Células Endócrinas del Estómago Humano Normal
Oscar Larraza
Servicio de Patología, Hospital Central Sur de Concentración Nacional, PEMEX, México, D. F.
Patología Revista Latinoamericana, Volumen 27, número 2, 1989, página 99.
INTRODUCCIÓN
El sistema celular endócrino difuso (SCED), a diferencia de las glándulas endócrinas, se encuentra distribuido en todo el organismo, en contacto estrecho con los epitelios. El SCED posee numerosas funciones, muchas de las cuales comparte con el sistema nervioso y explica interrelaciones de diversos sistemas corporales regulados a través de mensajeros químicos que actúan a distancia de su sitio de producción. De esta manera, la (s) sustancia (s) producidas por una célula pueden verterse a la circulación (endócrina), pueden actuar sobre células contiguas (paracrinia) o sobre la misma célula (autocrinia); ya que se están íntimamente relacionadas con el sistema nervioso, pueden agregarse a la lista anterior los procesos de neurocrinia (nervio - celular) y neuroendocrinia, (nervio - circulación).
La porción alta del tubo digestivo (estómago y duodeno) constituye la zona más prominente de SCED por la gran cantidad de células endócrinas que ahí se localizan. En 1902, Bayliss y Satrling descubrieron una sustancia, extraída del contenido duodenal, capaz de estimular la secreción de agua y electrolitos de páncreas denervado; la denominaron secreina en virtud de esta función y, posteriormente Hardy propuso el término de Hormona para designar las características de esta de esta sustancia y la de otros mensajeros con mecanismo de acción similar (la palabra hormona procede del griego - ormein -y significa estimulo). Si se piensa someramente en estos datos, la endocrinología moderna nació a partir de observaciones realizadas en el tubo digestivo.
Las células (q2a6 UFlo) responsables de las funciones endócrinas del SCED en general y las localizadas en el tubo digestivo en particular, se conocen desde 1870: Heidenham las identificó por primera vez. En 1891, Nicolas las denominó células amarillas, al estudiar el epitelio digestivo con las sales de cromo. Posteriormente, Kultschitzky las llamó células acidófilas en 1897. Su terminología sufrió nuevos cambios en 1905, 1906 y 1914, en los que fueron denominadas cromafines, enterocromafines y argentafines por Schmidt, Ciaccio y Masson, respectivamente. La terminología anterior refleja algunas de las propiedades histoquímicas de las células endócrinas del SCED; sin embargo, no fue sino hasta el advenimiento de nueva metodología de estudio que estas células y sus productos de secreción se han caracterizado adecuadamente; ello incluye la microscopía electrónica, las técnicas de inmunolocalización y más recientemente, la hibridización in situ de ácidos nucleicos.
CARACTERÍSTICAS DE LAS CÉLULAS ENDÓCRINAS DEL TUBO DIGESTIVO
Con tinciones convencionales, las células endócrinas se encuentran en relación estrecha con los epitelios superficiales y con las glándulas submucosas; se observan como células de citoplasma claro o eosinofilo granular. Al ser sometidas a sales de plata o de cromo, algunas de ellas revelan la presencia de gránulos intracitoplásmicos; esta propiedad se conoce como argentafinidad y depende probablemente del contenido de aminas presentes en el citoplasma; puede demostrarse con varias técnicas entre las que se encuentra la de Fontana - Masson y la inducción de fluorecencia con vapores de formol. Otra propiedad que tiene la mayoría de las células endócrinas del tubo digestivo alto es la argirofilia, que difiere de la argentafinidad en el hecho de que las células incorporan sales de plata previamente reducidas; en esta propiedad se basan las técnicas diseñadas por Grimelius, Bodian, Churukian y otras. Esta metodología histoquímica de estudio es accesible a la mayoría de los laboratorios de Patología; sin embargo, únicamente proporciona información genérica y no hace evidente la o las sustancias producidas y almacenadas por las células. Con el advenimiento de la microscopía electrónica resultó obvio que las células endócrinas gastrointestinales poseen gránulos intracitoplásmicos distintivos en condiciones normales, que permiten, en función de su morfología, separar a las diferentes células; de hecho, las primeras clasificaciones de las células endócrinas gástricas (CEG) se realizaron sobre bases ultraestructurales.
