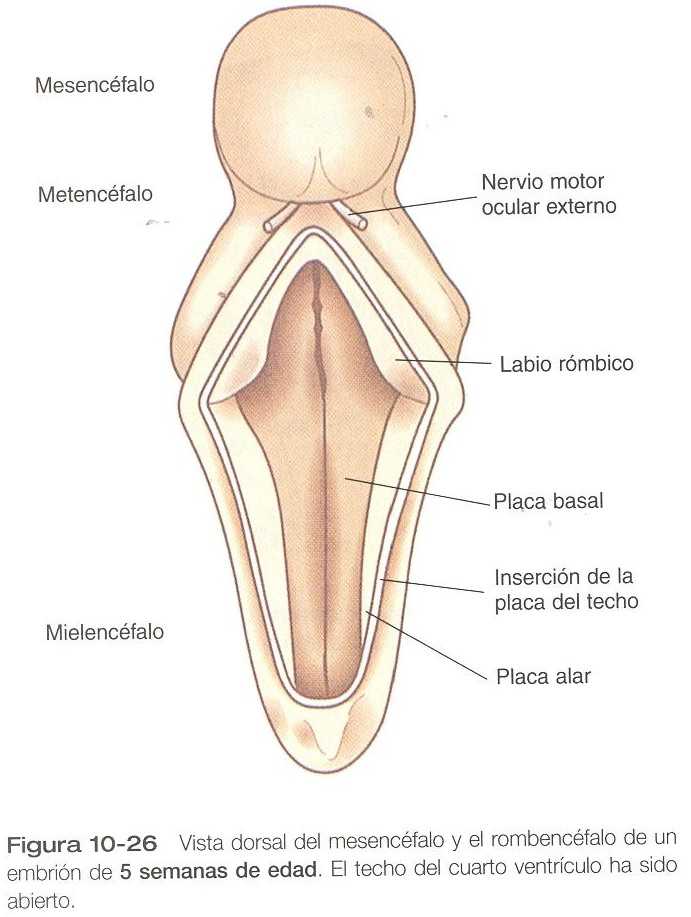{kind=link}
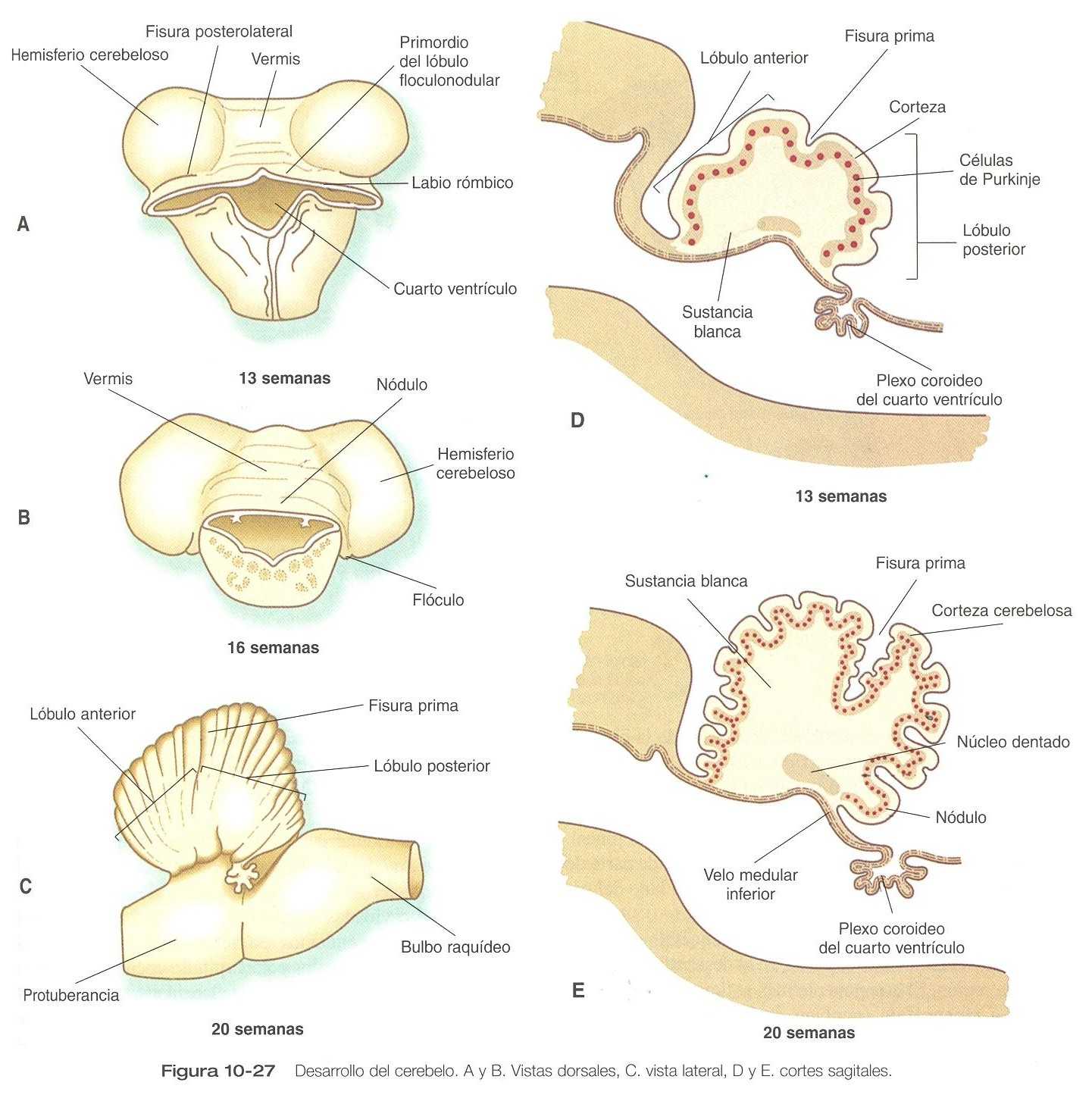{kind=link}
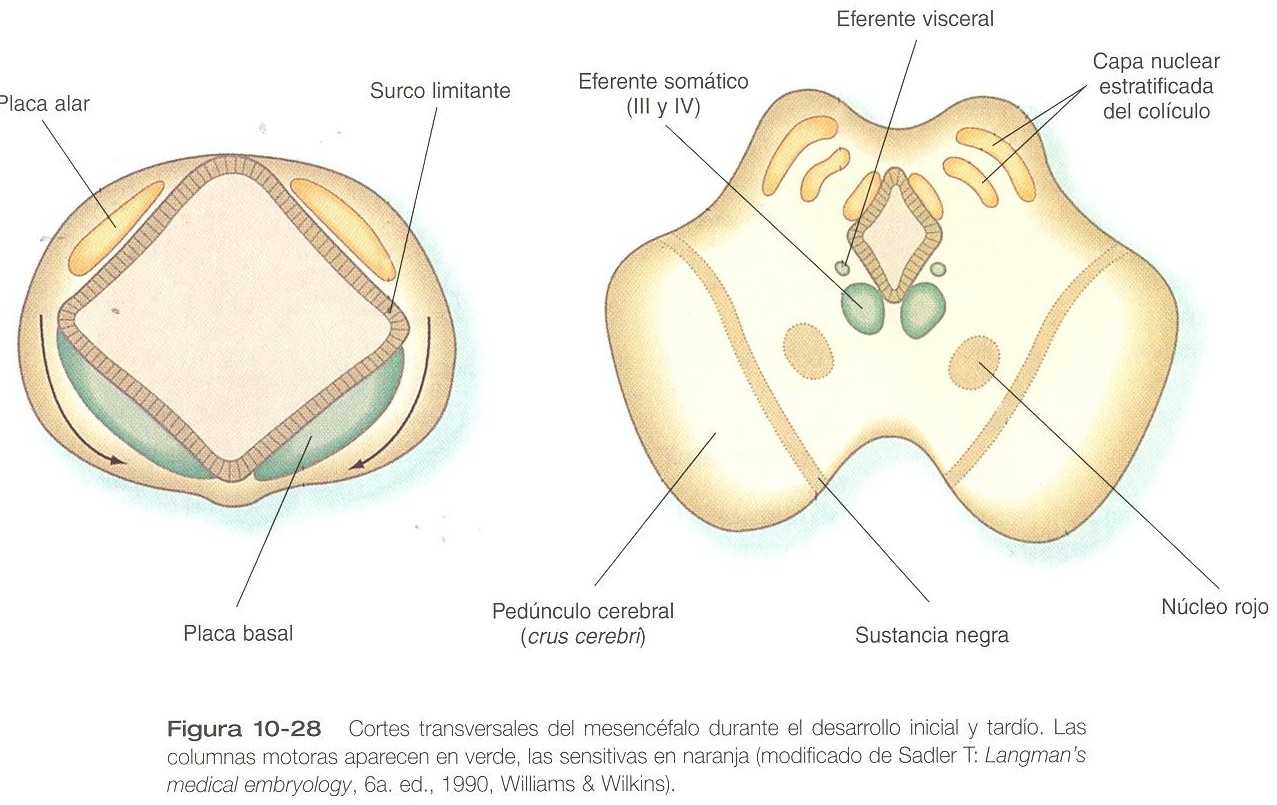