APARATO DIGESTIVO
Los alimentos contienen diversos nutrientes, los cuales son moléculas necesarias para que se formen nuevos tejidos, se reparen los dañados y tengan lugar las reacciones químicas imprescindibles. Además, son esenciales para la vida, ya que constituyen la fuente de energía para las reacciones químicas que ocurren en todas las células. Sin embargo, tal y como se ingieren, muchos alimentos no pueden utilizarse como fuente de energía celular. En primer término, se requiere su desdoblamiento en moléculas suficientemente pequeñas para cruzar la membrana plasmática de las células, proceso llamado digestión. El paso de estas moléculas más pequeñas por ciertas células a la linfa y sangre se denomina absorción. Los órganos que realizan conjuntamente estas funciones, es decir, el aparato digestivo, son el tema de este capítulo.
La rama de la medicina que estudia la estructura y el funcionamiento del estómago e intestinos, así como el diagnóstico y tratamiento de sus enfermedades, se llama gastroenterología. La disciplina médica relacionada con el diagnóstico y tratamiento de enfermedades del recto y ano es la proctología.
GENERALIDADES DEL APARATO DIGESTIVO
El aparato digestivo (fig. 24.1) se compone de dos grupos de órganos, el tubo digestivo y los órganos accesorios. El primero es un tubo continuo que se extiende de la en la cavidad corporal ventral. Sus órganos comprenden boca, gran parte de la faringe, esófago, estómago intestino delgado e intestino grueso. Su longitud, en un cadáver es de 9 m, mientras que en personas vivas es más corto, ya que el músculo liso de la pared del tubo digestivo guarda un estado tónico (de contracción sostenida). Los órganos abarcan dientes, lengua, glándulas salivales, hígado, vesícula biliar y páncreas. Los dientes participan en la descomposición física de los alimentos, y la lengua, en la masticación, deglución. En contraste, los demás órganos auxiliares de la digestión nunca tienen contacto directo con los alimentos. En vez de ello, producen o almacenan secreciones al tubo digestivo por diversos conductos y participan en el desdoblamiento químico de los alimentos.
El tubo digestivo contiene los alimentos desde que se ingieren hasta que se digieren y absorben o eliminan. Las contracciones de la pared del tubo desdoblan físicamente los alimentos al agitarlos. Además, las contracciones ayudan a disolverlos, al mezclarlos con líquidos que se secretan en el propio tubo digestivo. Las enzimas que liberan los accesorios y las células del revestimiento del tubo se encargan del desdoblamiento químico de los contracciones ondulantes del músculo liso de la pared del tubo digestivo impulsan los alimentos por éste desde el esófago hasta el ano.
En general, el aparato digestivo realiza seis básicas:
1. Ingestión. Es el proceso que consiste en llevar alimentos y líquidos a la boca (comer).
2. secreción. Cada día, las células de la pared del tubo digestivo y los órganos auxiliares secretan casi 7 L de agua, ácidos, amortiguadores y enzimas en la luz del tubo digestivo.
3. mezclado y propulsión. La contracción y relajación alternadas del músculo liso de la pared del tubo digestivo mezcla los alimentos y secreciones, además de impulsarlos en dirección al ano. Esta característica del tubo digestivo, de mezclar y mover material en su interior, se denomina motifidad.
4. digestión. Diversos procesos mecánicos y químicos desdoblan los alimentos ingeridos en moléculas más pequeñas. En la digestión mecánica, los dientes cortan y trituran los alimentos antes de deglutirlos, después de lo cual se mezclan por acción del músculo liso del estómago intestino delgado. Como consecuencia de todo ello, las moléculas de los alimentos se disuelven y mezclan con las enzimas digestivas. En la digestión química, las moléculas grandes de hidratos de carbono (carbohidratos), lípidos, proteínas y ácidos nucleicos de los alimentos se dividen en otras más pequeñas, por hidrólisis (fig. 2.16). Las enzimas digestivas, producidas por glándulas salivales, lengua, estómago, páncreas e intestino delgado, catalizan estas reacciones. Unas cuantas sustancias de los alimentos pueden absorberse sin digestión química, entre ellas aminoácidos, colesterol, glucosa, vitaminas, minerales y agua.
5. Absorción. Durante esta fase, los líquidos secretados y las moléculas pequeñas de iones, productos de la digestión, entran en las células epiteliales que revisten la luz del tubo digestivo, por transporte activo o difusión pasiva. Las sustancias absorbidas pasan a la sangre o linfa y circulan a las células de todo el cuerpo.
6. Defecación. Los desechos, sustancias no digeribles, bacterias, células que se esfacelan del revestimiento del tubo digestivo y materiales digeridos que no fueron absorbidos salen del cuerpo por el ano, en el proceso llamado defecación. El material eliminado en ésta se denomina heces o materia fecal.
CAPAS DEL TUBO DIGESTIVO
MUCOSA
La luz del tubo digestivo tiene revestimiento de una membrana, la mucosa.
Este tejido posee tres capas:
1) un
revestimiento de epitelio, que tiene contacto directo con el contenido
del tubo digestivo;
2) una capa subyacente del tejido conectivo areolar, y
3) una capa delgada de músculo liso.
1. El epitelio de boca, faringe, esófago y conducto anal es de tipo escamoso estratificado no queratinizado y desempeña una función protectora. El epitelio cilíndrico simple, que participa en la secreción y la absorción, reviste el estomago y los intestinos. Las células de este segundo tipo epitelio están unidas herméticamente una con otra de manera estrecha, lo cual impide el paso de material entre ellas. La renovación de las células epiteliales digestivas tiene ritmo acelerado: se esfacelan cada cinco a siete días y reponen nuevas células. Entre las epiteliales de absorción, se localizan células exocrinas, que secretan moco y líquido en la luz digestiva, además de varios tipos de ellas, llamadas colectivamente enteroendocrinas, secretan hormonas en la sangre.
2. Lámina propia es tejido conectivo areolar que contiene numerosos vasos sanguíneos y linfáticos, por los cuales se absorben los nutrientes del tubo digestivo para llegar a otros tejidos. Esta capa brinda sostén al epitelio y lo une con la muscular de la mucosa, que se analiza a continuación.
3. Además, la lámina propia incluye gran parte de las células tejido linfoide relacionado con mucosas (TLRM). Estos folículos linfáticos prominentes contienen células del sistema inmunitario, que protegen contra enfermedades. Es un tejido localizado en el tubo digestivo, particularmente en amígdalas, intestino delgado, apéndice e intestino grueso, el cual existen tantas células inmunitarias como las del resto del cuerpo. Los linfocitos y macrófagos del TLRM producen respuestas inmunitarias contra microbios, como las bacterias, que llegan a penetrar el epitelio.
4. Una delgada capa de fibras de músculo liso, la (lámina) muscular de la mucosa, hace que la mucosa de estómago e intestino delgado presente numerosos pliegues pequeños, los cuales incrementan el área de superficie para la digestión y la absorción. Los movimientos de la muscular hacen e todas las células de absorción estén expuestas al contenido del tubo digestivo.
SUBMUCOSA
La submucosa comprende
tejido conectivo areolar que une la mucosa con la tercera capa, la muscular.
Está muy vascularizada y contiene el plexo submucoso o plexo de meissner,
que es una porción del sistema nervioso entérico (SNE). Este último es el
“cerebro del tubo digestivo” y consta de casi 100 millones de neuronas en dos
plexos entéricos, que abarcan todo el tubo digestivo. El plexo de Meissner
incluye neuronas entéricas sensoriales y motoras, además de fibras
postganglionares simpáticas y parasimpáticas que inervan mucosa y submucosa.
Se encarga de regular los movimientos de la mucosa y la constricción de los
vasos sanguíneos. Además inerva las células secretoras de las glándulas de la
mucosa por lo que reviste importancia en el control de las secreciones del
tubo digestivo. Asimismo, la submucosa contiene glándulas y tejido linfático.
MUSCULAR
La muscular de boca, faringe y tercios medio y superior del esófago incluye músculo esquelético, para la deglución voluntaria. Además, dicho músculo forma el esfínter externo del ano, lo cual posibilita el control voluntario de la defecación. En el resto del tubo digestivo, la muscular está formada por músculo liso, generalmente dispuesto en dos capas: una lámina interna de fibras circulares y otra externa de fibras longitudinales. Las contracciones involuntarias del músculo liso ayudan al desdoblamiento físico de los alimentos, su mezclado con las secreciones digestivas y su propulsión por el tubo digestivo. Además, la muscular contiene otro plexo del sistema nervioso entérico, el mientérico o plexo de Auerbach, compuesto de neuronas entéricas, ganglios y fibras postganglionares parasimpáticos, y fibras postganglionares simpáticas que inervan la muscular. Este plexo regula gran parte de la motilidad del tubo digestivo, particularmente en lo relativo a la frecuencia y fuerza de las contracciones de la muscular.
SEROSA
La serosa es la capa superficial de las porciones del tubo digestivo que están suspendidas en la cavidad abdominopélvica. Se compone de tejido conectivo y epitelio escamoso sencillo. Como se verá poco más adelante, el esófago, que cruza el mediastino, tiene una capa superficial, la adventicia, formada por tejido conectivo areolar. En plano infradiafragmático, la serosa se denomina peritoneo visceral y es parte del peritoneo, que a continuación se analiza en detalle.
PERITONEO
El
peritoneo, la serosa más grande del cuerpo, consta de una capa de mesotelio
escamoso sencillo y otra subyacente, de sostén, de tejido conectivo. El
peritoneo parietal reviste la pared de la cavidad abdominopélvica, mientras
que el visceral recubre ciertos órganos de esa cavidad y es su serosa (fig.
24.3a). El angosto espacio entre el peritoneo parietal y el visceral es la
cavidad peritoneal, que contiene líquido seroso. En ciertas enfermedades,
dicha cavidad se distiende por la acumulación de varios litros de líquido,
estado al que se llama ascitis.
Como se verá más adelante, ciertos órganos se sitúan contra la pared abdominal posterior y tienen recubrimiento de peritoneo sólo en su superficie anterior. Esos órganos, como páncreas y riñones, son vísceras retroperitoneales.
A diferencia del pericardio y la pleura, que cubren de manera uniforme el corazón y los pulmones, el peritoneo posee grandes repliegues que se ubican entre las vísceras. Dichos repliegues unen unos órganos con otros y con la pared abdominal además de contener vasos sanguíneos y linfáticos, así como nervios, que irrigan e inervan los órganos abdominales. El mesenterio es un repliegue del peritoneo hacia afuera de la cubierta serosa del intestino delgado (fig. 24.3a, d), cuyo extremo une dicho intestino con la pared abdominal posterior. El mesocolon es otro repliegue peritoneal, que une el intestino grueso con la pared abdominal posterior; también proporciona irrigación a los intestinos mediante vasos sanguíneos y linfáticos. El mesenterio y mesocolon mantienen laxamente en su sitio a los intestinos, ya que permiten una gran movilidad a medida que las contracciones musculares mezclan y mueven el contenido luminal por el tubo digestivo.
Otros repliegues peritoneales de importancia son el ligamento falciforme del hígado y los epiplones menor y mayor. El ligamento falciforme del hígado une esta víscera con la pared abdominal anterior y el diafragma (fig. 24.3b). [El hígado es el único órgano digestivo que se fija a la pared abdominal anterior.] El epiplón menor, que nace de dos pliegues de la serosa de estómago y duodeno a los cuales suspende del hígado (fig. 24.3c), posee algunos ganglios linfáticos. El epiplón mayor, que es el repligue peritoneal más grande, cuelga laxamente como un delantal de grasa” sobre el colon transverso y las asas de intestino delgado (fig. 24.3b, d). Es una doble lámina que se pliega sobre sí misma, con lo que se constituye en una estructura de cuatro capas. Desde sus inserciones en estómago y duodeno, el epiplón mayor se extiende hacia abajo por delante del intestino delgado y luego se pliega y se dirige hacia arriba, hasta unirse al colon transverso. El epiplón mayor contiene tejido adiposo considerable y muchos ganglios linfáticos contribuyen con macrófagos y células plasmáticas productoras de anticuerpos, que ayudan a combatir las infecciones del tubo digestivo e impiden que éstas se diseminen.
APLICACIÓN CLÍNICA
PERITONITIS
La peritonitis es una inflamación aguda del peritoneo. Una de sus causas frecuentes es La contaminación del espacio peritoneal por microbios infecciosos, lo cual puede ser consecuencia de heridas accidentales o quirúrgicas de la pared abdominal, así como de perforación o rotura de vísceras abdominales. Por ejemplo, si las bacterias logran acceso a la cavidad peritoneal por una perforación intestinal o por la rotura del apéndice, pueden ocasionar una forma aguda de peritonitis, que pone en riesgo la vida. Una variante menos grave (aunque también dolorosa) de peritonitis puede ocurrir con la fricción entre superficies peritoneales inflamadas.
BOCA
La boca o cavidad
bucal se forma con los carrillos, paladares duro y blando, y lengua (fig.
24.4). Los carrillos, estructuras musculares cubiertas de piel por fuera y de
epitelio escamoso estratificado no queratinizado por dentro, forman las
paredes laterales de la boca. La porción anterior de los carrillos termina en
los labios.
Los labios son pliegues carnosos que rodean la abertura de la boca. Los cubre
piel en su parte exterior y mucosa en la interior. Hay una zona de transición,
donde se unen los dos tipos de tejido. Dicha área no está queratinizada y el
color de la sangre en los vasos sanguíneos subyacentes es visible a través de
la capa superficial transparente. La superficie interna de cada labio está
unida a la encía correspondiente por un pliegue de mucosa en la línea media,
el frenillo labial.
Entre la piel y la mucosa de la boca, están el músculo orbicular de los labios y el tejido conectivo. Durante la masticación, la contracción de los músculos buccinadores en los carrillos y del orbicular de los labios ayudan a mantener los alimentos entre los dientes superiores e inferiores. Asimismo, tales músculos participan en el habla.
El vestíbulo de la boca es un espacio delimitado externamente por los carrillos y labios, y de manera interna por las encías y los dientes. La boca o cavidad bucal propiamente dicha es el espacio que está entre los dientes y las encías, por un lado, y las fauces, o abertura entre la boca y la faringe, por el otro.
El paladar duro, que es la porción anterior del techo de la boca, se forma con los maxilares superiores y palatinos, tiene recubrimiento de mucosa y constituye una división ósea entre boca y nariz. El pa1adar b1ando, que forma la porción posterior del techo de la boca, es una división muscular en forma de arco con revestimiento de mucosa entre la bucofaringe y nasofaringe.
La úvula es una prolongación muscular cónica que cuelga del borde libre del paladar blando. Durante la deglución, este último y la úvula se retraen en dirección superior, lo cual cierra la nasofaringe e impide que los alimentos y líquidos deglutidos pasen a la nariz. En sentido lateral ala base de la úvula, se observa 2 pliegues musculares con trayecto descendente por los lados del paladar blando. En la parte anterior, el arco glosopalatino se extiende hacia un lado de la base de la lengua, y en la posterior, el arco gopalatino lo hace en dirección a un lado de la faringe. Las amígdalas palatinas se sitúan entre los arcos, y las linguales en la base de la lengua. En el borde posterior del paladar blando, la boca se abre en la bucofaringe a través de las fauces (fig. 24.4).
ESTRUCTURA Y FUNCIÓN DE LAS GLÁNDULAS SALIVALES
Una glándula salival es toda célula u órgano que libere la secreción llamada saliva en la boca. Por lo regular se produce apenas la saliva suficiente para mantener húmeda la mucosa de la boca y faringe, así como limpiar la boca y los dientes. Sin embargo, su producción aumenta cuando entran alimentos en la boca y se encarga de lubricarlos y disolverlos así como de iniciar su digestión química.
La mucosa de la boca y lengua contiene muchas glándulas salivales pequeñas, que se abren directamente en la boca o lo hacen de manera indirecta por conductos cortos. Entre ellas, se incluyen las glándulas labiales, bucales y palatales en los labios, carrillos y paladar, respectivamente, y las glándulas linguales en la lengua; todas ellas contribuyen poco a la producción de saliva. La mayor parte de esta secreción se produce en las glándulas salivales mayores, ubicadas en plano subyacente a la mucosa bucal y cuya secreción se vacía en conductos que se abren en la boca.
Son tres los
pares de glándulas salivales mayores: parótidas mandibulares y sublinguales
(fig. 24.5a). Las glándulas parótidas se localizan en plano anteroinferior a
los oídos, entre la
piel y el músculo masetero. Cada una secreta saliva en la boca por un conducto
parotídeo, que perfora el músculo buccinador y se abre en el vestíbulo frente
al segundo molar inferior. Las glándulas submandibulares se localizan bajo la
base de la lengua, en la parte posterior del suelo de la boca. Sus conductos
submandibulares siguen trayecto bajo la mucosa, a ambos lados de la línea
media del suelo de la boca y entran en la cavidad bucal propiamente dicha de
manera lateral al frenillo lingual. Las glándulas sublinguales tienen
ubicación superior a la de las submandibulares, y sus conductos sublinguales
menores se abren en la cavidad bucal propiamente dicha en el suelo de la boca.
COMPOSICIÓN Y FUNCIONES DE LA SALIVA
Desde el punto de vista químico, la saliva está constituida por 99.5% de agua y 0.5% de solutos. Entre éstos, se incluyen iones como el sodio, potasio, cloruro, bicarbonato y También contiene algunos gases disueltos y diversas sustancias orgánicas, como urea, ácido úrico, moco, inmunoglobulina A, enzima bacteriolítica lisozima y dos enzimas digestivas, la amilasa salival, que actúa en los almidones, y la lipasa lingual, que lo hace en los triglicéridos.
Cada una de las glándulas salivales mayores aporta proporciones distintas de ingredientes a la saliva. Las glándulas poseen células que secretan un líquido acuoso (seroso), el cual contiene amilasa salival. Las glándulas submandibulares poseen células similares a las de las parótidas, además de algunas células mucosas, de modo que liberan un líquido que contiene amilasa espesada con moco. Las sublinguales incluyen ante todo células mucosas, de modo que su secreción es mucho mas espesa y contribuye con poca amilasa a la saliva.
El agua de la saliva constituye un medio para disolver los alimentos, de modo que se puedan saborear y se inicien las reacciones digestivas. Los iones cloruro de la saliva activan la amilasa salival. Los iones bicarbonato y fosfato amortiguan los alimentos ácidos que entran a la boca, por lo cual la saliva es apenas levemente ácida (pH 6.35 a 6.85). La urea y el ácido úrico de la saliva se deben a que las glándulas salivales (al igual que las sudoríparas de la piel) ayudan a eliminar moléculas de desecho. El moco lubrica los alimentos, de modo que se puedan mover fácilmente en la boca, formen el bolo y se deglutan. La inmunoglobulina A es un tipo de anticuerpo que inhibe la proliferación bacteriana, mientras que la lisozima da muerte a bacterias. Aunque estas sustancias ayudan a proteger la mucosa contra infecciones y los dientes contra la caries, no las hay en suficiente cantidad para eliminar todas las bacterias de la boca.
SALIVACIÓN
La secreción de saliva o salivación es regulada por el sistema nervioso. Aunque el volumen de saliva que se secreta diariamente varía mucho, promedia 1 000 a 1 500 ml. En condiciones normales, la estimulación parasimpática fomenta la secreción continua de un volumen moderado de saliva, lo cual mantiene húmeda la mucosa y lubrica los movimientos de la lengua y los labios durante el habla. Luego, se deglute la Saliva y ello ayuda a humectar el esófago. En última instancia, se reabsorbe gran parte de sus componentes, lo cual previene la pérdida de líquidos. La estimulación simpática predomina durante el estrés, por lo que ocurre resequedad de la boca. Durante la deshidratación, las glándulas salivales dejan de secretar saliva para conservar agua, y la resequedad consecuente de la boca contribuye a la sensación de sed. En tales circunstancias, beber agua no sólo restaura la homeostasis corporal, sino que también humecta la boca.
Las acciones de tocar y saborear los alimentos son estímulos importantes de la
secreción de las glándulas salivales. Los compuestos químicos de los alimentos
estimulan a receptores de los botones gustativos de la lengua y los impulsos
correspondientes se transmiten a dos núcleos salivales del tronco encefálico.
Los impulsos parasimpáticos eferentes, que viajan por fibras de los nervios
facial (VII) y glosofaríngeo (IX), estimulan la salivación. Esta continúa de
manera abundante durante algún tiempo después de deglutir los alimentos, lo
cual enjuaga la boca, además de diluir y amortiguar los residuos de compuestos
irritantes.
