METABOLISMO
Las plantas utilizan un pigmento
verde, la clorofila, para captar la energía de la luz solar. A diferencia de
los vegetales, los seres humanos carecemos de un pigmento en la piel que
realice funciones similares, por ello la única fuente de energía de que
disponemos para efectuar el trabajo biológico son los alimentos que ingerimos.
Es necesario que nuestro cuerpo produzca a partir de precursores más simples
muchas moléculas que son necesarias para el mantenimiento de células y tejidos;
otras como los aminoácidos esenciales, vitaminas y minerales las obtenemos de
los nutrimentos, puesto que no podemos sintetizarlas. Las enzimas del conducto
gastrointestinal digieren los carbohidratos, lípidos y proteínas de los
alimentos. Los productos finales de la digestión que llega a las células del
cuerpo son monosacáridos, ácidos grasos, glicerol, monoglicéridos y
aminoácidos. Algunos minerales y muchas vitaminas son parte de sistemas
enzimáticos que catalizan el desdoblamiento y la síntesis de carbohidratos,
lípidos y proteínas. Las moléculas de los alimentos que absorbe el conducto
gastrointestinal (CG) tienen tres destinos principales:
La mayor parte de
ellas se utilizan para suministrar energía, ya que el cuerpo debe
realizar procesos vitales, como el transporte activo, la duplicación del ADN,
la síntesis de proteínas, la contracción muscular ya la mitosis.
Algunas moléculas
de los alimentos sirven como materia prima para sintetizar otras más
complejas desde el punto de vista estructural o moléculas funcionales como las
proteínas musculares, hormonas y enzimas.
Ciertas moléculas
de los alimentos se almacenan para su uso futuro. Por ejemplo, el
glucógeno se guarda en las células del hígado y los triglicéridos en las
células adiposas.
En este capítulo se estudia la
manera en que las reacciones metabólicas liberan la energía química de los
alimentos; como contribuye cada grupo de dichas moléculas al crecimiento, a la
reparación y a las necesidades energéticas del cuerpo, y la forma en que se
mantiene el equilibrio de calor y energía del organismo. Por último, se
analizan algunos aspectos de la nutrición.
REACCIONES
METABÓLICAS
OBJETIVO
Explicar el
papel del ATP en el anabolismo t el catabolismo.
El término metabolismo
comprende todas las reacciones químicas del cuerpo. Se trata de un proceso por
el cual se establece el equilibrio de energía entre reacciones catabólicas, que
son descomposición, y las anabólicas, que corresponden a la síntesis. En
conjunto, las reacciones catabólicas son exergónicas, es decir, producen
más energía de la que consumen. En contraste, las reacciones anabólicas son endergónicas,
pues consumen más energía de la que generan. La molécula que participa con
mayor frecuencia en los intercambios de energía en células vivas es el ATP (adenosintrifosfato),
que acopla las reacciones catabólicas que liberan energía a las anabólicas que
la necesitan.
Las reacciones metabólicas que se
llevan a cabo dependen de la enzima que se active en una célula específica en
un momento particular. A menudo, las reacciones catabólicas se realizan en un
compartimiento de una célula, por ejemplo, la mitocondria, en tanto que las de
síntesis ocurren en otro sitio, como el retículo endoplásmico.
Una molécula sintetizada en una reacción anabólica tiene una vida limitada. Con pocas excepciones, finalmente se desintegra y sus átomos componentes se reciclan en otras moléculas o se excretan del cuerpo. El reciclamiento de moléculas biológicas ocurre continuamente en los tejidos vivos, en unos con rapidez y en otros lentamente. Las células individuales pueden ser restauradas molécula a molécula o se puede reconstruir todo un tejido célula por célula.
Definición de catabolismo y
anabolismo
Se llama catabolismo a las reaccione
químicas por medio de las cuales las moléculas orgánicas complejas se
descomponen en otras más simples. Dichas reacciones liberan la energía química
almacenada en esas moléculas orgánicas. En la glucólisis, el ciclo de Krebs y
la cadena de transporte de electrones tienen lugar grupos importantes de
reacciones catabólicas, que se analizan posteriormente en este capítulo.
Las reacciones químicas que combinan
moléculas simples y monómeros para formar los complejos elementos estructurales
y funcionales se conocen en conjunto como anabolismo. Entre los complejos de
este tipo de reacciones están la formación de enlaces peptídicos entre
aminoácidos durante la síntesis de proteínas, la integración de ácidos grasos
en los fosfolípidos que forman las bicapas de la membrana plasmática y la unión
de monómeros de glucosa para formar glucógeno.
Las reacciones químicas de los
sistemas vivos dependen de la transferencia de cantidades adecuadas de energía
de una molécula a otra. De éstas, la que con mayor frecuencia ejecuta dicha
tarea es el ATP, que constituye la fuente de energía fundamental de una célula
viva y siempre está disponible para que se lleven a cabo actividades celulares;
se gaste y reconstituye una y otra vez. Una célula típica posee alrededor de
mil millones de moléculas de ATP, cada una de las cuales de manera
característica, dura menos de un minuto antes de ser utilizada. Por tanto, esta
molécula no es una forma de energía que se pueda almacenar por largo plazo,
sino que se puede usar en todo momento.
El lector recordará que en el
capítulo 2 se mencionó que una molécula de ATP consta de una molécula de
adenina, una de ribosa y tres grupos fosfatos unidos entre sí. Cuando el grupo
fosfato terminal se desprende del ATP en las reacciones anabólicas, se forma
tanto difosfato de adenosina (ADP) como un grupo fosfato (simbolizado por una
]). Parte de la energía liberada se usa
para generar reacciones anabólicas como la formación de glucógeno a partir de
glucosa. Subsecuentemente, la energía de las moléculas complejas se emplea en
reacciones catabólicas para combinar ADP con un grupo fosfato y sintetizar ATP:
ADP +
]
+ energía ![]() ATP
ATP
TRANSFERENCIA DE
ENERGÍA
OBJETIVOS
Describir las
reacciones de oxidación-reducción.
Explicar el
papel del ATP en el metabolismo.
Varias reacciones catabólicas alimentan los enlaces
fosfato de alta energía del ATP. Si bien la potencia energética de estas
uniones no es excepcionalmente grande, puede liberarse con rapidez y facilidad. Antes de analizar las vías
metabólicas es preciso considerar dos importantes aspectos de la transferencia
de energía: las reacciones oxidación reducción y los mecanismos que generan
ATP.
Reacciones oxidación-reducción
La oxidación es un proceso por el cual se extraen electrones de un átomo o una molécula; como resultado disminuye la energía potencial de uno y otra. Puesto que la mayor parte de las reacciones biológicas de oxidación conllevan la pérdida de átomos de hidrógeno, se les denomina reacciones de deshidrogenación. Un ejemplo de una reacción oxidativa es la conversión del ácido láctico en ácido pirúvico:
COOH
HC OH
![]()
CH3
Ácido
láctico
C O
Ácido pirúvico
COOH + - COOH
| añade 2H (H +
H)
|
C ==O
¾¾¾¾¾¾¾→ HC
¾OH
| (reducción)
|
CH3
CH3
Ácido
pirúvico Ácido
láctico
Cuando se oxida una
sustancia, los átomos de hidrógeno liberados no permanecen en la célula, sino
que son transferidos de inmediato por coenzimas a otro compuesto. Las células
animales suelen emplear dos coenzimas para transportar átomos de hidrógeno: dinucleótido
adenina nicotidamida (NAD+), un derivado de vitamina B niacina, y dinucleótido flavina adenina
(FAD) un derivado de vitamina B2 (riboflavina). Los estados de oxidación y
reducción de NAD+ y FAD se pueden
representar de la manera siguiente:
+
-
+2H (H +
H)
NAD +
NADH +
H-
oxidado
+ - reducido
-2H (H +
H)
+2H (H +
H)
FAD
FADH2
oxidado + - reducido
-2H (H
+ H)
Las
reacciones de oxidación y reducción siempre están acopladas; cada vez que una
sustancia se oxida, de manera simultánea
otra se reduce. Estos procesos pareados se denominan reacciones
oxidación reducción o redox. Por ejemplo, cuando el ácido láctico se
oxida para formar ácido pirúvico, los dos átomos de hidrógeno retirados en la
reacción se usan para reducir NAD+. Esta reacción redox aplicada puede
escribirse de la manera siguiente:
Ácido
láctico
NAD
reducido
oxidado
![]()
![]()
![]() +
+
Ácido
pirívico
NADH + H
oxidado
reducido
Parte de la
energía liberada en las reacciones de oxidación queda atrapada dentro de la
célula cuando se forma ATP. Poco después un grupo fosfato
] se añade al ADP, con ingreso de
energía, para formar ATP. Los dos enlaces fosfato de alta energía que pueden
usarse para transformarla se indican con:
Adenosina + energía →
ADP
Adenosina
La fosforilación en el nivel de sustrato genera ATP
al transferir un grupo de fosfato de alta energía de un compuesto metabólico
fosforilado intermedio (un sustrato) directamente al ADP. En las células
humanas este proceso tiene lugar en el citosol.
La fosforilación oxidativa retira electrones de
compuestos orgánicos y los pasa a través de un aserie de aceptores de
electrones que en conjunto reciben el nombre de cadena de transporte de
electrones a moléculas de oxígeno (O2). Este proceso ocurre en la parte interna
de la membrana mitocondrial de las células.
La fosforilación sólo tiene lugar en las células
vegetales que contienen clorofila o en ciertas bacterias que poseen otros
pigmentos que absorben la energía de la luz solar.
METABOLISMO DE CARBOHIDRATOS
OBJETIVO
Describir destino, metabolismo y funciones de los
carbohidratos.
Durante la
digestión, los polisacáridos y disacáridos se hidrolizan en monosacáridos,
glucosa (alrededor de 80%), galactosa y fructosa (parte de ésta se convierte en
glucosa a medida que la absorben las células del epitelio intestinal). Los
capilares de los vellos del intestino delgado captan estos tres monosacáridos y
los llevan al hígado a través de la vena porta. Los hepatocitos (células
hepáticas) convierten la mayor parte del resto de la fructosa y prácticamente
toda la galactosa en glucosa. Por tanto, se puede decir que el metabolismo de
los carbohidratos es el mismo es el mismo que el de la glucosa. Puesto que
sistemas de retroalimentación negativa mantienen las concentraciones de glucosa
en sangre en proporciones de 90mg/mL de plasma (5mmol/litro) normalmente
circula en la sangre un total de 2 a 3 g de glucosa.
Destino de
la glucosa
Puesto que
la glucosa es la fuente de energía que más utiliza el cuerpo para sintetizar ATP,
el destino de la glucosa absorbida en la dieta depende de las necesidades
energéticas de las células del cuerpo, que la emplean para:
Producir ATP. En las células que requieren energía
inmediata, la glucosa se oxida para producir ATP, la que no se requiere de
inmediato para esta función puede emplearse en alguna de las diversas vías
metabólicas.
Síntesis de aminoácidos. Las células de todo el
cuerpo pueden utilizar glucosa para formar varios aminoácidos, los cuales después
se incorporan a las proteínas.
Síntesis de glucógeno. Los hepatocitos y las fibras
musculares pueden efectuar la glucogénesis, en la cual se combinan
cientos de monómeros glucosa para formar el polisacárido glucógeno. La
capacidad total de almacenamiento de glucógeno es de unos 125 g en el hígado y
375 g en los músculos esqueléticos.
Síntesis de triglicéridos. Cuando los sitios de
reserva de glucógeno se llenan, los hepatocitos pueden transformar la glucosa
en glicerol y ácidos grasos, que después se usan para la lipogénesis, es
decir, la síntesis de triglicéridos, los cuales a continuación se depositan en
el tejido adiposo, que virtualmente tiene una capacidad ilimitada de
almacenamiento.
Movimiento
de la glucosa en las células
Antes que
las células del cuerpo puedan utilizar la glucosa, ésta primero debe pasar a
través de la membrana plasmática y entrar al citosol. En tanto que la absorción
de la glucosa en el CG (y los túmulos del riñón) tiene lugar por medio de
transporte activo secundario (Na+ -contransportadores de glucosa) su movimiento
desde la sangre al interior de la mayor parte de las otras células del cuerpo
se llevan a cabo mediante moléculas GluT, una familia de transportadores de
glucosa por difusión facilitada (véase capitulo 3). La hormona insulina
incrementa la introducción de GluT, en las membranas plasmáticas de casi todas
las células del cuerpo, con lo que aumenta la velocidad de difusión facilitada
de glucosa en la célula. Sin embargo, en neuronas y hepatocitos, siempre hay un gran número de moléculas GlutT en la membrana del plasma, de modo que la entrada de glucosa es un
proceso continuo. Al entrar a la célula, la glucosa sufre fosforilación. Puesto que las moléculas
GluT no pueden transportar glucosa
fosforilada, esta reacción la atrapa dentro de la célula.
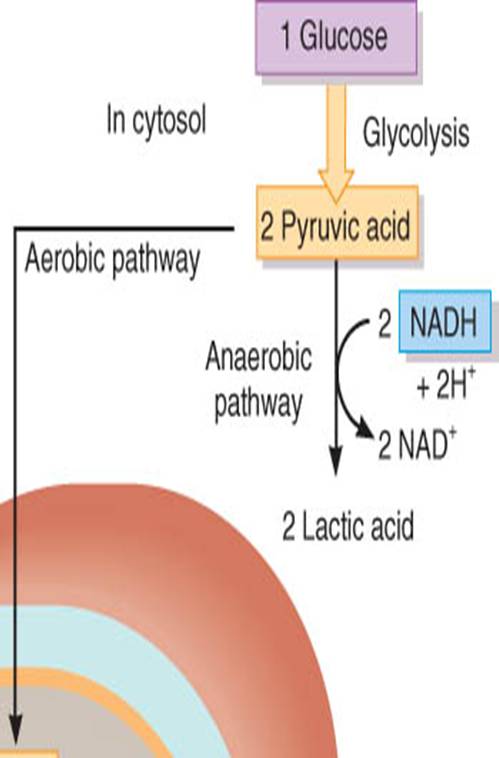
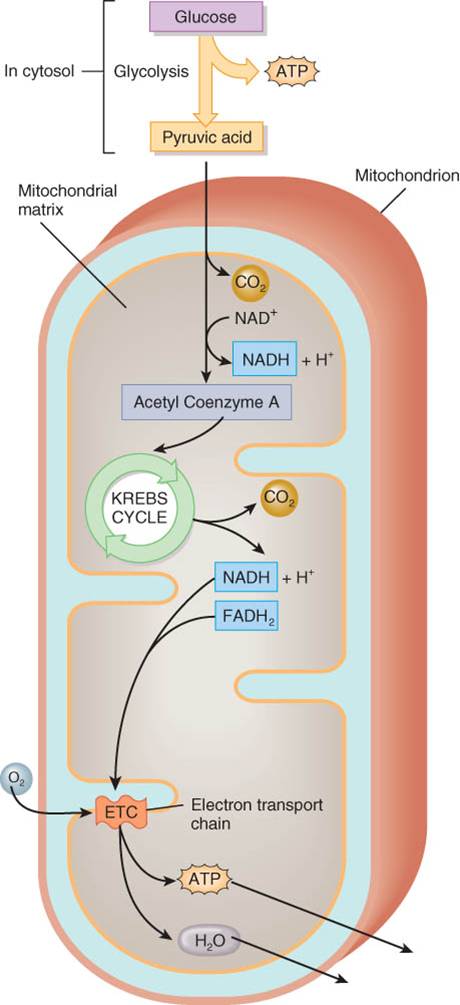
Catabolismo de la glucosa
La
oxidación de la glucosa para producir ATP también se conoce como respiración
celular; y comprende un conjunto de cuatro reacciones, la glucólisis, la
formación de acetil coenzima A, el ciclo de Krebs y la cadena de transporte de
electrones.