{kind=link}
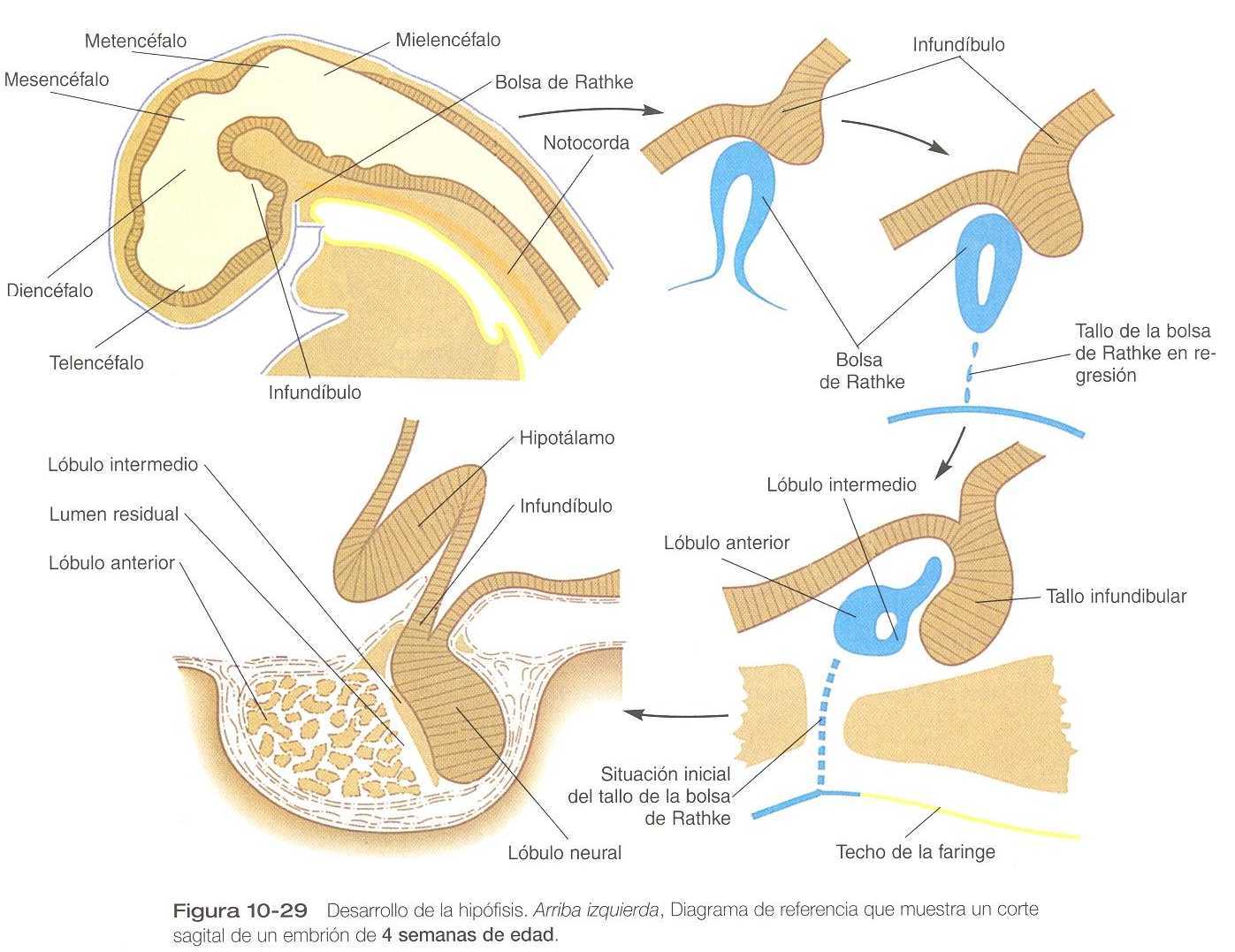{kind=link}
{kind=link}
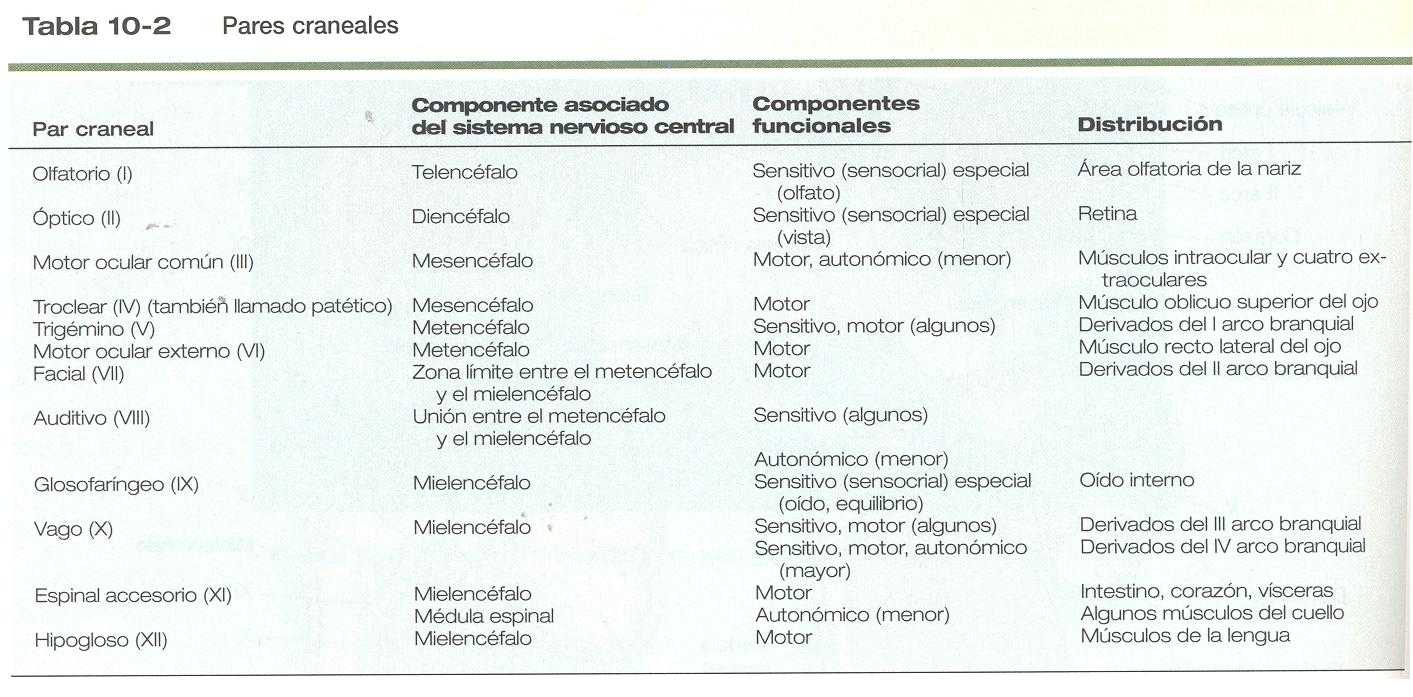{kind=link}