Oler alimentos, verlos, escuchar sonidos relacionados con ellos o pensar en
ellos también estimula la salivación. Estos estímulos constituyen una
activación psíquica que incluye conductas aprendidas. Cuando surgen recuerdos
que relacionan los estímulos con los alimentos, se propagan impulsos nerviosos
de la corteza a núcleos del tronco encefálico y se activan las glándulas
salivales. Esta activación psíquica resulta útil para el organismo, ya que
permite que se inicie la digestión química en la boca tan pronto se ingieren
los alimentos. Además, ocurre salivación en respuesta a la deglución de
alimentos irritantes o durante la náusea, como consecuencia de reflejos con
origen en el estómago y la porción proximal del intestino delgado. Se supone
que este mecanismo ayuda a diluir o neutralizar la sustancia irritante.
APLICACIÓN CLÍNICA
PAROTIDITIS
Aunque cualquiera de las glándulas salivales puede verse afectada por infecciones de la nasofaringe, es común que el mixovirus ataque las glándulas parotidas. La parotiditis (paperas) es la inflamación y agrandamiento de estas glándulas, con fiebre moderada, malestar general y dolor muy intenso en la garganta, en especial al deglutir alimentos amargos o jugos ácidos. La inflamación ocurre en uno o ambos lados de la cara, justo en plano anterior al ramo del maxilar inferior. En casi 30% de varones adolescentes o adultos afectados, también se inflaman los testículos, si bien pocas veces ocasiona esterilidad, ya que la afección suele ser unilateral. La incidencia de esta enfermedad ha disminuido desde 1967 con la disponibilidad de una vacuna contra el virus.
ESTRUCTURA Y FUNCIÓN DE LA LENGUA
La lengua es un órgano accesorio digestivo que se compone de músculo esquelético cubierto con mucosa. Junto con los músculos relacionados con ella, forma el suelo de la cavidad bucal. Se divide en mitades laterales simétricas gracias a un tabique que se extiende a todo lo largo de su línea media y se inserta de manera inferior en el hioides, apófisis estiloides del temporal y maxilar inferior. Cada mitad consta de los mismos músculos extrínsecos e intrínsecos.
Los músculos extrínsecos de la lengua, que se originan fuera de ella (es
decir, en huesos del área) y se insertan en ella, son el hiogloso, genio gloso
y estiogloso (fig. 11.7). Estos músculos mueven la lengua de un lado a otro y
de adelante atrás para desplazar los alimentos durante la masticación, dan
forma a la masa redondeada llamada bolo alimenticio y fuerzan su paso a
la parte posterior de la boca para su deglución. Además, forman el suelo de la
boca y mantienen la lengua en su posición. Los músculos intrínsecos, que se
originan e insertan en el tejido conectivo de la lengua, modifican el tamaño y
la forma de ésta para el habla y la deglución. Son el lingual superior,
lingual inferior, transverso y vertical de la lengua. El frenillo lingual es
un pliegue de mucosa en la línea media de la cara inferior de la lengua que la
fija al suelo de la boca y participa en la limitación de sus movimientos
posteriores (figs. 24.4 y 24.5). Cuando este frenillo es anormalmente corto o
rígido, trastorno llamado anquiloglosia, comer y hablar se dificultan tanto
que se dice que ala persona “se le traba la lengua”.
El dorso o cara superior y las caras laterales de la lengua están recubiertos de papilas, las cuales son prolongaciones de la lámina propia recubiertas de epitelio queratinizado (fig. 16.2a). Muchas papilas contienen botones gustativos, que son los receptores del gusto. Las papilas fungiformes son proyecciones a manera de hongo distribuidas entre las papilas filiformes y resultan más numerosas cerca de la punta de la lengua. Se observan como puntos rojos en la superficie de este órgano y muchas de ellas contienen botones gustativos. Las papilas circunvaladas forman una V invertida en la cara posterior de la lengua y todas poseen botones gustativos. Por último, las papilas filiformes son proyecciones cónicas blancuzcas distribuidas en filas paralelas sobre los dos tercios anteriores de la lengua. Aunque carecen de botones gustativos, aumentan la fricción entre la lengua y los alimentos, lo cual facilita a dicho órgano mover las partículas de alimentos en la boca. Las glándulas linguales de la lámina propia secretan moco y un líquido seroso (acuoso), que contiene lipasa lingual, que es parte de la saliva.
ESTRUCTURA Y FUNCIÓN DE LOS DIENTES
Los dientes (fig. 24.6) son órganos digestivos que se localizan en los alvéolos de las apófisis del mismo nombre de ambos maxilares. Dichas apófisis están cubiertas por las encías, que se extienden un poco en cada alvéolo y forman el surco gingival. Los alvéolos tienen revestimiento del ligamento periodontal, el cual esta formado de tejido conectivo fibroso denso y se inserta en las paredes alveolares y en la superficie de cemento de la raíz de los dientes. De tal suerte, fija a los dientes en su posición y actúa como amortiguador de impactos durante la masticación.
Un diente consta
de tres partes principales. La corona es la porción visible por arriba del
nivel de las encías. Posee unas tres raíces, incrustadas en el alvéolo. El
cuello es la porción angosta en la unión de la corona y las raíces, cerca del
contorno gingival.
Los dientes se componen principalmente de dentina tejido conectivo calcificado
que les confiere su forma y rigidez fundamentales. La dentina es más dura que
los huesos por su contenido más alto de sales de calcio (70% de su peso seco).
Dicho tejido envuelve la cavidad pulpar, situada dentro de la corona y llena
de pulpa, tejido conectivo que contiene vasos sanguíneos y linfáticos, así
como nervios. Las prolongaciones angostas de la cavidad pulpar, los conductos
radiculares, tienen trayecto a lo largo de las raíces y presentan un orificio
en su base, el agujero apical, por el cual pasan los vasos sanguíneos,
linfáticos y nervios.
La dentina de la
corona tiene recubrimiento de esmalte, que consta principalmente de fosfato y
carbonato de calcio. El esmalte, la sustancia más dura del cuerpo y la de
mayor contenido de sales de calcio (casi 95% de su peso seco) protege los
dientes del desgaste propio de la masticación, además de ser una barrera
contra ácidos que disolverían fácilmente la dentina. Esta última tiene, en la
raíz, recubrimiento de cemento, también parecido al tejido óseo, que une la
raíz con el ligamento periodontal.
La rama de la odontología que estudia la prevención, el diagnóstico y
tratamiento de enfermedades que afectan pulpa, raíces, ligamento periodontal y
alveolos es la endodoncia. La disciplina que se encarga de la prevención y
corrección de la alineación anormal de los dientes es la ortodoncia, mientras
que el tratamiento de anormalidades de los tejidos adyacentes a los dientes
corresponde a la periodoncia. Los seres humanos poseen dos denticiones o
conjuntos de dientes: decidua y permanente. La primera que forma los dientes
deciduos, primarios o de leche, empieza a brotar hacia los seis meses de edad
ya partir de entonces aparece un par de dientes aproximadamente cada mes hasta
que hay 20 (fig. 24.7a). Los incisivos son los más cercanos a la
línea media,
tienen forma de bisel y están adaptados cortar los alimentos. Se les denomina
incisivos centrales o laterales con base en su posición. Junto a ellos, en
dirección posterior, están los caninos (cuspídeos), provistos de una
superficie puntiaguda, llamada cúspide, que se usa para desgarrar los
alimentos. Tanto incisivos como caninos poseen una sola raíz.
En el plano posterior a ellos, están el primero y segundo molares, que tienen cuatro cúspides. Los molares superiores poseen tres raíces, y los inferiores, dos. Estos dientes se utilizan para triturar los alimentos.
Todos los dientes deciduos se caen, generalmente entre los seis y 12 años reemplazan los dientes permanentes o secundarios (fig. 24.7b).
La dentición permanente incluye 32 dientes que brotan entre la edad señalada y la adulta. La distribución es similar a la de los dientes primarios, con las excepciones siguientes. El primero y segundo premolares (bicúspides), provistos de dos cúspides y una raíz (los premolares superiores poseen dos raíces), que se usan para triturar, sustituyen a los molares deciduos. Los molares permanentes, que brotan en plano distal a los premolares, no reemplazan dientes primarios y aparecen a medida que los maxilares crecen para darles cabida: los primeros molares a los seis años, los segundos molares a los 12 años y los terceros molares (muelas del juicio) después de los 17 años.
Es frecuente que en seres humanos no haya espacio suficiente, en plano posterior a los segundos molares, para que broten los terceros molares. En tal caso, éstos quedan incluidos en el hueso alveolar y se dice que están “impactados”. Ello suele causar presión y dolor, lo cual obliga a extirparlos quirúrgicamente. Además, en algunas personas el tercer molar es muy pequeño o no se desarrolla en absoluto.
DIGESTIÓN MECÁNICA Y QUÍMICA EN LA BOCA
La digestión mecánica en la boca es resultado de la masticación, en que la lengua mueve los alimentos y los dientes los trituran, además de que se mezclan con la saliva. En consecuencia, se reducen a una masa blanda, flexible y de fácil deglución, llamada bolo alimenticio. Los alimentos empiezan a disolverse en el agua de la saliva, lo cual reviste importancia porque las enzimas sólo pueden reaccionar con las moléculas alimenticias en un medio líquido.
Dos enzimas contribuyen a la digestión química en la boca, la amilasa salival y la lipasa lingual. La amilasa salival inicia la digestión de los almidones. Los hidratos de carbono de los alimentos son monosacáridos y disacáridos o polisacáridos complejos, como los almidones. Aunque la mayoría de los hidratos de carbono ingeridos son almidones, solo los monosacáridos pueden absorberse en la sangre. Así pues los disacáridos y almidones deben convertirse en monosacáridos. La función de la amilasa salival es romper ciertos enlaces químicos entre las unidades de glucosa de los almidones , lo cual reduce polisacáridos de cadena larga al disacárido maltosa, trisacárido maltotriosa y los polímeros de glucosa de cadena corta llamados a-dextrinas. Aun cuando por lo general se degluten los alimentos con rapidez excesiva para que ocurra su conversión bucal en disacáridos, la amilasa salival de los alimentos deglutidos continúa actuando durante casi una hora, después de lo cual se inactiva por efecto los ácidos gástricos. Por otra parte, la saliva contiene lipasa lingual, que empieza la digestión de los triglicéridos de los alimentos en ácidos grasos y monoglicéridos. Esta enzima se activa en el medio ácido del estómago, por lo que comienza a ejercer su acción después de deglutir los alimentos. Las actividades digestivas en la boca se resumen en el cuadro 24.1.
FARINGE
Al deglutir los alimentos, pasan de la boca a la faringe, o conducto ahusado que va de los orificios posteriores de la nariz al esófago en el plano posterior y a la laringe en el anterior (fig. 23.4). La faringe se compone de músculos esqueléticos con revestimiento de mucosa. La nasofaringe participa únicamente en la respiración, mientras que la bucofaringe y laringofaringe tienen funciones digestivas y respiratorias. Los alimentos deglutidos pasan de la boca a la bucofaringe y laringofaringe, cuyas contracciones musculares ayudan a impulsar los alimentos hacia el esófago.
El movimiento de los alimentos de la boca al estómago se logra con el acto de la deglución (fig. 24.8). Esta, que se facilita con la saliva, incluye la participación de la boca, faringe y esófago. Hay tres etapas en la deglución: 1) voluntaria, en que pasa el bolo a la bucofaringe;
2) faríngea, que es el movimiento involuntario del bolo alimenticio por la faringe al esófago, y 3) esofágica, que se analiza en la sección sobre el esófago y es el desplazamiento también involuntario del bolo por el esófago al estomago.
La deglución se inicia cuando la persona desplaza el bolo alimenticio a la parte posterior de la cavidad bucal y de ésta a la bucofaringe gracias al movimiento de la lengua hacia arriba y atrás, contra el paladar, acciones que constituyen la etapa voluntaria. Una vez que el bolo está en la bucofaringe, comienza la etapa faríngea (involuntaria) [fig. 24.8b]. Se cierra el acceso a las vías respiratorias inferiores y se interrumpe transitoriamente su función. El bolo alimenticio estimula receptores de la bucofaringe, que envían impulsos al centro de la deglución, del bulbo raquídeo y la porción inferior del puente, en el tronco encefálico. Los impulsos diferentes que llegan de esos centros hacen que el paladar blando y la úvula se desplacen hacia arriba para ocluir la nasofaringe, además de que la laringe se mueve en dirección anterosuperior, bajo la lengua. Al ascender la laringe, la epiglotis se mueve hacia atrás y abajo, con lo que se sella la rima glottidis. El movimiento de la laringe también tira de las cuerdas vocales, con lo que se sella el acceso a las vías respiratorias inferiores, además de ensanchar la abertura entre la laringofaringe y el esófago. El bolo cruza la laringofaringe y entra al esófago en uno a dos segundos. Luego, se abren de nuevo vías respiratorias y se reanuda su función.
ESÓFAGO
El esófago es un conducto muscular colapsable que se localiza en plano posterior a la tráquea y tiene unos 25 cm de longitud. Se inicia en el extremo inferior de la laringofaringe, cruza el mediastino en plano anterior a la columna vertebral, perfora el diafragma en el orificio 11amado hiato esofágico y termina en el extremo superior del estómago(fig. 24.1).
CARACTERÍSTICAS HISTOLÓGICAS DEL ESÓFAGO
La mucosa del esófago está formada por epitelio escamoso estratificado no queratinizado, lámina propia (tejido conectivo areolar) y muscular de la mucosa (músculo liso) (fig. 24.9). Cerca del estómago, la mucosa esofágica también contiene glándulas mucosas. El epitelio escamoso estratificado de labios, lengua, boca, bucofaringe, laringofaringe y esófago brinda protección considerable contra la abrasión y el uso y desgaste relacionados con la masticación, mezclado de las partículas de alimentos con secreciones, y deglución. La submucosa incluye tejido conectivo areolar, vasos sanguíneos y glándulas mucosas. La muscular del tercio esofagico superior consta de músculo esquelético y liso; y la del tercio inferior, de músculo liso. La capa superficial se denomina adventicia, no serosa, ya que no tiene recubrimiento de mesotelio y el tejido conectivo se fusiona con el de estructuras circundantes del mediastino, el cual cruza. La adventicia une el esófago con dichas estructuras.
FISIOLOGÍA DEL ESÓFAGO
El esófago secreta moco y transporta los alimentos al estomago. No produce enzimas digestivas ni lleva a cabo la absorción. El paso de los alimentos de la laringofaringe al esófago ésta regulado en el extremo superior del esófago por un esfínter (banda circular o anillo de músculo que normalmente está contraído), el esfínter esofágico superior, que se compone músculo faríngeo inferior y su inserción en el cartílago cricoides. La elevación de la laringe durante la etapa faríngea de la deglución hace que se relaje dicho esfínter y el bolo alimenticio pase al esófago. El esfínter también se relaja durante la exhalación.
En la etapa esofágica de la deglución, una sucesión de iones y relajaciones coordinadas de las capas circular y longitudinal de la muscular, proceso llamado peristaltismo, desplaza el bolo alimenticio hacia abajo (fig.24.10). El peristaltismo también ocurre en otras estructuras tubulares, lo que incluye otras partes del tubo digestivo, uréteres, vías biliares y trompas de falopio; en el esófago, está bajo regulación del bulbo raquídeo. En la porción del esófago inmediatamente proximal al bolo alimenticio, se contraen las fibras musculares circulares, lo cual constriñe la pared esofagica e impulsa el bolo hacia el estómago. Entretanto, también ocurre la contracción de las fibras longitudinales dístales al propio bolo, lo cual acorta esta sección inferior y dilata la esofágica, de modo que pueda recibir el bolo. Las contracciones se repiten en una onda que impulsa los alimentos hacia el estómago. El moco que secretan las glándulas esofágicas lubrica el bolo y reduce la fricción. El paso de alimentos sólidos o semisólidos de la boca al estómago tarda de cuatro a ocho segundos, y el de los alimentos muy blandos y líquidos, apenas un segundo.
En plano inmediatamente superior al diafragma, el esófago se angosta levemente como resultado de la contracción sostenida de la muscular en esa parte de dicho órgano. Se trata de un esfínter fisiológico, llamado esfínter esofágico inferior, el cual se relaja durante la deglución para que el bolo pase del esófago al estómago.
Las actividades digestivas de la faringe y el esófago se resumen en el cuadro 24.2.
APLICACIÓN CLÍNICA
ENFERMEDAD POR REFLUJO GASTROESOFÁGICO
Cuando el esfínter esofágico inferior no se cierra adecuadamente después de que los alimentos entran en el estómago, es posible el reflujo del contenido gástrico a la porción inferior del esófago, trastorno llamado enfermedad por reflujo gastroesofágico (ERGE). EL ácido clorhídrico (HCI), que proviene del estómago, puede irritar la pared esofágica y causar una sensación de quemadura, llamada pirosis. La ingestión de alcohol y el tabaquismo relajan el esfínter, lo cual agrava el problema. Los síntomas de la ERGE generalmente pueden controlarse al evitar alimentos que estimulen intensamente la secreción de ácidos gástricos, como café, chocolate, jitomates, alimentos grasosos, menta, menta verde y cebollas. Otras estrategias para reducir la acidez es el uso, unos 30 a 60 minutos antes de comer, de bloqueadores de la histamina H2 de venta libre, además de neutralizar el ácido ya secretado mediante antiácidos. Los síntomas son menos probables si se ingieren pequeñas raciones y la persona no se acuesta inmediatamente después de las comidas. La ERGE suele guardar relación con el cáncer del esófago.
ESTÓMAGO
El estómago es una dilatación característica del tubo digestivo en forma de J, situada directamente abajo del diafragma en el epigastrio, hipocondrio izquierdo y región umbilical del abdomen (fig. l.12a). Se ubica entre el esófago y el duodeno; este último es la primera porción de intestino delgado (fig. 24.11). La ingestión de alimentos, puede ser mucho mayor que la capacidad de los intestinos para digerirlos y absorberlos, por lo que una función del estómago es servir como sitio de mezclados y depósito de retención. A intervalos apropiados después de ingeridos los alimentos, el estómago transfiere una pequeña parte de su contenido a la porción inicial del intestino delgado. La posición y tamaño gástricos varían de manera continua; el diafragma empuja el estomago hacia abajo con cada inspiración y tira de el hacia arriba con cada espiración. Cuando está vacío, tiene un tamaño como el de un salchichón; pero constituye la porción más distensible del tubo digestivo y puede dar cabida a un gran volumen de alimentos. En el estómago, continúa la digestión de los almidones, se inicia la de proteínas y triglicéridos, el bolo alimenticio semisólido se convierte en Líquido y se absorben ciertas sustancias.
ANATOMÍA DEL ESTÓMAGO
El estómago tiene cuatro regiones principales: cardias, fondo, cuerpo y píloro (fig. 24.11). El cardias rodea la abertura superior del estómago. La porción redondeada que está arribaba y a la izquierda del cardias es el fondo. En plano inferior a éste, está la gran porción central del estómago, su cuerpo. La parte que conecta este órgano con el duodeno es el píloro, que incluye dos partes: el antro pilórico, conectado el cuerpo del estómago, y el conducto pilórico, que se conecta con el duodeno. Cuando el estómago está vacío, la mucosa presenta grandes pliegues, llamados arrugas, observables a simple vista. El píloro se comunica con el duodeno del intestino delgado mediante el llamado esfínter pilórico. El borde medial cóncavo del estómago es su curvatura menor, y su borde convexo lateral, la curvatura mayor.
APLICACIÓN CLINICA
PILOROESPASMO Y ESTENOSIS PILÓRICA
En lactantes, puede haber dos anormalidades del esfínter pilórico. En el píloro espasmo, las fibras musculares del esfínter no se relajan de manera normal, de modo que alimentos no pasan fácilmente del estómago al intestino delgado; el primero se llena excesivamente y el lactante tiende al vómito para aliviar la presión. Este trastorno se trata con medicamentos que relajan las fibras musculares del esfínter. La estenosis pilórica es el angostamiento del esfínter pilórico y debe corregir quirúrgicamente. Su signo cardinal es el vómito en proyectil, es decir, el que se lanza a cierta distancia del lactante.
CARACTERÍSTICAS HISTOLÓGICAS DEL ESTÓMAGO
La pared gástrica se compone de las mismas cuatro capas básicas que el resto del tubo digestivo, con ciertas modificaciones (fig. 24.12a). La superficie de la mucosa es una capa de células epiteliales cilíndricas sencillas, llamadas células de la superficie de la mucosa. La mucosa incluye lámina propia (tejido conectivo areolar) y muscular de la mucosa (músculo liso). Las células epiteliales llegan hasta la lámina propia, donde forman columnas de células secretoras, llamadas glándulas gástricas, que revisten muchos conductos angostos, las criptas o fositas gástricas. Las secreciones de varias glándulas gástricas fluyen a cada cripta gástrica y luego a la luz del estómago.
Las glándulas gástricas poseen tres tipos de células de glándulas exocrinas, que secretan su producto en la luz del estómago: células mucosas del cuello, las principales y las parietales. Tanto las células de la superficie de la mucosa como las del cuello de la mucosa secretan moco (fig. 24.12b). Las células principales (zimógenas) liberan pepsinógeno y lipasa gástrica.
