SISTEMA LINFÁTICO, RESISTENCIA INESPECÍFICA A
ENFERMEDADES E INMUNIDAD
La capacidad del organismo para evitar enfermedades se llama resistencia, y la falta de ésta o la vulnerabilidad se denomina susceptibilidad. La resistencia a los trastornos es de dos tipos, inespecífica y específica. La primera comprende mecanismos de defensa que brindan protección inmediata a la vez que general contra la invasión por una amplia gama de microorganismos patógenos, como bacterias, virus y parásitos, que son microbios productores de enfermedades. La primera línea de defensa en la resistencia inespecífica corresponde a las barreras físicas y químicas que constituyen la piel y mucosas; a manera de ejemplo, la acidez del contenido gástrico mata muchas bacterias ingeridas con los alimentos. La resistencia específica o inmunidad surge más lentamente e incluye la activación de linfocitos específicos, que combaten microbios patógenos o sustancias extrañas particulares. El sistema corporal que se encarga de la inmunidad es el linfático. En este capítulo, se describen los mecanismos que proporcionan defensas contra los invasores e inducen la reparación de los tejidos corporales dañados.
SISTEMA LINFÁTICO
OBJETIVOS
El sistema linfático consta de un líquido, la linfa, que fluye por los vasos linfáticos, diversos órganos y tejido, y la médula ósea roja, la cual contiene las células madre de las que se originan los linfocitos (fig. 22.1). La composición del líquido intersticial y la linfa es, en lo fundamental, idéntica, y la diferencia entre ellos radica en su localización. Después de que el líquido recorre los espacios intersticiales y pasa a los vasos linfáticos, su nombre cambia a linfa. El tejido linfático es un tipo especializado de tejido conectivo que contienen numerosos linfocitos.
Funciones del sistema linfático
El sistema linfático desempeña tres funciones principales:
Vasos linfáticos y circulación linfática
Los vasos linfáticos principian como capilares linfáticos, los cuales son conductos cerrados en un extremo que se localizan en los espacios intercelulares (fig. 22.2). Justo en el punto en que los capilares sanguíneos convergen y forman vénulas y venas, los linfáticos se unen y dan origen a los vasos linfáticos más grandes (fig. 22.1), similares a las venas en su estructura, si bien de paredes más delgadas y con más válvulas. A intervalos variables en dichos vasos hay estructuras de tejido linfático, los ganglios linfáticos. En la piel, los vasos mencionados están en el tejido subcutáneo y generalmente acompañan a las venas, mientras que en las vísceras suelen seguir el trayecto de las arterias y formar plexos (redes) alrededor de ellas.
Capilares linfáticos
Estos capilares, cuyo diámetro es un poco mayor que los sanguíneos, se hallan en todo el cuerpo, salvo en los tejidos avasculares (cartílago, epidermis y córnea), sistema nervioso central, ciertas partes del bazo y médula ósea roja. La estructura singular de los capilares linfáticos permite que el líquido intersticial fluya al interior de ellos, pero no hacia afuera. Los bordes de las células endoteliales que componen la pared de un capilar linfático están sobrepuestos; cuando la presión es mayor en el líquido intersticial que en la linfa, las células se separan ligeramente, como al abrirse una válvula unidireccional, y el líquido entra en el capilar (fig. 22.2b). Si la presión es mayor en este último, las células se adhieren estrechamente entre sí e impiden que la linfa regrese al líquido intersticial. Los filamentos de anclaje, dispuestos en ángulo recto al capilar, fijan las células endoteliales de éste al tejido circundante. La acumulación excesiva de líquido en los tejidos y el edema consecuente empuja dichos filamentos, lo cual agranda los espacios entre las células endoteliales y permite que fluya más líquido dentro del capilar linfático.
En el intestino delgado, existen capilares linfáticos especializados, los vasos quilíferos, por los que circulan los lípidos alimentarios a la linfa y, en última instancia, a la sangre. Dichas sustancias hacen que la linfa que drena del intestino delgado, denominada quilo, sea de color blanco cremoso.
En el resto del cuerpo, la linfa es un líquido transparente de color amarillo pálido.
Troncos y conductos linfáticos
La linfa pasa de los capilares linfáticos a los vasos y llega a los ganglios linfáticos. Los vasos que nacen de los ganglios la llevan a otro del mismo grupo o a otro grupo de ganglios.
En el grupo más proximal de cada cadena de ganglios, los vasos de salida se unen y forman los troncos linfáticos. De estos, los principales son: lumbar, intestinal, broncomediastínico, subclavio y yugular (fig. 22.3). Los troncos más importantes vacían la linfa en dos vasos de gran calibre, los conductos torácico y linfático derecho, de los cuales la linfa pasa a la sangre venosa.