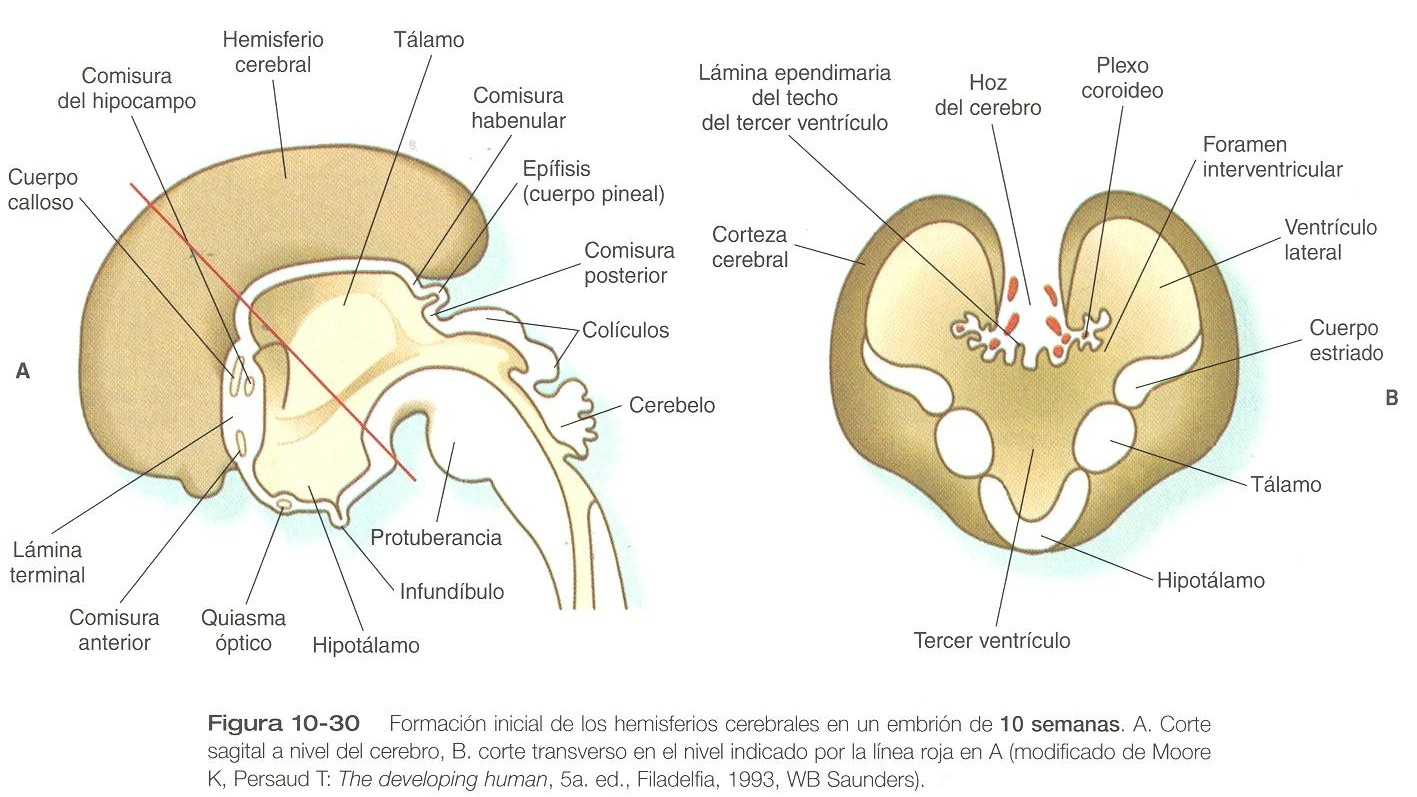{kind=link}
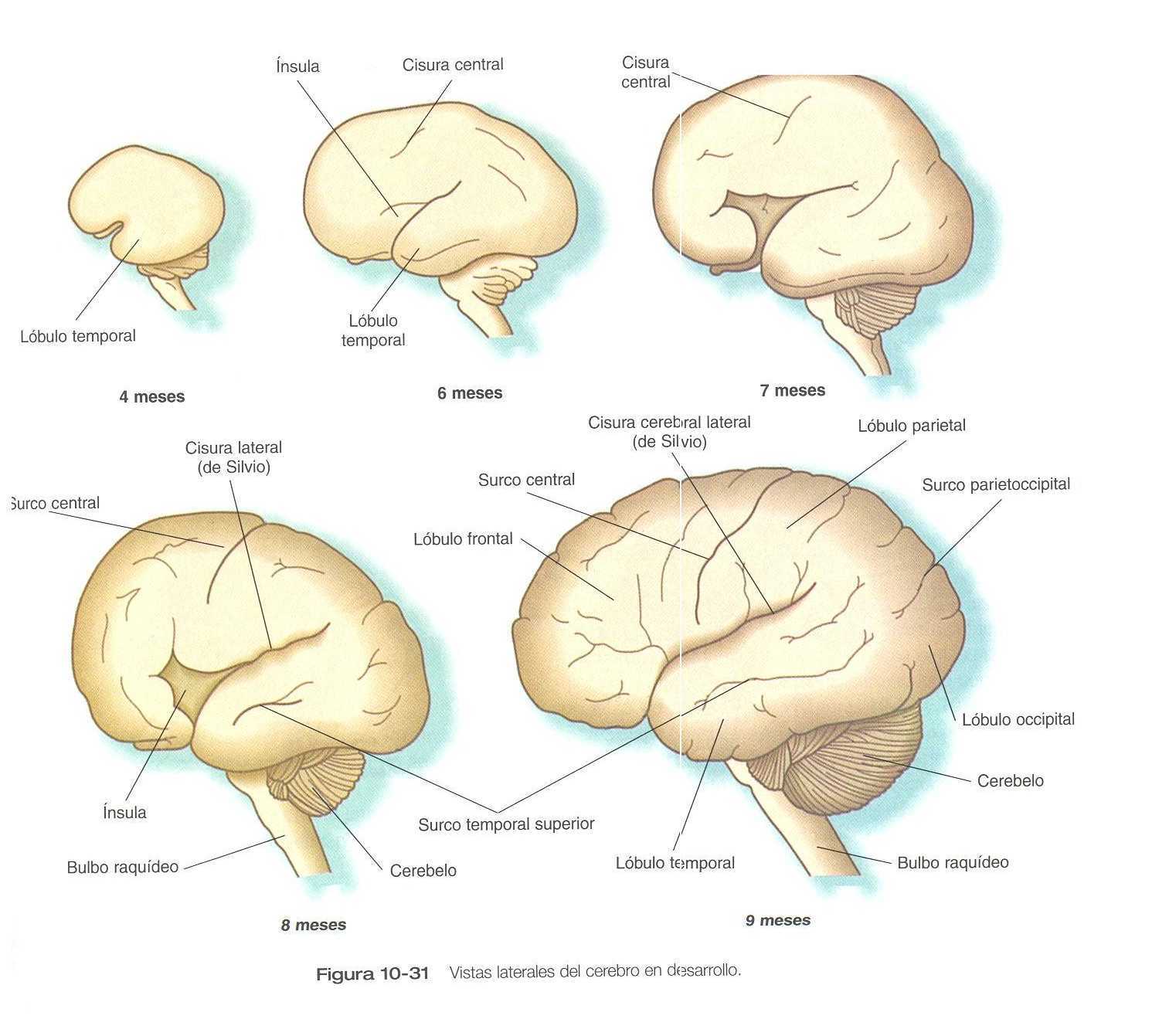{kind=link}