La glucólisis consiste en un conjunto de reacciones
en las cuales se oxida una molécula de glucosa y se producen dos moléculas de
ácido pirúvico. Además, genera dos moléculas de ATP y dos NADH + H+
que contienen energía. Puesto que la glucólisis no requiere oxígeno, es un
proceso anaerobio (sin oxígeno) de producir ATP y se le conoce como respiración
celular anaerobia.
La formación de acetil coenzima A es un paso de
transición, en el cual se prepara el ácido pirúvico para su ingreso al ciclo de
Krebs. Este paso también produce NADH
+ H+ que contiene energía más dióxido de carbono
(CO2).
Las reacciones de ciclo de Krebs oxidan acetil
coenzima A y producen CO2, ATP, NADH
+ H+, que contienen energía y
FADH2.
Las reacciones de la cadena de transporte de electrones
oxidan NADH + H+ y FADH2 y transfieren sus electrones a
una serie de transportadores. Tanto el ciclo de Krebs como la cadena de
transporte de electrones requieren oxígeno para producir ATP y por ello este
proceso se denomina respiración celular aerobia.
Glucólisis
La glucólisis
consta de diez reacciones químicas que desintegran una molécula de glucosa con
seis carbonos en dos moléculas de tres carbonos de ácido pirúvico. (Advertencia:
no se debe confundir glucólisis con glucogenólisis, que es la
desintegración de glucógeno en glucosa). Una enzima específica cataliza cada
una de las reacciones. Los diez pasos de la glucólisis se llevan a cabo en el
siguiente orden:
En la primera reacción se fosforila glucosa mediante el uso
de un grupo fosfato de una molécula de ATP.
La segunda reacción convierte glucosa 6-fosfato en fructosa
6-fosfato.
La tercera reacción emplea un segundo ATP para añadir un
segundo grupo fosfato. La fosfofructocinasa, enzima que cataliza este
paso, es la llave que regula la velocidad de la glucólisis. Su actividad es
alta cuando la concentración de ADP es elevada, en cuyo caso se produce ATP con
rapidez, y su actividad es baja cuando la concentración de ATP es alta. Si la
actividad de fosfofructosinasa es reducida, la mayor parte de la glucosa no
participa en las reacciones de glucólisis, sino que se convierte en glucógeno
para su almacenamiento.
5) La molécula de fructosa doblemente
fosforilada se desdobla en dos compuestos de tres carbonos, gliceraldehído
3-fosfato (G 3-P) y dihidroxiacetona fosfato. Ambos compuestos pueden
convertirse el uno en el otro, pero su G -3P es sometido a oxidación adicional
para convertirlo en ácido pirúvico.
La oxidación se
presenta como dos moléculas de NAD+ que aceptan dos pares de
electrones y iones de hidrógeno procedentes de dos moléculas de G 3-P,
cada una de las cuales a su vez forma dos moléculas de NADH y de ácido
1,3 bifosfogligérico (BFG). Muchas células del cuerpo usan los dos NADH
producidos en este caso para genera cuatro ATP en la cadena de transporte de
electrones. Unos cuantos tipos de células, como los hepatocitos, las células
renales y las fibras de músculo cardiaco, pueden generar seis ATP a partir de
los dos NADH.
10) Estas reacciones generan cuatro moléculas
de ATP y producen dos moléculas de ácido pirúvico (piruvato).
En resumen,
aunque la glucólisis utiliza dos moléculas de ATP con una ganancia neta de dos
moléculas de ATP por cada una de glucosa que se oxida.
Destino
del ácido pirúvico
El destino del ácido pirúvico que se produce durante la glucólisis depende de la disponibilidad de oxígeno. Si el oxígeno es escaso (condiciones anaerobias), por ejemplo en las fibras del músculo esquelético durante ejercicio extenuante, entonces el ácido pirúvico se reduce mediante la vía aerobia por adición de dos átomos de hidrógeno para formar ácido láctico (lactato):
2 Ácido
pirúvico + 2NADH
+ 2H → 2 Ácido láctico +
2NAD
(oxidado)
(reducido)
Cuando el
oxígeno es abundante (condiciones aerobias), la mayor parte de las células
convierten ácido pirúvico en acetil coenzima A. Esta molécula vincula la
glucólisis, que ocurre en el citosol, con el ciclo de Krebs, que se lleva a
cabo en la matriz de la mitocondria. El ácido pirívico entra a la matriz
mitocondrial con ayuda de una proteína transportadora especial. Puesto que los
eritrocitos carecen de mitocondrias,
sólo pueden producir ATP a través de glucólisis.
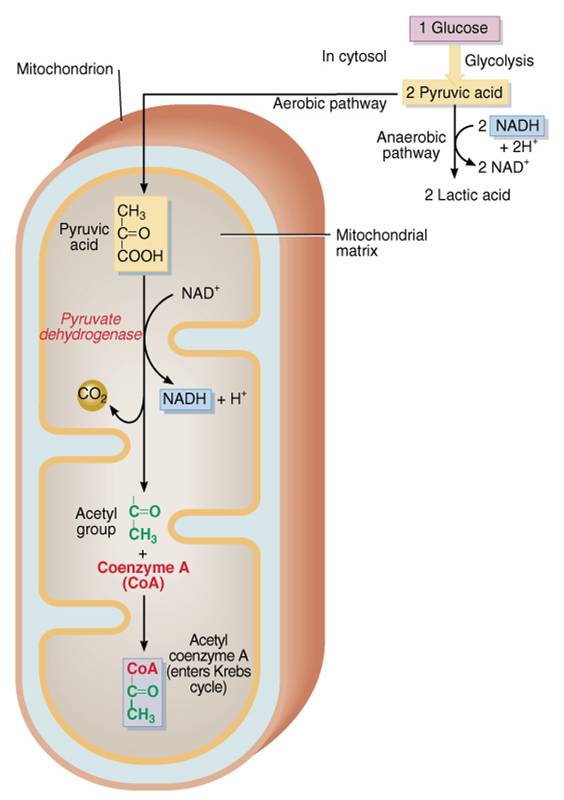
Formación
de acetil coenzima A
En
la oxidación de la glucosa cada paso requiere una enzima diferente, y a menudo
también coenzima. La que se emplea en este punto de la respiración celular es
la coenzima A (CoA), la cual se deriva de ácido pantoténico, una
vitamina B. Durante el paso de transición entre glucólisis y ciclo de Krebs se
prepara el ácido pirúvico para que entre a dicho ciclo. La enzima deshidrogenasa
de piruvato, que se localiza de manera exclusiva en la matriz mitocondrial,
convierte piruja en un fragmento de dos carbonos mediante el retiro de una
molécula de dióxido de carbono. La pérdida de una molécula de CO2 por una
sustancia se llama descarboxilación. Ésta es la primera reacción de la
respiración celular que libera CO2. Durante ella también se oxida el ácido
pirúvico. Cada molécula de éste ácido pierde dos átomos de hidrógeno en la
forma de un ión hidruro (H-) más un ión hidrógeno (H+). La coenzima NAD+ se
reduce conforme capta el H del ácido pirúvico; el H+ se libera en la matriz
mitocondrial. Debe recordarse que la oxidación de una molécula de glucosa
produce dos de ácido pirúvico, de modo que por cada molécula de glucosa, se
pierden dos de dióxido de carbono y se producen dos de NADH + H+.
El fragmento de dos carbonos, llamado grupo acetil se une a la coenzima
y todo el complejo se denomina acetil coenzima A (acetil CoA). Una vez
que el ácido pirúvico ha sufrido descarboxilación y el resto del grupo se
enlaza a CoA el compuesto resultante (CoA) se incorpora rápidamente al ciclo de
Krebs.
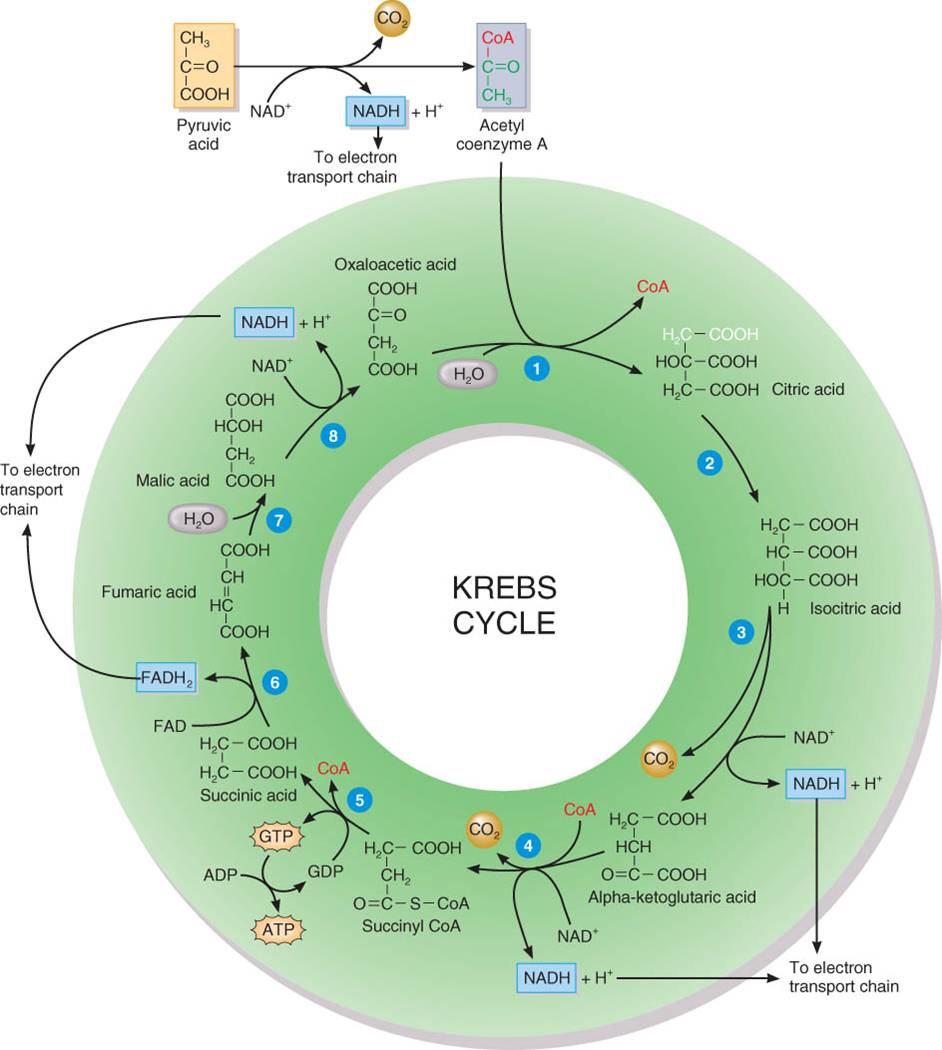
Ciclo
de Krebs
EL ciclo
de Krebs-así llamado por el bioquímico Hans Krebs, quien describió estas
reacciones en el decenio de 1930 también se conoce como el ciclo del ácido
nítrico, por la primera molécula que se forma cuando un grupo acetil se une
al ciclo. En su mayor parte, el ciclo de Krebs es una serie de reacciones de oxidación reducción y de descarboxilación
cada una cataliza por una enzima específica en la matriz de la mitocondria.
En el ciclo
de Krebs, la gran cantidad de energía potencial química almacenada en sustancias
intermedias derivadas de ácido pirúvico se libera de manera gradual. Las
reacciones de oxidación reducción transfieren energía química en forma de
electrones, a dos coenzimas NAD+ y FAD. Los derivados del ácido pirúvico se
oxidan, en tanto que las coenzimas se reducen. Además, en un paso se genera
ATP. Cada reacción descrita en el texto sea relaciona con las numeradas en la
figura 25.5 a. La enzima que cataliza cada reacción aparece entre paréntesis.
{kind=link}
Entrada del grupo acetil. La unión química que une el
grupo acetil a la coenzima A (CoA) se rompe, y el grupo acetil de dos carbonos
se enlaza a una molécula de cuatro carbonos de ácido oxalacético para formar
una de seis carbonos denominada ácido cítrico. La CoA queda libre para
combinarse con otro grupo acetil de ácido pirúvico y repetir el proceso (sintasa
cítrica).
Isomerización. El ácido cítrico sufre este proceso
para convertirse en ácido isocítrico que tiene la misma fórmula molecular que
el citrato. Nótese, sin embargo, que el
grupo hidroxilo (-OH) se encuentra unido a un carbono diferente (aconitasa).
Descarboxilación oxidativa. El ácido isocítrico se
oxida y pierde una molécula de CO2, forma una molécula de cinco carbonos
llamada ácido cetoglutárico-alfa. El H- de la oxidación se pasa a un NAD+, que
se reduce a NADH + H+. Ésta es la segunda reacción celular que
libera CO2 (deshidrogenasa isocírtica).
Descarboxilación oxidativa. El ácido cetoglutárico-alfa
se oxida, pierde una molécula de CO2 y capta CoA para formar una de cuatro
carbonos llamada succinil CoA. Se trata de la tercera reacción en la
respiración celular que libera Co2 (deshidrogenasa cetoglutarato-alfa).
Fosforilación en el nivel de sustrato. En este punto
del ciclo, ocurre una fosforilación en el nivel de sustrato. CoA es desplazada
por un grupo fosfato, que luego se transfiere a difosfato de guanosina (GDP)
para formar trifosfato de guanosina (GTP), que es similar al ATP y puede donar
un grupo fosfato al ADP para integrar ATP. Estas reacciones también convierten
succinil CoA en ácido succínico (sintetasa succinil CoA).
Deshidrogenación. El ácido succínico se oxida en
ácido fumárico cuando dos de sus átomos de hidrógeno de transfieren a la
coenzima nucleótido flavina adenina (FAD), que se reduce a FADH2 (deshidrogenasa
succinato).
Hidratación. El ácido fumárico se convierte en ácido
málico por adicción de una molécula de agua (fumarasa).
Deshiodrogenación. En el paso final del ciclo, el
ácido málico se oxida para volver a formar ácido oxilacético. En el proceso se
retiran dos átomos de hidrógeno y uno se transfiere a NAD+, que así se reduce a
NADH +
H+. El ácido oxalacético regenerado puede combinarse con otra molécula
de acetil CoA e iniciar un nuevo ciclo (deshidrogenasa málica).
Las
coenzimas reducidas (NADH y FADH2) constituyen el producto más importante en el
ciclo de Krebs puesto que contienen la energía almacenada originalmente en la
glucosa y luego en el ácido pirúvico. En total, por cada acetil CoA que entra
al ciclo de Krebs, se producen tres NADH tres H+ y un FADH2 mediante reacciones
de oxidación-reducción y se genera una molécula de ATP por fosforilación en el
nivel de sustrato (fig. 25.5b). En la cadena de transporte de electrones, los
tres NADH + tres H+ producen más tarde
nueve moléculas de ATP y el FADH2 genera posteriormente dos moléculas de ATP.
Así, cada que concluye un ciclo de Krebs finalmente se producen 12 moléculas de
ATP. Dado que cada molécula de glucosa suministra dos acetil CoA, el
catabolismo de la glucosa por la vía del ciclo de Krebs y la cadena de
transporte de electrones rinde 24 moléculas de ATP por molécula de glucosa. La
liberación de CO2 tiene lugar conforme el ácido pirúvico se convierte en acetil
CoA y durante las dos reacciones de descarboxilación del ciclo de Krebs (véase
fig 25.5b). Pero en virtud de que cada molécula de glucosa genera dos de ácido pirúvico, se libera un total de seis moléculas de CO2 por cada molécula de
glucosa original catabolizada a lo largo de esta vía. Las moléculas de CO2
abandonan las mitocondrias, difunden a través del citosol hacia la membrana del
plasma y luego pasan a la sangre. Con el tiempo, la sangre transporta el CO2 a
los pulmones, que lo exhalan.