Las células parietales producen ácido clorhídrico y factor intrínseco (necesario para la absorción de la vitamina B12). Las secreciones de los tres tipos celulares forman el jugo gástrico, cuyo volumen es de 2 a 3 L diarios. Además, las glándulas gástricas incluyen un tipo de células enteroendocrinas, las células G, que se localizan ante todo en el antro pilórico y que secretan la hormona gastrina en la sangre. Como se verá poco más adelante, esta hormona estimula varios aspectos de la actividad gástrica.
Hay otras tres capas en plano profundo a la mucosa. La submucosa del estómago está compuesta de tejido conectivo areolar. La muscular tiene tres capas (no dos) de músculo liso, a saber, longitudinal externa, circular intermedia y oblicua interna. De ellas, la tercera se limita principalmente al cuerpo del estómago. La serosa (mesotelio escamoso sencillo y tejido conectivo areolar) que recubre el estómago es parte del peritoneo visceral. En la curvatura menor, este último se continúa hacia arriba, en dirección al hígado, con el nombre de epiplón menor. En la curvatura mayor, dicho peritoneo se continúa hacia abajo con el nombre de epiplón mayor, que recubre los intestinos.
DIGESTIÓN MECÁNICA Y QUÍMICA EN EL ESTÓMAGO
Varios minutos después de que los alimentos entran en el estómago, se producen movimientos peristálticos suaves, llamados ondas de mezclado, a intervalos de 15 a 25 segundos, las cuales maceran los alimentos, los mezclan con las secreciones de las glándulas gástricas y los reducen a un líquido parecido a una sopa, el quimo. Se observan unas cuantas ondas de mezclado en el fondo, que desempeña principalmente funciones de almacenamiento. Al ocurrir la digestión en el estómago, se inician ondas de mezclado más intensas en el cuerpo gástrico y se intensifican a medida que se acercan al píloro. El esfínter de este último normalmente está casi cerrado, de modo que al llegar los alimentos a esta abertura cada onda de mezclado fuerza el paso de varios mililitros de quimo al duodeno por el esfínter. La mayor parte del quimo regresa al cuerpo del estómago, donde continúa su mezclado. La siguiente onda lo empuja de nuevo hacia el duodeno y fuerza el paso de un pequeño volumen de ese líquido al intestino delgado. Estos movimientos anterógrados y retrógrados son el origen de gran parte del mezclado gástrico.
Los alimentos suelen permanecer en el fondo durante poco más o menos una hora sin mezclarse con el jugo gástrico. En dicho lapso, continúa la acción digestiva de la amilasa salival. Sin embargo, poco después el quimo se mezcla con el jugo gástrico, lo cual inactiva la amilasa salival y activa la lipasa lingual, de modo que se inicia la digestión de los triglicéridos en ácidos grasos y diglicéridos.
Aunque las células parietales secretan por separado los iones hidrógeno (H+) y cloruro (Cl-) en la luz gástrica, su efecto neto es la secreción de ácido clorhídrico (HC1). La bomba de protones que alimenta la ATPasa de H+/K+ transporta activamente los H+ a la luz gástrica y lleva los iones potasio (K+) a las células. Al mismo tiempo, los iones Cl- y K+ difunden fuera de las células por los canales respectivos de la membrana apical, situados junto a la luz gástrica. En la membrana basolateral, que se encuentra frente a la lámina propia, los antitransportadores llevan los Cl- a las células parietales, a cambio de HCO-3, que difunde hacia los capilar sanguíneos. El resultado es una “marea alcalina” de iones bicarbonato, que entran al torrente sanguíneo después de las comidas. Además, el líquido muy ácido del estómago da muerte a muchos microbios de los alimentos, mientras que el HCl desnaturaliza (desdobla) parcialmente las proteínas de los alimentos y estimula la secreción de hormonas que promueven el flujo de bilis y jugo pancreático.
La digestión enzimática de las proteínas se inicia en el estómago. En éste, la única enzima proteolítica (que digiere proteínas) es la pepsina, que secretan las células principales. Esta desdobla ciertos enlaces peptídicos de los aminoácidos que componen las proteínas, por lo que una cadena técnica de muchos aminoácidos se desdobla en fragmentos peptídicos más pequeños. La pepsina posee eficacia máxima en el ambiente muy ácido del estómago (pH 2) y se inactiva cuando disminuye la acidez.
¿Qué evita la digestión de las células gástricas por la pepsina, junto con los alimentos? En primer término esta enzima es secretada en una forma inactiva, llamada pepsinógeno, en el cual no puede digerir las proteínas de las células principales que la producen. El pepsinógeno no se convierte en pepsina activa hasta que tiene contacto con otras moléculas de la misma o con el ácido clorhídrico que tan las células parietales. En segundo lugar, las células del epitelio gástrico están protegidas contra los jugos gástricos por una capa de 1 a 3 mm de espesor de moco alcalino, que secretan las células superficiales de la mucosa y las del cuello de la mucosa.
Otra enzima del estómago es la lipasa gástrica, que desdobla los triglicéridos de cadena corta de las moléculas de grasa que se encuentran en la leche en ácidos grasos y monoglicéridos. Esta enzima, de actividad limitada en el estómago de adultos, funciona óptimamente con pH 5 a 6. De mayo importancia que las lipasas lingual o gástrica es la pancreática enzima que secreta el páncreas en el intestino delgado.
Es mínima la
absorción que ocurre en el estómago, ya que sus células epiteliales son
impermeables a la mayoría de los materiales. Sin embargo, las células mucosas
gástricas absorben algo de agua, iones y ácidos grasos de cadena corta, al
igual que ciertos medicamentos (en particular, el ácido a acetilsalicílico) y
el etanol.
Las actividades digestivas del estómago se resumen cuadro 24.3.
REGULACIÓN DE LA SECRECIÓN Y MOTILIDAD GÁSTRICAS
Tanto mecanismos neurales como hormonales regulan la secreción de jugo gástrico y la contracción del músculo liso de la pared estomacal. Los fenómenos de la digestión gástrica ocurren en tres fases que se sobreponen: cefálica, gástrica e intestinal (fig. 24.13).
FASE CEFÁLICA
La fase cefálica de la digestión gástrica consiste en reflejos que inician receptores sensoriales de la cabeza. Incluso antes de que los alimentos entren en el estómago, dichos reflejos se activan al ver, oler o saborear alimentos y al pensar en ellos. La corteza cerebral y el centro de la alimentación hipotalamico envían impulsos nerviosos al bulbo raquídeo, que a vez los transmite a fibras preganglionares parasimpáticas del nervio vago (X), lo cual estimula fibras postganglionares parasimpáticas del plexo submucoso. Luego, las fibras parasimpáticas transmiten impulsos a las células parietales, principales y mucosas, y se incrementa la secreción de todas las glándulas gástricas. En éstas, los impulsos estimulan la liberación de pepsinógeno, ácido clorhídrico y moco en el quimo gástrico, así como de gastrina en la sangre. Los impulsos de las fibras parasimpáticas también incrementan la motilidad gástrica. Emociones como ira, miedo y ansiedad tienden a acelerar la digestión, ya que estimulan el sistema nervioso simpático, que inhibe la actividad gástrica
FASE GÁSTRICA
Una vez que los alimentos llegan al estómago, los receptores sensoriales de éste activan mecanismos neurales y hormonales para que continúen la secreción y motilidad gástricas, lo que se conoce como fase gástrica (fig. 24.13). Todo tipo de alimentos distiende (estira) el estómago y estimula los receptores de estiramiento en sus paredes, así como quimiorreceptores que vigilan el pH del quimo gástrico. Cuando distienden las paredes gástricas o aumenta el pH porque entran proteínas al estómago y amortiguan una parte de los ácidos gástricos, se activan los receptores de estiramiento y quimiorreceptores, y con ellos, un ciclo de retroalimentación negativa neural (fig. 24.14). Desde los receptores, se propagan impulsos nerviosos al plexo submucoso, donde activan las fibras entéricas y parasimpáticas. Los impulsos nerviosos resultantes causan ondas de peristaltismo y continúan estimulando el flujo de jugo gástrico que proviene de las células parietales, principales y mucosas.
Las ondas de peristaltismo mezclan los alimentos con el jugo gástrico, y cuando adquieren intensidad suficiente, un pequeño volumen de quimo, aproximadamente 10 a 15 mL (dos a tres cucharaditas) cruza el esfinter pilórico hacia el duodeno. Al volverse nuevamente más ácido el quimo gástrico y reducirse la distensión de las paredes del estómago con el paso del quimo al intestino delgado, este ciclo de retroalimentación negativa inhibe la secreción de jugo gástrico.
La retroalimentación negativa hormonal también regula la secreción gástrica durante la fase del mismo nombre (fig. 24.13). Las proteínas digeridas parcialmente amortiguan los H+, con lo que incrementan el pH, mientras que los alimentos ingeridos distienden el estómago. Los quimiorreceptores y receptores de estiramiento detectan estos cambios y estimulan la liberación de acetilcolina de las fibras parasimpáticas. A su vez, la acetilcolina induce la secreción de la hormona gastrina de las células G, que son las enteroendocrinas de la mucosa del antro pilórico. (Una pequeña cantidad de gastrina es también secretada por las células enteroendocrinas de] intestino delgado, además de que ciertos compuestos de lo alimentos, como la cafeína, estimulan en forma directa la liberación de gastrina.) Esta pasa al torrente sanguíneo y finalmente llega a sus células blanco, las glándulas gástricas.
La gastrina estimula el crecimiento de las glándulas gástricas y la secreción de grandes volúmenes de jugo gástrico. Además, fortalece las contracciones del cardias, aumenta la motilidad gástrica y relaja los esfínteres pilórico e ileocecal que se analizan más adelante. La secreción de gastrina se inhibe cuando el pH de jugo gástrico disminuye a menos de 2, mientras que su aumento la estimula. Este mecanismo de retroalimentación negativa ayuda a lograr el pH bajo óptimo para el funcionamiento de la pepsina, la acción microbicida y la desnaturalización de proteínas en el estómago.
La acetilcolina que liberan las fibras parasimpáticas y la que secretan las células G estimulan, en las parietales, secreción de más HC1 en presencia de histamina. En otras palabras, la histamina, que es una sustancia paracrina que liberan las células cebadas de la lámina propia y actúan en las células parietales adyacentes, tiene efecto sinérgico con la acetilcolina y gastrina, cuyas acciones intensifica. Hay receptores de las tres sustancias en la membrana plasmática de las células parietales. Los de histamina en dichas células se llaman receptores H2 y median respuestas distintas a las de los receptores H1 que participan en las reacciones alérgicas.
FASE INTESTINAL
La fase intestinal de la digestión gástrica se debe a la acción de receptores en el intestino delgado. Los reflejos que se inician en las fases cefálica y gástrica estimulan la actividad secretora y motilidad en el estómago, mientras que los los reflejos de la fase intestinal tienen efectos inhibitorios (flg.24.13), que desaceleran la salida de quimo del estómago para evitar que el duodeno se sobrecargue con este líquido excesivo. Además, las respuestas de la fase intestinal promueven la digestión continuada de los alimentos que llegan al intestino delgado. Cuando el quimo que contiene ácidos grasos y glucosa pasa del estómago al duodeno, hace que las células enteroendocrinas de la mucosa del intestino delgado liberen en la sangre dos hormonas con efecto en el estómago, la secretina colecistocinina (CCC). En el estómago, la secretina tiene como efecto principal la disminución de las secreciones gástricas, mientras que la CCC inhibe ante todo el vaciado gástrico. Ambas hormonas ejercen otros efectos de importancia en el páncreas, hígado y la vesícula biliar (los cuales se explican poco más adelante) y contribuyen a la regulación de los os digestivos.
REGULACIÓN DEL VACIADO GÁSTRICO
El vaciado gástrico, o paso periódico del quimo del estomago al duodeno, está regulado por reflejos neurales y hormonales, como sigue (fig. 24.15a):
1. Estímulos como la distensión del estómago y la presencia de proteínas digeridas parcialmente, etanol y cafeína inician el vaciado gástrico.
2. Estos estímulos hacen que aumente la secreción de gas y se generen impulsos parasimpáticos en el nervio vago (X).
3. La gastrina y los impulsos nerviosos estimulan la contracción del cardias, aumentan la motilidad gástrica y relajan el esfínter pilórico.
4. El efecto neto de estas acciones es el vaciado gástrico.
Los reflejos neurales y hormonales también hacen que el estomago no transfiera al intestino delgado más quimo que el que pueda procesar. El reflejo neural llamado reflejo enterogástrico y la colecistocinina inhiben el vaciado gástrico, de la manera siguiente (fig. 24.15b):
1. Estímulos como la distensión duodenal y la presencia de ácidos grasos, glucosa y proteínas digeridas parcialmente en el quimo duodenal inhiben el vaciado gástrico.
2. Tales estímulos inician el reflejo enterogástrico, en que se propagan impulsos nerviosos del duodeno al bulbo raquídeo, donde inhiben la estimulación parasimpática y excitan la actividad simpática en el estómago. Esos mismos estímulos también incrementan la secreción de colecistocinina.
3. El aumento de los impulsos simpáticos y de la secreción de colecistocinina disminuye la motilidad gástrica.
4. El
efecto neto de esas acciones es la inhibición del vaciado gástrico.
En las dos a cuatro horas que siguen a la ingestión de una comida, el estómago termina de vaciar su contenido en el duodeno. Los alimentos con gran cantidad de hidratos de carbono son los que están menos tiempo en el estómago, mientras que los ricos en proteínas permanecen un poco más, y el vaciado tiene lentitud máxima después de una comida grasosa, que contenga muchos triglicéridos. La razón de esto último es que los ácidos grasos del quimo estimulan la liberación de colecistocinina, lo cual desacelera el vaciado gástrico.
APLICACIÓN CLINICA
VÓMITO
El vómito o emesis es la expulsión con fuerza del contenido gastrointestinal superior (estómago y, a veces, duodeno) por la boca. El estímulo más importante para el vómito es la irritación y distensión del estómago; otros son escenas desagradables, anestesia general, mareos y ciertos medicamentos, como la morfina y los derivados de la digital. En dicho fenómeno, se transmiten impulsos nerviosos al centro del vómito, en el bulbo raquídeo, y los impulsos eferentes se propagan a Los órganos gastrointestinales superiores, al diafragma y a los músculos abdominales. En lo fundamental, el vómito implica la compresión del estómago entre el diafragma y los músculos abdominales, con la expulsión de su contenido por los esfínteres esofágicos abiertos. El vómito prolongado, ante todo en lactantes y personas de edad avanzada, puede revestir gravedad porque la pérdida de jugo gástrico ácido genera alcalosis (pH sanguíneo mayor que el normal).
PANCREAS
Desde el estomago, el quimo pasa al intestino delgado. La digestión química en este depende de la actividad del páncreas, hígado y vesícula biliar, por lo que primero se consideran las actividades de estos órganos accesorios y su contribución a la digestión en el intestino delgado.
ANATOMÍA DEL PÁNCREAS
El páncreas es una glándula retroperitoneal de unos 12 a 15 cm de longitud y 2.5 cm de espesor, situada en el plano posterior a la curvatura mayor del estómago. Consta de cabeza, cuerpo y cola; se conecta al duodeno, generalmente por dos conductos (fig. 24.16). La cabeza es la porción expandida del órgano cerca de la curva del duodeno, mientras que en el plano superior a ella y a la izquierda están el cuerpo o porción central y la cola de forma ahusada.
Las secreciones pancreáticas pasan de las células secretoras del páncreas a pequeños conductos, que en ultima instancia se unen para formar dos de gran calibre, que vacían secreciones en el intestino delgado. El mayor de los dos es el conducto pancreático o de Wirsung. En la mayoría de las personas, este conducto se une al colédoco del hígado y la vesícula biliar, con el cual entra al duodeno en la ampolla de Vater. Ésta se abre en una prominencia de la mucosa, la llamada papila duodenal, unos 10 cm en sentido distal al esfínter pilórico. El menor de los dos conductos pancreáticos, el de Santorini, se vacía en el duodeno unos 2.5 cm en sentido proximal a la ampolla de Vater.
CARACTERÍSTICAS HISTOLÓGICAS DEL PÁNCREAS
El páncreas se compone de pequeños grupos de células epiteliales glandulares, de las cuales casi 99% esta dispuesto en grupos llamados acinos, que constituyen la porción exocrina de la glándula (fig. 18.18b). Las células de los acinos secretan una mezcla de líquido y enzimas digestivas, el jugo pancreático. El 1% restante de las células esta organizado en los grupos llamados islotes de Langerhans, porción endocrina del páncreas. Dichas células secretan glucagón, insulina, somatostatina y polipéptido pancreático, hormonas cuyas funciones son tema del capítulo 18.
COMPOSICIÓN Y FUNCIONES DEL JUGO PANCREÁTICO
Cada día, el páncreas produce 1200 a 1500 ml de jugo pancreático, líquido
transparente e incoloro formado en su mayor parte de agua, algunas sales,
bicarbonato de sodio y enzimas. El bicarbonato de sodio le confiere pH
levemente alcalino (7.1 a 8.2), que amortigua el jugo gástrico ácido en el
quimo, detiene la acción de la pepsina y crea el pH apropiado para el efecto
de las enzimas digestivas en el intestino delgado.
Entre las enzimas del jugo pancreático, se incluyen la amilasa pancreática,
que desdobla hidratos de carbono; varias enzimas proteolíticas, a saber,
tripsina, quimotripsina, carboxipeptidasa y elastasa; la lipasa pancreática,
que es la principal enzima en la digestión de triglicéridos en adultos y la
ribonucleasa y desoxirribonucleasa, que se encargan de desdoblar ácidos
nucleicos.
De igual modo que la pepsina gástrica se produce en forma inactiva (pepsinógeno),
lo mismo ocurre con las más proteolíticas del páncreas. Dado su estado
inactivo, no digieren las células pancreáticas mismas. La tripsina se secreta
en la forma inactiva llamada tripsinógeno. Además, las células acinares
pancreáticas secretan el inhibidor tripsina, proteína que se combina con la
tripsina formada accidentalmente en el páncreas o en el jugo pancreático y
bloquean su actividad enzimática. Cuando el tripsinógeno llega a la luz del
intestino delgado, tiene contacto con una enzima de las células en borde en
cepillo, la enterocinasa, que separa una parte de la molécula de tripsinógeno
para formar la tripsina. A su vez, ésta
actúa en los precursores inactivos quimotripsinogeno,
procarboxipeptidasa y proelastasa para producir quimotripsina,
carboxipeptidasa y elastasa,
respectivamente.
APLICACIÓN CLÍNICA
PANCREATITIS
Se denomina pancreatitis a la inflamación del páncreas, como la que ocurre con el abuso del etanol o los cálculos biliares crónicos. En una forma más grave, la pancreatitis aguda, relacionada con la ingesta abundante de etanol o la obstrucción de las vías biliares, las células pancreáticas liberan tripsina en lugar de tripsinógeno o producen cantidades insuficientes del inhibidor de la tripsina, con lo que esta enzima tiene acción digestiva en las propias células pancreáticas. Aunque los pacientes con pancreatitis aguda por lo regular responden al tratamiento, la norma es que haya recurrencias.
Regulación de las secreciones pancreáticas
Las secreciones pancreáticas, al igual que las gástricas, están reguladas por mecanismos neurales y hormonales, como sigue (fig. 24.17):
1. Durante las fases cefálica y gástrica de la digestión gástrica, se transmiten impulsos parasimpáticos al páncreas por el nervio vago (X).
2. Esos impulsos parasimpáticos estimulan el aumento de la secreción de enzimas pancreáticas.
3. El quimo ácido, que contiene proteínas y grasas digeridas parcialmente, entra al intestino delgado.
4. En respuesta a los ácidos grasos y aminoácidos, algunas células enteroendocrinas del intestino delgado secretan colecistocinina (CCC) en la sangre. Ante la presencia del quimo ácido, otras células enteroendocrinas de la mucosa del intestino delgado liberan secretina en la sangre.
5. La secretina estimula el flujo de jugo pancreático, que contiene grandes cantidades de iones bicarbonato.
6. La colecistocinina estimula la secreción del jugo pancreático con alto contenido de enzimas digestivas.
HÍGADO Y VESÍCULA BILIAR
El hígado es la glándula con mayor peso en el cuerpo, cercano a 1.4 Kg. en un adulto de talla promedio, y ocupa el segundo lugar, después de la piel, como órgano más grande. Se localiza en plano inferior al diafragma y ocupa gran parte del hipocondrio derecho y una porción del epigastrio, en la cavidad abdominopélvica (fig. 1.12a).
La vesícula biliar es un saco en forma de pera situado en una depresión de la cara posterior del hígado. Tiene 7 a 10 cm. de longitud y habitualmente cuelga del borde anteroinferior del hígado (fig. 24.16).