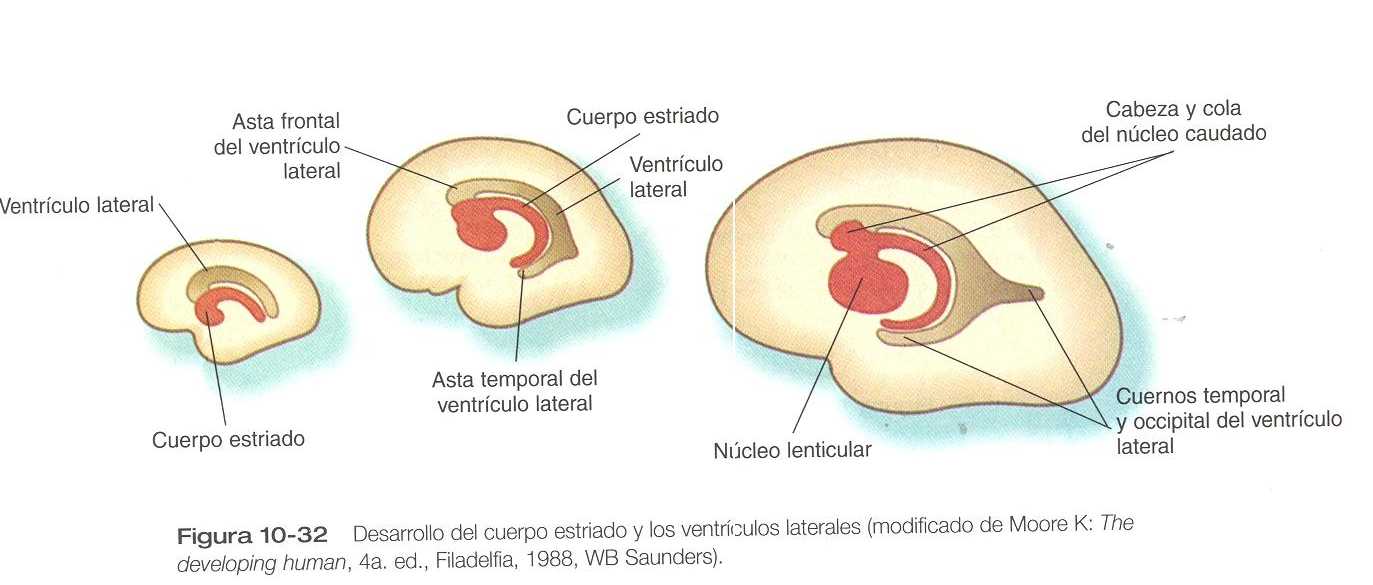{kind=link}
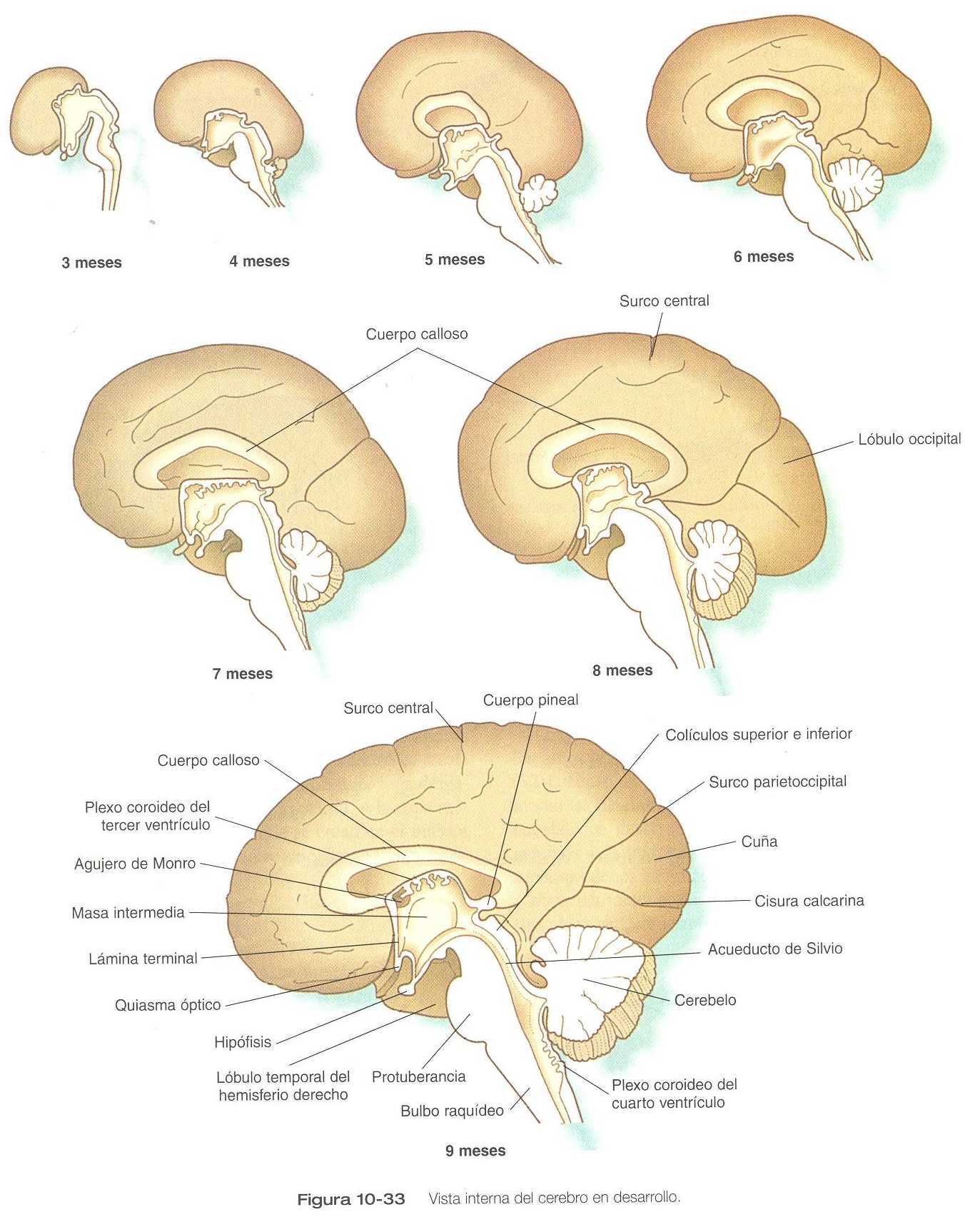{kind=link}
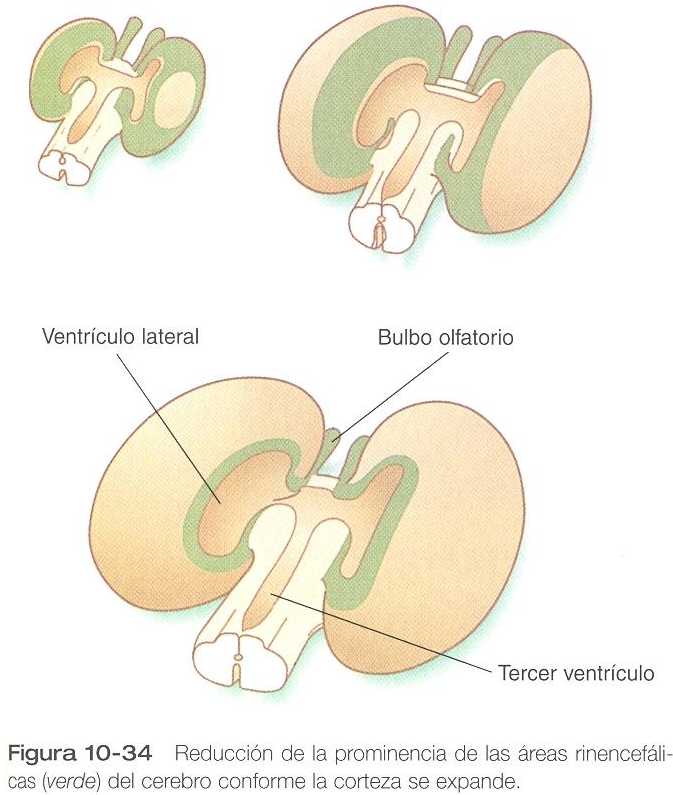{kind=link}