{kind=link}
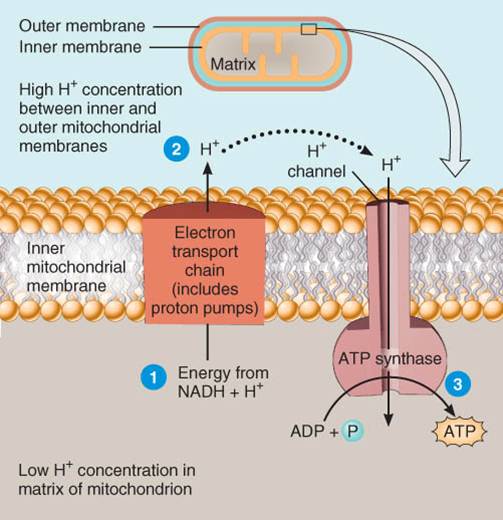
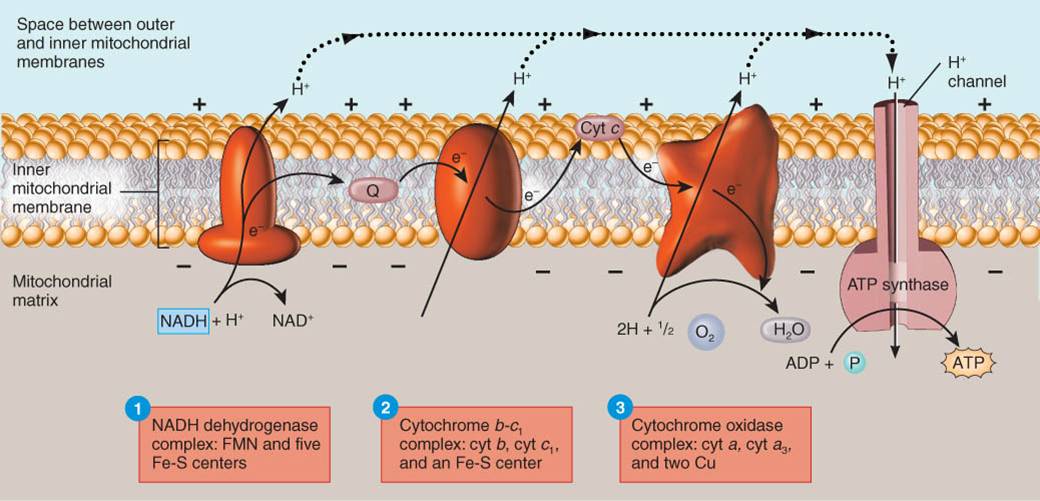
Cadena
de transporte de electrones
La
cadena de transporte de electrones es una serie de transportadores de
electrones que son proteínas integradas a la membrana mitocondrial interna, la
cual se halla plegada en crestas que incrementan el área de superficie, y de
ésta manera acomodan miles de copias de la cadena de transporte en cada
mitocondrión. En dicha cadena, cada transportador se reduce conforme capa
electrones y se oxida a medida que libera electrones. Cuando éstos pasan a
través de la cadena, las reacciones exergónicas liberan pequeñas cantidades de
energía en una serie de pasos y esta energía se emplea para formar ATP. En la
respiración celular aerobia, el receptor final de electrones de la cadena es el
oxígeno. Dado que este mecanismo de generación de ATP vincula reacciones
químicas (el paso de electrones a lo largo de la cadena de transporte) con el
bombeo de iones hidrógeno, se le llama quimiósmosis. En resumen, la
quimiósmosis funciona de la manera siguiente (fig. 25.6):
{kind=link}
La energía de NADH
+ H+ pasa a lo largo de la cadena
de electrones y se usa para bombear H+
(protones ) desde la matriz del
mitocondrión al interior del espacio situado entre las membranas mitocondriales
interna y externa.
H+ se acumula en concentración elevada entre las membranas mitocondriales
interna y externa.
A continuación tiene lugar la síntesis de ATP conforme los
iones hidrógeno difunden de regreso al interior de la matriz mitocondrial a
través de un tipo especial de canal de H+ en la membrana interna.
Primero se
examinarán los transportadores de electrones y posteriormente se analizará en
detalle la quimiósmosis.
Transportadores de electrones. Varios tipos de
moléculas y átomos sirven como transportadores de electrones:
Mononucleótido flavina (FMN), al igual que FAD (dinucleótido
flavina adenina), es una flavoproteína derivada de riboflavina (vitamina B2).
Citocromos. Son proteínas con un grupo que contiene
hierro (hem) capaz de existir de manera alternada en una forma reducida (Fe2+)
y una oxidada (Fe3+). Los distintos citocromos que participan ene.
transporte de la cadena de transporte de electrones son citocromo b (cit
b), citocromo c1 (cit c1), citocromo
c (cit c), citocromo a (cit a) y citocromo a3
(cit a3).
Los centros hierro-azufre (Fe-S) contienen dos o
cuatro átomos de hierro que se unen a átomos de azufre para formar un centro de
transferencia de electrones de una proteína.
Los átomos de cobre (Cu) enlazados a dos proteínas en
la cadena también participan en la transferencia de electrones.
La coenzima Q, es un transportador no proteína, de
bajo peso molecular, móvil en la bicapa lipídica de la membrana interna.
Pasos en el transporte de electrones y la generación
quimismótica de ATP. En el interior de la membrana mitocondrial
interna, los transportadores de la cadena de transporte de electrones se
agrupan en tres complejos; cada uno actúa como bomba de protones que
expulsa H+ de la matriz mitocondrial y ayuda a crear un gradiente
electroquímico de H+. Cada un a de las tres bombas de protones
constituye un complejo que transporta electrones y bombea H+, de la
manera siguiente (fig 25.7):
{kind=link}
La primera bomba de protones es el complejo
deshidrogenasa NADH que contiene mononucleótido flavina y cinco o más
centros Fe-S. NADH + H+ se oxida a NAD+ y
FMN se reduce a FMNH2, que a su vez se oxida conforme pasa
electrones a los centros hierro, azufre, que tiene movilidad en la membrana,
pasa electrones al segundo complejo bomba.
La segunda bomba de protones es el complejo citocromo b-c1,
que contiene citocromos y un centro hierro azufre. Los electrones pasan
sucesivamente de Q a cit b, a Fe-S, a cit c1. El Q
móvil (de ida y vuelta) que pasa electrones del segundo complejo bomba al
tercero es el citocromo c (cit c).
La tercera bomba de protones es el complejo citocromo
oxidasa que contiene citocromos a y a3 y dos átomos de cobre. Los
electrones pasan de cit c, a Cu, a cit a y por último a cit a3. éste
último transfiere electrones a la mitad de una molécula de oxígeno (O2),
que adquiere carga negativa y capta dos H+ del medio circundante
para formar H2O. Ésta es la única parte de la respiración celular
aerobia en la cual se consume O2. El cianuro es un veneno mortal
porque se une al complejo citocromo oxidasa y bloque éste último paso del
transporte de electrones.
El bombeo
de H+ produce un gradiente concentración de protones y un gradiente
eléctrico. El incremento de H+ en untado de lado de la membrana
mitocondrial interna le confiere a esta parte una carga positiva en comparación
con el otro. El gradiente electroquímico resultante posee energía potencial,
denominada fuerza motriz protónica. Dentro de la membrana mitocondrial
interna se encuentran canales para protones que permiten la difusión de H+
de regreso a través de la membrana, impulsado por una fuerza motriz protónica,
conforme H+ vuelve a la membrana, genera ATP debido a que los
canales H+ también incluyen una enzima llamada sintasa ATP,
al cual usa la fuerza motriz protónica para sintetizar ATP a partir de ADP
y
].
Este proceso de quimiósmsis explica casi todo el ATP producido durante la
respiración.
Resumen
de la respiración celular
Las
distintas transferencias de electrones en la cadena de transporte generan 32 o
34 moléculas de ATP por cada molécula de glucosa oxidada: 28 a 30+ a
partir de 10 moléculas de NADH + H+ y dos de cada una de las dos
moléculas de FADH2 (cuatro en total). Por tanto, durante la
respiración celular, se pueden generar o
38 ATP de una molécula de glucosa. Nótese que dos de estos ATP provienen de la
fosforilación en el nivel de sustrato en la glucólisis, y dos de la
fosforilación en el nivel de sustrato en el ciclo de Krebs. La reacción total
es:
C6H12O6
+ 6 O2 + 36 o 38 ADP +
36 o 38
]
Glucosa Oxígeno
![]() 6 CO2 + 6 H2O + 36
o 38 ATP
6 CO2 + 6 H2O + 36
o 38 ATP
Dióxido Agua
de carbono
El cuadro
25.1 muestra un resumen del rendimiento de ATP durante la respiración celular.
En la figura 25.8 aparece un esquema de las principales reacciones de la
respiración celular. El verdadero rendimiento de ATP puede ser menor de 36 a 38
ATP, por molécula de glucosa. Se desconoce el número exacto de H+
que debe ser bombeado hacia fuera para generar un ATP durante la quimiósmosis.
Más aún, el ATP generado en la mitocondria ha de transportarse fuera de estos
organelos hacia el citosol para su uso en cualquier otra parte de la célula. La
salida de ATP en el intercambio que se realiza en el movimiento de entrada del
ATP que se forma a partir de las reacciones metabólicas en el citosol, utiliza
parte de la fuerza motriz protónica.
{kind=link}
La
glucólisis, el ciclo de Krebs y en especial la cadena de transporte de
electrones suministran todo el ATP para las actividades de la célula y dado que
los dos últimos procesos son aeróbicos, las células no pueden realizar sus
actividades por mucho tiempo sin un suministro suficiente de oxígeno.
APLICACIÓN CLÍNICA
Carga de carbohidratos
La cantidad
de glucógeno almacenado en el hígado y los músculos esqueléticos varía y puede
agotarse por completo durante procesos atléticos prolongados. Así, muchos
corredores del maratón y otros atletas de resistencia siguen un régimen preciso
de ejercicio y dieta que incluye la ingestión de grandes cantidades de
carbohidratos complejos, como pasta y patatas, en los tres días previos a la
competencia. Esta práctica, llamada carga de carbohidratos, tiene por
objeto incrementar al máximo la concentración de glucógeno disponible en el
músculo para producir ATP. Se ha demostrado que la carga de carbohidratos
incrementa la resistencia del deportista en las justas atléticas que duran más
de una hora
Los NADH producidos en el citosol durante la glucólisis no
pueden entrar a la mitocondria. En vez de ello donan sus electrones a una de
dos moléculas de transferencia, conocidas como malato de transferencia y
fosfato de glicerol de transferencia. En las células del hígado, riñón y
corazón, el uso del malato de transferencia da como resultado la síntesis de
tres ATP por cada NAH. En otras células del cuerpo, como fibras del músculo
esquelético y neuronas, el uso de fosfato de glicerol de transferencia da como
resultado la síntesis de dos ATP por cada NADH.
Cuadro 25.1 Resumen de ATP producido en la
respiración celular.
|
FUENTE
|
RENDIMIENTO DE ATP (RENDIMIENTO) |
|
Glucólisis |
|
|
Oxidación de una molécula de glucosa a dos moléculas de
ácido pirúvico |
2 ATP (fosforilación en el nivel de sustrato) |
|
Producción de 2 NADH + H+ |
4 a 6 ATP (fosforilación oxidativa en la cadena de
transporte de electrones) |
|
Formación de acetil coenzima A |
|
|
2 NADH + H+ |
6 ATP (fosforilación oxidativa en la cadena de transporte
de electrones) |
|
Ciclo de Krebs y cadena de transporte de electrones |
|
|
Oxidación de succinil CoA en ácido succínico |
2 GTP que se convierten en 2 ATP (fosforilación en el
nivel de sustrato) |
|
Producción de 6 NADH + 6H+ |
18 ATP (fosforilación oxidativa en la cadena de transporte
de electrones) |
|
Producción de 2 FADH2 |
4 ATP (fosforilación oxidativa en la cadena de transporte
de electrones) |
|
Total |
36 o 38 ATP por molécula de glucosa (máximo teórico) |
La mayor
parte de la glucosa en el cuerpo se cataboliza para generar ATP, pero la
glucosa puede tomar parte en varias reacciones anabólicas. Una es la síntesis
de glucógeno; otra es la de nuevas moléculas de glucosa a partir de alguno de
los productos del desdoblamiento de proteínas y lípidos.
Almacenamiento
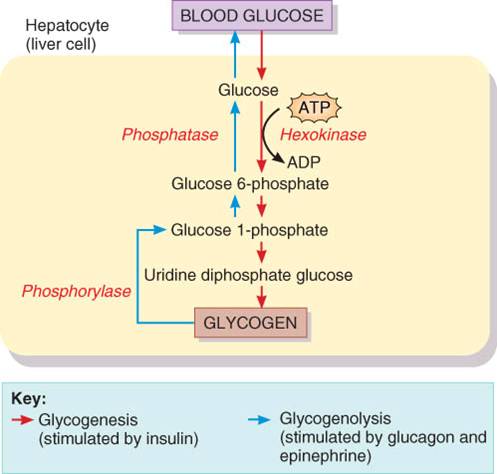
de glucosa: glucogénesis.
Si no se
necesita de inmediato la glucosa para producir ATP, se combina con muchas otras
moléculas de glucosa a fin de formar el polisacárido glucógeno. En el proceso
de glucogénesis (fig. 25.9), la
glucosa que entra a los hepatocitos o a las células del músculo esquelético es
fosforilada por la acción de la hexocinasa y se convierte en glucosa 6-fosfato,
que luego se transforma en glucosa 1-fosfato, después en difosfato glucosa
uridina y por último en glucógeno. La insulina de las células beta del páncreas
estimula la glucogénesis.
{kind=link}
Liberación
de glucosa: glucogenólisis
Cuando las
actividades del cuerpo requieren ATP, el glucógeno almacenado en los
hepatocitos se desdobla en glucosa que se libera en la sangre para que sea
transportada a las células, donde se cataboliza. El proceso por el cual el
glucógeno se convierte de nuevo en glucosa se denomina glucogenólisis, que no es la simple revisión de los pasos de la
glucogénesis (véase
fig. 25.9).
Principia cuando las moléculas de glucosa se separan de la molécula ramificada
de glucógeno por medio de fosforilación para formar glucosa 1-fosfato. Tanto
las hormonas glucagon de las células alfa del páncreas como la adrenalina de la
médula suprarrenal activan la
fosforilasa, la enzima que cataliza esta reacción. Enseguida, la glucosa
1-fosfato se convierte en glucosa 6-fosfato y por último en glucosa. Dado que
la fosfatasa, enzima que transforma la glucosa 6-fosfato en glucosa, se
encuentra en los hepatocitos pero no en las células del músculo esquelético,
aquellos pueden liberar la glucosa sderivada del glucógeno en el torrente
sanguíneo, en tanto que las células del músculo esquelético no realizan esta
función. En estas últimas, el glucógeno se desdobla en glucosa 1-fosfato, que
luego se cataboliza para producir ATP por medio de glucólisis y el ciclo de
Krebs. Sin embargo, el hígado tiene la capacidad de convertir en glucosa el
ácido láctico producido por la glucólisis en las células del músculo. De esta
manera, el glucógeno del músculo puede ser una fuente indirecta de glucosa
sanguínea.
Formación
de glucosa a partir de proteínas y grasas: gluconeogénesis
Cuando el
suministro de glucógeno hepático es bajo, las células del cuerpo comienzan a
catabolizar más triglicéridos y proteínas. Aunque en condiciones normales
siempre se cataboliza algo de triglicéridos y proteínas, el desdoblamiento de
unos y otras a gran escala no ocurre a menos que el individuo esté en ayunas,
en estado de inanición, coma alimentos con muy pocos carbohidratos o sufra
alguna enfermedad endocrina.