Anatomía del hígado y la vesícula biliar
El hígado está cubierto casi completamente por el peritoneo visceral y lo está de manera total por una capa de tejido conectivo denso e irregular situada en plano profundo al peritoneo. El ligamento falciforme del hígado (fig. 24.16) lo divide en dos lóbulos principales: el derecho, grande, y el izquierdo, más pequeño. Aunque muchos anatomistas consideran que el lóbulo derecho comprende los lóbulos cuadrado inferior y caudado posterior, con base en su morfología interna (principalmente, la distribución de vasos sanguíneos) es más apropiado incluirlos en el lóbulo izquierdo. El ligamento falciforme es un repliegue del peritoneo parietal que se extiende desde la cara inferior del diafragma, entre los dos lóbulos principales del hígado, hasta la cara superior de dicha glándula, y ayuda a suspenderlo. En el borde libre del ligamento falciforme, está el ligamento redondo del hígado, cordón fibroso que es un residuo de la vena umbilical fetal (fig. 21.31a, b) y se extiende del hígado al ombligo. Los ligamentos coronarios derecho e izquierdo son repliegues angostos del peritoneo parietal que suspenden el hígado del diafragma.
Las partes de la vesícula biliar son afondo amplio, que se proyecta hacia abajo del borde inferior del hígado; el cuerpo o porción central, y el cuello, que es la parte ahusada. Estas dos últimas se proyectan hacia arriba.
CARACTERÍSTICAS HISTOLÓGICAS DEL HÍGADO Y LA VESÍCULA BILIAR
Los lóbulos del hígado se componen de numerosas unidades funcionales, llamadas lobulillos (fig. 24.18), que consisten en células epiteliales especializadas, los hepatocitos, dispuestas en láminas ramificantes e irregulares conectadas unas con otras, alrededor de una vena central. En lugar de capilares, el hígado posee grandes espacios epiteliales con revestimiento de endotelio, los sinusoides, por los cuales circula la sangre. Además, contienen fagocitos fijos, las células reticuloendoteliales estrelladas (de Kupffer), que se encargan de la destrucción de leucocitos y eritrocitos viejos, bacterias y otros materiales extraños en la sangre venosa que proviene del tubo digestivo.
La bilis, que secretan los hepatocitos, pasa a los canalículos biliares, que son conductos intercelulares angostos que vierten en los conductillos biliares (fig. 24.18a). De ellos, pasa a las vías biliares en la periferia de los lobulillos. Estos conductos se fusionan y en última instancia forman los conductos hepáticos derecho e izquierdo, los cuales se unen y salen del hígado como conducto hepático común (fig. 24.16); más adelante, este último se une al conducto cístico que proviene de la vesícula biliar y forman el conducto colédoco. La bilis pasa al conducto cístico y se almacena temporalmente en la vesícula biliar.
La mucosa de la vesícula biliar consta de epitelio cilíndrico simple, dispuesto en pliegues semejantes a los del estómago. La vesícula carece de submucosa. La capa muscular de su pared se compone de fibras de músculo liso, cuya contracción expulsa el contenido de la vesícula en el conducto cístico. La capa externa de la vesícula biliar es el peritoneo visceral. Las funciones del saco biliar comprenden el almacenamiento y la concentración (hasta del décuplo) de la bilis hasta que es necesaria en el intestino delgado. En el proceso de concentración, se absorben iones y agua en la mucosa vesicular.
VASCULATURA DEL HÍGADO
El hígado recibe sangre de dos fuentes (fig. 24.19). Obtiene sangre oxigenada de la arteria hepática y recibe, de la vena porta hepática, sangre desoxigenada que contiene los nutrientes, fármacos y, posiblemente, microbios y toxinas recién absorbidos del tubo digestivo (fig. 21.29). Las ramas de ambos vasos entran en los sinusoides hepáticos, donde los hepatocitos captan el oxígeno, casi todos los nutrimentos y ciertas sustancias tóxicas. Los compuestos que producen los hepatocitos y los nutrientes necesarios para las células del cuerpo se secretan de nuevo en la sangre, que luego drena en la vena central y finalmente pasa a una vena hepática. La sangre del tubo digestivo pasa a través del hígado como parte de la circulación porta hepática, de modo que esta víscera es sitio frecuente de metástasis con origen en cánceres del tubo digestivo. Las ramas de la vena porta hepática, arteria hepática y vías biliares generalmente se acompañan en su distribución por el hígado. En forma conjunta, a esas tres estruturas se les ha denominado tríada porta (fig. 24.18).
Funciones y composición de la bilis
Cada día, los hepatocitos secretan 800 a 1 000 mL de bilis, la cual es un líquido de color amarillo, parduzco o verde oliva. Su pH es de 7.6 a 8.6 y está compuesto principalmente de agua y ácidos biliares, sales biliares, colesterol, el fosfolípido lecitina, pigmentos biliares y diversos iones.
La bilis es al mismo tiempo una excreción y una secreción digestiva. Las sales biliares, que son sales de sodio y de potasio de los ácidos biliares (en su mayor parte, ácidos cólico y quenodesoxicólico), participan en la emulsión, es decir, el desdoblamiento de grandes glóbulos de lípidos en una suspensión de microgotas de casi 1 u,m de diámetro, así como la absorción de los lípidos después de su digestión. Las diminutas microgotas de lípidos poseen una gran área de superficie, lo cual permite que la lipasa pancreática logre más rápidamente la digestión de los triglicéridos. El colesterol se vuelve soluble en la bilis gracias a las sales biliares y la lecitina.
El principal pigmento biliar es la bilirrubina conjugada. La fagocitosis de los eritrocitos viejos libera hierro, globina y bilirrubina. El hierro y la globina se reciclan, mientras que una parte de la bilirrubina se transforma en conjugada, es decir, unida a moléculas de ácido glucurónico. Luego, la bilirrubina conjugada se secreta en la bilis y se desdobla en el intestino. Uno de los productos de su catabolismo, la estercobilina, confiere su color parduzco normal a las heces.
REGULACIÓN DE LA SECRECIÓN DE BILIS
Luego de desempeñar sus funciones como agentes de emulsión, la mayoría de las sales biliares se reabsorben por transporte activo en la porción final del intestino delgado (íleon) y pasan a la sangre porta que fluye hacia el hígado. Aunque los hepatocitos liberan constantemente la bilis, incrementan la producción y secreción de ésta cuando la sangre porta contiene más ácidos biliares, es decir, aumenta la liberación de bilis mientras continúen la digestión y absorción en el intestino delgado. Entre comidas, después de que ha ocurrido gran parte de la absorción, la bilis fluye hacia la vesícula biliar para su almacenamiento porque el esfínter de Oddi (fig. 24.16) cierra la entrada al duodeno. Tras las comidas, diversos estímulos neurales y hormonales promueven la síntesis y liberación de bilis (fig. 24.20):
1. Los impulsos parasimpáticos que se transmiten por las fibras del nervio vago (X) pueden estimular en el hígado el aumento de la producción de bilis a más del doble del ritmo basal.
2. La presencia de ácidos grasos y aminoácidos en el quimo que llega al duodeno estimula, en ciertas células enteroendocrinas duodenales, la secreción de la hormona colecistocinina (CCC) en la sangre. El quimo ácido que pasa al duodeno también estimula, en otras células enteroendocrinas, la secreción de la hormona secretina en la sangre.
3. La CCC origina la contracción de la pared de la vesícula biliar, lo cual hace que la bilis almacenada pase al conducto cístico y de éste al colédoco. Además, la CCC causa la relajación del esfínter de la ampolla de Valer, lo cual permite el flujo de bilis al duodeno.
4. La secretina, que estimula la secreción de jugo pancreático con alto contenido de HCOJ, también estimula la secreción de la misma sustancia en la bilis por los hepatocitos.
Funciones del hígado
Además de secretar bilis, necesaria para la absorción de las grasas alimenticias, el hígado realiza muchas otras funciones vitales:
• Metabolismo de los hidratos de carbono. El hígado reviste importancia especial en el mantenimiento de la glucemia normal. Cuando este parámetro es bajo, el hígado puede desdoblar el glucógeno en glucosa, que libera en el torrente sanguíneo. Además, esta glándula puede convertir ciertos aminoácidos, ácido láctico y otros azúcares, como la fructosa y galactosa, en glucosa. Si la glucemia es alta, como ocurre justo después de una comida, el hígado convierte la glucosa en glucógeno y triglicéridos para su almacenamiento.
ü Metabolismo de los lípidos. Los hepatocitos almacenan ciertos triglicéridos; desdoblan los ácidos grasos para generar ATP; sintetizan lipoproteínas, que transportan ácidos grasos, triglicéridos y colesterol hacia las células y desde éstas; sintetizan colesterol, y lo usan en la producción de sales biliares.
ü Metabolismo de las proteínas. Los hepatocitos desaminan (separan el grupo amino, NH2) los aminoácidos, de modo que puedan utilizarse para la producción de ATP o convertirse en hidratos de carbono o grasas. Luego, el amoniaco (NH3) tóxico resultante se transforma en urea, mucho menos tóxica, que se excreta en la orina. Además, los hepatocitos sintetizan muchas proteínas plasmáticas, como las globulinas alfa y beta, albúmina, protrombina y fibrinógeno.
ü Procesamiento de fármacos, hormonas y otras sustancias. El hígado puede destoxificar sustancias como el etanol o excretar, en la bilis, fármacos como penicilina, eritromicina y sulfonamidas. Además, modifica químicamente o excreta las hormonas tiroideas y esteroideas, como estrógenos y aldosterona.
ü
Excreción de
bilirrubina. Como se mencionó, la bilirrubina obtenida del grupo hem de
eritrocitos viejos se absorbe en el hígado desde la sangre y se secreta en la
bilis.
Gran parte de la bilirrubina de la bilis se metaboliza en el intestino delgado
por acción de bacterias y se elimina en las heces.
Síntesis de sales biliares. Se utilizan en el intestino delgado para la emulsión y absorción de líquidos, colesterol, fosfolípidos y lipoproteínas
ü Almacenamiento. Además de glucógeno, el hígado es un sitio importante de almacenamiento de ciertas vitaminas (A, B12, D, E y K) y minerales (hierro y cobre), que libera cuando se necesitan en otras partes del cuerpo.
ü Fagocitosis. Las células reticuloendoteliales estrelladas (o de Kupffer) del hígado fagocitan a los eritrocitos y leucocitos viejos, así como a ciertas bacterias.
ü Activación de la vitamina D. Piel, hígado y riñones participan en la síntesis de la forma activa de la vitamina D.
Las funciones del hígado relacionadas con el metabolismo son tema de análisis detallado en el capítulo 25.
APLICACIÓN CLÍNICA CÁLCULOS BILIARES (COLELITIASIS)
Cuando la bilis contiene sales biliares o lecitina insuficientes, o colesterol en exceso, este último tiende a cristalizarse y formar cálculos biliares. Al aumentar de tamaño y número, dichos cálculos pueden originar obstrucción mínima, intermitente o completa del flujo de bilis de la vesícula al duodeno. Su tratamiento consiste en usar agentes que disuelvan los cálculos, litotricia (terapia con ondas de choque) o intervención quirúrgica. En personas con colelitiasis recurrente o en quienes no estén indicados los fármacos o la Litotricia, se requiere la colecistectomía, es decir, la excisión de la vesícula biliar y su contenido. A manera de ejemplo, cada año se practican en Estados Unidos más de 500 000 colecistectomías.
RESUMEN: HORMONAS DIGESTIVA
Los efectos de las tres hormonas digestivas principales (gastrina, secretina y colecistocinina) y los estímulos que promueven su liberación se resumen en el cuadro 24.4. Las células enteroendocrinas de la mucosa gastrointestinal secretan estas tres hormonas en la sangre. La gastrina ejerce sus efectos principales en el estómago, mientras que la secretina y colecistocinina lo hacen en el páncreas, hígado y vesícula biliar.
El estiramiento del estómago cuando recibe los alimentos y el efecto amortiguador de las proteínas alimenticias en los ácidos gástricos desencadenan la liberación de gastrina. A su vez, ésta promueve la secreción de jugo gástrico y aumenta la motilidad del estómago, de modo que los alimentos ingeridos se mezclen adecuadamente y formen el quimo espeso, parecido a una sopa. El reflujo del quimo ácido al esófago se previene gracias a la contracción del cardias, en la cual tiene un efecto estimulante la gastrina.
El estímulo principal para la liberación de la secretina es el quimo ácido (concentración alta de H"1") que entra en el intestino delgado. A su vez, la secretina promueve la secreción de iones bicarbonato (HCOJ) en el jugo pancreático y la bilis (figs. 24.17 y 24.20). Los HCO+" actúan como amortiguadores ("esponjas") del exceso de H+. Además de estos efectos importantes, la secretina inhibe la liberación del jugo gástrico, promueve el crecimiento y mantenimiento normales del páncreas e intensifica los efectos de la CCC. En general, la secretina amortigua el ácido del quimo que llega al duodeno y desacelera la producción de ácido en el estómago.
Los aminoácidos de las proteínas digeridas parcialmente y los ácidos grasos de los triglicéridos digeridos parcialmete estimulan la secreción de colecistocinina de las células enteroendocrinas de la mucosa del intestino delgado. Por su parte, la CCC estimula la secreción de jugo pancreático con contenido alto de enzimas digestivas (fig. 24.17) y el paso de bilis al duodeno (fig. 24.20), desacelera el vaciado gástrico al promover la contracción del esfínter pilórico y produce la sensación de saciedad mediante sus efectos en el hipotálamo encefálico. Al igual que la secretina, la colecistocinina estimula el crecimiento y mantenimiento normales del páncreas, amén de intensificar los efectos de la propia secretina.
Además de esas tres hormonas, se secretan al menos otras 10 hormonas gastrointestinales que tienen efecto en el propio tubo, entre ellas la motilina, sustancia P y bombesina, que estimulan la motilidad intestinal; el polipéptido intestinal vasoactivo, que estimula la secreción de iones y agua por los intestinos e inhibe la secreción de acido gástrico; el péptido liberador de gastrina, que estimula su liberación y la somatostatina, que inhibe la secreción de gastrina. Se piensa que algunas actúan como hormonas locales (paracrinas), mientras que otras se secretan en la sangre o incluso en la luz gastrointestinal. Todavía se investigan las funciones fisiológicas de éstas y otras hormonas gastrointestinales.
INTESTINO DELGADO
Una vez examinadas las fuentes de las diversas enzimas y hormonas digestivas, se procede al análisis de la parte siguiente del tubo digestivo y se reanuda el tema de la digestión y absorción. Los fenómenos principales de estos dos procesos tienen lugar en un largo conducto, el intestino delgado. Casi toda la absorción y digestión de nutrientes ocurre en él, por lo que su estructura está adaptada especialmente para tales funciones. Su gran longitud constituye un área de superficie igualmente grande para la digestión y absorción, área que se incrementa en virtud de los pliegues circulares, vellosidades y microvellosidades. El intestino delgado comienza en el esfínter pilórico gástrico, describe curvas en las porciones central e inferior del abdomen y se continúa con el intestino grueso. En promedio, tiene 2.5 cm. de diámetro y longitud de unos 3 m en personas vivas y de casi 6.5 m en cadáveres, esta última debida a la pérdida del tono del músculo liso tras la muerte.
ANATOMÍA DEL INTESTINO DELGADO
El intestino delgado se divide en tres partes (fig. 24.21). El duodeno, la más corta de ellas, tiene posición retroperitoneal. Se inicia en el esfínter pilórico del estómago y tiene longitud de 25 cm. hasta su unión con el yeyuno. Su nombre significa "doce", porque su longitud equivale a la anchura de 12 dedos. El yeyuno, de casi 1 m de longitud, cuyo nombre significa "vacío", estado que guarda en los cadáveres, se continúa con el íleon. Éste es la parte final y más larga del intestino delgado, de casi 2 m de longitud, y termina en el esfínter ileocecal, donde se une al intestino grueso.
Las proyecciones llamadas pliegues circulares son rebordes permanentes de la mucosa intestinal, de 10 mm de altura (fig. 24.22c). Se inician cerca de la porción proximal del duodeno y terminan en el tercio medio del íleon; algunas se extienden por toda la circunferencia del intestino, y otras, sólo por una parte de ella. Mejoran la absorción al incrementar el área de superficie y hacer que el quimo describa un trayecto en espiral, no recto, a su paso por el intestino delgado.
CARACTERÍSTICAS HISTOLÓGICAS DEL INTESTINO DELGADO
La pared del intestino delgado se compone de las mismas cuatro capas que la mayor parte del resto del tubo digestivo; pero ciertas características especiales de la mucosa y submucosa facilitan la digestión y absorción en él. La mucosa forma una serie de vellosidades digitiformes, las cuales son proyecciones de 0.5 a I mm de altura (fig. 24.22a). Su gran número, de 20 a 40/mm2, incrementa considerablemente el área de superficie epitelial disponible para la absorción y digestión, además de conferir a la mucosa intestinal un aspecto aterciopelado. Cada vellosidad posee un centro de lámina propia (tejido conectivo areolar), que incluye arteriola, vénula, red de capilares sanguíneos y vaso quilífero, que es un capilar linfático. Los nutrientes que absorben las células epiteliales de recubrimiento de las vellosidades cruzan la pared del capilar sanguíneo o el vaso quilífero para llegar a la sangre o linfa, respectivamente.
El epitelio de la mucosa es de tipo cilíndrico sencillo e incluye células de absorción, caliciformes, enteroendocrinas y de Paneth (fig. 24.22b). La membrana apical (libre) de las células de absorción tiene micro vellosidades, cada una de las cuales es una proyección cilíndrica de 1 um de longitud con recubrimiento de membrana y unos 20 a 30 filamentos de actina. En una microfotografía obtenida con un microscopio de luz, las microvellosidades son demasiado pequeñas para verlas una por una, en vez de lo cual forman una línea difusa, llamada borde en cepillo, que se extiende en la luz del intestino delgado (fig. 24.23b). Se calcula en 200 millones el número de microvellosidades por milímetro cuadrado de intestino delgado. Puesto que dichas estructuras incrementan considerablemente el área de superficie de la membrana plasmática, grandes cantidades de nutrientes digeridos se pueden difundir a las células de absorción en un periodo dado. El borde en cepillo también contiene diversas enzimas con funciones digestivas, que se analizan poco más adelante. La mucosa tiene muchas depresiones profundas llenas de epitelio glandular. Las células de revestimiento de tales depresiones forman las criptas de Lieberkühn, que secretan el jugo intestinal. Muchas células epiteliales de la mucosa son caliciformes, secretoras de moco. Las células de Paneth, situadas en la parte más profunda de las criptas, secretan lisozima, que es una enzima bactericida, además de que pueden realizar la fagocitosis. Desempeñan una función clave en la regulación de la población microbiana intestinal. Tres tipos de células enteroendocrinas, también de la parte más profunda de las criptas, secretan hormonas: las células S, secretina; las CCC, colecistocinina, y las K, péptido insulinotrópico dependiente de la glucosa. La lámina propia del intestino delgado tiene abundante tejido linfoide relacionado con mucosa. Los folículos linfáticos solitarios son más numerosos en la porión distal del íleon; los grupos de estos folículos, llamados placas de Peyer, son también numerosos en el íleon. La muscular de la mucosa consta de músculo liso. La submucosa duodenal contiene las glándulas de Brunner (fig. 24.23a), las cuales secretan un moco alcalino que ayuda a neutralizar el ácido gástrico del quimo.La muscular del intestino delgado está formada por dos capas de músculo liso. La externa, más delgada, tiene fibras longitudinales, y la interna, más gruesa, fibras circulares. Excepto una porción importante del duodeno, la serosa (peritoneo visceral), rodea por completo el intestino delgado.
Funciones del jugo intestinal y de las enzimas del borde en cepillo
El jugo intestinal es un líquido amarillo transparente, del cual se secretan 1 a 2 L diarios. Contiene agua y moco y es ligeramente alcalino (pH 7.6). En forma conjunta, los jugos pancreático e intestinal constituyen un medio líquido que facilita la absorción de sustancias del quimo a su contacto con las microvellosidades. Las células epiteliales de absorción sintetizan diversas enzimas digestivas, las enzimas del borde en cepillo, y las insertan en la membrana plasmática de las microvellosidades. De tal suerte, ocurre cierta digestión enzimática en la superficie de las células epiteliales de revestimiento de las vellosidades, no sólo en la luz intestinal, al igual que en otras partes del tubo digestivo. Entre las enzimas en cuestión, se cuentan la a-dextrinasa, maltasa, sacarasa y lactasa, que digieren carbohidratos; las peptidasas (aminopeptidasa y dipeptidasa), que son proteolíticas, y las nucleosidasas y fosfatasas, que son dos tipos de enzimas de digestión de nucleótidos. Además, a medida que se esfacelan células en la luz del intestino delgado, se desintegran y liberan enzimas que ayudan a la digestión de nutrientes del quimo.