A partir del empleo de técnicas de inmunolocalización, sobre todo de las que emplean inmunoperoxidasa para revelar el producto almacenado, el panorama se ha ampliado notablemente. Existen técnicas que informan sobre productos genéricos de las CEG, tales como la enolasa neuronal específica, que se localiza en el citosol; la cromogranina, que es una proteína asociada a los gránulos de secreción y la sinaptofisina, que se encuentra dentro de los gránulos; con estas técnicas sencillas de realizar y reproducibles fácilmente, es posible asegurar la naturaleza endócrina de las CEG en condiciones normales o patológicas. De mayor especificidad resulta el empleo de anticuerpos dirigidos contra productos almacenados por las células; así se puede saber si una célula posee gastrina o serotonina o secretina, somatostatina u otra sustancia hormonal (vide infra). Con las técnicas de inmunolocalización ha sido posible evidenciar, además, la existencia de sustancias que normalmente no están presentes en las CEG; este fenómeno se observa en muy diversas neoplasias endócrinas de esta zona y permite postular que las Ceg poseen una programación genética común.
El panorama se ha ampliado aín más con el decubrimiento y aplicación de las técnicas de hibridización de ácidos nucleicos, en las que se emplean guías de DNA o RNA biotinadas o marcadas con sustancias radioactivas, con lo que se obtiene información del programa genético de la célula en estudio, a diferencia de las otras técnicas, que únicamente evidencian al producto almacenado.
LOCALIZACIÓN Y NOMENCLATURA
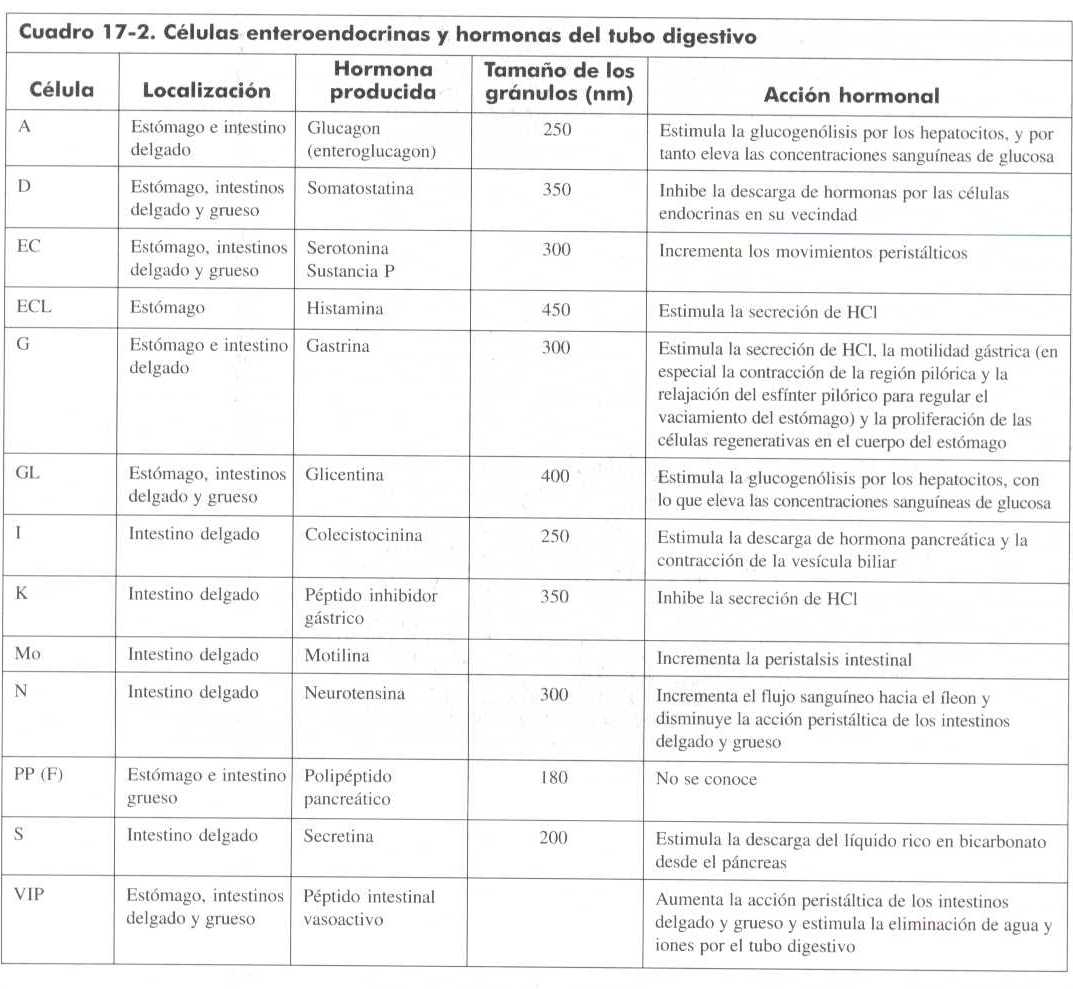
El empleo de las técnicas descritas en la sección precedente ha permitido saber que las CEG se encuentran en diversas localizaciones. En la mucosa gástrica se ubican en dos sitios preferentemente: la mucosa especializada u oxíntica (cuerpo y fondo) o en el antro gástrico. en la primera zona están relacionadas con las glándulas gástricas de la porción media e inferior del epitelio; en la segunda, se encuentra asociadas con la porción profunda de las glándulas. Algunas sustancias se producen en los dos territorios; tal es el caso de la somatostatina y la serotina; otras como la gastrina, son exclusivas de la porción antral. Como puede verse en la tabla 1, hay numerosas sustancias almacenadas en las células endócrinas, cuya identidad aún es desconocida; se sabe de su existencia gracias a los estudios ultraestructurales. En la misma tabla aparece la nomenclatura de las CEG, aceptada mundialmente. Esta terminología es confusa y difícil de aprender ya que algunas células se designan de acuerdo con su similitud con las células endócrinas del páncreas (por ejemplo, la célula D, productora de somatostatina); otras llevan la inicial de su producto de secreción (célula G, productora de gastrina); algunas se clasifican de acuerdo con la morfología ultraestructural de sus gránulos (EC, enterocromafines; ECL, similares a enterocromafines) y, por último, una célula no clasificable de acuerdo con los criterios anteriores y cuyo producto de secreción se desconoce, se denomina X. Parece razonable suponer que esta clasificación cambiará y se hará sobre bases más uniformes.