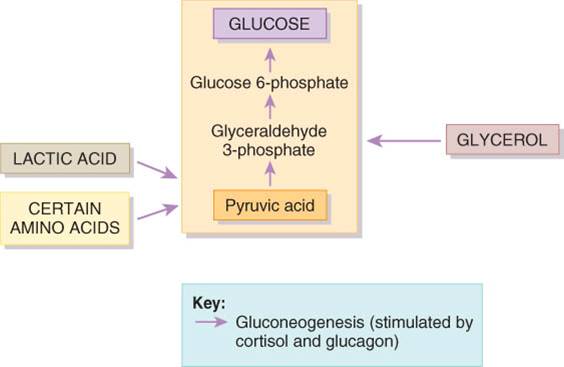
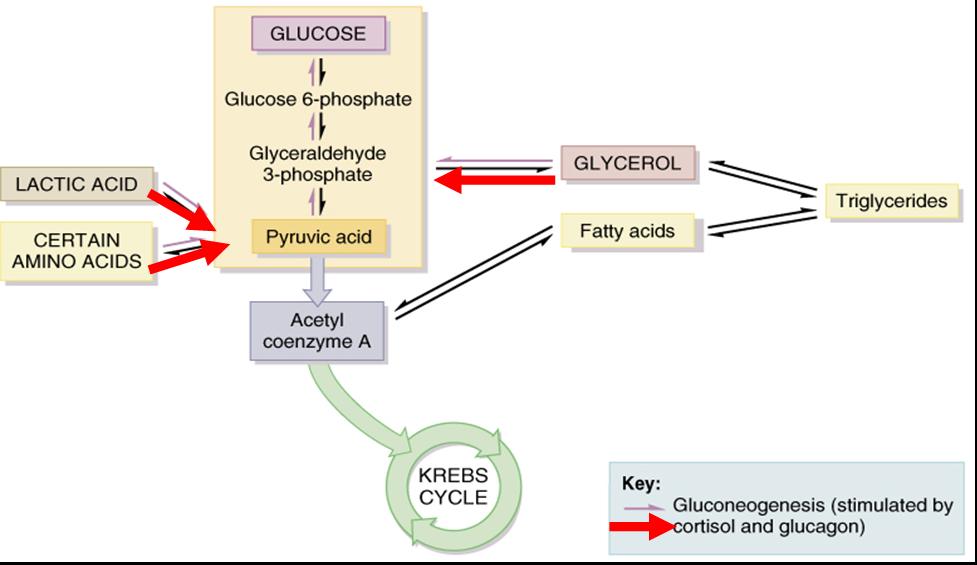
Ciertas moléculas pueden fragmentarse y convertirse en glucosa en el hígado. El proceso mediante el cual se forma glucosa nueva a partir de fuentes que no son carbohidratos recibe el nombre de gluconeogénesis. Es posible usar ácido láctico, ciertos aminoácidos y la porción glicerol de las moléculas de triglicéridos para formar nuevas moléculas de glucosa a través de la gluconeogénesis (fig. 25.10). Alrededor de 60% de los aminoácidos del cuerpo pueden sufrir esta conversión. Los aminoácidos como alanita, cisterna, glicina, serina y treonina, junto con el ácido láctico se convierten en ácido pirúvico, el cual puede entonces sintetizarse en glucosa o entrar al ciclo de Krebs. El glicerol se convierte entonces en gliceraldehído 3-fosfato, el cual forma ácido pirúvico o se emplea para sintetizar glucosa. El cortisol, principal hormona glucocorticoide de la corteza suprarrenal y el glucagon del páncreas estimulan la gluconeogénesis. Además el primero estimula el desdoblamiento de proteínas en aminoácidos, con lo que amplia la reserva disponible de éstos para la gluconeogénesis. Las hormonas tiroideas (tiroxina y triyodotironina) también movilizan proteínas y tienen la capacidad de hacerlo con triglicéridos del tejido adiposo; de esta manera hacen que el glicerol quede disponible para la gluconeogénesis.
{kind=link}
METABOLISMO
DE LOS LÍPIDOS
OBJETIVOS
Conocer las
lipoproteínas que transportan los lípidos en la sangre.
Explicar el destino,
el metabolismo y las funciones de los lípidos.
Transporte de lípidos
por lipoproteínas.
La mayor
parte de los lípidos, como el colesterol y los triglicéridos, son moléculas no
polares (y por tanto resultan muy
hidrófobas). Por ello, para que puedan ser transportadas en la sangre acuosa
primero deben hacerse hidrosolubles al combinarse con las proteínas producidas
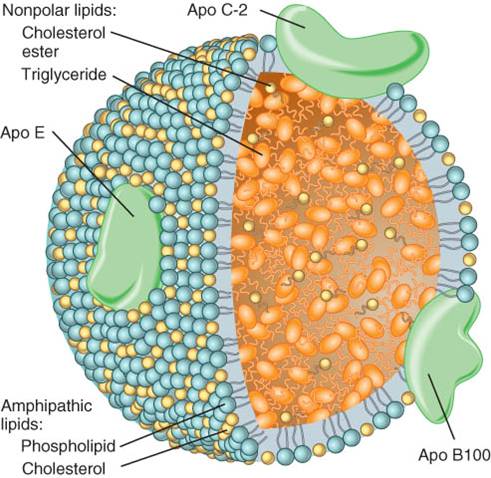
por el hígado y el intestino. Las combinaciones así formadas son las lipoproteínas, partículas específicas
que contienen cientos de moléculas (fig. 25.11). En una lipoproteína, un núcleo
interno de triglicérido hidrófobo y de moléculas de éster de colesterol se
encuentra rodeado por una capa externa de proteínas polares, además de
fosfolípidos antipáticos y moléculas de colesterol. Las proteínas de la
cubierta externa se denominan apoproteínas
(apo) y se identifican con las letras A, B, C, D y E más un número. Además
de ayudar a solubilizar la lipoproteína en los lípidos del cuerpo, cada
aporpoteína también realiza funciones específicas.
{kind=link}
Hay varios
tipos de lipoproteínas, cada uno de los cuales tiene funciones diferentes, pero
prácticamente todas son vehículos de transporte: proporcionan una especie de
servicio de paquetería, es decir, reciben y entregan, de modo que los distintos
tipos de lípidos se hallan disponibles para las células que los necesitan o se
retiran de la circulación cuando no se requieren. Las lipoproteínas se
clasifican y denominan principalmente de acuerdo con su densidad, la cual varía
con la proporción de lípidos (de baja
densidad) en relación con las proteínas (de alta densidad). De la más grande y
ligera a la más pequeña y pesada, las cuatro clases principales de
lipoproteínas son quilomicrones, lipoproteínas de muy baja densidad (VLDL),
lipoproteínas de baja densidad (LDL) y lipoproteínas de alta densidad (HDL).
Los quilomicrones se forman en las células
epiteliales de la mucosa del intestino delgado y contienen lípidos exógenos (de la dieta). Asimismo poseen
alrededor de 1 2 % de proteínas, 85% de triglicéridos, 7% de fosfolípidos y de
Las lipoproteínas de muy baja densidad (VLDL)
se forman en los hepatocitos y contienen triglicéridos endógenos. Las VLDL
están formadas aproximadamente por 10% de proteínas, 50% de triglicéridos, 20%
de fosfolípidos y 20% de colesterol. Transportan los triglicéridos sintetizados
en los hepatocitos a los adipositos donde son almacenados. Al igual que los
quilomicrones, pierden triglicéridos a medida que su apo C-2 activa la lipasa lipoproteína endotelial y los ácidos
grasos resultantes son captados por adipositos para almacenamiento y por
células musculares para producción de AYP. Después de depositar una parte de
sus triglicéridos en sus células adiposas, las VLDL se convierten en LDL.
Las lipoproteínas de
baja densidad (LDL) contienen 25% de proteínas, 5% de triglicéridos, 20% de
fosfolípidos y 50% de colesterol. Transportan cerca de 75% de colesterol total
en la sangre y lo llevan a las células de todo el cuerpo para que se utilicen
en reparación de membranas celulares, así como en la síntesis de hormonas
esteroides y sales biliares. La única proteína que contiene LDL es apo B100 y constituye la proteína que
se desprende y se une a receptores LDL para que estas lipoproteínas penetren en
el cuerpo celular por endocitosis mediada por receptor. Una vez en la célula
los LDL son desdoblados y liberan colesterol, el cual sirve para satisfacer las
necesidades de esta unidad vital. Cuando la célula tiene suficiente colesterol
para realizar sus actividades, un sistema de retroalimentación negativa inhibe
la síntesis de nuevos receptores LDL en la célula.
Cuando la
cantidad de LDL e excesiva, estas lipoproteínas también depositan colesterol
dentro y alrededor de las fibras del músculo liso en las arterias, con lo que
forman placas lípidas ateroéscleróticas que incrementan el riesgo de actividad
arteria coronaria (véase capítulo
20). Por esta razón el colesterol de los LDL, llamado colesterol LDL, se
considera malo. Algunas personas poseen muy pocos receptores LDL; por esta
razón las células de su cuerpo no pueden retirar las lipoproteínas de baja
densidad de la sangre eficazmente; como resultado, su concentración retorna muy
alta en el plasma y hay mayores posibilidades de que desarrollen placas
lipídas. Más aún, ingerir una dieta rica en grasas aumenta la producción de
VLDL, lo que eleva la concentración de LDL e incrementa la formación de placas
lipídas.
Las lipoproteínas de alta densidad (HDL) constan de
proteínas (
Origen y
significado del colesterol en la sangre
Hay dos fuentes que aportan colesterol al cuerpo. Una se
encuentra en los alimentos (huevos, productos lácteos, vísceras, carne de
bovino, cerdo y embutidos), aunque la mayor parte se sintetiza en el hígado.
Incluso los alimentos grasos que no contienen colesterol en absoluto pueden
incrementar de manera espectacular las concentraciones de éste en la sangre de
dos maneras: primero, la ingestión abundante de grasas en la dieta estimula la
reabsorción del colesterol que contiene la bilis que regresa, con lo cuales
desecha menos colesterol en las heces. Segundo, cuando las grasas saturadas se
desdoblan en el cuerpo, los hepatocitos usan una parte de los productos del
desdoblamiento para generar colesterol.
La prueba de perfil de lípidos en general mide colesterol
total (CT), colesterol HDL y triglicéridos (VLDL). A continuación se puede
calcular el colesterol LDL por medio de la siguiente fórmula: LDL = TC – HDL –
(triglicéridos/5). En Estados Unidos, el colesterol sanguíneo suele medirse en
miligramos por decilitro (mg/dL); un decilitro es 0.1 litro o 100 mL. Para los
adultos, las concentraciones deseables en sangre son: colesterol total menos de
200 mg/100mL, LDL menor de 130 mg/mL y HDL más de 40 mg/100mg/mL. Normalmente
los triglicéridos se encuentran en el intervalo de 10 a 190 mg/mL.
A medida que aumenta la concentración total de colesterol,
también es mayor el riesgo de enfermedad de arteria coronaria. Cuando el
colesterol total se eleva a más de 200mg/100mL (5.2mmol/litro), se duplica el
riesgo de un ataque cardiaco con cada 50mg/100mL (1.3 mmol/litro) de incremento
en el colesterol total. Cuado éste es de 200 a 239mg/100 mL y LDL de 130 a 159
mg/mL se encuentran en el límite alto, en tanto que un colesterol mayor de
239mg/100mL y LDL arriba de 159 se clasifica como colesterol sanguíneo alto. La
proporción entre colesterol total y colesterol HDL predice el riesgo de
desarrollar enfermedad de arteria coronaria. Por ejemplo, una persona con
colesterol, total de 180 y HDL de 60 tiene una proporción de riesgo de 3. Cuado
éste es mayor de 4 se considera que el sujeto tiene un nivel indeseable; cuando
más alta sea la proporción, mayor será el riesgo de que la persona desarrolle enfermedad
de arteria coronaria.
Hay diversos procedimientos que se utilizan para predecir la
concentración de colesterol en sangre, entre ellos, el ejercicio, la dieta y
los fármacos. La actividad física regular en niveles aeróbicos y subaeróbicos
elevan sus niveles de HDL. Los cambios en la dieta tienen como objeto reducir
la ingestión total de grasas, grasas saturadas y colesterol. Los medicamentos
usados para tratar las concentraciones elevadas
de colesterol en sangre incluyen colestiramina (Questran) y colistripol
(Colestid) que promueven la excreción de bilis en las heces; ácido nicotínico
(Liponicin), lovastatina (Mevacor) y sinvastatina (Zocor) los cuales bloquean
la enzima básica (HMGCoA reductasa) que se requiere para la síntesis de
colesterol.
Destino de
los lípidos
Los lípidos, al igual que los carbohidratos, pueden oxidarse
para producir ATP. Si el cuerpo no necesita utilizar grasas de inmediato, éstas
se almacenan en el tejido adiposo (depósito de grasa) en el hígado y en todo el
cuerpo. Unos pocos lípidos se emplean como moléculas estructurales o para
sintetizar otras sustancias esenciales. Por ejemplo, los fosfolípidos son
constituyentes de las membranas plasmáticas, las lipoproteínas transportan el
colesterol a las diferentes partes del cuerpo; la tromboplastina es necesaria
para la coagulación de la sangre, y las vainas de mielina aceleran la
conducción de los impulsos nerviosos. En el cuadro 2.7 (capítulo 2) se
describen las distintas funciones que desempeñan los lípidos en el cuerpo.
Almacenamiento
de los triglicéridos
Una función fundamental del tejido adiposo consiste en
retirar los triglicéridos de los quilomicrones y de las VLDL para almacenarlos
hasta que se requiera para producir ATP en otras partes del cuerpo. Los
adipositos del tejido subcutáneo contienen cerca del 50% de la reserva de
triglicéridos. Otros tejidos adiposos explican la otra mitad (cerca del 21% en
torno a los riñones, 10 a 15% en los epiplones, 15% en áreas genitales, 5 a 8%
en los músculos y 5% de tras de los ojos, en los surcos del corazón y unidos al
exterior del intestino grueso). Los triglicéridos del tejido adiposo son
continuamente desdoblados y sintetizados. Por ello, los que almacena el tejido
adiposo un día no son las mismas moléculas del mes pasado, puesto que son
liberadas de dicha reserva continuamente, transportadas en la sangre y vueltas
a depositar en otras células del tejido graso.
Catabolismo
de lípidos: lipólisis
Los triglicéridos almacenados en el tejido adiposo
constituyen 98% de todas las reservas energéticas de todo el cuerpo. Se guardan
con más facilidad que el glucógeno, en parte debido a que son hidrófobos y no
ejercen presión osmótica sobre las membranas celulares. Varios tejidos
(músculo, hígado y tejido adiposo) oxidan de manera rutinaria ácidos grasos
derivados de triglicéridos para producir ATP; sin embargo, antes que los
triglicéridos puedan catabolizasre para producir ATP, deben desdoblarse en
glicerol y ácidos grasos, un proceso llamado lipólisis, que es
catalizado por las enzimas llamadas lipasas. Dos hormonas que
incrementan el desdoblamiento de triglicéridos en ácidos grasos y glicerol son
adrenalina y noradrenalina, que se liberan cuado aumenta el tono simpático,
como ocurre, por ejemplo, durante el ejercicio. Otras hormonas lipolíticas son
cortisol, hormona tiroidea y factores de crecimiento parecidos a insulina
(insulinoides). Por el contrario, esta hormona inhibe la lipólisis.
El glicerol y los ácidos grasos resultantes de la lipólisis
se catabolizan a través de diferentes vías (fig. 25.12). Muchas células del
cuerpo convierten el glicerol en gliceraldehído 3-fosfato, uno de los
compuestos que también se forman durante el catabolismo de la glucosa. Si el
suministro de ATP a una célula es alto, el gliceraldehído 3- fosfato se convierte
en glucosa, un ejemplo de gluconeogénesis. Sin embargo, cuando una célula
necesita producir más ATP, el gliceraldehído 3-fosfato entra en la secuencia
catabólica para producir ácido pirúvico.