Dos tipos de movimientos del intestino delgado, las segmentaciones y el tipo de peristaltismo llamado complejos de motilidad migratoria, están principalmente bajo regulación del plexo mientérico. Las segmentaciones son un tipo localizado de contracción de mezclado que ocurre en porciones intestinales distendidas por un gran volumen de quimo. Mezclan éste con los jugos digestivos y hacen que las partículas de alimentos entren en contacto con la mucosa, para su absorción, sin desplazar el contenido intestinal en sentido distal. Una segmentación se inicia con las contracciones de las fibras musculares circulares en una porción del intestino delgado, acción que constriñe el órgano en segmentos. Luego, también se contraen las fibras musculares que rodean la porción central de cada segmento, que se divide una vez más. Por último, se relajan las fibras que se contrajeron en primer término y cada segmento pequeño se une con el adyacente, de modo que se forman de nuevo los segmentos grandes. Al repetirse esta sucesión de fenómenos, el quimo se desplaza hacia delante y hacia atrás. Las segmentaciones ocurren más rápidamente en el duodeno, unas 12 veces por minuto, y disminuyen gradualmente hasta ocho por minuto en el íleon. Son similares a comprimir de manera alternada la porción central y luego los extremos de un tubo de pasta de dientes tapado.
Después de absorbida la mayor parte de una comida, lo cual reduce la distensión de la pared del intestino delgado, se detienen las segmentaciones y se inicia el peristaltismo. El tipo de éste que ocurre en el intestino delgado, los llamados complejos de motilidad migratoria, se inicia en la porción inferior del estómago y desplaza el quimo en dirección anterógrada un breve trecho del intestino delgado, antes de desaparecer. Estos complejos emigran lentamente por el intestino delgado en sentido distal, hasta alcanzar el extremo del íleon en 90 a 120 min. Luego, se inicia otro complejo de este tipo en el estómago. En total, el quimo permanece en el intestino delgado de tres a cinco horas.
DIGESTIÓN QUÍMICA EN EL INTESTINO DELGADO
En la boca, la amilasa salival convierte el almidón (polisacárido) en maltosa (disacárido), maltotriosa (trisacárido) y adextrinas (fragmentos de cadena corta ramificados de almidón, de cinco a 10 unidades de glucosa). En el estómago, la pepsina convierte las proteínas en péptidos (fragmentos pequeños de proteínas), y las lipasas lingual y gástrica transforman algunos triglicéridos en ácidos grasos, diglicéridos y monoglicéridos. Así pues, el quimo que llega al intestino delgado contiene hidratos de carbono, proteínas y lípidos digeridos parcialmente. La digestión de estas sustancias se completa gracias a la acción conjunta del jugo pancreático, bilis y jugo intestinal en el intestino delgado.
DIGESTIÓN DE HIDRATOS DE CARBONO
Aunque la acción de la amilasa salival suele continuar algún tiempo en el estómago, el pH gástrico ácido la destruye e interrumpe su actividad. Así pues, sólo algunos almidones están reducidos a maltosa cuando el quimo sale del estómago. Los que no están desdoblados en maltosa, maltotriosa y adextrinas se descomponen por acción de la aniuasa pancreática, enzima del jugo pancreático que actúa en el intestino delgado. Aunque la amilasa ejerce sus efectos en el glucógeno y almidones, no actúa en otro polisacárido, la celulosa, que es una fibra vegetal no digerible. Después de que la amilasa (salival o pancreática) ha desdoblado el almidón en fragmentos más pequeños, la enzima del borde en cepillo adextrinasa se encarga de la catabolia de las adextrinas resultantes, separando las unidades de glucosa una por una.
Las moléculas de sacarosa, lactosa y maltosa (que son tres disacáridos) ingeridas no son objeto de catabolia hasta que llegan al intestino delgado. Tres enzimas del borde en cepillo digieren estos disacáridos en monosacáridos: la sacarasa desdobla la sacarosa en una molécula de glucosa y otra de fructosa; la lactasa metaboliza la lactosa en una molécula de glucosa y otra de galactosa, y la maltasa digiere la maltosa y maltotriosa en dos y tres moléculas de glucosa, respectivamente. La digestión de los carbohidratos termina al producirse monosacáridos, ya que hay mecanismos para su absorción.
APLICACIÓN CLÍNICA
INTOLERANCIA A LA LACTOSA
En algunas personas, las células de la mucosa del intestino delgado no producen lactasa suficiente para la digestión de la lactosa, lo cual ocasiona la intolerancia a la lactosa, en que la lactosa no digerida del quimo retiene líquido en las heces y su fermentación bacteriana origina la producción de flatos. Los síntomas de la intolerancia a la lactosa incluyen diarrea, flatulencia y distensión y cólicos abdominales después de la ingestión de leche y productos lácteos. Los síntomas varían desde relativamente leves hasta de gravedad suficiente para justificar la atención médica. Las personas con este problema pueden tomar complementos dietéticos que faciliten la digestión de este hidrato de carbono.
DIGESTIÓN DE PROTEÍNAS
La digestión de las proteínas se inicia en el estómago, donde se fragmentan en péptidos por acción de la pepsina. Las enzimas del jugo pancreático (tripsina, quimiotrípsina, carboxipeptidasa y elastasa) continúan su desdoblamiento. Aunque estas enzimas convierten proteínas enteras en péptidos, sus acciones difieren hasta cierto punto porque cada una deshace los enlaces peptídicos entre aminoácidos distintos. La tripsina, quimotripsina y elastasa destruyen el enlace peptídico entre un aminoácido específico y el adyacente, y la carboxipeptidasa, el enlace peptídico que une el aminoácido terminal con el extremo carboxilo (ácido) del péptido. La digestión de las proteínas se completa por el efecto de dos peptídasas del borde en cepillo, la aminopeptidasa y dipeptidasa. La aminopeptidasa actúa en los péptidos al romper el enlace peptídico que une el aminoácido terminal con el extremo amino del péptido, mientras que la dipeptidasa separa los dipéptidos (dos aminoácidos unidos por un enlace peptídico) en los aminoácidos componentes.
DIGESTIÓN DE LÍPIDOS
Los lípidos más abundantes en los alimentos son los triglicéridos, que constan de una molécula de glicerol unida a tres de ácidos grasos (fig. 2.17). Las enzimas que desdoblan los triglicéridos y fosfolípidos reciben el nombre de lipasas. En adultos, gran parte de la digestión de los lípidos ocurre en el intestino delgado, si bien una pequeña porción tiene lugar en el estómago por acción de las lipasas lingual y gástrica. Cuando el quimo entra en el intestino delgado, las sales biliares emulsionan los glóbulos de triglicéridos en microgotas de casi 1 un micra de diámetro, lo cual aumenta el área de superficie expuesta a la lipasa pancreática, que es otra enzima del jugo pancreático. Esta sustancia hidroliza los triglicéridos en ácidos grasos y monoglicéridos, que son los productos terminales principales de la digestión de los triglicéridos. Las lipasas pancreática y gástrica separan dos de los tres ácidos grasos del glicerol, mientras que el otro permanece unido a él y se forma un monoglicérido.
DIGESTIÓN DE ÁCIDOS NUCLEICOS
El jugo pancreático contiene dos nucleasas, la ribonucleasa, que digiere al RNA, y la desoxirribonudeasa, que desdobla al DNA. Los nucleótidos resultantes de la acción de estas dos nucleasas son objeto de digestión adicional por enzimas del borde en cepillo, las nucleosidasas y fosfatasas, en pentosas, grupos fosfato y bases nitrogenadas. Estos productos se absorben mediante transporte activo. Las fuentes, sustratos y productos de las enzimas digestivas se resumen en el cuadro 24.5
REGULACIÓN DE LAS SECRECIONES Y MOTILIDAD INTESTINALES
Los mecanismos reguladores más importantes de las secreciones y motilidad del intestino delgado son reflejos entéricos que responden a la presencia del quimo, además de que el péptido intestinal vasoactivo (PIV) estimula la producción del jugo intestinal. Los movimientos de segmentación dependen principalmente de la distensión del intestino, lo cual genera impulsos nerviosos que se transmiten a los plexos entéricos y al sistema nervioso central. Los reflejos entéricos y los impulsos parasimpáticos eferentes del SNC aumentan la motilidad, mientras que los simpáticos la disminuyen. Los complejos de motilidad migratorios son más intensos cuando se ha absorbido gran parte del agua y nutrientes, es decir, cuando las paredes del intestino delgado están menos distendidas. Con el peristaltísmo más intenso, el quimo se desplaza hacia el intestino grueso con velocidad de 10 cm./s. Los primeros residuos de una comida llegan al intestino grueso luego de cuatro horas
ABSORCIÓN EN EL INTESTINO DELGADO
Todas las fases mecánicas y químicas de la digestión, desde la boca hasta el intestino delgado, están encaminadas a convertir los alimentos en formas que puedan cruzar las células epiteliales de revestimiento de la mucosa hacia los vasos sanguíneos y linfáticos subyacentes. Estas formas son los monosacáridos (glucosa, fructosa y galactosa), en el caso de los hidratos de carbono; los aminoácidos, dipéptidos y tripéptidos, en el de las proteínas, y los ácidos grasos, glicerol y monoglicéridos, en el de los triglicéridos. Se llama absorción al paso de estos nutrientes digeridos desde el tubo digestivo hacia la sangre o linfa.
La absorción de los nutrimentos ocurre por difusión, difusión facilitada, osmosis y transporte activo. Casi 90% de la absorción de nutrientes tiene lugar en el intestino delgado, y el otro 10%, en el estómago e intestino grueso. Los materiales no digeridos o no absorbidos que permanezcan en el intestino delgado pasan al grueso.
ABSORCIÓN DE MONOSACÁRIDOS
Todos los hidratos de carbono se absorben en la forma de monosacáridos. La capacidad del intestino delgado para su absorción es considerable, de unos 120 g/h. En consecuencia, lo normal es que se absorban todos los hidratos de carbono digeridos de los alimentos, de modo que en las heces sólo queden las fibras y celulosa no digeribles. Los monosacáridos se absorben desde la luz. intestinal, a través de la membrana apical, por difusión facilitada o transporte activo. La fructosa, monosacárido de las frutas, se transporta por difusión facilitada, mientras que la glucosa y galactosa pasan a las células epiteliales de las vellosidades por transporte activo secundario, el cual está acoplado al transporte activo de los Na+ (fig. 24.24a). La molécula transportadora tiene sitios receptores para una molécula de glucosa y dos iones sodio; si no están ocupados los tres sitios, no se transporta ninguna de las dos sustancias. La galactosa compite con la glucosa por la misma molécula transportadora. (Los Na+ y la glucosa o galactosa se mueven en la misma dirección, de modo que se trata de un cotransporte o transporte asociado. El mismo tipo de cotransporte de Na+-glucosa reabsorbe la glucosa sanguínea filtrada en los túbulos renales [fig. 26.12].) Luego, los monosacáridos salen de las células epiteliales por la superficie basolateral mediante difusión facilitada y entran en los capilares de las vellosidades (fig. 24.24a, b).
ABSORCIÓN DE AMINOÁCIDOS, DIPÉPTIDOS Y TRIPÉPTIDOS
La mayoría de las proteínas se absorben en la forma de aminoácidos mediante procesos de transporte activo que ocurren principalmente en el duodeno y yeyuno. Casi la mitad de los aminoácidos absorbidos es parte de los alimentos, mientras que el resto corresponde a proteínas de los jugos digestivos y células muertas que se esfacelan de la mucosa. En condiciones normales, se digiere y absorbe 95 a 98% de las proteínas que están en el intestino delgado. Varios transportadores participan en la absorción de distintos tipos de aminoácidos. Algunos entran en las células epiteliales de las vellosidades gracias a procesos de transporte activo secundario dependiente de los Na+, similares a los de la glucosa, mientras que otros se transportan activamente a sí mismos. Al menos un cotransporte lleva a los dipéptidos y tripéptidos con los H+, después de lo cual se hidrolizan los péptidos en aminoácidos, en el interior de las células epiteliales. Los aminoácidos salen de dichas células por difusión y pasan a los capilares de las vellosidades (fig. 24.24a, b). Tanto los monosacáridos como los aminoácidos se transportan al hígado en la sangre del sistema porta hepático. Si los hepatocitos no los extraen de la sangre, pasan a la circulación general.
ABSORCIÓN DE LÍPIDOS
Tocios los lípidos de la alimentación se absorben por difusión sencilla. En adultos, ocurre la absorción de casi 95% de los lípidos que hay en el intestino delgado, mientras que los neonatos absorben tan sólo 85%, lo cual se debe a su baja producción de bilis. Como resultado de su emulsión y digestión, los triglicéridos se separan en monoglicéridos y ácidos grasos. Recuérdese que las lipasas lingual y pancreática separan dos de los tres ácidos grasos del glicerol durante la digestión de un triglicérido, mientras que el otro permanece unido al glicerol y forma un monoglicérido. La reducida proporción de ácidos grasos de cadena corta (con menos de 10 a 12 átomos de carbono) de los alimentos pasa a las células epiteliales por difusión sencilla y sigue el mismo trayecto que los monosacáridos y aminoácidos hacia los capilares sanguíneos de las vellosidades (fig. 24.24a, b).
Sin embargo, la mayoría de los ácidos grasos de los alimentos son de cadena larga. Junto con los monoglicéridos, llegan a la sangre en virtud de un proceso distinto y requieren la bilis para su absorción satisfactoria. Las sales biliares son anfipáticas, es decir, tienen porciones polar (hidrófila) y no polar (hidrófoba). De tal suerte, pueden formar diminutas esferas, las micelas, de 2 a 10 nm de diámetro, que contienen 20 a 50 moléculas de sales biliares. A causa de su pequeño tamaño y de la porción polar de las moléculas de sales biliares en su superficie, las micelas pueden disolverse en el agua del líquido intestinal. En contraste, los lípidos de los alimentos digeridos parcialmente pueden disolverse en el centro no polar de las micelas. De esta manera, los ácidos grasos y monoglicéridos llegan a las células epiteliales de las vellosidades.
En la superficie apical de las células epiteliales, los ácidos grasos y monoglicéridos se difunden hacia dichas células, mientras que las micelas permanecen en el quimo. Luego, , continúan esta función de transporte. Cuando el quimo llega al íleon, se ha reabsorbido 90 a 95% de las sales biliares, que regresa al hígado por el sistema porta hepático para su reciclaje. Se denomina circulación enterohepática a este ciclo de secreción de sales biliares por los hepatocitos en la bilis, su reabsorción en el íleon y su ulterior secreción en la bilis. El volumen insuficiente de sales biliares, ya sea por obstrucción de las vías biliares o por la excisión de la vesícula biliar, puede hacer que se pierda en las heces hasta 40% de los lípidos de los alimentos como resultado de su absorción disminuida. Por añadidura, cuando los lípidos no se absorben correctamente, ocurre lo mismo con las vitaminas liposolubles (A, D.EyK).
En las células epiteliales, numerosos monoglicéridos son objeto de digestión adicional por la lipasa, en glicerol y ácidos grasos. Luego, estas sustancias se recombinan para formar los triglicéridos, se agregan en glóbulos con los fosfolípidos y colesterol, y después quedan recubiertos con proteínas. Estas masas esféricas grandes, de unos 80 mm de diámetro, se llaman quilomicrones. La cubierta hidrófila de proteínas las mantiene suspendidas e impide que se adhieran unas con otras. Los quilomicrones salen de las células epiteliales por exocitosis. A causa de su gran tamaño, no pueden entrar en los capilares sanguíneos del intestino delgado, en vez de lo cual pasan a los vasos quilíferos, mucho más permeables. Desde tales vasos, se transportan por el sistema linfático al conducto torácico y se incorporan a la sangre en la vena subclavia izquierda (fig. 24.24b).
En los 10 minutos que siguen a su absorción, casi la mitad de los quilomicrones ha sido extraída de la sangre a su paso por los capilares sanguíneos del hígado y tejido adiposo. Ello se logra gracias a una enzima de las células endoteliales de los capilares, la lipoproteína lipasa, que desdobla los triglicéridos de los quilomicrones y otras lipoproteínas en ácidos grasos y glicerol. Los ácidos grasos se difunden a los hepatocitos y las células adiposas, donde se combinan con el glicerol durante la síntesis de triglicéridos. Luego de dos o tres horas de una comida, son pocos los quilomicrones que permanecen en la sangre.
Absorción de electrólitos
Muchos de los electrólitos que se absorben en el intestino delgado provienen de las secreciones gastrointestinales, y el resto, de los líquidos y alimentos ingeridos. Los iones sodio salen de las células epiteliales intestinales por transporte activo de bombas de sodio (ATPasa de Na+/K+) después de entrar en dichas células por difusión y transporte activo secundario. Así pues, gran parte de los iones sodio de las secreciones gastrointestinales se reciclan, en vez de perderse en las heces. Los iones cloruro, yoduro y nitrato, de carga negativa, pueden seguir pasivamente a los Na+ o ser objeto de transporte activo. Los iones calcio se absorben de manera activa, en un proceso que estimula el calcitriol. Otros electrólitos, como el hierro, potasio, magnesio y fosfato, también se absorben por mecanismos de transporte activo.
ABSORCIÓN DE VITAMINAS
Las vitaminas liposolubles A, D, E y K se incluyen con los lípidos de los alimentos en las micelas y se absorben por difusión sencilla. La mayoría de las vitaminas hidrosolubles, entre ellas casi todas las del complejo B y la vitamina C, también se absorben por difusión sencilla. Sin embargo, la vitamina B12 se combina con el factor intrínseco, que se produce en el estómago, y se absorbe en el íleon por un mecanismo de transporte activo.
ABSORCIÓN DE AGUA
El volumen total de líquido que llega al intestino delgado todos los días, de unos 9.3 L, proviene de la ingestión de líquidos (unos 2.3 L) y de las diversas secreciones gastrointestinales (cerca de 7 L). En la figura 24.25, se muestran los volúmenes de líquidos ingeridos, secretados, absorbidos y excretados por el tubo digestivo. El intestino delgado absorbe casi 8.3 L, mientras que el resto pasa al intestino grueso, donde también se absorbe la mayor parte (0.9 L), y apenas 0.1 L de agua se excreta diariamente en las heces.
La absorción del agua en el tubo digestivo ocurre por osmosis desde la luz intestinal hacia los capilares sanguíneos, a través de las células epiteliales. El agua puede cruzar la mucosa intestinal en ambas direcciones, de modo que su absorción en el intestino delgado depende de la correspondiente a electrólitos y nutrientes, para mantener el equilibrio osmótico con la sangre. Los electrólitos, monosacáridos y aminoácidos absorbidos producen un gradiente de concentración para el agua que fomenta la absorción de ésta por osmosis.
Las actividades digestivas de páncreas, hígado, vesícula biliar e intestino delgado se resumen en el cuadro 24.6.
INTESTINO GRUESO
El intestino grueso es la porción Terminal del tubo digestivo y se divide en cuatro partes principales. En general, sus funciones son completar la absorción, producir ciertas vitaminas, formar las heces y expulsarlas del cuerpo.
ANATOMÍA DEL INTESTINO GRUESO
El intestino grueso, de unos 1.5 m de longitud y 6.5 cm. de diámetro, se extiende entre el íleon y el ano, que se une a la pared abdominal posterior por el mesocolon, que es una doble capa de peritoneo. Desde el punto de vista estructural, sus cuatro porciones principales son ciego, colon, recto y conducto anal (fig. 24.26a).
La comunicación del íleon con el intestino grueso está protegida por un pliegue de mucosa, el esfínter ileocecal, que permite el paso de materiales del intestino delgado al grueso. En plano inferior a dicha válvula, cuelga el ciego, saco de unos 6 cm. de longitud que ésta cerrado en su extremo distal. El apéndice, tubo contorneado de unos 8 cm. de longitud, esta unido al ciego. El mesenterio del apéndice, llamado mesoapéndice, lo fija en la parte inferior del mesenterio ileal. El extremo abierto del ciego guarda continuidad con un largo conducto, el colon, que se divide en porciones ascendente, transversa, descendente y sigmoide. Las partes ascendente y descendente son retroperitoneales, no así la transversa y sigmoide. El colon ascendente sigue trayecto superior en el lado derecho del abdomen, llega a la cara interior del hígado y describe un giro abrupto a la izquierda, lo cual forma el ángulo hepático. Luego, continúa hacia el otro lado del abdomen con el nombre de colon transverso. Presenta una curva bajo el extremo inferior del bazo, en el lado izquierdo del abdomen, donde forma el ángulo esplénico, para luego seguir hacia abajo, hasta la altura de la cresta iliaca, como colon descendente. El colon sigmoide se inicia cerca de la cresta iliaca izquierda, se proyecta hacia la línea media y termina en el recto, poco más o menos a la altura de la vértebra S3.