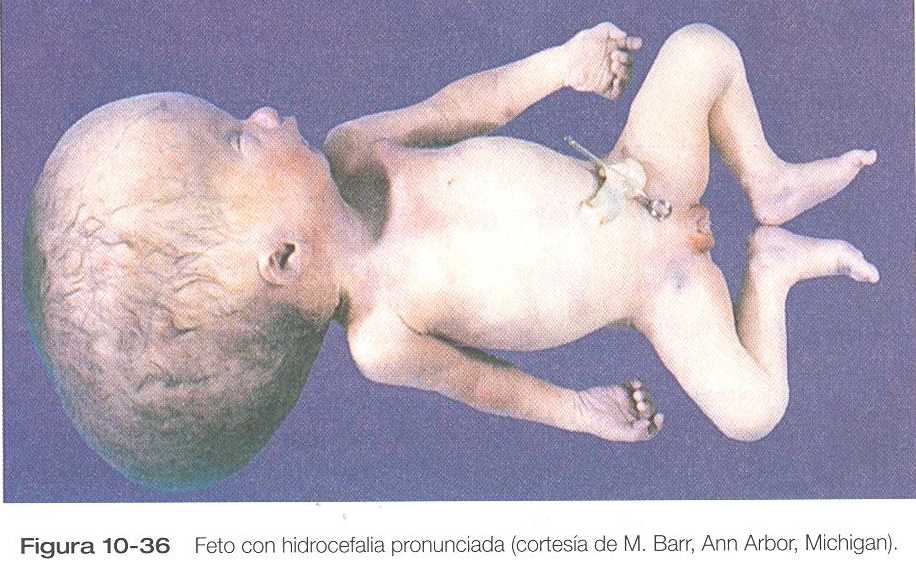{kind=link}
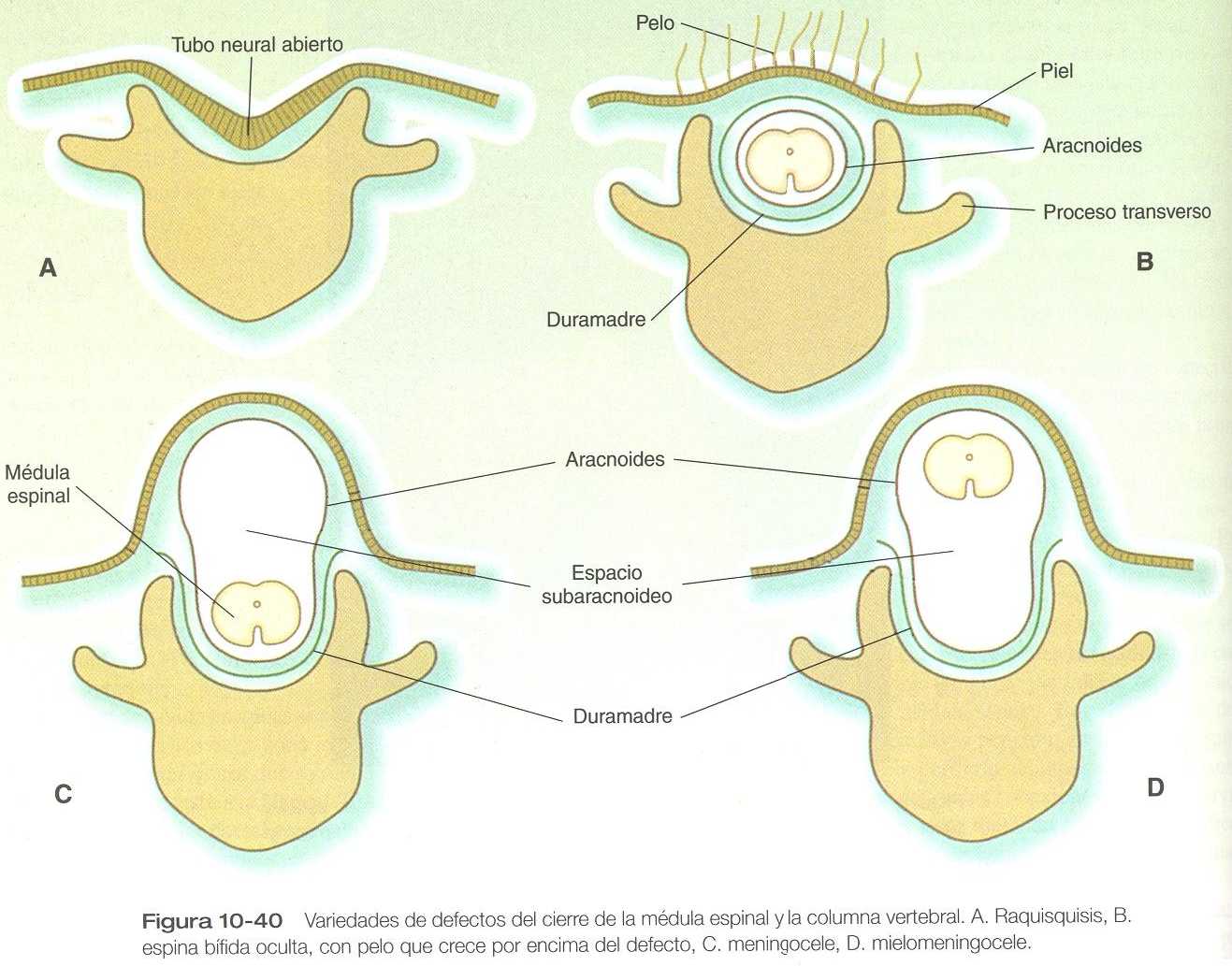{kind=link}