{kind=link}
El catabolismo de los ácidos grasos es diferente al glicerol
y produce más ATP. El primer paso en el catabolismo de ácidos grasos comprende
una serie de reacciones denominadas oxidación beta, que tienen lugar en
la matriz de la mitocondria. A la vez, las enzimas retiran dos átomos de
carbono de la cadena larga de átomos de este elemento que componen un ácido
graso y unen el fragmento resultante de carbonos que unen a la coenzima A, para
formar acetil coenzima (CoA). En la segunda etapa del catabolismo de ácidos
grasos, el acetil CoA, formada como resultado de la oxidación beta, entra al
ciclo de Krebs (véase fig 25.12). Un ácido graso de 16 carbonos como el
ácido palmítico puede rendir una ganancia neta de 129 moléculas de ATP por su
oxidación completa a través de la oxidación beta, el ciclo de Krebs y la cadena
de transporte de electrones. Como parte del catabolismo normal de ácidos
grasos, y hepatocitos pueden captar dos moléculas de acetil CoA a la vez y
condensarla para formar ácido acetanóico. Esta reacción libera la porción
voluminosa de CoA que no puede rendir fuera de las células. Parte del ácido
acetanóico se convierte en ácido hidroxibutírico-beta y acetona.
La formación de estas tres sustancias, conocidas en conjunto como cuerpos
cetónicos, se llama cetogénesis (véase fig. 25.12). Puesto
que los cuerpos cetónicos difunden libremente a través de las membranas del
plasma, abandonan los hepatocitos y entran en la corriente sanguínea.
Otras células captan ácido acetanóico y unen sus cuatro
carbonos a dos moléculas de coenzima A para forman dos moléculas de CoA, que
después se incorporan al ciclo de Krebs para su oxidación. El músculo cardiaco
y la corteza renal (parte externa) usan ácido acetanóico en vez de glucosa para
genera ATP. Los hepatocitos, forman ácido acetoacéico, no pueden emplearlo para
producir ATP porque carecen de la enzima que transfiere el ácido acetoácetico
de regreso a la coenzima A.
Anabolismo
de lípidos: lipogénesis
Las células del hígado y las células adiposas pueden
sintetizar lípidos a través de glucosa o aminoácidos por medio de un proceso
llamado lipogénesis (véase fig. 25.12), el cual es estimulado por
la insulina. La lipogénesis tiene lugar cuando el individuo consume más
carbohidratos de los que necesita para satisfacer sus necesidades de ATP. El
exceso carbohidratos, proteínas y grasas en la dieta tiene un destino común: se
convierten en triglicéridos. Muchos aminoácidos pueden transformarse en acetil
CoA, que a su vez se convierte en triglicéridos. Los pasos en la conversión de
glucosa en lípidos implican la formación de gliceraldehído 3 fosfato, que puede
transformarse en glicerol y acetil CoA, que se convierte en ácidos grasos.
Éstos y el glicerol pueden ser sometidos a reacciones anabólicas que los
transforman en triglicéridos para su almacenamiento, o pasar por una serie de
reacciones anabólicas a fin de producir otros lípidos como lipoproteínas,
fosfolípidos y colesterol.
APLICACIÓN CLÍNICA
Cetosis
Normalmente el nivel de cuerpos cetónicos en la sangre es muy bajo, puesto que otros tejidos los usan para producir ATP con la misma rapidez con que se generan por el desdoblamiento de ácidos grasos en los hepatocitos. Sin embargo, durante periodos de oxidación beta excesiva, la producción de cuerpos cetónicos excede su captación y uso por las células del cuerpo. La oxidación beta excesiva puede ocurrir luego de una comida rica en triglicéridos, o durante ayuno en inanición, porque hay pocos carbohidratos disponibles para el catabolismo. Asimismo, la oxidación beta excesiva puede ocurrir en los pacientes con diabetes mellitus mal controlada o sin tratamiento por dos razones: 1) la glucosa adecuada no puede entrar a las células, por ende éstas usan los triglicéridos para producir ATP, y 2) la insulina inhibe normalmente la lipólisis y su carencia acelera el ritmo de este proceso. Cuando la concentración de cuerpos cetónicos en la sangre se eleva por arriba de lo normal, condición llamada cetosis, dichos cuerpos que, en su mayoría son ácidos deben ser amortiguados. Si se acumulan demasiados, reducen la concentración de amortiguadores, como los iones de bicarbonato, y el pH de la sangre desciende. En consecuencia, una cetosis extrema o prolongada puede conducir a la acidosis (cetoacidosis), un pH sanguíneo normalmente bajo. Cuando un diabético sufre deficiencia grave de insulina, uno de los signos característicos es el olor dulce en la respiración causado por el cuerpo cetónico o acetona.
METABOLISMO DE PROTEÍNAS
OBJETIVO
Describir el destino, el metabolismo y las funciones de
las proteínas.
Durante la
digestión, las proteínas se desdoblan en sus aminoácidos constituyentes, los
cuales en seguida son absorbidos por los capilares de las vellosidades del
intestino y transportados al hígado por la vena porta hepática. A diferencia de
los carbohidratos y triglicéridos, que se almacenan, las proteínas no se
guardan para su uso futuro; en vez de ello, los aminoácidos se oxidan para
producir ATP o se emplean para sintetizar nuevas proteínas para el crecimiento
y la reparación del cuerpo. El exceso de aminoácidos en la dieta no se excreta
en la orina o las heces, sino que se convierte en glucosa (gluconeogénesis) o
triglicéridos (lipogénesis).
Destino
de las proteínas
Factores de
crecimiento isulinoides (IGF) y la insulina estimulan el transporte activo de aminoácidos
al interior de las células del cuerpo. Casi inmediatamente después de su
ingreso, los aminoácidos se integran a las proteínas. Muchas de ellas funcionan
como enzimas; otras participan en el transporte (hemoglobina) o sirven como
anticuerpos, sustancias para la coagulación (fibrinógeno), hormonas (insulina)
o electos contráctiles en las fibras musculares (actina y miosina). Diversas
proteínas son componentes estructurales del cuerpo (colágeno, elastina y
queratina). Las distintas funciones de las proteínas se exponen en el cuadro
2.8.
Catabolismo
de proteína
Cada día
cierta porción del catabolismo de proteína tiene lugar en el cuerpo. Las
proteínas se extraen de células destruidas (como los eritrocitos) y se
fragmentan en aminoácidos libres. Algunos de ellos se convierten en
aminoácidos, vuelven a formar enlaces peptídicos y sintetizan nuevas proteínas
como parte de un continuo estado de recambio en todas las células. Una fracción
significativa de los aminoácidos absorbidos del CG, proviene de las proteínas
en células gastadas que se han desprendido de la mucosa intestinal hacia la
luz.
Las
proteínas recicladas primero se descomponen en aminoácidos. A continuación, los
hepatocitos los convierten en ácidos
grasos, cuerpos cetónicos o glucosa o bien los oxidan para transformarlos en
dióxido de carbono y agua. Sin embargo, antes que los aminoácidos puedan
catabolizarse primero deben convertirse en algunas sustancias que pueden
incorporarse al ciclo de Krebs.
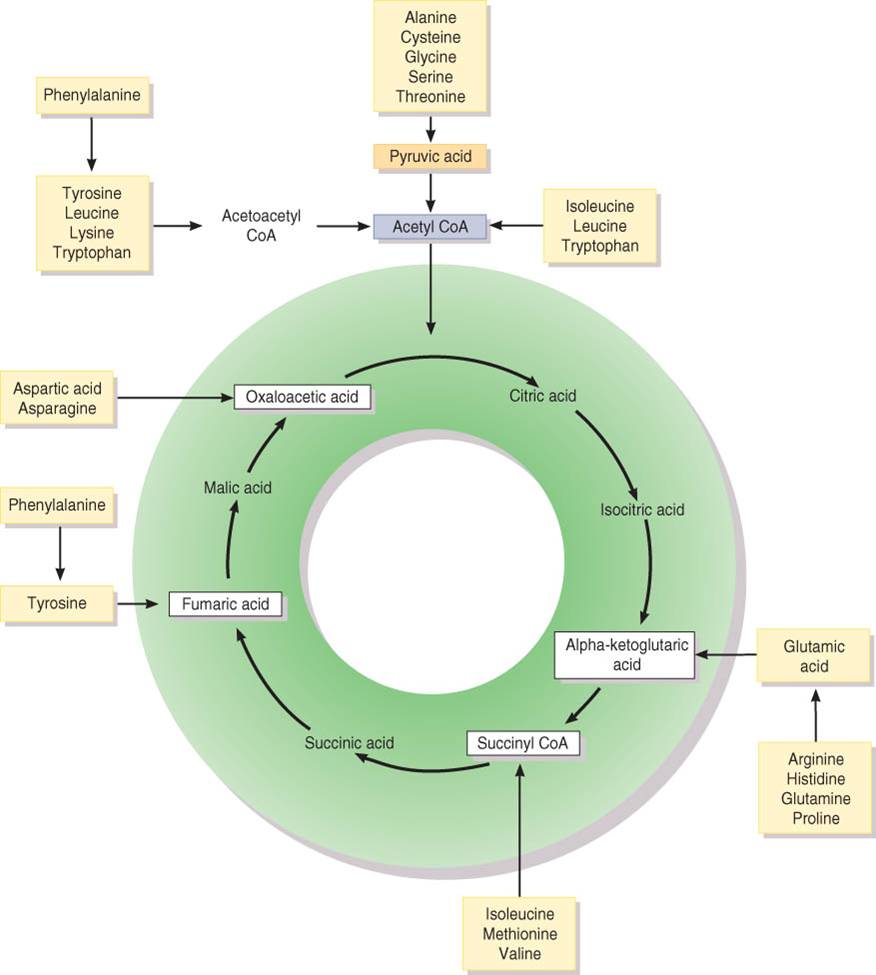
Una de estas conversiones consiste en retirar el grupo amino (NH2) del aminoácido, un proceso llamado desaminación, para transformarlo en amoniaco (NH3). Los hepatocitos convierten el amoniaco en urea, que se excreta en la orina. Otros procesos de cambio son la descarboxilación y la deshidrogenación. La figura 25.13 muestra cuales aminoácidos particulares participan en el ciclo de Krebs en diferentes puntos. La conversión de aminoácidos en glucosa (gluconeogénesis) puede analizarse en la figura 25.10; la transformación de aminoácidos en ácidos grasos (lipogénesis) o cuerpos cetónicos (cetogénesis) se muestra en la figura 25.12.
{kind=link}
Anabolismo
de proteína
El
anabolismo de proteína comprende la formación de enlaces peptídicos entre los
aminoácidos para producir nuevas proteínas. El anabolismo o síntesis de
proteína se produce en los ribosomas de casi todas las células del cuerpo y es
dirigido por el ADN y el ARN de las células (véase fig.
3.30). Los
factores de crecimiento insulinoides, las hormonas tiroideas T3 y T4), la
insulina, el estrógeno y la testosterona estimulan la síntesis de proteína.
Dado que son el principal componente de la mayor parte de las estructuras
celulares, es esencial que durante el crecimiento, el embarazo y cuando los
tejidos sufren daño por enfermedad o lesión, la dieta incluya cantidades
suficientes de proteínas. No obstante, cabe aclarar que su ingestión en exceso
no incrementa la masa ósea o la muscular; sólo un programa regular de
ejercitación con pesas puede lograr ese objetivo.
{kind=link}
De los 20 aminoácidos
que requiere el cuerpo humano, 10 son esenciales; dado que el cuerpo no
puede sintetizarlos, se deben obtener por medio de la dieta; de ellos, ocho no
pueden sintetizarse en absoluto (isoleucina, leucina, metionina, lecina.
fenilalanina, treonina, triptófano y valina) y los otros dos reproducen en
cantidades inadecuadas, principalmente en la niñez (arginina e histidina). En
virtud de que las plantas sí los sintetizan, los humanos pueden adquirir los
aminoácidos esenciales al ingerir plantas o carne de animales herbívoros. La
síntesis de aminoácidos no esenciales se lleva a cabo por un proceso
denominado transaminación, o sea, la transferencia de un grupo amino de
un aminoácido al ácido pirúvico o algún ácido del ciclo de Krebs. Una vez que
los aminoácidos apropiados, esenciales y no esenciales, están presentes en las
células, la síntesis de proteína se lleva a cabo con rapidez.
APLICACIÓN
CLÍNICA
Fenilcetonuria
La
fenilcetonuria o PKU es un error genético del metabolismo de proteína,
caracterizado por concentraciones elevadas en sangre del aminoácido
fenilalanina. La mayoría d los niños con este trastorno tienen una mutación del
gen que codifica la enzima hidroxilasa fenialalanina, la cual es necesaria para
convertir la fenilalanina en el aminoácido triocina, que participa en el ciclo
de Krebs. Puesto que hay deficiencia de la enzima, no se puede metabolizar
fenilalanina y lo que no se usa en la síntesis de proteína se acumula en la
sangre. Sin tratamiento, la enfermedad causa vómito, convulsiones, erupciones,
deficiencia de crecimiento y retraso mental grave. Se realizan pruebas a los
recién nacidos para determinar si padecen PKU, a que puede prevenirse el
retraso mental restringiendo las cantidades de fenilalanina necesarias para el
crecimiento al mínimo indispensable, aunque aún así el menor puede tener
deficiencia de aprendizaje. Edulcorante artificial aspartame (Nutra Sweet)
contiene fenilalanina y por ello debe restringirse su consumo en niños con este
trastorno.
MOLÉCULAS
BÁSICAS EN LAS ENCRUCIJADAS METABÓLICAS
OBJETIVO
Identificar las moléculas básicas en el metabolismo y
conocer las reacciones en los productos que pueden formar.
Aunque las
células tienen miles de sustancias químicas diferentes, sólo tres moléculas (glucosa
6-fosfato, ácido pirúvico y acetil coenzima A) desempeñan una función
fundamental en el metabolismo. Estas moléculas se encuentran en las
encrucijadas metabólicas; las reacciones que ocurren dependen del estado
nutricional o de las actividades del individuo. Las reacciones de 1 a 7 de la
figura 25.14 se desarrollan en el citosol, en tanto que las de 8 a 10 tienen
lugar dentro de la mitocondria.
{kind=link}
Papel
de la glucosa fosfato
Poco
después de que la glucosa entra en una célula del cuerpo, una cinasa la
convierte en glucosa 6-fosfato, que tiene cuatro destinos posibles
(véase fig. 25.14):
Síntesis de glucógeno. Cuando la glucosa es abundante
en el torrente sanguíneo, como ocurre después de ingerir alimentos, se emplea
una cantidad considerable de glucosa 6-fosfato para sintetizar glucógeno, que
es la forma de almacenamiento de los carbohidratos en los animales. El
desdoblamiento subsecuente del glucógeno en glucosa 6-fosfato tiene lugar
después de una serie de reacciones ligeramente diferentes. La síntesis y el
desdoblamiento de glucógeno se lleva a cabo principalmente en las fibras de
músculo esquelético y los hepatocitos.
Liberación de glucosa en la sangre. Si la enzima
glucosa 6-fosfatasa está presente y activa, la glucosa 6-fosfato puede
desfosfoilarse. Una vez que la glucosa es liberada del grupo fosfato, abandona
la célula y entra al torrente sanguíneo. Los hepatocitos son las principales
células que suministran glucosa a la circulación general.
Síntesis de ácidos nucleicos. La glucosa 6-fosfato es
el precursor que utilizan las células en el cuerpo para formar ribosa
5-fosfato, un azúcar de 5 carbonos necesario para la síntesis de ARN (ácido
ribonucleico) y ADN (ácido desoxirribonucleico). La misma secuencia de
reacciones que produce la ribosa 5-fosfato también genera NADPH, que se
requiere para ciertas reacciones de reducción.
Glucólisis. Parte del ATP se produce de manera
anaerobia por medio de glucólisis, en la cual la glucosa 6-fosfato se convierte
en ácido pirúvico, otra molécula básica que participa en el metabolismo. La
mayor parte de las células del cuerpo realizan glucólisis.
El
papel de ácido pirúvico
Cada
molécula de glucosa con 6 carbonos que sufre glucólisis produce dos moléculas de tres carbonos de ácido
pirúvico. Esta molécula se encuentra en una encrucijada metabólica: en
presencia de suficiente oxígeno, las reacciones aerobias (que consumen oxígeno)
de la respiración celular pueden llevarse a cabo; pero si dicho gas es escaso,
ocurren reacciones anaerobias (véase fig. 25-14).