El conducto torácico ( o linfático izquierdo) tiene unos 38 a 45 cm. de longitud y se inicia como una dilatación, la cisterna del quilo, a la altura de la segunda vertebre lumbar. Es el más importante recolector del sistema linfático y recibe linfa de la mitad izquierda de la cabeza, cuello y tórax, extremidad superior izquierda y todo el cuerpo en plano inferior a las costillas. Vacía su linfa en la sangre venosa por medio de la vena subclavia izquierda.
La cisterna del quilo recibe linfa de los troncos lumbares derecho e izquierdo, así como del tronco intestinal. Los lumbares drenan linfa de las extremidades inferiores, pared y vísceras pélvicas, riñones, suprarrenales y vasos linfáticos profundos; estos últimos la drenan de casi toda la pared abdominal. A los troncos intestinales llega linfa del estómago, intestinos, páncreas, bazo y parte del hígado.
En el cuello, el conducto torácico recibe linfa de los troncos yugular, subclavio, broncomediastínico izquierdos.
El yugular izquierdo la drena de la cabeza y el cuello, y el subclavio izquierdo, de la extremidad superior izquierda. La del tronco broncomediastínico proviene de la mitad izquierda de partes profundas de la pared torácica anterior, parte superior de la pared abdominal anterior, parte frontal del diafragma, pulmón izquierdo y lado izquierdo del corazón.
El conducto linfático derecho (fig. 22.3) tiene cerca de 1.25 cm. de longitud y drena linfa de la mitad superior derecha del cuerpo, la cual se incorpora a la sangre venosa en la vena subclavia derecha. Tres troncos linfáticos se vacían en ese conducto: el yugular derecho, con linfa de la mitad derecha de cabeza y cuello; el subclavio derecho, al cual llega el linfa de la extremidad superior del mismo lado, y el broncomediastínico derecho, en el cual se vierte la linfa del lado derecho del tórax, pulmón derecho, mitad derecha del corazón y una parte del hígado.
Formación y flujo de la linfa
La mayoría de los componentes del plasma sanguíneo se filtran irrestrictamente a través de la pared de los capilares y forman el líquido intersticial. Sin embargo, el volumen de líquido filtrado de los capilares es mayor que el devuelto a ellos por reabsorción (fig.21.7). El exceso de líquido filtrado, unos 3 L/día, drena a los vasos linfáticos y se convierte en linfa. Muchas proteínas plasmáticas son demasiado grandes para salir de los vasos sanguíneos, de modo que el líquido intersticial contiene cantidades mínimas de proteínas. No obstante, las que sí salen del plasma no pueden regresar directamente a la sangre por difusión, ya que lo impide el gradiente de su concentración (alta en los capilares sanguíneos y baja en el espacio intersticial). Así pues, una importante función de los vasos linfáticos consiste en regresar proteínas plasmáticas al torrente sanguíneo.
En última instancia, la linfa drena en la sangre venosa por los conductos torácico y linfático derecho, en la unión de las venas yugular interna y subclavia (fig. 22.3). Este orden de flujo, así como la relación de los sistemas linfático y cardiovascular, se ilustra en la figura 22.4.
Recuérdese que las bombas muscular y respiratoria facilitan el retorno de la sangre venosa el corazón. De igual modo, estas dos bombas impulsan el flujo de la linfa, proveniente de los espacios intersticiales a los grandes conductos linfáticos y luego a las venas subclavias. Las contracciones musculares comprimen los vasos linfáticos y desplazan su contenido hacia los vasos subclavios. Los vasos linfáticos poseen válvulas unidireccionales, semejantes a las venosas, que impiden el flujo de la linfa. La respiración hace que cambie la presión en el interior del sistema linfático. Con cada inhalación, la linfa fluye de la región abdominal, donde la presión es mayor, hacia el tórax, donde es menor. De igual modo, la caída de la presión abdominal durante la exhalación facilita el flujo de linfa de los vasos distales a los del abdomen. Además, la dilatación de los vasos linfáticos hace que se contraiga el músculo liso de su pared, lo cual ayuda a mover la linfa de un segmento del vaso a otro
Órganos y tejidos linfáticos
Los órganos y tejidos del sistema linfático, distribuidos ampliamente en todo el cuerpo, se clasifican en dos grupos con base en sus funciones. Los órganos linfáticos primarios constituyen el ambiente idóneo para la división de las células madres y su maduración en células B y T, o sea, los linfocitos que se encargan de las respuestas inmunitarias. Dichos órganos son la médula ósea roja, que se encuentra en los huesos planos y en la epífisis de los huesos largos de adultos, y el timo. Las células madre pluripotenciales de la médula ósea roja son el origen de las células B maduras y las células pre-T, de las cuales estas últimas emigran al timo y maduren en él. Los órganos y tejidos linfáticos secundarios, donde tiene lugar gran parte de las respuestas inmunitarias, comprenden los ganglios y folículos linfáticos y el Bazo. Se considera que el timo, los ganglios y el bazo son órganos por que los rodea una capsula de tejido conectivo, mientras que los folículos no lo son por carecer de ella.