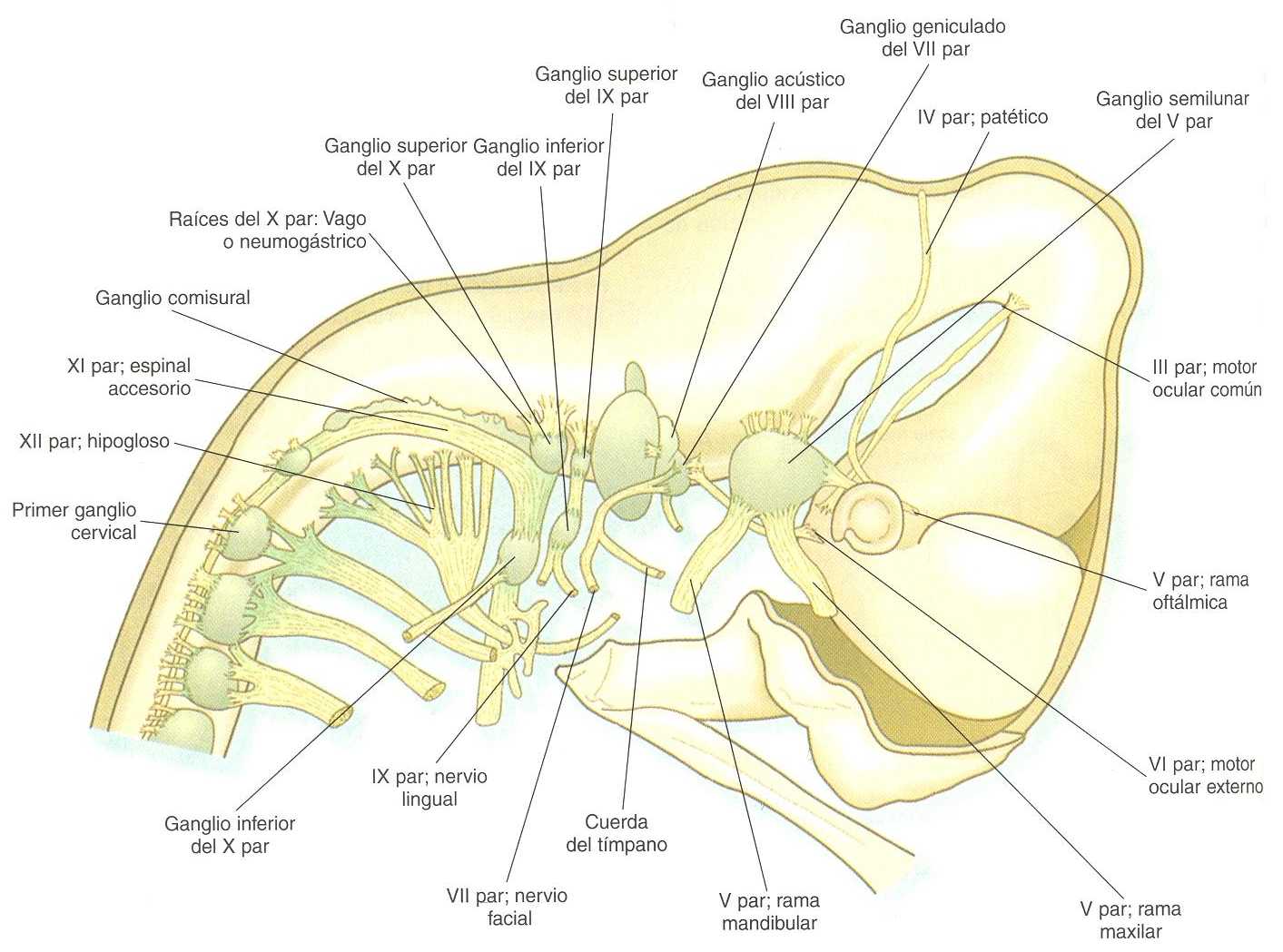{kind=link}
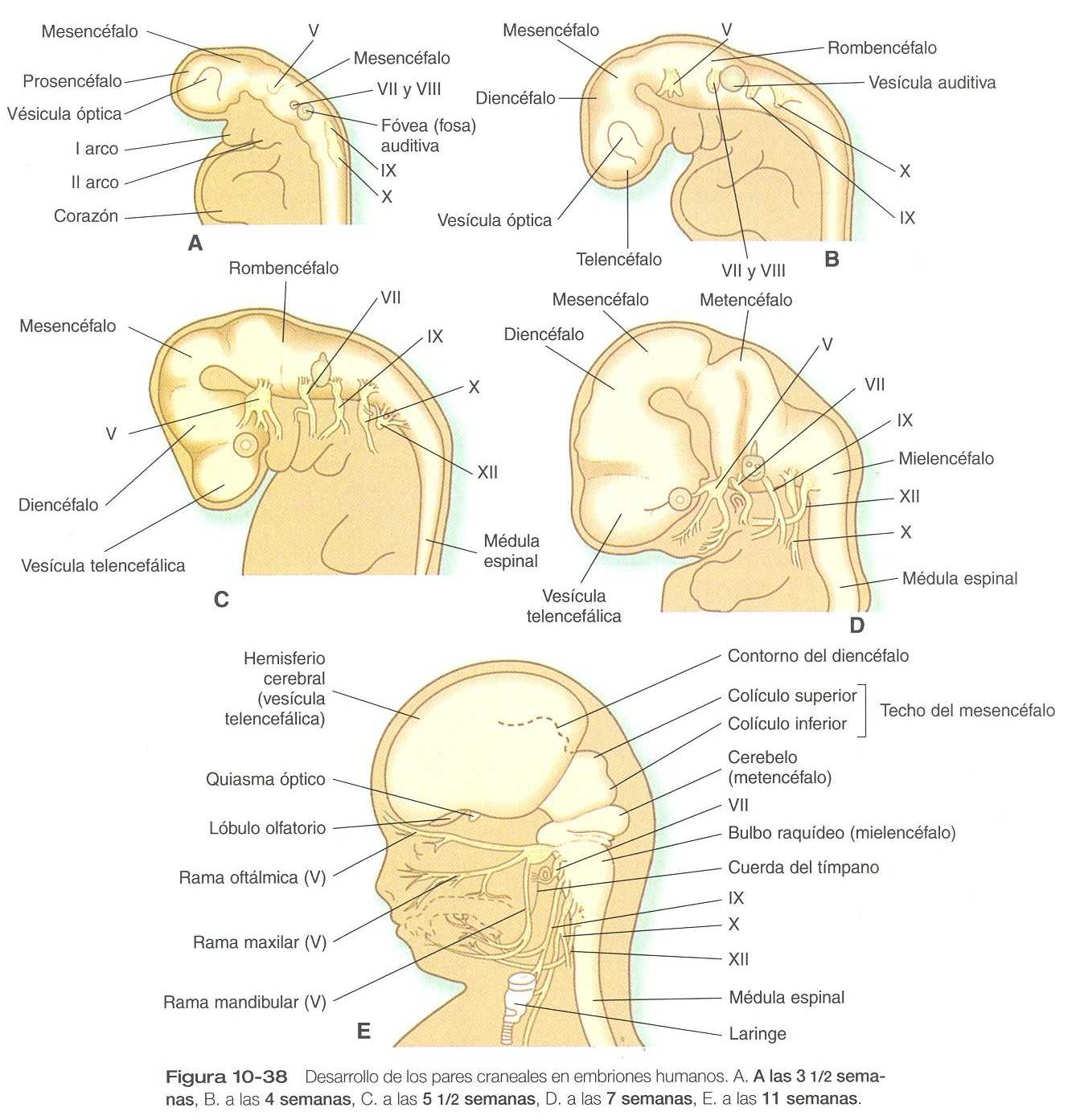{kind=link}
{kind=link}
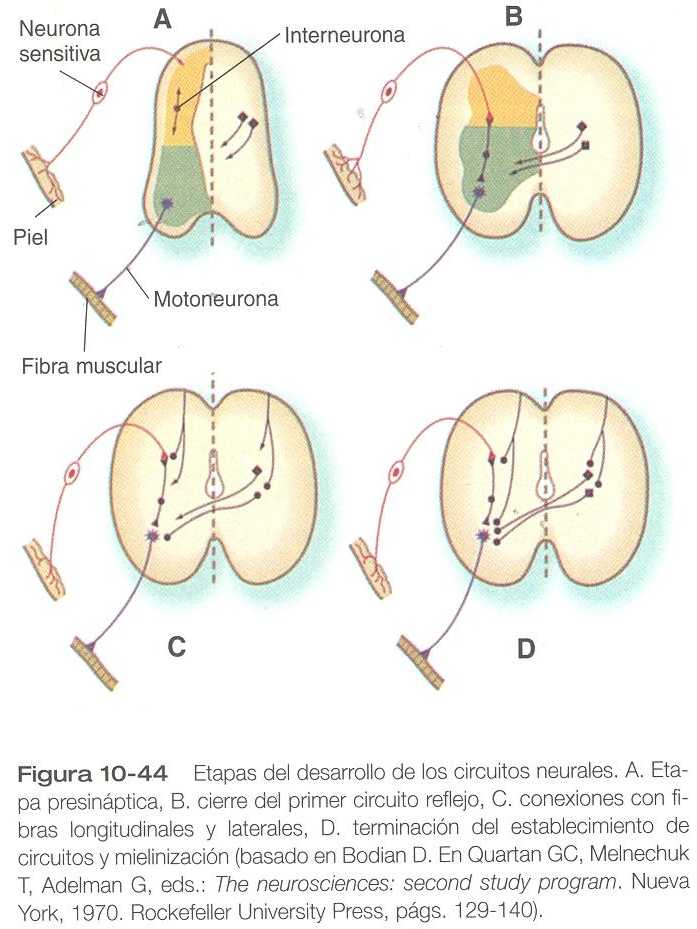