Producción de ácido láctico. Cuando no hay suficiente
suministro de oxígeno a un tejido como en los músculos esquelético y cardiaco
durante su contracción activa, parte del ácido pirúvico se transforma en ácido
láctico, el cual pasa entonces a la circulación general y es captado por los
hepatocitos, los cuales lo convierten finalmente de nuevo en ácido pirúvico.
Producción de alanina. Al formarse el ácido pirúvico
se establece un vínculo entre el metabolismo de carbohidratos y de proteínas.
Un grupo amino (-NH3) puede añadirse al ácido pirúvico por
transaminación (un carbohidrato) para producir el aminoácido alanina, o bien
ser retirado de la alanina para generar ácido pirúvico.
Gluconeogénesis. El ácido pirúvico y ciertos
aminoácidos también pueden convertirse en ácido oxaloacético, uno de los
intermediarios del ciclo de Krebs, el cual, a su vez, puede utilizarse para
formar glucosa 6-fosfato. Esta secuencia de reacciones de gluconeogénesis pasan
por alto ciertas reacciones de una vía de glucólisis.
Función
de acetil coenzima A
Cuando el nivel de
ATP en una célula es bajo, pero el oxígeno es abundante, la mayor parte del
ácido pirúvico, se desvía hacia reacciones que producen ATP (ciclo de Krebs y
cadena de transporte de electrones) a través de la conversión de acetil
coenzima A.
Entrada al ciclo d Krebs. Acetil CoA es el vehículo
para que los grupos acetil de dos carbonos ingresen al ciclo de Krebs, cuyas
reacciones oxidativas convierten el acetil CoA en CO2 y producen
enzimas reducidas (NADH y FADH2) que transfieren electrones a la
cadena de transporte de electrones. A su vez, las reacciones oxidativas en esta
cadena generan ATP. La mayor parte de las moléculas que se oxidan para producir
ATP (glucosa, ácidos grasos y cuerpos cetónicos) primero se transforman en
acetil CoA.
Síntesis de lípidos. El acetil CoA también se emplea
en la síntesis de ciertos
lípidos, entre ellos ácidos grasos, cuerpos cetónicos y colesterol.
Puesto que el ácido pirúvico puede convertirse en acetil CoA, es posible transformar
carbohidratos en triglicéridos; esta es la vía metabólica para almacenar la
mayor parte del exceso de calorías en forma de grasa. Sin embargo, los
mamíferos, como los humanos no pueden reconvertir la acetil CoA en ácido
pirúvico por lo cual los ácidos grasos no de utilizan para generar glucosa u
otras moléculas de carbohidrato.
En el
cuadro 25.2 resume el metabolismo de carbohidratos, lípidos y proteínas.
ADAPTACIONES
METABÓLICAS
OBJETIVO
Comparar el metabolismo durante los estados absorbente y
postabsorbente.
La
regulación de las reacciones metabólicas depende del ambiente químico dentro de
las células del cuerpo, como las concentraciones de ATP y oxígeno, y de señales
de los sistemas nervioso y endocrino. Algunos aspectos del metabolismo dependen
del tiempo que ha transcurrido desde que se ingirió la última comida. Durante
el estado de absorción, los nutrimentos ingeridos entran en la circulación
sanguínea y la glucosa queda disponible para la producción de ATP. Durante el
estado de postabsorción, la asimilación de nutrientes en el CG se completa y
las necesidades energéticas deben satisfacerse con los combustibles del cuerpo.
De manera característica es necesario que pasen cuatro horas para que se lleve
acabo la absorción completa de los alimentos y dado que se ingieren tres
comidas cada 24 horas, el estado de absorción se prolonga 12 horas cada día. Si
se supone que no se comen refrigerios entre comidas, el estado de postabsorción
abarca las doce horas restantes (al final de la mañana, de la tarde y casi toda
la noche).
En virtud
de que el sistema nervioso y los eritrocitos dependen de la glucosa para la
producción de ATP durante el estado de postabsorción, es fundamental mantener
un estado estable de glucosa en sangre durante este periodo. Las hormonas son
los principales reguladores del metabolismo en ambos estados. Los efectos de la
insulina predominan en el estado de absorción, mientras que diversas hormonas
regulan el metabolismo del estado de postabsorción. Durante el ayuno y la
inanición, muchas células del cuerpo incrementan el uso de cuerpos cetónicos
para la producción de ATP.
Metabolismo
durante el estado de absorción
Poco
después de una comida, los nutrimentos comienzan a entrar en la sangre. Se debe
recordar que lo hacen principalmente en forma de glucosa, aminoácidos, y
triglicéridos (en quilomicrones).
Dos puntos de referencia metabólicos del estado de absorción
son la oxidación de la glucosa para producción de ATP, que ocurre en la mayor
parte de las células del cuerpo, y el almacenamiento de exceso de moléculas
combustibles par uso futuro entre comidas, lo que tiene lugar sobre todo en
hepatocitos, adipositos y fibras del músculo esquelético.
Reacciones
del estado de absorción
Las
siguientes reacciones predominan durante el estado de absorción (fig. 25.15):
{kind=link}
Alrededor de 50% de la glucosa absorbida de una comida
normal se oxida en las células de todo el cuerpo para producir ATP por vía de
la glucólisis, el ciclo de Krebs y la cadena de transporte de electrones.
La mayor parte de la glucosa que entra a los hepatocitos se
convierte en triglicéridos o glucógeno.
Parte de los ácidos grasos y los triglicéridos sintetizados
en el hígado permanecen allí, pero los hepatocitos los concentran
principalmente en VLDL, las cuales transportan los lípidos al tejido adiposo
para su almacenamiento.
Los adipositos también captan glucosa que no retiene el
hígado y la convierten en triglicéridos para el almacenamiento. En total, cerca
del 40% de la glucosa absorbida de los alimentos se transforma en triglicéridos
y alrededor del 10% se guarda como glucógeno en los músculos esqueléticos y los
hepatocitos.
La mayor parte de los lípidos de la dieta (principalmente
triglicéridos y ácidos grasos) se almacenan en tejido adiposo; sólo una pequeña
porción se emplea para reacciones de síntesis. Loas adipositos obtienen los
lípidos de los quilomicrones, de VLDL y de sus propias reacciones de síntesis.
Muchos aminoácidos absorbidos que entran a los hepatocitos
son desaminados para convertirse en cetoácidos, los cuales pueden incorporarse
al ciclo de Krebs para producir ATP o emplearse para sintetizar glucosa o
ácidos grasos.
Algunos de los aminoácidos que entran a los hepatocitos se
usan para sintetizar proteínas (proteínas del plasma).
Los aminoácidos no captados por los hepatocitos se usan en
otras células del cuerpo (como las musculares) para sintetizar proteínas o
sustancias químicas reguladoras como hormonas o enzimas.
Regulación del metabolismo durante el estado de absorción.
Poco
después de una comida, el péptido insulinotrópico dependiente de glucosa (GIP),
además de la creciente concentración de glucosa en sangre y ciertos
aminoácidos, estimula las células beta del páncreas para que liberen insulina.
En general, esta hormona incrementa la actividad de las enzimas necesarias para
el anabolismo y la síntesis de moléculas de reserva; al mismo tiempo disminuye
la actividad de las enzimas necesarias para reacciones catabólicas o de
desdoblamiento. La insulina favorece la entrada de glucosa y aminoácidos a las
células de muchos tejidos y estimula la fosforilación de glucosa en
hepatocitos, así como la conversión de glucosa 6-fosfato en glucógeno, tanto en
hígado como en células musculares. Una vez que llega al hígado y al tejido adiposo,
esta hormona aumenta la síntesis de triglicéridos y estimula la síntesis de proteínas en todas
las células del cuerpo (véase
capítulo 18).
La
concentración baja de glucosa en sangre también activa la rama simpática del
SNA. Las neuronas del hipotálamo sensibles a glucosa detectan su disminución e
incrementan los estímulos simpáticos. Como resultado, las terminaciones
nerviosas simpáticas liberan neurotransmisor noradrenalina y médula
suprarrenal, dos hormonas catecolaminas (adrenalina y noradrenalina) en la
sangre. Al igual que el glucagon, la adrenalina estimula el desdoblamiento de
glucógeno y en combinación con noradrenalina son potentes estimulado res de la lipólisis. Estas acciones de las
catecolaminas contribuyen a aumentar las concentraciones de glucosa y ácidos
grasos libres en el torrente sanguíneo. Como resultado, el músculo emplea más
ácidos grasos para producir ATP y hay disponibles mayores cantidades de glucosa
para el sistema nervioso. En el cuadro 25.4 se resume la regulación hormonal
del metabolismo durante el estado de postabsorción.
Metabolismo durante
ayuno e inanición
El término ayuno significa
no ingerir alimentos por muchas horas o algunos días, en tanto que inanición
implica semanas o meses de privación de alimentos o ingesta inadecuada de
nutrientes. Las personas pueden sobrevivir sin comer durante dos meses o más si
beben suficiente agua para evitar la deshidratación. Aunque las reservas de
glucógeno se agotan a las pocas horas de iniciar un ayuno, el catabolismo de
triglicéridos almacenados y de las proteínas estructurales pueden proporcionar
energía durante varias semanas. La cantidad de tejido adiposo que contiene el
cuerpo determina el tiempo que se puede vivir sin alimento.
Durante el ayuno y la
inanición, e! tejido nervioso y los eritrocitos continúan usando glucosa para
producir ATP. El suministro de aminoácidos para gluconeogénesis es rápido,
puesto que el descenso de insulina y el incremento de la concentración de
cortisol reduce la velocidad de síntesis de proteína y promueve su
catabolismo. La mayor parte de las células del cuerpo, en especial las del
músculo esquelético, pueden ahorrar una buena cantidad de proteína antes que su
función sea afectada de manera adversa debido a que contienen suficiente proteína
desde su origen. Durante los primeros días de ayuno, el catabolismo de proteína
supera su síntesis por casi
Hacia el segundo día de
ayuno, se estabilizan los niveles de glucosa en sangre en 65 mg/100 mL (3.6
mmol/litro), en tanto que la concentración de ácidos grasas en plasma se ha
cuadruplicado. La lipólisis de triglicéridos en tejido adiposo libera glicerol,
que se emplea para la gluconeogénesis, y ácidos grasoso Éstos difunden al
interior de las fibras musculares y otras células de! cuerpo, donde se usan
para elaborar acetil CoA que entra al ciclo de Krebs. A continuación, se
sintetiza ATP a medida que se realiza la oxidación por e! ciclo de Krebs y la
cadena de transporte de electrones.
El cambio metabólico más
notable que aparece con el ayuno y la inanición es el incremento en la
formación de cuerpos cetónicos por los hepatocitos. Durante el ayuno, sólo
pequeñas cantidades de glucosa sufren glucólisis para convertirse en ácido
pirúvico, que a su vez puede transformarse en ácido oxaloacético. El acetil
CoA entra al ciclo de Krebs al combinarse con el ácido oxaloacetico (véase
fig.
25.14); cuando éste es escaso por la falta de alimento, sólo una parte de acetil CoA disponible puede incorporarse al ciclo de Krebs. El exceso de acetil
CoA se emplea en la cetogénesis, sobre todo en los hepatocitos. De esta manera,
la producción de cuerpos cetónicos aumenta conforme se eleva el catabolismo de
ácidos grasas. Los cuerpos cetónicos liposolubles pueden difundir a través de
las membranas plasmáticas
y de la barrera
hematoencefálica para aprovecharse como .¡ combustible alterno en la producción
de ATP, especialmente por las fibras cardiacas musculoesqueléticas y las
neuronas. Normalmente, sólo en la sangre hay cantidades mínimas de cuerpos
cetónicos (0.01 mmol/litro) y por tanto Son una fuente insignificante de
combustible. Sin embargo, luego de dos días de ayuno, la concentración de
cetonas es
APLICACION CLÍNICA
Absorción de alcohol
Los efectos intoxicantes
e incapacitantes del alcohol dependen de su concentración en la sangre.
Dado que es liposoluble
se empieza a absorber en el estómago. Sin embargo, en el intestino delgado es
mayor el área de absorción que en el estómago; así, cuando el etilo pasa al
duodeno se capta más rápidamente. Cuanto más tiempo permanezca el alcohol en el
estómago, se eleva con mayor lentitud su nivel en la sangre. En virtud
de que los ácidos grasas en el quimo disminuyen la velocidad de
vaciamiento gástrico, la concentración de alcohol en la sangre se eleva con
mayor lentitud cuando se consumen alimentos ricos en grasa como pizza,
hamburguesas o nachos con las bebidas alcohólicas. Además, la enzima deshidrogenasa
de alcohol, presente en las células de la mucosa gástrica, desdobla una parte
del etilo en acetaldehído, que no es tóxico. Cuando la velocidad de vaciamiento
gástrico es más lenta, se absorbe proporcionalmente más alcohol y se convierte
en acetaldehído en el estómago, y por tanto menos cantidad llega a la
circulación general. Con la misma cantidad de alcohol, las mujeres suelen
tener niveles más altos de alcohol en la sangre (por tanto, experimentan mayor
intoxicación que los varones de la misma talla) por dos razones: 1) por
lo regular las mujeres poseen menor volumen de sangre y 2) la actividad
de la deshidrogenasa de alcohol en el estómago es hasta 60% menor en mujeres
que en varones. Los asiáticos también tienen menores niveles de esta
enzima gástrica .
CALOR Y BALANCE DE ENERGIA
OBJETIVOS
Definir el índice
metabólico basal (IMB) y conocer los diversos factores que lo afectan.
Describir los factores
que influyen en la producción del
calor corporal. '
Explicar como se
mantiene la temperatura normal del cuerpo mediante el circuito de
retroalimentación negativa que incluye el termostato hipotalámico.
El cuerpo produce más o
menos calor según el índice de las reacciones metabólicas. La homeostasis de la
temperatura corporal sólo puede mantenerse si el índice de pérdida de calor
del cuerpo es igual al índice de producción de calor metabólico. Por tanto, es
importante entender cómo se pierde, gana o conserva el calor.
Índice metabólico
La velocidad total a la
cual las reacciones metabólicas usan energía se denomina índice metabólico. Una
parte de la energía se emplea para producir ATP y otra se libera como calor,
que es una forma de energía cinética, la cual puede medirse como temperatura
y expresarse en unidades llamadas calorías. Una caloría, representada
con una c minúscula, es la cantidad de energía calórica que se requiere para
elevar la temperatura de
Dado que muchos factores
afectan el índice metabólico, se debe medir en condiciones estándares, con el
cuerpo en reposo y ayuno, lo que se denomina el estado basal. La medición
que se obtiene es el índice metabólico basal (1MB). La manera más común
de determinar el 1MB consiste en medir la cantidad de oxígeno que se consume
por kilocaloría de alimento metabolizada. Cuando el cuerpo usa
Homeostasis de la
temperatura corporal
No obstante las amplias
fluctuaciones en la temperatura ambiental, los mecanismos homeostásicos pueden
mantener Un intervalo normal de temperatura interna del cuerpo. Si el índice de
la producción del calor corporal es igual al índice de pérdida de calor, el
organismo mantiene una temperatura central constante cercana a
Producción de calor
La producción de calor
del cuerpo es proporcional al índice metabólico que se ve afectado por varios
factores, que también influyen de manera indirecta en el índice de producción
de calor:
Ejercicio.
Durante la actividad física extrema, el índice
metabólico puede aumentar hasta 15 veces la tasa basa!. En atletas de alto
rendimiento, dicho índice se incrementa hasta 20 veces.