Timo
Esta glándula por lo regular consta de dos lóbulos y se localiza en el mediastino, detrás del esternón (fig. 22.5a). Una capa de tejido conectivo envuelve y mantiene unidos los dos lóbulos tímicos; mientras que una cápsula de tejido conectivo delimita por separado cada lóbulo. Las trábeculas son prolongaciones de las cápsulas que penetran en los lóbulos y los dividen en lobulillos (fig. 22.5b). Cada uno consta de corteza externa, que se tiñe de color oscuro, y médula de color claro a la tinción. La corteza se compone de linfocitos estrechamente apiñados, células epiteliales denominadas epiteliales reticulares que rodean a grupos de linfocitos, y macrófagos. La médula contiene ante todo células epiteliales reticulares, además de linfocitos muy dispersos. Aunque se conocen solo algunas de sus funciones, las células reticulares producen hormonas tímicas, que, según se piensa, contribuyen a la maduración de las células T. además en la médula existen los corpúsculos del timo (o de Hassall) característicos, los cuales son capas concéntricas de células epiteliales reticulares aplanadas y llenas de gránulos queratohialina y queratina (fig. 22.5c).
En los lactantes, el timo es una glándula de gran tamaño, con masa de unos 70g. Después de la pubertad, se inicia el remplazó del tejido tímico con tejido adiposo y conectivo areolar. Cuando la persona alcanza la madurez, la glándula se ha atrofiado considerablemente, mientras que en la edad avanzada pesa apenas unos 3g.
Ganglios linfáticos
Se llaman ganglios linfáticos a casi 600 órganos en forma de frijol dispuestos a lo largo de los vasos linfáticos. Están dispersos en todo el cuerpo, tanto en capas superficiales como profundas, usualmente en grupos (fig. 22.1). Los ganglios son abundantes cerca de las glándulas mamarias y en las axilas e ingles.
Por lo regular miden de 1a 25mm de largo y los cubre una cápsula de tejido conectivo denso, que penetra en el ganglio (fig. 22.6). Sus prolongaciones, llamadas trábeculas, dividen el ganglio en compartimientos, le brindan sostén y constituyen una ruta para los vasos sanguíneos que penetran en el ganglio. En plano interno a la cápsula, existe una red de sostén que forman fibras reticulares y fibroblastos. Cápsula, trábeculas, fibras reticulares y fibroblastos constituyen el estroma o estructura del ganglio. El parénquima de los ganglios linfáticos se especializa en dos regiones, la corteza superficial y la médula interna. La corteza externa contiene folículos linfáticos de células (linfocitos)B. Las áreas de los folículos que se tiñen de claro, llamadas centros germinativos, son el sitio donde las células B se diferencian en células plasmáticas, secretoras de anticuerpos. Las células reticulares de dichos centros, llamadas dendríticas, desempeñan funciones como antígenos, que ayudan a iniciar las respuestas inmunitarias. Además, los centros germinativos contienen macrófagos. En plano profundo a la corteza externa, está la interna, que contiene células (linfocitos)T. La médula de los ganglios linfáticos incluye células B y plasmáticas en filamentos estrechamente apiñados, los cordones medulares.
La linfa fluye por los ganglios linfáticos en una sola dirección. Entra por los vasos linfáticos aferentes, que penetran la superficie convexa del ganglio en varios puntos. Dichos vasos poseen válvulas que se abren en dirección al centro del ganglio, de modo que la linfa fluye hacia el interior del mismo. Una vez dentro, la linfa llega a los senos, conjunto de conductos irregulares que incluyen macrófagos, linfocitos y fibras reticulares que se ramifican. Desde los vasos linfáticos aferentes, la linfa fluye hacia el seno subcapsular, situado inmediatamente bajo la cápsula. Luego, se dirige a los senos trabeculares, que se extienden por la corteza, con trayecto paralelo a las trábeculas. Desde ellos, la linfa fluye a los senos medulares, que se extienden por la médula. Los senos medulares drenan en uno o más vasos linfáticos eferentes, de mayor calibre y menos numerosos que los aferentes. Poseen válvulas que se abren en dirección contraria al centro del ganglio para sacar la linfa. Estos vasos nacen de un lado del ganglio en una pequeña depresión, llamada hilio. Además, entran y salen vasos sanguíneos del ganglio por el hilio.