{kind=link}
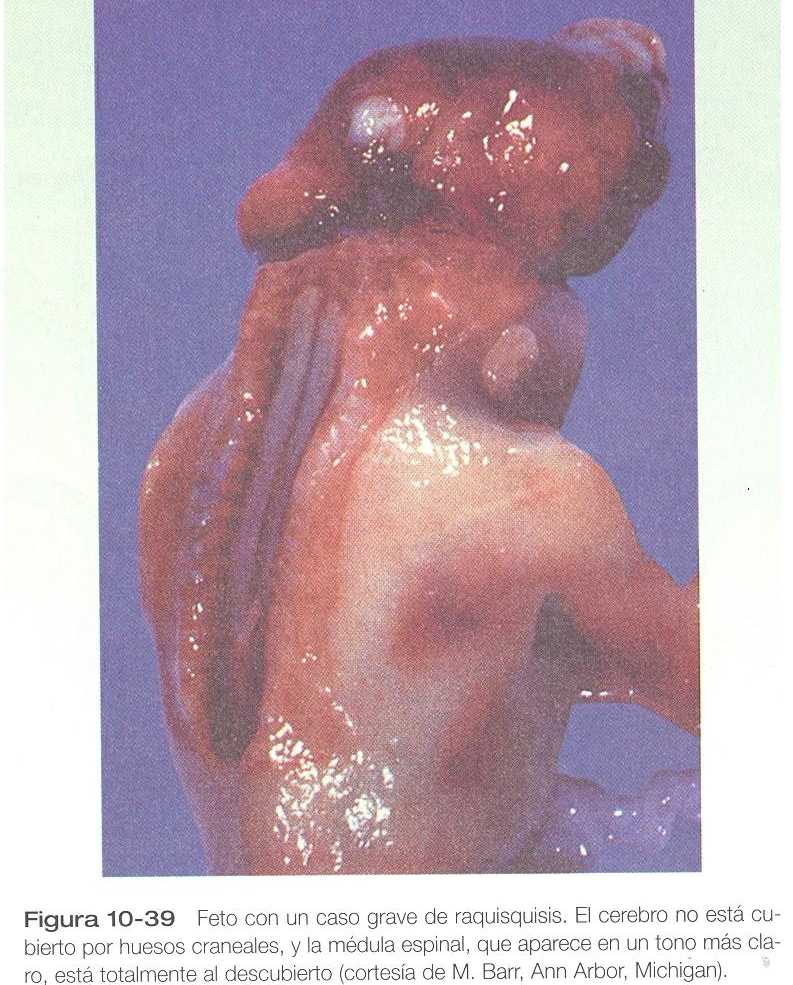{kind=link}
{kind=link}
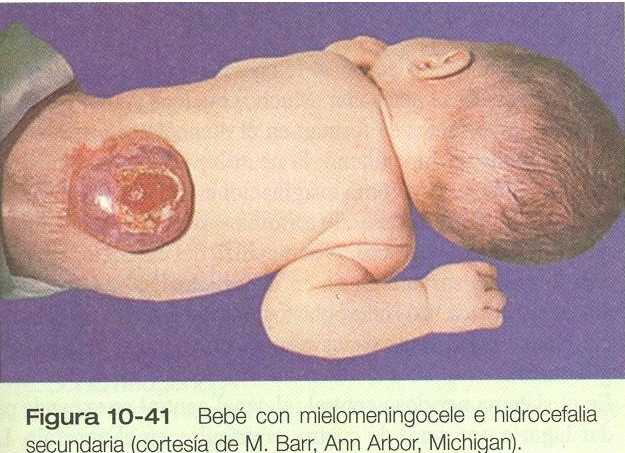{kind=link}