Hormonas. Las hormonas tiroideas (tiroxina y
triyodotironina) son los principales reguladores del 1MB, el cual se incrementa
conforme se elevan los niveles de esta hormona. Sin embargo, la respuesta a
los cambios de concentración de hormonas tiroideas es lenta, tarda varios días
en aparecer. En parte incrementan el 1MB al estimular la respiración celular
aerobia. Puesto que las células usan más oxígeno para producir ATP, se
desprende más calor, y la temperatura del cuerpo se eleva. Otras hormonas tienen
menor efecto sobre el 1MB. La testosterona, la insulina y la hormona del
crecimiento humano pueden aumentar el índice metabólico de
Sistema nervioso.
Al hacer el ejercicio o en situaciones de estrés,
se estimula la división simpática del sistema nervioso autónomo. Sus neuronas
postganglionares liberan noradrenalina (NA), que a su vez estimula a la médula
suprarrenal que libera adrenalina y noradrenalina. Ambas incrementan el índice
metabólico de las células del cuerpo. Temperatura corporal. Cuanto más
alta sea la temperatura del cuerpo, mayor será el índice metabólico. Por cada
grado que aumenta la temperatura central se incrementa el índice de las
reacciones bioquímicas en casi 10%. Por tal razón, cuando una persona tiene
fiebre, se incrementa significativamente su índice metabólico.
Ingestión de
alimentos. Puede elevar el índice
metabólico en
Edad.
El índice metabólico de un niño, en relación con
su talla, es alrededor del doble de la de un anciano debido a la elevada
frecuencia de reacciones que se relacionan con ti crecimiento.
Factores diversos.
Otras variables que afectan el índice metabólico
son género (menor en mujeres, excepto durante el embarazo y la lactancia),
clima (más bajo en regiones tropicales), suei10 (menor) y malnutrición (más
baja).
Mecanismos de
transferencia de calor
El mantenimiento normal
de la temperatura corporal depende de la capacidad para eliminar el calor a la
misma velocidad con que se realizan las reacciones metabólicas. El calor se
transfiere del organismo al ambiente de cuatro maneras: por conducción,
convección, radiación y evaporación.
Conducción.
Es el intercambio de calor que tiene lugar entre
moléculas de dos materiales que están en contacto directo entre si. En reposo,
cerca de 3% del calor del organismo se pierde a través de la conducción a
materiales sólidos que están en contacto con el cuerpo, como una silla, la ropa
o joyas. Por conducción también es posible aumentar la temperatura, por
ejemplo, al sumergirse en una tina de agua caliente. Dado que este líquido
conduce el calor con eficacia 20 veces mayor que el aire, la pérdida o ganancia
de calor por conducción es mucho mayor cuando el cuerpo se halla inmerso en agua
fría o caliente.
Convección.
Es la transferencia de calor por el movimiento de
un fluido (gas o líquido) entre áreas de diferente temperatura. El contacto de
aire o agua con el cuerpo da como resultado la transferencia del calor por
conducción y convección. Cuando el aire entra en contacto con el cuerpo, se
calienta y por tanto se hace menos denso y se eleva. Cuanto más rápido se mueva
el aire (por ejemplo, por una brisa o un abanico), mayor será el índice de convección.
En reposo, se pierde cerca de 15% del calor corporal por convección y
conducción.
Radiación. Es la transferencia de calor en forma de rayos
infrarrojos entre el objeto más caliente y el más frío sin que haya contacto
físico. El cuerpo pierde calor al irradiar más ondas infrarrojas de las que
absorbe de objetos más fríos. Si éstos tienen mayor temperatura que el cuerpo,
la persona adquiere más calor del que pierde por radiación. En una habitación a
Evaporación.
Es la conversión de un líquido en vapor.
Cada mililitro de agua
que se evapora consume una gran cantidad de calor (cerca de 0.58 Cal/mL). En
condiciones normales de reposo, cerca de 22% de la pérdida de calor ocurre por
la evaporación de unos 700 mL de agua por día (300 mL en el aire espirado y 400
mL en la superficie de la piel). Dado que normalmente no nos percatamos de esta
pérdida de agua a través de la piel y las membranas mucosas de la boca y vías
respiratorias, este proceso se llama pérdida insensible de agua. El
índice de evaporación tiene una relación inversa con la humedad relativa, es decir,
la razón de la verdadera cantidad de humedad que el aire puede mantener a una
temperatura determinada. Cuanto más alta resulte la humedad relativa, menor
será el índice de evaporación. Cuando el ambiente tiene 100% de humedad, la
ganancia de calar por condensación de agua en la superficie cutánea es tan
rápida como su pérdida por evaporación, la cual constituye la principal defensa
contra el sobrecalentamiento al practicar un deporte. En condiciones extremas,
se puede producir Un máximo de casi
Termostato
hipotalámico
El centro de control que
funciona como termostato del cuerpo es un grupo de neuronas en la parte al
interior del hipotálamo, la región preóptica. Esta área recibe impulsos
de termorreceptores situados en la piel y membranas mucosas, así como el
hipotálamo. Las neuronas de la región preóptica generan impulsos nerviosos a
una mayor frecuencia cuando aumenta la temperatura de la sangre, y cuando ésta
disminuye, se reduce dicha frecuencia.
Los impulsos nerviosos de
la región preóptica se propagan a otras dos partes del hipo tálamo conocidas
como centro de pérdida de calor y centro promotor de calor, que, cuando
son estimulados por la región preóptica, ponen en marcha una serie de
respuestas que reducen y elevan la temperatura corporal, según se requiera.
Termorregulación
Si la temperatura central
del cuerpo desciende, actúan mecanismos que ayudan a conservar o incrementan la
producción de calor por medio de varios circuitos de retroalimentación negativa
que normalizan la temperatura del cuerpo (fig. 25.17). Los termorreceptores en
la piel y el hipotálamo envían impulsos nerviosos a la región preóptica y al
centro promotor de calor en el hipotálamo, así como a las células
neurosecretoras que producen hormona liberadora de tirotropina o HLT, que por
su parte estimula los tirotrofos en la hipófisis anterior para liberar hormona
estimulante de la tiroides (HET). Los impulsos nerviosos del hipotálamo y la
HET activan entonces varios efecto res.
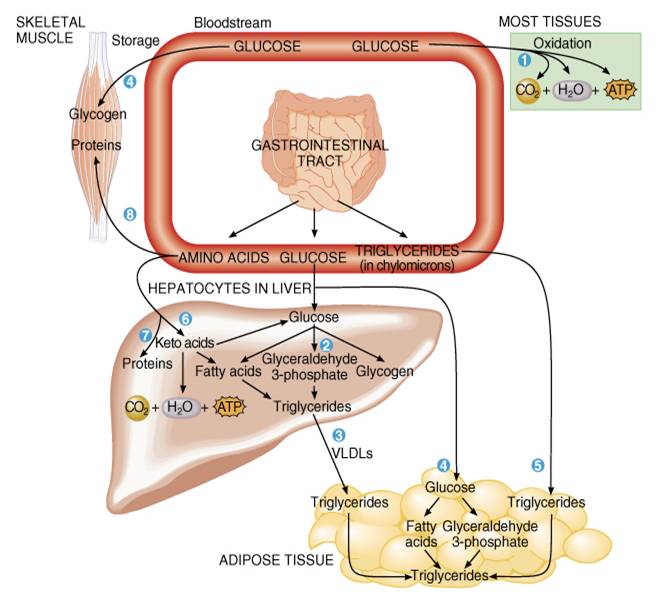{kind=link}
En respuesta, cada uno de
ellos ayuda a incrementar la temperatura central en niveles normales:
Los impulsos nerviosos
del centro promotor de calor estimulan nervios simpáticos que causan
constricción de los vasos sanguíneos de la piel, con lo cual disminuye el flujo
de sangre caliente y por tanto se efectúa la transferencia de calor de los
órganos internos a la piel. La disminución del índice de pérdida de calor
permite que se incremente la temperatura interna corporal mientras las
reacciones metabólicas producen calor.. Impulsos nerviosos en los nervios
simpáticos que llegan a la médula suprarrenal estimulan la liberación de
adrenalina y noradrenalina en la sangre. Las hormonas, a su vez, aumentan el
metabolismo celular, que a su vez incrementa la producción de calor. . i El
centro promotor de calor estimula partes del encéfalo que incrementan el tono
muscular y por tanto la producción de calor. Conforme se eleva el tono en un
músculo (ej. agonista), pequeñas contracciones estiran los huesos musculares en
su antagonista, para iniciar un reflejo de estiramiento. La contracción
resultante en el antagonista estira los husos musculares en el agonista, y esto
también da lugar a un reflejo de estiramiento. Este ciclo repetitivo
llamado temblor, aumenta en gran medida el índice de producción de calor.
Durante el temblor máximo, la temperatura corporal puede elevar hasta cuatro
veces el índice basal en pocos minutos.
La glándula tiroides
responde a la estimulación de la HET, liberando más hormonas tiroideas en la sangre.
Conforme se eleva su nivel, también aumenta lentamente el Índice metabólico y
se incrementa la temperatura corporal.
Si la temperatura del
cuerpo sube más de lo normal, un circuito de retroalimentación negativa opuesto
al que se muestra en la figura 25.17 entra en acción. La temperatura más
elevada de la sangre estimula termorreceptores que envían impulsos nerviosos a
Ia región preóptica, lo que por su parte estimula el centro de pérdida de calor
e inhibe la acción del centro promotor de calor. Los impulsos nerviosos del
centro de pérdida de calor causan dilatación de los vasos sanguíneos de la
piel, la cual se torna caliente y pierde el exceso de calor por radiación y
conducción hacia el ambiente y al aumentar el volumen de sangre que fluye desde
el centro del cuerpo, que es más caliente, hacia la piel, cuya temperatura es
menor. Al mismo tiempo, disminuye el índice metabólico y no ocurre el temblor.
La temperatura elevada de la sangre estimula las glándulas sudoríparas de la
piel por medio de activación hipotalámica de nervios simpáticos. Conforme el
agua de la transpiración se evapora en la superficie cutánea, ésta se enfría.
Todas estas respuestas contrarrestan los efectos promotores de calor y ayudan a
que el cuerpo recupere su temperatura normal.
APLICACIÓN CLÍNICA
Hipotermia
La hipotermia es
la disminución de la temperatura central
del cuerpo a
Regulación de la
ingesta de alimentos
Cuando el contenido
energético de los alimentos es igual a la energía empleada por las células del cuerpo,
se mantiene constante el peso corporal. Esto es verdad salvo si existe
ganancia o pérdida de agua. Sin embargo, en las naciones más ricas, una
fracción significativa de la población muestra sobrepeso, un problema que
aumenta el riesgo de muerte por varias enfermedades cardiovasculares y metabólicas,
por ejemplo, diabetes mellitus.
La mayoría de los
animales, hombres y mujeres adultos mantienen un peso corporal estimu1ule
durante periodos muy prolongados. La estabilidad ponderal puede persistir a pesar
de grandes variaciones
cotidianas en la actividad y la ingestión de alimentos. Pero no existen
receptores sensoriales que vigilen el peso corporal ni la talla. ¿Cómo se
regula entonces la ingestión de alimentos? Resulta difícil dar una respuesta inequívoca
a esta pregunta. Según parece depende de muchos factores, como los niveles de
ciertos nutrientes en la sangre, ciertas hormonas, elementos psicológicos Como
estrés o depresión, estímulos provenientes del conducto gastrointestinal y de
los sentidos especiales, así como las conexiones neuronales entre el hipotálamo
y otras partes del encéfalo.
En el hipotálamo se
encuentran núcleos que desempeñan papeles clave en la regulación de la
ingestión de alimentos (véase fig. 14.10,
). Cuando se estimula un grupo de neuronas en animales el centro de alimentación, que
se localiza en los núcleos hipotalámicos laterales-, éstos comienzan a comer
vorazmente, aunque estén totalmente llenos. Un segundo centro, un conjunto de
neuronas denominado el centro de saciedad, se encuentra en los núcleos
ventromediales del hipotálamo. Saciedad es lo opuesto al apetito; consiste en
una sensación de plenitud acompañada por una falta de deseo de comer. La estimulación
de este centro ocasiona que los animales dejen de ingerir alimentos, aun si
han ayunado por varios días.
{kind=link}
¿Cómo pueden las neuronas
del hipotálamo percibir si una persona está llena o necesita alimento? Una
posibilidad es que los cambios en la composición química de la sangre después
de comer y durante el ayuno alerten a las neuronas hipotalámicas.
Las moléculas
estimuladoras en la sangre que actúan sobre el hipotálamo para disminuir el
apetito e incrementar el consumo de energía incluyen varias hormonas (glucagon,
colecistocinina y adrenalina que actúan a través de los receptores beta),
glucosa y leptina, la cual es liberada por adipocitos conforme sintetizan
triglicéridos. Las moléculas estimulado ras que incrementan el apetito y
disminuyen el consumo de energía son las siguientes: opioides, hormona
liberadora de la hormona del crecimiento (HLHC), glucocorticoides, adrenalina
que actúa por medio de los receptores alfa, insulina, progesterona y
somatostatina.
La ingestión de alimento
también es regulada por la distensión del CG, en particular estómago y
duodeno. El estiramiento de estos órganos inicia un reflejo que activa el
centro de la saciedad e inhibe el de la alimentación. Factores psicológicos
pueden contrarrestar los mecanismos habituales de ingestión, como ocurre, por ejemplo,
en la anorexia nerviosa, bulimia y obesidad (véase más adelante en este
capítulo).
NUTRICION
OBJETIVO
Describir de qué
numera se pueden seleccionar los alimentos para
mantener una dicta saludable.
Comparar las fuentes,
las funciones y la importancia de minerales y vitaminas en el metabolismo.
i
Los alimentos contienen
las sustancias, llamadas nutrimentos, que el cuerpo usa para su
crecimiento, mantenimiento y reparación de tejidos. Los nutrimentos incluyen
agua, carbohidratos, lípidos, proteínas, vitaminas y minerales. El nutriente
que se requiere en mayor cantidad es el agua: alrededor de
A continuación, se
analizan algunos lineamientos para una dieta saludable y se exponen las
funciones de minerales y vitaminas en el metabolismo.
Lineamientos para una
dieta saludable
Considerando la
alimentación, cada día muchas mujeres y los ancianos requieren cerca de 1 600
calorías; los niños, las adolescentes, las mujeres activas y la mayoría de los
varones requieren alrededor de 2 200 calorías; los adolescentes y varones
activos precisan cerca de 2 800 calorías. Cada gramo de proteína o carbohidrato
en los alimentos suministra casi 4 calorías, en tanto que un gramo de grasa
([lípidos) proporciona cerca de 9 calorías.
Aún no se sabe con
certeza qué concentración y tipo de carbohidratos, grasa y proteínas son
óptimos en la dieta; diferentes poblaciones en todo el mundo consumen alimentos
radicalmente diferentes adaptados a su particular estilo de vida. Los expertos
recomiendan la siguiente distribución de calorías:
Los lineamientos para una
dieta saludable son:
Comer alimentos variados.
Mantener un peso corporal
adecuado.
Elegir alimentos bajos en
grasa, grasa saturada y coles te rol. Conmer abundantes vegetales, frutas y
gramíneas. Consumir azúcares con moderación.
Consumir sal)' sodio con
moderación (menos de 6 gramos diarios).
Si se ingieren bebidas
alcohólicas, hacerla con moderación (menos de
Para ayudar a la gente a
lograr un buen balance de vitaminas, minerales, carbohidratos, grasas)'
proteínas en su alimentación, el U.S. Departmento of Agriculture desarrolló la
guía de la pirámide alimentaria (fig. 25.18). Las secciones de dicha pirámide
indican cuántas razones de cada uno de los cinco principales grupos de
alimentos deben comerse cada día. El menor número de raciones corresponde a una
dieta de 1 600 Cal/día en tanto que el mayor número de raciones corresponde a
una dieta de 2 800 Cal/día. Puesto que deben consumirse en la mayor cantidad,
los alimentos ricos en carbohidratos complejos (el grupo del pan, cereales,
arroz y pastas) constituyen la base de la pirámide. Los vegetales y las frutas
forman el siguiente nivel. Los beneficios para la salud por comer abundante
cantidad de estos alimentos están bien comprobados. Los alimentos del siguiente
nivel son los que se comen en cantidades menores (el grupo de leche, yogur y
queso y la carne, aves, pescados, granos secos, huevos y nueces). Estos dos
grupos de alimentos tienen contenido más alto de grasa y proteínas que los de
alimentos situados por debajo de ellos. Para reducir la ingestión diaria de
grasas, deben elegirse los alimentos que la contienen en poca cantidad (leche y
yogur descremados, queso bajo en grasa, pescado y aves [sin piel]).