El recto, que conforma los últimos 20 cm. del tubo digestivo, está por delante del sacro y cóccix. Sus 2 a 3 cm. terminales se denominan conducto anal (fig. 24.26b), La mucosa de éste tiene pliegues longitudinales, llamados columnas anales, que contienen una red de arterias y venas. La apertura del conducto al exterior, el ano, presenta un esfínter interno de músculo liso (involuntario) v otro externo de músculo esquelético (voluntario.). En condiciones normales, el ano está cerrado, salvo durare la eliminación de las heces.
APLICACIÓN CLÍNICA APENDICITIS
La inflamación del apéndice o apendicitis va precedida de la obstrucción de la luz apendicular por el quimo, inflamación, cuerpo extraño, carcinoma cecal, estenosis o acodadura del propio apéndice. Se caracteriza por fiebre alta y cuenta leucocitaria elevada, con recuento de neutrófilos mayor de 75%. La infección subsiguiente puede causar edema e esquemia, que progresan a la gangrena y perforación en 24 a 36 ñoras. Por lo regular, la apendicitis comienza con dolor irradiado a la región umbilical del abdomen, seguido de anorexia (inapetencia), náusea y vómito. Luego de varias horas, el dolor se tocaza en el cuadrante inferior derecho y es continuo, sordo y se agrava con tos, estornudos o movimientos corporales. Se recomienda realizar apendicetomía.(excision del apéndice) a la brevedad posible, en vez de arriesgarse a la rotura apendicular, peritonitis y gangrena.
CARACTERÍSTICAS HISTOLÓGICAS DEL INTESTINO GRUESO
La pared del intestino grueso difiere de la correspondiente al intestino delgado en varios aspectos. En primer término, la mocosa, que consta de epitelio cilíndrico simple, lámina propia (tejido conectivo areolar) y muscular de la mucosa(músculo liso), está desprovista de vellosidades y pliegues circulares permanentes (fig. 24.27a). El epitelio contiene principalmente células de absorción y caliciformes (fig. 24.27b, c). Las células de absorción se encargan ante todo de absorber agua, y las caliciformes, de secretar moco que lubrica el tránsito del contenido de colon. Ambos tipos celulares se localizan en glándulas intestinales tubulares rectas y largas, que abarcan todo e! espesor de la mucosa. Esta también posee folículos linfáticos solitarios. La submucosa del intestino grueso es como la del resto del tubo digestivo. La muscular está formada por una cara externa de músculo liso longitudinal y otra interna y circular, también de músculo liso.
A diferencia de lo que ocurre en otras partes del aparato digestivo, ciertas porciones de los músculos longitudinales están engrosadas, lo cual forma tres bandas longitudinales prominentes, llamadas tenias del colon, que se encuentran en gran parte del intestino grueso. Estas se hallan separadas por porciones de la pared con poco músculo liso longitudinal, o sin el. La contracción tónica de las tenias hace que el colon se convierta en una sucesión de bolsas, llamadas haustras, que le confieren su aspecto abollonado característico. Una sola capa de músculo liso circular está dispuesta entre las tenias del colon. La serosa del intestino grueso es parte del peritoneo visceral. Los apéndices epiploicos son pequeños sacos de peritoneo visceral llenos de grasa y unidos a las tenias del colon.
DIGESTIÓN MECÁNICA EN EL INTESTINO GRUESO
El paso del quimo del ileon al ciego esta regulado por el esfínter ileocecal. En condiciones normales, esta válvula permanece cerrada parcialmente, de modo que el quimo suele cruzarla con lentitud. Después de las comidas, el reflejo gastroileal intensifica el peristaltismo del íleon y fuerza el paso del quimo al ciego. La gastrina también relaja el esfínter ileocecal. Cuando el ciego esta distendido, se intensifica la contracción del esfínter mencionado.
Los movimientos del colon se inician cuando el quimo leocecal. El quimo se mueve en el intestino delgado con velocidad relativamente constante, de modo que el tiempo necesario para qué pase al colon depende del tiempo cié vaciado gástrico. Una vez que cruza el esfínter ileocecal, se acumula en el colon ascendente.
Un movimiento característico del intestino grueso es la propulsión haustral, en que las haustras permanecen relajadas y se distienden al llenarse. Cuando la distensión alcanza cierto punto, las paredes se contraen y desplazan el contenido a la haustra siguiente. También ocurre peristaltismo, si bien con ritmo más lento (tres a 12 contracciones por minuto) que en porciones más proximales del tubo digestivo. Un tercer tipo de movimiento es el peristaltismo masivo, potente onda peristáltica que comienza en la porción media del colon transverso y desplaza rápidamente el contenido del colon al recto. La presencia de alimentos en el estómago inicia este reflejo gastrocólico en el colon, de modo que hay peristaltismo masivo por lo regular tres o cuatro veces al día, durante las comidas o inmediatamente después de ellas.
DIGESTIÓN QUÍMICA EN EL INTESTINO GRUESO
La etapa final de la digestión ocurre en el colon por la actividad de las bacterias presentes en su luz. Las glándulas del intestino grueso secretan moco, no así enzimas. El quimo se prepara para la defecación mediante la acción de bacterias, que fermentan los hidratos de carbono residuales y liberal, hidrógeno, dióxido de carbono y metano. Estos gases contribuyen a que haya flatos en el colon, que se denominan flatulencia cuando resultan excesivas. Además, las bacterias convierten las proteínas residuales en aminoácidos, los cuales desdoblan en sustancias más sencillas: indol, escatol, sulfuro de hidrógeno y ácidos grasos. Una parte de los dos primeros se elimina en las heces y contribuye al olor caracteristico de estas, mientras que el resto se absorbe y transporta al hígado donde tales compuestos se convierten en otros menos tóxicos, y se excretan en la orina. Por añadidura, las bacterias descomponen la bilirrubina en pigmentos más sencillos, corno la estercobilina, que confiere su color pardo característico a las heces. Se requieren diversas vitaminas para el metabolismo normal, entre ellas algunas del complejo B y la K, las cuales son productos bacterianos que se absorben en el colon.
ABSORCIÓN Y FORMACIÓN DE HECES EN EL INTESTINO GRUESO
Cuando el quimo ha permanecido en el intestino grueso 3 a 10 h, adquiere consistencia sólida o semisólida como resultado de la absorción de agua y su nombre cambia a heces, las cuales químicamente están compuestas de agua, sales inorgánicas, células epiteliales esfaceladas de la mucosa gastrointestinal, bacterias, productos de la descomposición bacteriana, materiales digeridos y no absorbidos y partes no digeribles de los alimentos.
Aunque el intestino delgado absorbe gran parte del agua, el grueso lo hace en volumen suficiente para ser un órgano de importancia en la conservación del equilibrio hídrico del cuerpo. De los 0.5 a 1 L que entran al intestino grueso, solo o.11 a 0.2 l no se absorben por osmosis. Además el intestino grueso absorbe electrolitos, incluidos el sodio y cloruro, y algunas vitaminas
REFLEJO DE DEFECACIÓN
El peristaltismo masivo empuja las heces del colon sigmoide al recto. La distensión resultante de la pared rectal estimula receptores de estiramiento, lo cual inicia un reflejo de defecación, con el que se vacía el recto. Este fenómeno ocurre como sigue. En respuesta a la distensión de la pared rectal, los receptores envían impulsos nerviosos a la porción sacra de la médula espinal. Desde ésta, viajan impulsos motores por las fibras parasimpáticas al colon descendente y sigmoide, recto y ano. La contracción subsiguiente del músculo liso longitudinal del recto lo acorta, con lo que aumenta la presión en este último. Ello, aunado a la contracción voluntaria del diafragma y de los músculos abdominales, así como a la estimulación parasimpática, abre el esfínter interno del ano.
El esfínter externo del ano está bajo regulación voluntaria. Al relajarlo, ocurre la defecación y se expulsan las heces por el ano, mientras que su contracción voluntaria permite postergar la evacuación. La contracción también voluntaria del diafragma y los músculos abdominales facilita la defecación al incrementar la presión intraabdominal, lo cual empuja la pared del colon sigmoide y recto hacia la luz de éstos. En caso de no ocurrir la defecación, las heces regresan al colon sigmoide hasta que la siguiente onda de peristaltismo masivo estimule a los receptores de estiramiento y genere nuevamente la necesidad de defecar. En lactantes, dicho reflejo causa el vaciado automático del recto porque todavía no se ha desarrollado la regulación voluntaria del esfínter externo del ano.
La diarrea es el aumento de la frecuencia, el volumen y contenido de líquido de las heces como resultado de mayor motilidad y menor absorción intestinales. Cuando el quimo transita muy rápidamente por el intestino delgado y las heces lo hacen de igual manera por el grueso, no se dispone de tiempo suficiente para la absorción. La diarrea frecuente puede causar deshidratación y desequilibrios electrolíticos. La motilidad excesiva puede ser consecuencia de intolerancia a la lactosa, estrés o microbios que irriten la mucosa gastrointestinal.
El estreñimiento es la defecación infrecuente o difícil como resultado de motilidad intestinal disminuida. Las heces permanecen en el colon durante largo tiempo, lo cual vuelve excesiva la absorción de agua y hace que las heces se resequen y endurezcan. Este problema puede ser resultado de hábitos inadecuados (postergar la defecación), espasmo del colon, fibra insuficiente en los alimentos, ingesta deficiente de líquidos, falta de ejercicio, estrés emocional y ciertos medicamentos. Por lo regular se contrarresta con laxantes leves, como leche de magnesio, que induce la defecación. Sin embargo, muchos médicos consideran que los laxantes forman hábito y que aumentar la fibra en la dieta, el ejercicio y la ingesta de líquidos son formas más seguras de controlar este problema común.
Las actividades digestivas del intestino grueso se resumen en el cuadro 24.7.
APLICACIÓN CLÍNICA FIBRA ALIMENTICIA
Fibra alimenticia consta de hidratos de carbono no digeribles de origen vegetal, como celulosa, lignina y pectina, de las frutas, verduras, gramíneas y frijoles. La fbra indisoluble, es decir, la que no se disuelve en agua, comprende las partes leñosas o estructurales de las plantas, como la cáscara de frutas y verduras o el salvado de los granos de trigo y maíz. Esta fibra pasa por el tubo digestivo sin sufrir grandes cambios y acelera el tránsito gastrointestinal. La fibra soluble se disuelve en agua y forma un gel, lo cual desacelera el tránsito gastrointestinal; es abundante en frijoles, avena, centeno, brócoli, ciruela pasa, manzanas y cítricos. Las personas que optan por una dieta de alto contenido de fibra suelen disminuir el riesgo de obesidad, diabetes, ateroesclerosis, calculos biliares, hemorroides, diverticulistis, apendicitis y cancer colorrectal. Además, la fibra soluble ayuda a reducir la colesterolemia, dado que se une a las sales biliares y previene su absorción, con lo cual se utiliza más colesterol para reponer las sales biliares perdidas en las heces.
ANATOMÍA DEL DESARROLLO DEL APARATO DIGESTIVO
Hacia el decimocuarto día después de la fecundación, las células del endodermo forman una cavidad, el intestino primitivo (fig. 24.28a). Poco después, se forma el mesodermo y se divide en dos capas, somática y esplácnica, de las cuales la segunda se relaciona con el endodermo del intestino primitivo, con lo que éste adquiere una pared de doble capa. La capa endodérmica constituye el origen del revestimiento epitelial y glándulas de la mayor parte del tubo digestivo, mientras que la capa mesodérmica produce el músculo liso y tejido conectivo del propio tubo digestivo.
El intestino primitivo se alarga, de modo que en la tercera semana de gestación se diferencia en intestinos anterior, medio y posterior (fig. 24.28b). Hasta la quinta semana de desarrollo embrionario, el intestino medio se abre en el saco vitelino, que ulteriormente se constriñe y se desprende del intestino medio, con lo que éste se sella. En la región del intestino anterior, aparece una depresión de ectodermo, el estomodeo (fig. 24.28c), que es el origen de la boca. La membrana bucal, que separa el intestino anterior del estomodeo, se rompe en la cuarta semana embrionaria, de modo que el intestino anterior presenta continuidad con el exterior a través de la boca. Otra depresión de ectodermo, el proctodeo, se forma en el intestino posterior y finalmente se convierte en el ano. Se rompe la membrana cloacal, que separa el intestino posterior del proctodeo, con lo que el intestino posterior presenta continuidad con el exterior a través del ano. De esta manera, el tubo digestivo se vuelve continuo desde la boca hasta el ano.
El intestino anterior constituye el origen de la faringe, esófago, estómago y una porción del duodeno. El intestino medio se transforma en el resto del duodeno, yeyuno, íleon y ciertas porciones del intestino grueso (ciego, apéndice, colon ascendente y la mayor parte del colon transverso). El intestino posterior se desarrolla en el resto del intestino grueso, excepto una porción del conducto anal derivada del proctodeo.
A medida que avanza el desarrollo, el endodermo de diversos sitios del intestino anterior presenta primordios huecos, que crecen hacia el mesodermo, los cuales se transforman en las glándulas salivales, hígado, vesícula biliar y páncreas (fig. 24.28d). Cada una de las glándulas mantiene su conexión con el tubo digestivo mediante conductos.
ENVEJECIMIENTO Y APARATO DIGESTIVO
Los cambios generales del aparato digestivo relacionados con el envejecimiento comprenden disminución de sus secreciones y de la motilidad de los órganos digestivos, pérdida de tuerza y tono del tejido muscular y sus estructuras de sostén, cambios en la retroalimentación neurosensorial correspondiente a la liberación de enzimas y hormonas, y menor respuesta al dolor y sensaciones internas. En la porción superior del tubo digestivo, son comunes la sensibilidad disminuida a las irritaciones y úlceras bucales, menor sensibilidad gustativa, enfermedad periodontal, dificultades para deglutir (disgeusia) hernia hiatal, gastritis y enfermedad por úlcera péptica. En el intestino delgado, pueden aparecer úlceras duodenales, apendicitis, mala absorción y digestión deficiente. Otros trastornos cuya incidencia aumenta son los de la vesícula biliar, ictericia, cirrosis y pancreatitis aguda. También pueden ocurrir cambios del intestino grueso, como el estreñimiento, hemorroides y enfermedad diverticular. Así mismo, es muy frecuente el cáncer del colon o recto.
TRASTORNOS: DESEQUILIBRIOS HOMEOSTÁTICOS
CARIES DENTAL
La caries dental consiste en la desmineralización (reblandecimiento) gradual de esmalte y dentina. A falta de tratamiento, es factible la invasión de la pulpa por microorganismos, lo cual produce inflamación e infección, con necrosis subsiguiente de la pulpa y absceso del alveolo que rodea el vértice de la raíz. En tales casos, se requiere terapéutica del conducto radicular.
La caries dental se inicia cuando las bacterias, que actúan en los azúcares, producen ácidos que desmineralizan el esmalte. El dextrano, polisacárido adherente que se origina en la sacarosa, hace que las bacterias se adhieran a los dientes. Las masas de células bacterianas, dextrano y otros desechos adheridos a los dientes constituyen la placa dentó bacteriana. La saliva no puede llegar a la superficie de los dientes para amortiguar los ácidos cuando la placa los cubre. El cepillado de dientes inmediatamente después de comer elimina la placa de las superficies planas, antes de que las bacterias produzcan ácidos. Además, los odontólogos recomiendan eliminar la placa cada 24 horas, mediante el hilo dental, de la superficie entre uno y otros dientes.
ENFERMEDAD PERIODONTAL
El término enfermedad periodontal abarca de manera conjunta diversos padecimientos que se caracterizan por inflamación y degeneración de las encías, del hueso alveolar, ligamento periodontal y cemento. Uno de estos trastornos es la piorrea, cuyos síntomas iniciales consisten en hinchazón e inflamación de los tejidos blandos y hemorragia de las encías. A falta de tratamiento, se deterioran los tejidos blandos y ocurre resorción del hueso alveolar, lo cual origina aflojamiento de los dientes y retroceso de las encías. Es frecuente que las enfermedades periodontales sean ocasionadas por higiene bucal deficiente; irritantes locales, como bacterias, alimentos impactados o humo de cigarrillos, o bien por una "mordida" inadecuada.
ENFERMEDAD POR ÚLCERA PÉPTICA
En Estados Unidos, 5 a 10% de la población sutfa la enfermedad por úlcera péptica (EUP). Una úlcera es una lesión a modo de cráter en una membrana; las que surgen en áreas del tubo digestivo expuestas al jugo gástrico ácido se denominan úlceras pépticas. La complicación más común de este trastorno es la hemorragia, que puede causar anemia si se pierde suficiente sangre. En casos agudos, la úlcera péptica es posible origen de choque y muerte. Son tres las causas conocidas de la EUP: 1) la bacteria Helicobacter pylori; 2) el uso de antiinflamatorios no esteroideos (AINE), como el ácido acetilsalicílico, y 3) la hipersecreción de HC1, como ocurre en el síndrome de Zollinger-Ellison, tumor productor de gastrina, por lo común pancreático.
La Helicobacter pylori (antes llamada Campylobacter pylori) es la causa más frecuente de úlcera péptica. La bacteria produce una enzima, la ureasa, que desdobla la urea en dióxido de carbono y amoniaco. Este último protege la bacteria contra la acidez gástrica y al mismo tiempo daña la mucosa protectora del estómago y las células gástricas subyacentes. La H. pylori también produce catalasa, en zima que la protege contra la acción fagocitaría de los neutrófilos, así como diversas proteínas de adhesión que permiten a la bacteria unirse a las células gástricas.
Son varías las opciones de tratamiento útiles contra la úlcera péptica. Deben evitarse sustancias como el etanol, la cafeína, los AINE y el humo de los cigarrillos ya que obstaculizan los mecanismos de defensa de la mucosa y la hacen más susceptible a los efectos dañinos del HC1. En los casos relacionados con H. pylori, es frecuente que la administración de un antibiótico resuelva el problema. Los antiácidos orales también ayudan transitoriamente a amortiguar el ácido gástrico. Cuando el trastorno se debe a la hipersecreción de HC1, suelen administrarse bloqueadores de la histamina H2 o inhibidores de la bomba de protones, como el omeprazol, que bloquean la secreción de H+ en las células parietales.
DIVERTICULITIS
La diverticulosis es la formación de divertículos, protuberancias en forma de saco de la pared colónica, en sitios donde está debilitada la muscular. Muchas personas con este trastorno no experimentan síntomas ni complicaciones; pero en 15% de los casos surge, en última instancia, la inflamación llamada diverticuíitis. Esta se caracteriza por dolor, estreñimiento o aumento en la frecuencia de evacuaciones, náusea, vómito y febrícula. La dieta baja en fibra contribuye a la diverticuíitis, de modo que el cambio a una alimentación rica en fibra suele generar alivio considerable de los síntomas. En casos graves, puede ser necesaria la excisión quirúrgica de las porciones afectadas del colon. Si ocurre la rotura de los divertículos, la liberación de bacterias en la cavidad abdominal puede causar peritonitis.
CÁNCER COLORRECTAL
El cáncer colorrectal se cuenta entre los tumores malignos de mortalidad más alta, ya que ocupa el segundo lugar en varones, después del cáncer pulmonar, y el tercero en mujeres, luego de los cánceres pulmonar y mamario. La herencia desempeña una función muy importante, puesto que la predisposición hereditaria contribuye con más de la mitad de los casos de cáncer colorrectal. El consumo de etanol y las dietas ricas en grasas y proteínas de origen animal se relacionan con riesgo incrementado de este trastorno, mientras que la fibra de los alimentos, retinoides, calcio y selenio tendrían efectos protectores contra él. Los signos y síntomas del cáncer colorrectal abarcan diarrea, estreñimiento, cólicos, dolor abdominal y hemorragia rectal visible u oculta. Las pruebas de detección de este padecimiento incluyen análisis en busca de sangre en las heces, palpación rectal digital, sigmoidoscopia, colonoscopia y enema de bario. El tumor puede extirparse mediante endoscopia o intervención quirúrgica abierta.
HEPATITIS
La hepatitis es una inflamación del hígado que puede estar causada por virus, fármacos y otras sustancias químicas, incluido el etanol. Desde el punto de vista clínico, son varios los tipos de hepatitis viral. La hepatitis A (hepatitis infecciosa) se debe al virus de la hepatitis A y se disemina por la contaminación fecal de alimentos, ropa, juguetes y utensilios para comer (vía fecaloral). En general, se trata de una enfermedad leve en niños y adultos jóvenes, caracterizada por inapetencia, malestar general, náusea, diarrea, fiebre, escalofríos y, en última instancia, ictericia. Esta forma de hepatitis no causa daño duradero al hígado. La mayoría de los pacientes se recupera en cuatro a seis semanas.
La hepatitis B, que causa el virus de la hepatitis B, se disemina principalmente por contacto sexual o por jeringas y equipo de transfusión contaminados. También puede transmitirse por la saliva y las lágrimas. El virus puede estar presente en la persona durante años o incluso toda su vida y causar cirrosis y, posiblemente, cáncer hepático. Las personas infectadas por el virus de la hepatitis B activo están en riesgo de cirrosis y de volverse portadoras. Se cuenta con vacunas, producidas con la tecnología de DNA recombinante, para prevenir esta infección.