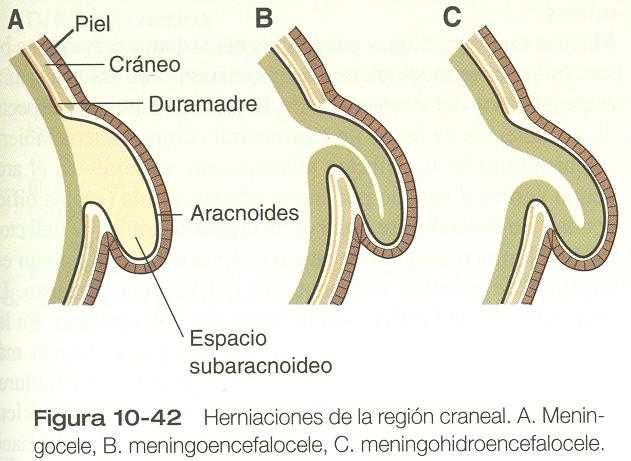{kind=link}
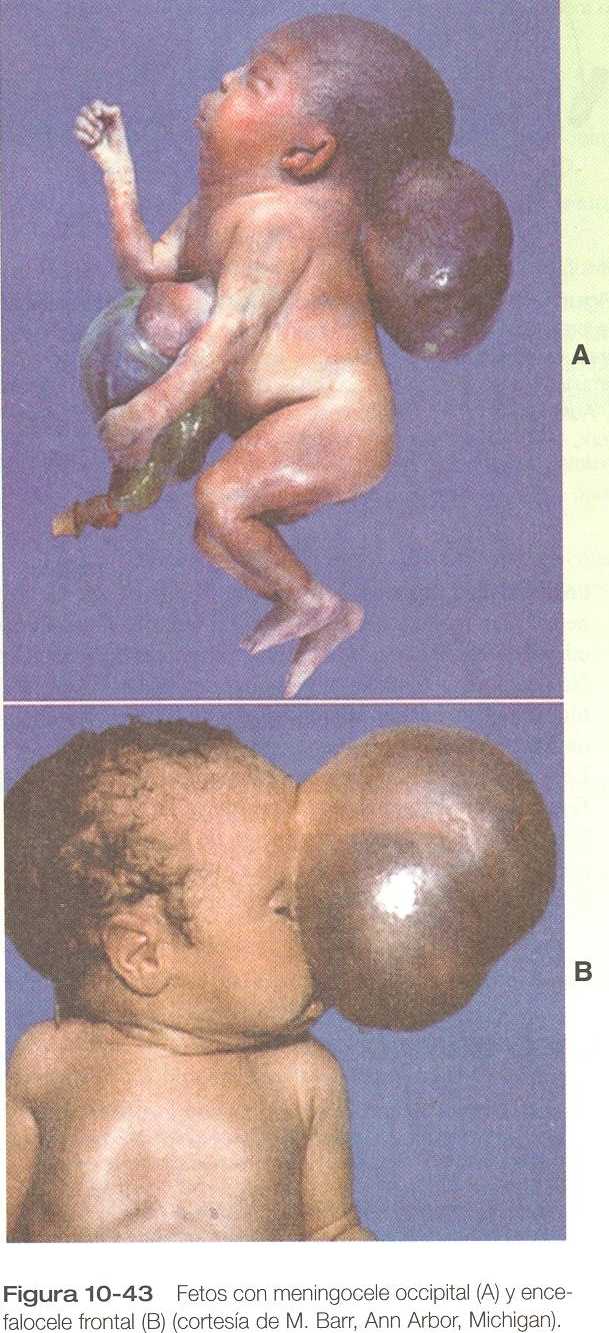{kind=link}
{kind=link}