INTERACCIONES DE LAS CÉLULAS ENDÓCRINAS
Por medio de los mecanismos de endocrinia, paracrinia y neuroendocrinia, la CEG interactúan con células endócrinas de otras localizaciones, son influenciadas por secreción nerviosa y se influencian recíprocamente en la misma área. Por ejemplo, la gastrina se secreta por influencia neural t también por distensión gástrica, aminoácidos y el pH del estómago; a su vez, una vez secretada actúa sobre las células parietales del estómago, con lo que favorece la producción del ácido clorhídrico. Además, promueve la secreción de insulina y calcitonina por lo que no es infrecuente encontrar hiperolasia de células beta del páncres y de células C de la tiroides en los estados de hipergastrinemia. Por otro lado, también estimula la producción y liberación de somatostatina, secretina, glucagon, péptido intestinal vasoactivo y péptido inhibitorio gástrico; todas estas sustancias, a su vez inhiben por vía sanguínea la secreción de gastrina. En el síndrome de Zollinger . Ellison, cuya característica primordial es la hipergastrinemia, se observa incremento de células endócrinas del estómago (D, ECL), del duodeno (D, S, K) páncreas (alfa, beta) y de la glándula tiroides (C); hay también incremento (hiperplasia) de las células epiteliales gástricas y duodenales. Estas interrelaciones complejas se han documentado también en el resto de las CEG y explican, en parte, la existencia de más de una célula endócrina en neoplasias endócrinas del tubo digestivo.
EMBRIOLOGÍA
Feyter, en 1938 postuló que las CEG se derivan del endodermo. En 1955 Pages sugirió un origen neuroectodérmico, que fue reforzado por Pearse en 1968; este autor es el creador del concepto APUD, que unifica las características funcionales del SCED en relación con su capacidad de captación y descarboxilación de aminiácidos.
Un concepto unitario, propuesto también por Pearse, fue el atribuir el origen del SCED específicamente a la cresta neural primitiva. Sin embargo, evidencias proporcionadas por diferentes fenómenos biológicos y por modelos experimentales, han desacreditado este concepto; Así en procesos inflamatorios, displásicos y neoplásicos del tubo digestivo alto, has incremento de células endócrinas aun en aquellos sitios en los que no se observan en condiciones normales, como es el caso de la vesícula biliar; por otro lado, la extirpación de las crestas neurales en fases tempranas del desarrollo embrionario no impide la aparición de células endócrinas en diversos modelos animales de experimentación, tal como lo ha demostrado Fontaine y cols. Argumentos adicionales en contra del origen neural de las CEG se han sugerido al observar neoplasias que contienen tanto células endócrinas como epiteliales en distintas áreas de la lesión o aun ambas diferenciaciones en la misma célula. Por todo lo anterior, la mayoría de los autores, actualmente, postulas un origen endodérmico de las CEG. (cuadro Gartner 17-2c)
Célula |
Mucosa oxintica |
Antro |
Sustancia producida |
D |
+ |
+ |
Somastostátina |
D1 |
+ |
+ |
Desconocida |
EC |
+ |
+ |
Serotonina |
ECL |
+ |
- |
Desconocida |
G |
- |
+ |
Gastrina |
P |
+ |
+ |
Desconocida |
X |
+ |
- |
Desconocida |
| Adaptado de Lechago, J: Am J Surg Phat 1987; 11:63 | |||
{kind=link}