La hepatitis C, debida al virus de la hepatitis C, guarda similitud clínica con la hepatitis B y suele diseminarse por transfusiones sanguíneas. Es posible origen de cirrosis y cáncer hepático.
La hepatitis D resulta de la infección por el virus de la hepatitis D. Se transmite como la hepatitis B y, de hecho, una persona puede estar infectada por el virus de esta última antes de contraer la hepatitis D. Produce daño hepático grave y su tasa de mortalidad es más alta que la de la infección por el virus de la hepatitis B solo.
La hepatitis E, debida al virus de la hepatitis E, se disemina como la hepatitis A. Pese a que no produce enfermedad hepática crónica, este virus origina una tasa de mortalidad muy alta en embarazadas.
ANOREXIA NERVIOSA
La anorexia nerviosa es una enfermedad crónica caracterizada por disminución ponderal que se induce la propia persona, percepción negativa de la imagen corporal de sí misma y cambios fisiológicos resultantes de déficit nutricionales. Los individuos con anorexia nerviosa tienen una fijación de controlar su peso y suelen insistir en la necesidad de evacuar los intestinos diariamente pese a la ingesta deficiente de alimentos. Abusan de los laxantes, lo cual agrava los desequilibrios hidroelectrolíticos y los déficit nutricionales. Es un trastorno que afecta principalmente a mujeres jóvenes solteras y podría ser hereditario. Las anormalidades de la menstruación, incluida la amenorrea (ausencia de menstruación), y la disminución del índice del metabolismo basal, reflejan los efectos depresivos de la inanición. La persona sufre emaciación y puede, finalmente, morir