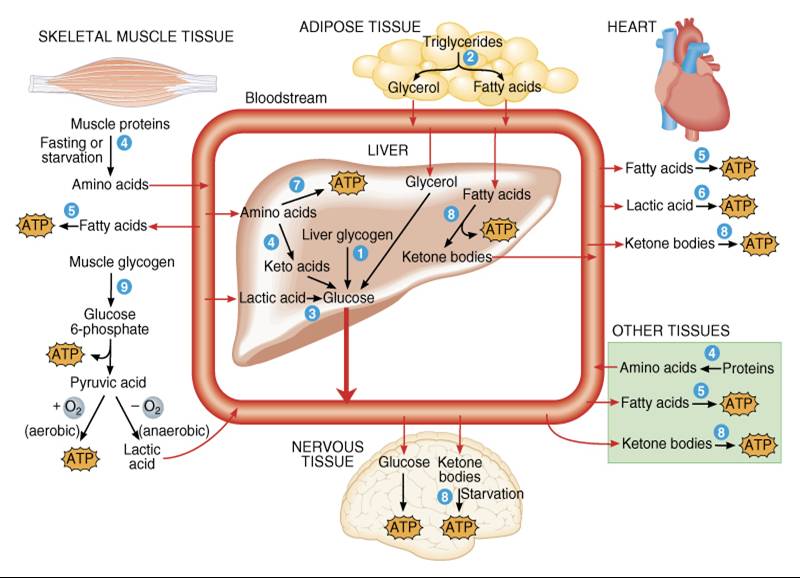{kind=link}
El vértice de la pirámide
no es un grupo de alimentos sino que advierte que se debe comer con moderación
grasas, aceites y azúcares. Aunque la pirámide de alimentos no distingue entre
diferentes tipos de ácidos grasas (saturados, poliinsaturados y
monoinsaturados) en las grasas de la dieta, debe advertirse que la
ateroesclerosis y enfermedad de arteria coronaria prevalecen en poblaciones que
consumen grandes cantidades de grasas saturadas y colesterol.
Las poblaciones que viven
a orillas del Mar Mediterráneo, por el contrario, presentan porcentajes bajos
de enfermedad de arteria coronaria a pesar de que su dieta contiene hasta 40%
de las calorías como grasas. Sin embargo, la mayor parte de sus grasos proviene
del aceite de oliva que es rico en ácidos grasas no saturados) no tiene
colesterol. Aceite de canola, aguacates, nueces y de cacahuate también son
ricos en ácidos grasas no saturados.
Minerales
Algunos minerales y
muchas vitaminas son componentes de los sistemas enzimáticos que catalizan
reacciones metabólicas. Los minerales constituyen elementos inorgánicos
naturales que se encuentran en la corteza terrestre. El cuerpo los contiene
combinados entre sí, con compuestos orgánicos o como iones en solución. Los
minerales constituyen cerca de 4% del peso total del cuerpo y se concentran
principalmente en el esqueleto. Los que el organismo contiene en mayor
cantidad son calcio, fósforo, potasio, azufre, sodio, cloro, magnesia,
hierro y yodo (véase cuadro 2.1, capítulo 2). Otros minerales que son esenciales para la vida comprenden
manganeso, cobre, cobalto, cinc, flúor, selenio y cromo. Los minerales
aluminio, sílice, arsénico y níquel se encuentran en el cuerpo, pero se
desconoce su función, si es que tienen alguna.
El calcio y el fósforo forman parte de la matriz ósea.
Puesto que los minerales no forman compuestos de cadena larga, no son óptimos
materiales estructurales Su función principal consiste en regular algunas
reacciones neumáticas. El calcio, hierro, magnesia y manganeso son
elementos constituyentes de algunas coenzimas. El magnesio también sirve como catalizador para la conversión de ADP
en ATP. Minerales como so dio y fósforo trabajan en sistemas amortiguadores. El sodio también ayuda a regular la
ósmosis de agua y, junto con otros iones, participa en la generación de
impulsos nerviosos. El cuadro 25.5 describe' las funciones vitales de minerales
seleccionados. Nótese que el cuerpo generalmente usa los iones de los minerales
en vez de la forma no ionizada. Algunos minerales, como el cloro, son tóxicos o
inclusa mortales cuando se ingieren en la forma no ionizada.
Vitaminas
Los nutrimentos orgánicos requeridos en cantidades pequeñas para mantener el crecimiento y metabolismo
normales se llaman vitaminas. A diferencia de los carbohidratos,
lípidos o proteínas, no suministran energía ni sirven como materiales
estructurales del organismo. De las vitaminas cuyas funciones se conocen, la
mayoría sirve como coenzimas.
El cuerpo no puede
sintetizar un gran número de vitaminas, por lo que deben ingerirse por medio
de la dieta. Otras vitaminas, como la K, son producidas por bacterias en el CG
y luego se absorben. El cuerpo puede ensamblar algunas vitaminas si se le
suministran los materiales crudos, llamados provitaminas. Por ejemplo, el
cuerpo produce vitamina A partir de la
provitamina carotenobeta, una sustancia química presente en vegetales
amarillos como zanahorias y de color verde oscuro como espinacas. No existe un
alimento que contenga todas las vitaminas requeridas, lo cual constituye una
de las razones para tener una dieta variada.
Las vitaminas se dividen
en dos grupos principales: las lipa solubles y las hidrosolubles. Las vitaminas
liposolubles se absorben junto con otros lípidos de la dieta en el intestino
delgado y se concentran en quilomicrones. No pueden absorberse en cantidades
adecuadas a menos que se ingieran con otros lípidos. Las vitaminas liposolubles
pueden almacenarse en células, en particular hepatocitos. Entre los ejemplos
de ellas están las vitaminas A, D, E y K. En contraste, el CG absorbe las
vitaminas hidrosolubles, junto con el agua en el conducto gastrointestinal y se
disuelven en los líquidos corporales. Cantidades excesivas de estas vitaminas
no se almacenan, sino por el contrario, se excretan en la orina. Entre los
ejemplos se pueden citar las vitaminas B y C.
Además de sus otras
funciones, tres vitaminas (C, E y caroteno-beta, una provitamina) se
denominan vitaminas antioxidantes porque inactivan radicales libres de oxígeno,
los cuales son partículas sumamente reactivas que transportan un electrón impar. Los radicales libres dañan las membranas
celulares, el ADN Y otras estructuras celulares y contribuyen a la formación
de placas arteroescleróticas que estrechan las arterias. Algunos radicales
libres se originan de manera natural en el cuerpo y otros se derivan de
factores adversos ambientales como humo de tabaco y radiación. Se cree que las
vitaminas antioxidantes desempeñan una función en la protección contra algunos
tipos de cáncer, reducción de la formación de placas arteroescleróticas,
retraso de algunos efectos del envejecimiento y reducción de la probabilidad de
la formación de cataratas en el cristalino. El cuadro 25.6 muestra una lista de
las principales vitaminas, su origen, funciones y enfermedades relacionadas con
su deficiencia.
APLICACIÓN CLÍNICA
Suplementos de
vitaminas y minerales
La mayoría de los
nutricionistas recomiendan consumir una dieta balanceada que incluya una
variedad de alimentos en vez de ingerir suplementos vitamínicos o de minerales,
excepto en circunstancias especiales. Ejemplos comunes de suplementan
justificada son: hierro para mujeres con sangrado menstrual excesivo; hierro y
calcio para embarazadas o mujeres que amamantan; ácido fálico (fosfato) en el
caso de todas las embarazadas, para reducir el riesgo de defectos del tubo
neural en el feto; calcio para la mayoría de los adultos, puesto que no reciben
la cantidad recomendada en su dieta; y vitamina B1Z para
vegetarianos estrictos. Puesto que la mayoría de los estadounidenses no
ingieren en sus alimentos las elevadas concentraciones de vitaminas
antioxidantes que se cree tienen efectos benéficos, algunos especialistas
recomiendan vitaminas e y E suplementarias. Sin embargo, no se debe exceder el
consumo de éstas, ya que las megadosis de vitaminas o minerales resultan muy
dañinas.
ANORMALIDADES DE LA
TEMPERATURA CORPORAL
Fiebre
La fiebre es una
elevación de la temperatura central del cuerpo como resultado de un reajuste
del termostato hipotalámico. La causa más común de fiebre es una infección
viral o bacteriana (o por toxinas de bacterias); otras causas son ovulación,
secreción excesiva de hormonas tiroideas, tumores y reacciones a vacunas. Una
sustancia que produce fiebre se denomina pirógeno y la induce de la
siguiente manera: cuando los fagocitos ingieren ciertas bacterias, son estimulados
para secretar interleucina, que actúa como pirógeno. La circulación la lleva
al hipo tálamo donde hace que las neuronas de la región preóptica secreten
prostaglandinas, en particular de la serie E. Las prostaglandinas reajustan el
termostato hipotalámico a una temperatura más alta y los mecanismos reflejos
reguladores de la temperatura actúan entonces para aumentar la temperatura
central del cuerpo. Los antipirético, son agentes que alivian o
reducen la fiebre; entre ellos están: aspirina, acetaminofeno (Tylenol) e
ibuprofeno (Advil), que reducen la fiebre al inhibir la síntesis de
prostaglandinas.
Supongamos que como
resultado de la acción de pirógenos el termostato se reajusta a
La fiebre es benéfica
hasta cierto punto. Una elevación de la temperatura intensifica el efecto del
interferón y de la actividad fagocitaria de los macrófagos en tanto que
dificulta la reproducción de algunos patógenos. Puesto que la fiebre incrementa
la frecuencia cardiaca, los leucocito s que combaten la infección llegan a los
sitios infectados con mayor rapidez. Además, se incrementa la proliferación de
anticuerpos y células T; más aún, el calor acelera la velocidad de las
reacciones químicas, lo que contribuye a que las células del cuerpo se reparen
con mayor prontitud durante una enfermedad. Las complicaciones de la fiebre son
deshidratación, acidosis y daño cerebral permanente. Como regla, si la
temperatura central se eleva arriba de
Calambres por
calor, agotamiento por calor y golpe de calor
Los calambres por
calor se presentan como resultado de sudación profusa que elimina agua y
sal (NaCl) del cuerpo. La pérdida de sal causa contracciones dolorosas de los
músculos, que tienden a ocurrir en los tejidos musculares que se usan durante
el trabajo, pero que no se manifiestan hasta que la persona se relaja una vez
que ha concluido su labor. Los líquidos salados en general producen rápida
mejoría.
En e! agotamiento por
calor (postración por calor), la temperatura central suele ser normal, o
un poco menor, y la piel se muestra fría y húmeda a causa de transpiración
profusa. El agotamiento por calor normalmente se caracteriza por pérdida de
líquidos y electrólitos, en especial sales. La pérdida de sal provoca calambres
musculares, vértigo, vómito o desmayo; la de líquido causa descenso de la
presión arteria!. Se recomienda reposo completo y la ingestión de tabletas de
sal.
Cuando ocurre un golpe
de calor la temperatura y humedad relativa son altas, lo que hace difícil
que el cuerpo pierda calor por radiación, conducción o evaporación, y como
resultado hay deficiencia neurológica. El riego sanguíneo a la piel disminuye,
la transpiración se reduce mucho y la temperatura central se eleva bruscamente.
Por tanto, la piel se muestra seca y caliente, puede alcanzar una temperatura
hasta de 43 0c. Puesto que las células del cerebro se ven afectadas, el
termostato hipotalámico deja de funcionar. El tratamiento, que debe iniciarse
de inmediato, consiste en enfriamiento del cuerpo por inmersión de la víctima
en agua fría y administración de líquidos y electrólitos.
OBESIDAD
Obesidad,
peso corporal más de 20% por arriba de algún estándar
deseable a causa de una acumulación excesiva de tejido adiposo (afecta a una
tercera parte de la población adulta estadounidense). Un atleta puede tener
sobrepeso debido a cantidades mayores de lo normal de tejido muscular sin ser
obeso. Incluso la obesidad moderada es peligrosa para la salud; es un factor
de riesgo de enfermedad cardiovascular, hipertensión, enfermedad pulmonar,
diabetes mellitus no dependiente de insulina, artritis, ciertos tipos de
cánceres (mama, útero y colon), venas varicosas y enfermedad de la vesícula
biliar. Además, se ha demostrado que la pérdida de grasa corporal en individuos
obesos eleva el colesterol HDL, e! tipo relacionado con prevención de enfermedad
cardiovascular.
En pocos casos, la
obesidad puede ser resultado de traumatismo o de tumores en el centro
regulador de la alimentación en e! hipotálamo. En casi todos los casos, no se
puede identificar una causa. Las variables que contribuyen a este trastorno
incluyen factores genéticos, hábitos alimentarios aprendidos en la infancia,
sobrealimentación para aliviar tensiones y costumbres sociales. Los estudios
indican que algunas personas obesas queman menor número de calorías durante la
digestión y la absorción de una comida. Además, los obesos que reducen su peso
necesitan consumir 15% menos de calorías para mantener un peso normal en
comparación con las personas que nunca han engordado. Aunque la leptina suprime
el apetito y produce saciedad en experimentos con animales, no se encuentra
disminuida en la mayoría de las personas obesas.
MALNUTRICIÓN
Malnutrición,
desequilibrio de la ingestión de nutrimentos (ya
sea ingestión calórica total insuficiente o excesiva o deficiencia de
nutrimentos específicos). Una causa de mal nutrición (subnutrición, o ingestión
inadecuada de alimento) puede ser resultado de condiciones como ayuno, anorexia
nerviosa, privación de alimento, cáncer, obstrucción gastrointestinal,
incapacidad para la deglución, enfermedad renal y dentadura en malas
condiciones. Otras causas de mal nutrición son el desequilibrio y la
malabsorción de nutrimentos, su distribución o incapacidad para aprovechados
(por ejemplo, en la diabetes mellitus), mayor re.9uerimiento de nutrimentos
(debido a fiebre, infecciones, quemaduras, fracturas, estrés y exposición a
calor o frío), pérdida excesiva de nutrimentos (diarrea, hemorragia o
glucosuria) y sobrenutrición (exceso de vitaminas, minerales y calorías).
Uno de los principales
tipos de subnutrición es resultado de la ingestión inadecuada de proteína,
calorías o ambas necesarias para satisfacer los requerimientos nutritivos de
una persona. Esta subnutrición de proteínas y calorías puede clasificarse en
dos tipos con base en las carencias de la dieta. En un tipo, llamado cuasiorcor,
la ingestión de proteínas es deficiente a pesar de una ingestión calórica
normal o casi normal. Debido a que la principal proteína en el maíz, la zeína,
carece de los aminoácidos esenciales triptófano y lisina, necesarios para el
crecimiento y la reparación de tejidos, muchos niños africanos cuya dieta
consta principalmente de harina de maíz desarrollan cuasiorcor. Los principales
signos de cuasiorcor son edema del abdomen, hígado hipertrofiado, hipotensión
arterial, bradicardia, hipotermia, anorexia, letargo, piel seca e
hiperpigmentada, caída del cabello y a veces retraso mental.
Otro tipo de subnutrición
de proteínas calorías, llamado marasmo, se debe a ingestión inadecuada
de proteínas y calorías. Sus características incluyen retraso del crecimiento,
peso bajo, desgaste muscular, emaciación, piel seca y cabello mate, delgado,
seco.
UNIVERSIDAD AUTÓNOMA SE
ZACATECAS
CIENCIAS MORFOLÓGICAS
Dr.: SIXTO SOSA DÍAZ
ALUMNA: MARÍA ESPERANZA GALLEGOS RANGEL