de la propia inanición o de alguna de sus complicaciones. El trastorno suele ir acompañado de osteoporosis, depresión y anormalidades encefálicas relacionadas con deterioro de los procesos mentales. El tratamiento consiste en psicoterapia y tratamiento dietético.
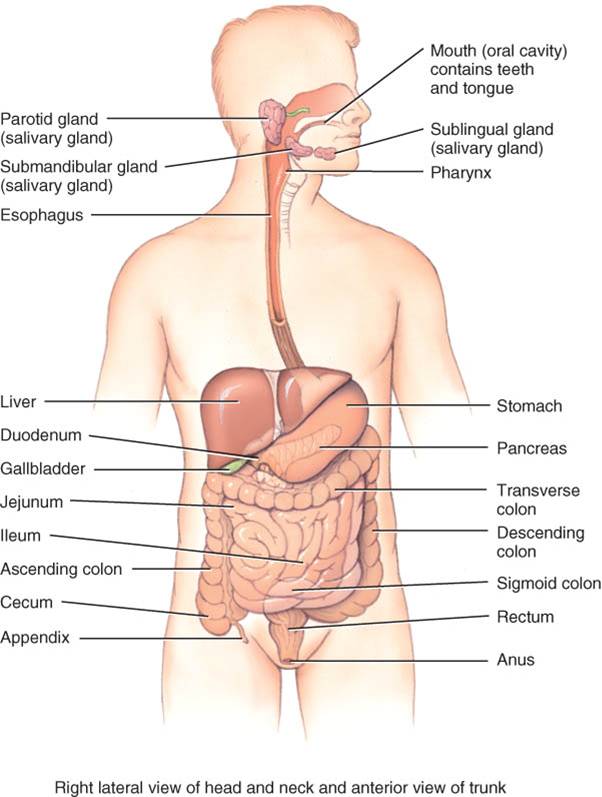{kind=link}
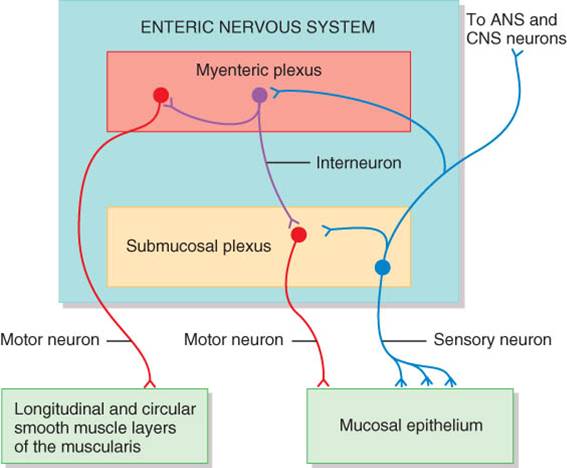{kind=link}
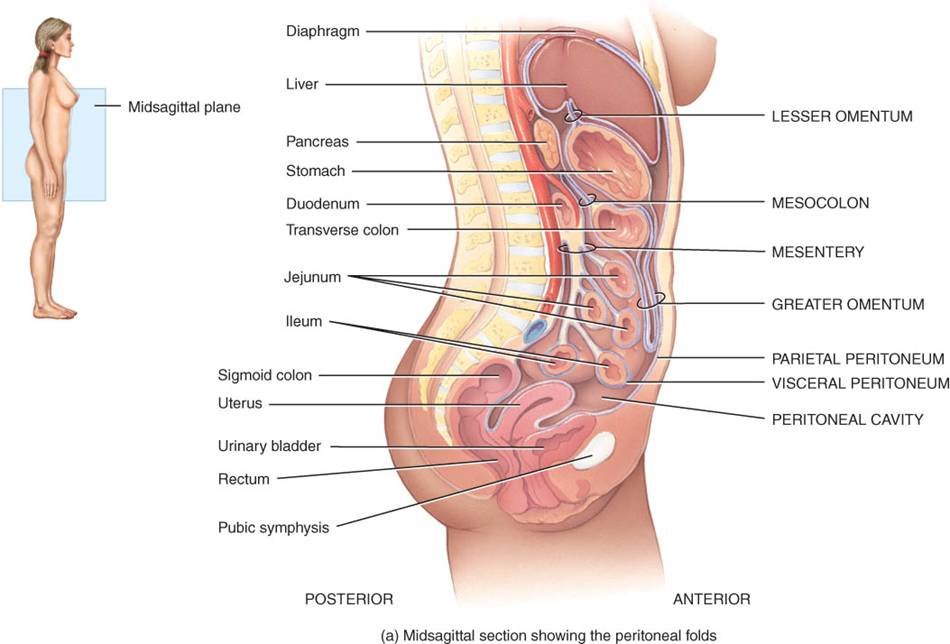{kind=link}
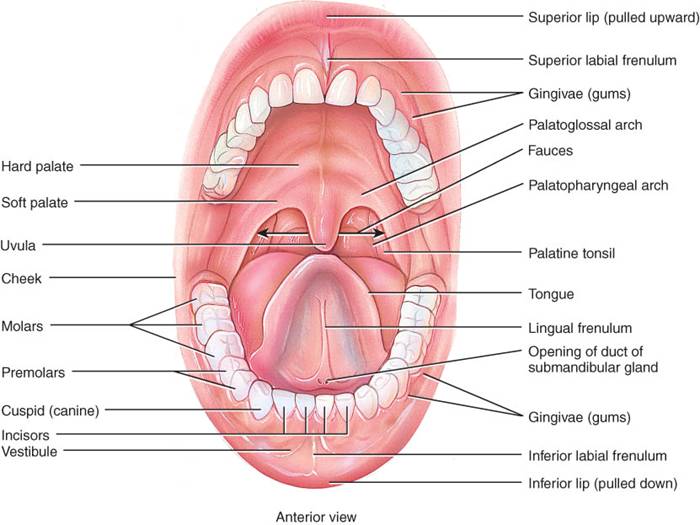{kind=link}
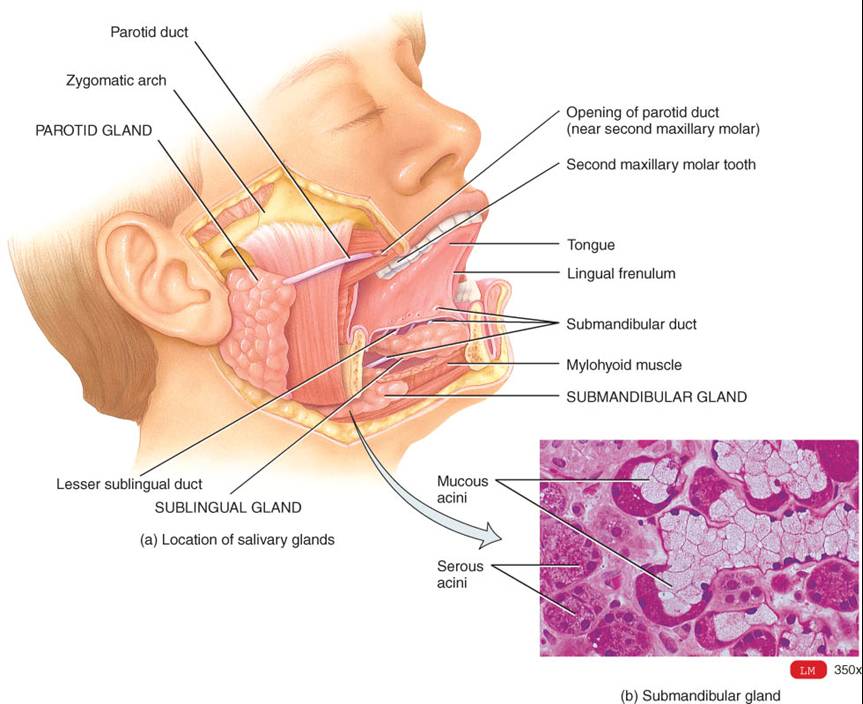{kind=link}
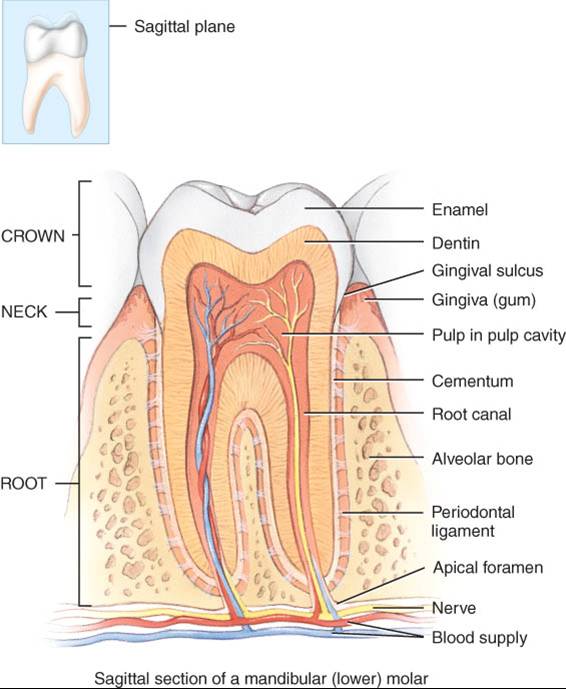{kind=link}
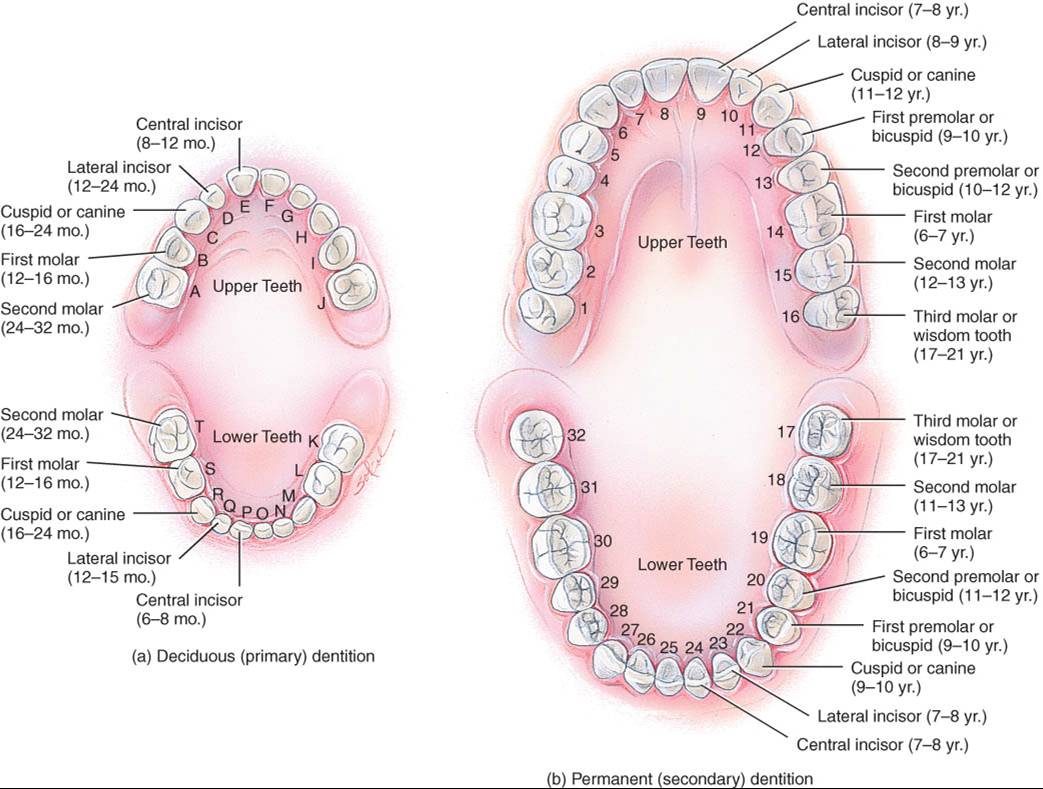{kind=link}
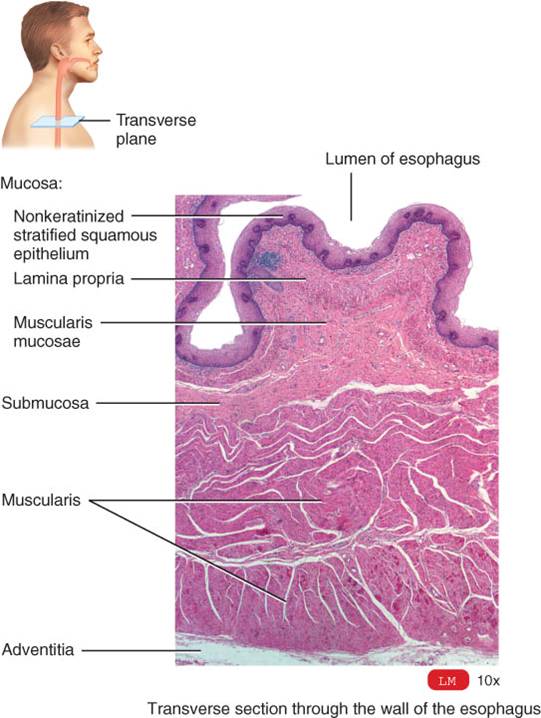{kind=link}
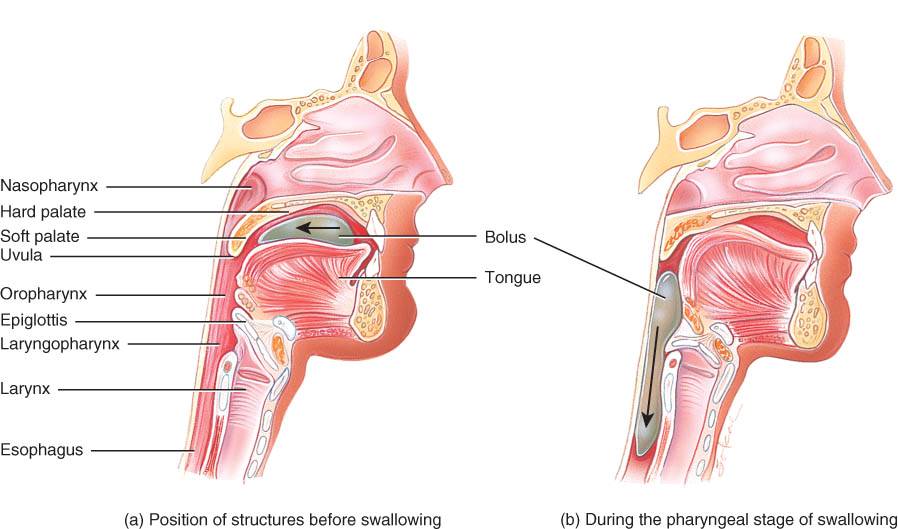{kind=link}
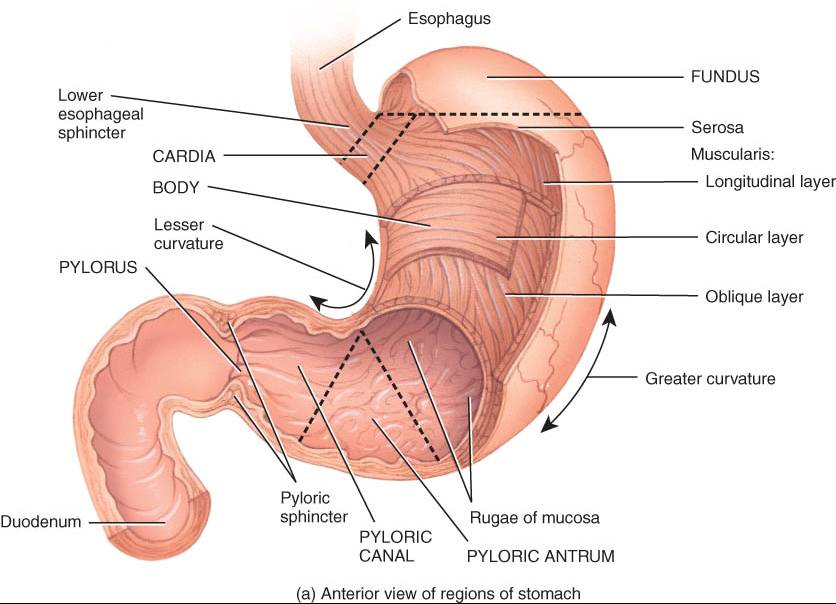{kind=link}
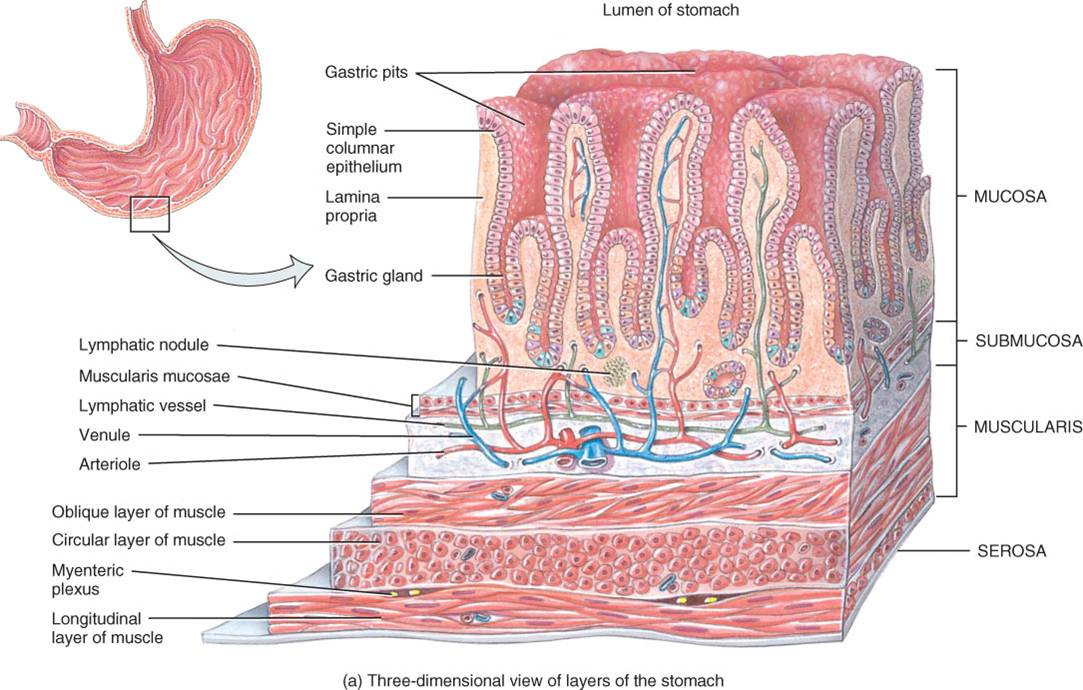{kind=link}
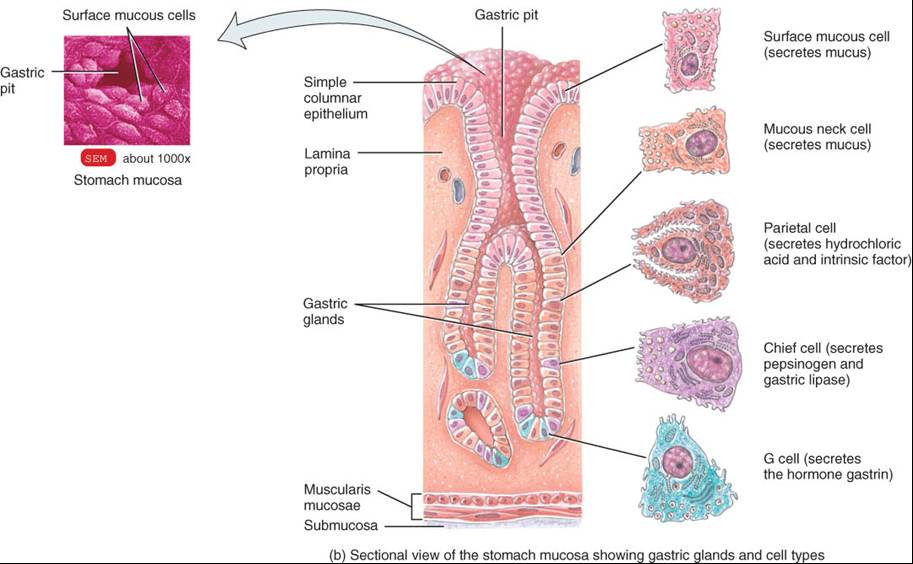{kind=link}
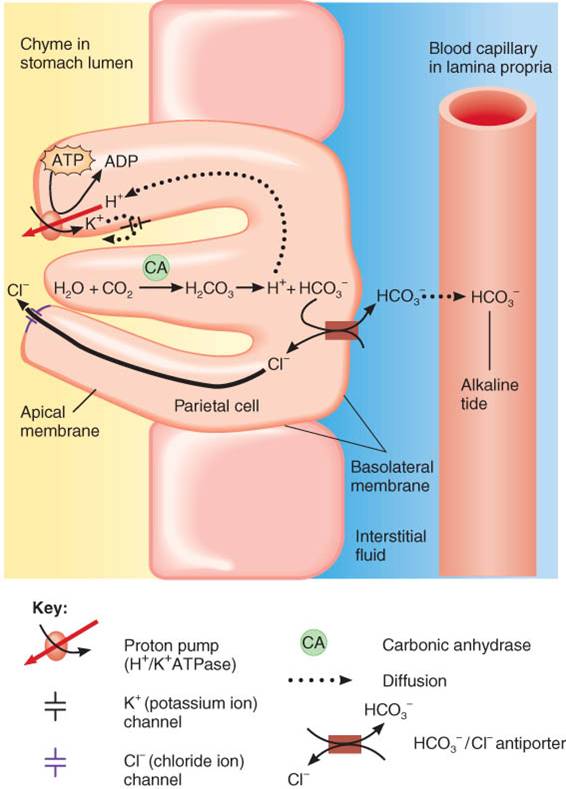{kind=link}
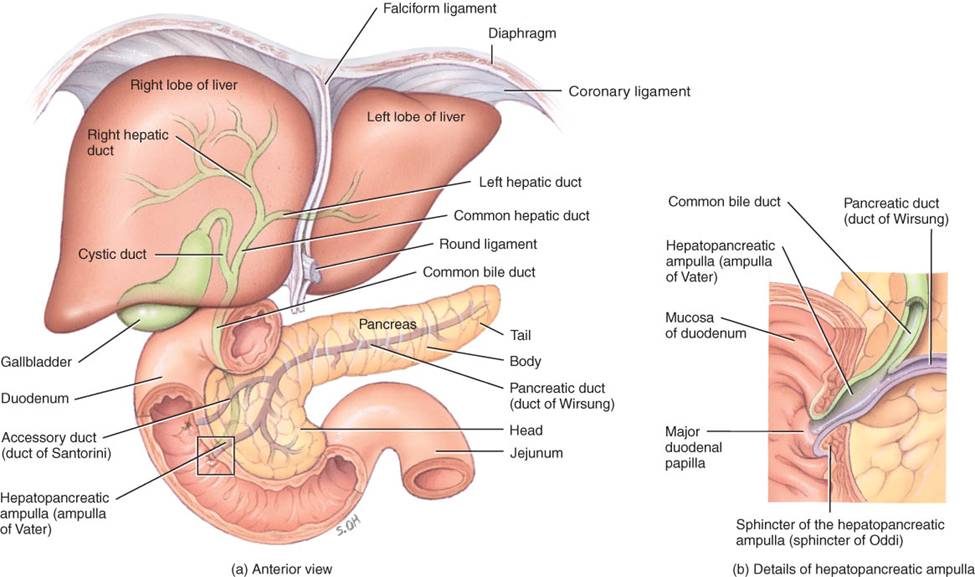{kind=link}
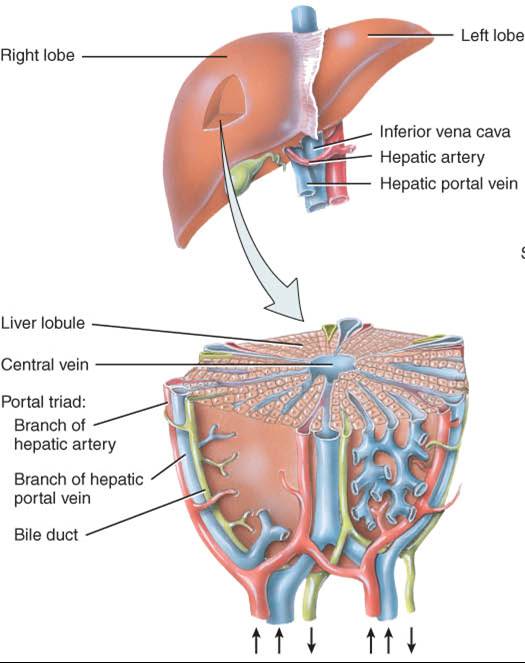{kind=link}
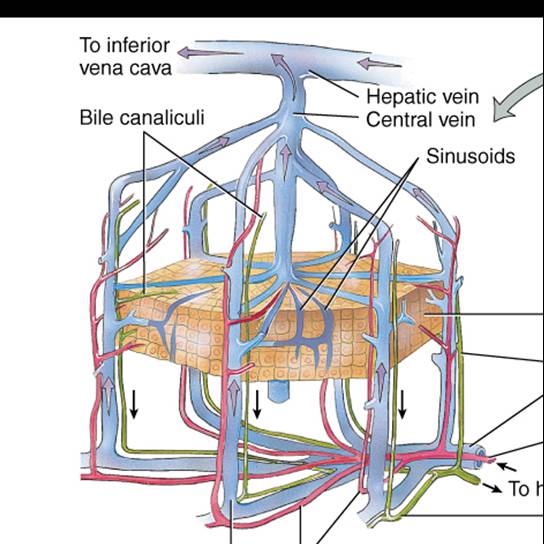{kind=link}
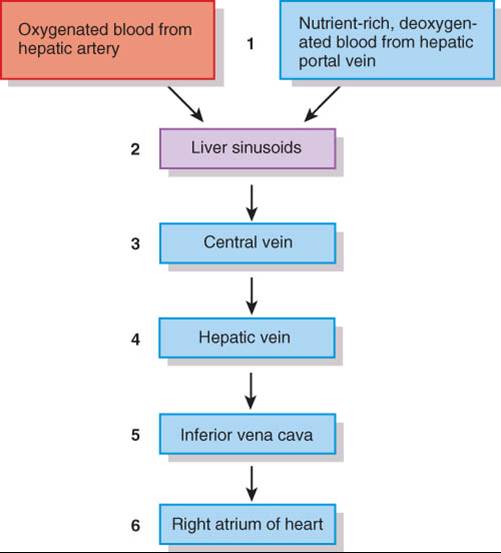{kind=link}
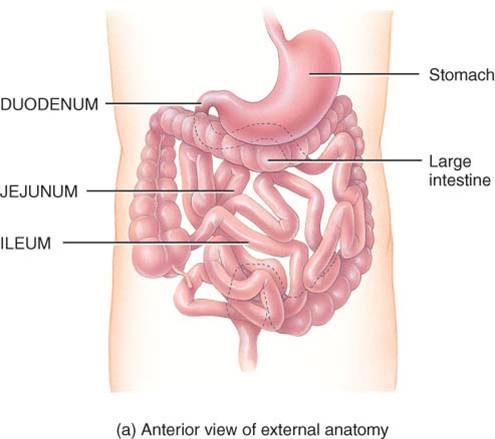{kind=link}
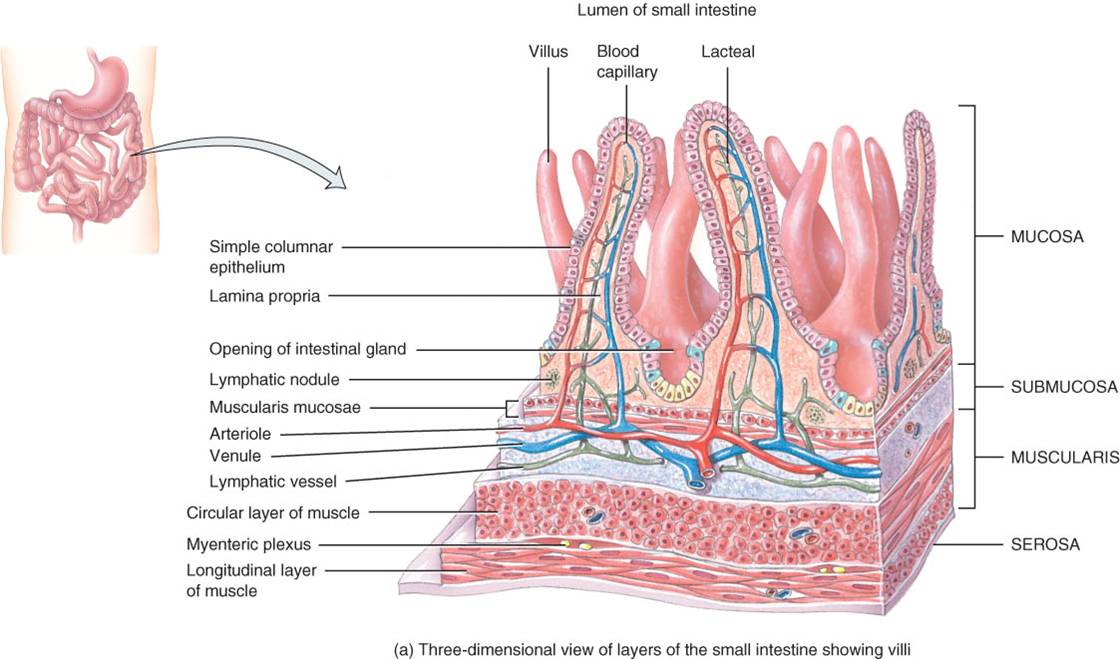{kind=link}
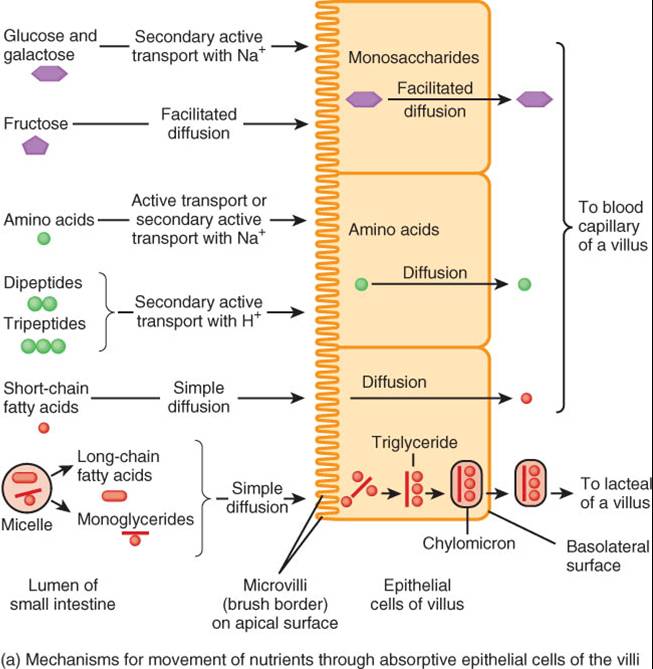{kind=link}
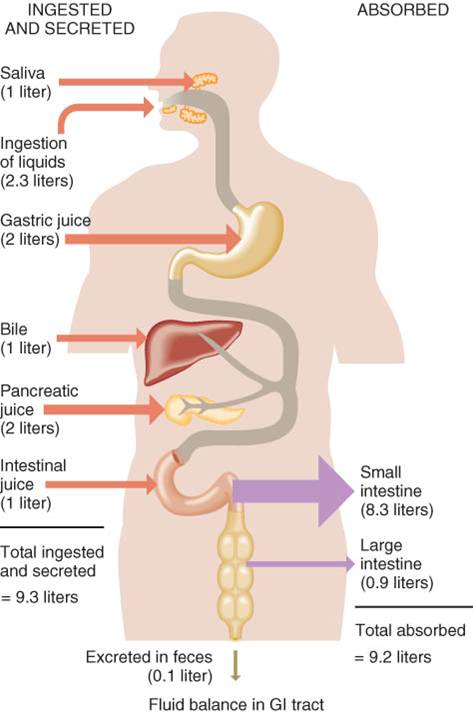{kind=link}
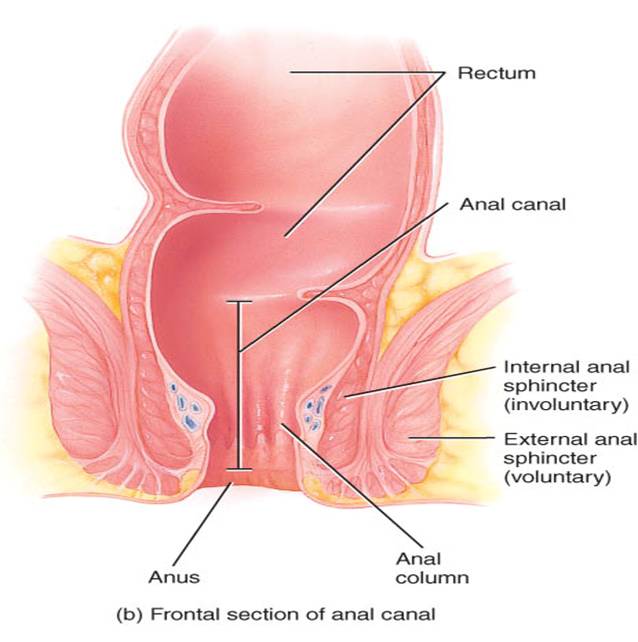{kind=link}
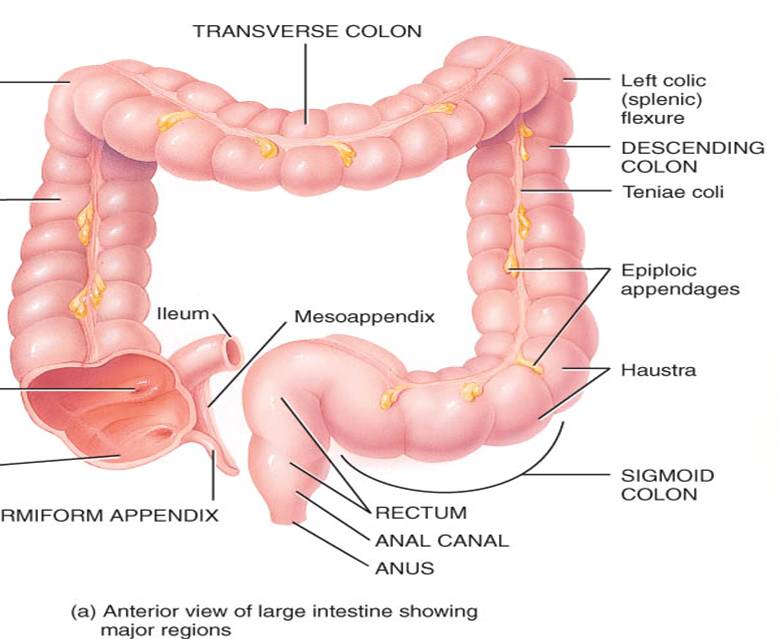{kind=link}
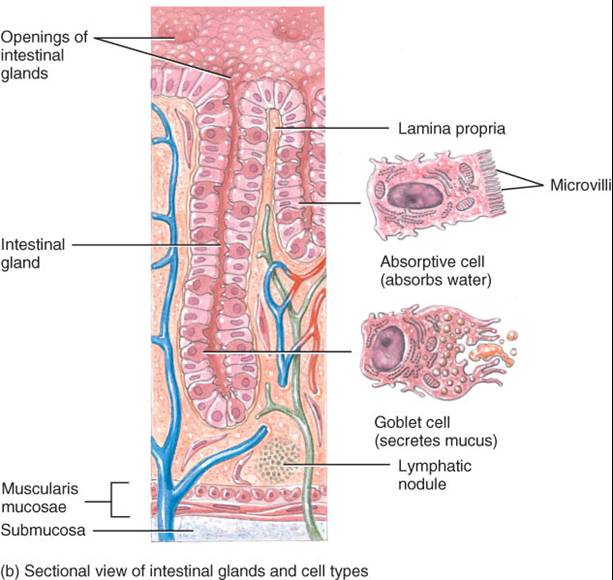{kind=link}
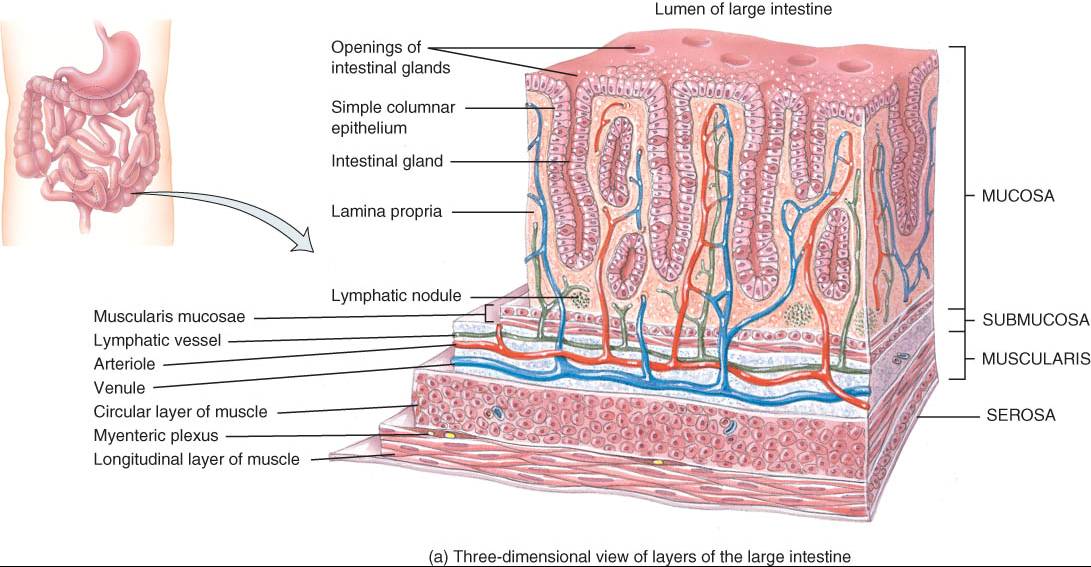{kind=link}
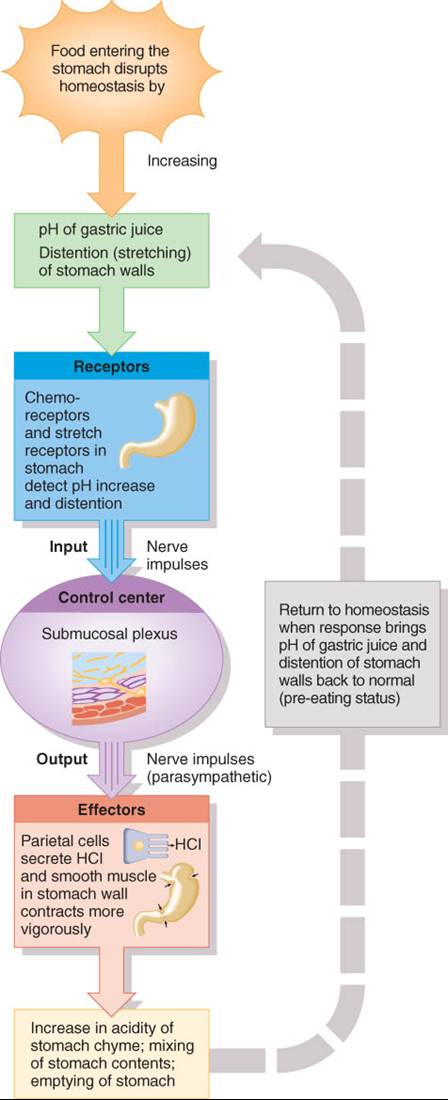{kind=link}