Los ganglios son las únicas estructuras que filtran la linfa. Dentro de los senos ganglionares, las fibras reticulares atrapan las sustancias extrañas que flotan en la linfa. Luego, los macrófagos destruyen algunas de esas sustancias por fagocitosis, mientras que los linfocitos lo hacen mediante diversas respuestas inmunitarias. Después, el líquido filtrado sale por el otro extremo del ganglio. Las células T y plasmáticas que proliferan en los ganglios también pueden salir en la linfa y circular a otras partes del cuerpo.
APLICACIÓN CLÍNICA
Metástasis a través del sistema linfático
Una metástasis es la diseminación de una enfermedad de una órgano a otro que no están conectados de manera directa. Es un fenómeno que caracteriza todos los tumores malignos. Las células cancerosas suelen emigrar por la sangre o linfa y formar nuevos tumores en el sitio donde se alojan. Cuando las metástasis se diseminan por el sistema linfático, es factible predecir los sitios de tumores secundarios con base en la dirección de flujo de la linfa desde el tumor primario. Al palparlos, se advierte que los ganglios linfáticos cancerosos están agrandados, con consistencia firme, sin dolor a la palpación y fijos a las estructuras subyacentes. En contraste, los ganglios linfáticos que se agrandan como resultado de infecciones no son de consistencia firme y son móviles y muy sensibles a la palpación.
Bazo
El bazo es un órgano oval y la masa más grande de tejido linfático del cuerpo, con unos 12cm de longitud (fig. 22.7a). Se sitúa en el hipocondrio izquierdo, entre el estómago y el diafragma. Su cara superior, lisa y convexa, se conforma a la superficie cóncava del diafragma. Los órganos adyacentes producen depresiones en la cara visceral del bazo: la impresión gástrica para el estómago, la renal para el riñón y la cólica para el ángulo esplénico del colon. Al igual que los ganglios linfáticos, el bazo posee un hilio, que cruzan la arteria y vena esplénicas, así como vasos linfáticos eferentes.
Una cápsula de tejido conectivo denso envuelve al bazo. Las trabéculas penetran desde la cápsula, que a su vez tiene el recubrimiento de una membrana serosa, el peritoneo visceral. Cápsula, trabéculas, fibras reticulares y fibroblastos constituyen el estroma del bazo, mientras que su parénquima consiste en dos tipos de tejido, las pulpas blanca y roja (fig. 22.7b). La pulpa blanca consta de tejido linfático, principalmente linfocitos y macrófagos, dispuestos alrededor de ramas de la arteria esplénica, llamadas arterias centrales. La pulpa roja está formada por senos venosos llenos de sangre y cordones de tejido esplénico, llamados cordones esplénicos (de la pulpa roja o de Billroth). Éstos se componen de eritrocitos, macrófagos, linfocitos, células plasmáticas y granulocitos. Las venas guardan relación estrecha con la pulpa roja.
La sangre que fluye hacia el bazo por la arteria esplénica llega a las arterias centrales de la pulpa blanca. En ésta, las células T y B llevan a cabo funciones inmunitarias, al tiempo que los macrófagos destruyen los microbios patógenos provenientes de la sangre por fagocitosis. La pulpa roja del bazo se encarga de tres funciones relacionadas con los elementos formes de la sangre: 1) eliminación de células sanguíneas y plaquetas defectuosas, que efectúan los macrófagos; 2) almacenamiento de plaquetas, que pueden ser hasta un tercio del total, y, 3) producción de células sanguíneas (hemopoyesis) durante la vida fetal.
El bazo es el órgano afectado con mayor frecuencia en caso de traumatismo abdominal. Su rotura produce hemorragia intraperitoneal grave y choque. La extirpación pronta del órgano (esplectomía) es necesaria para evitar que el paciente se desangre hasta morir. Otras estructuras, en particular la médula ósea roja y el hígado, pueden encargarse de las funciones que normalmente realiza el bazo.
Folículos linfáticos
Son concentraciones ovales de tejido linfático que no están rodeadas por una cápsula. Debido a que se encuentran dispersos en la lámina propia (tejido conectivo) de la mucosa que reviste los aparatos digestivo, urinario, reproductivo y respiratorio, los folículos linfáticos también se denominan tejido linfoide relacionado con mucosas (TLRM).
Aunque muchos ganglios linfáticos son pequeños y están aislados, otros forman grandes agregados en partes específicas del cuerpo. Entre estos se incluyen las amígdalas en la región faríngea y las placas de Peyer en el íleon. También se encuentran tales cúmulos en el apéndice. Por lo general hay cinco amígdalas en la unión de la boca con la bucofaringe y de la nariz con la nasofaringe (fig. 23.2b). Así pues, las amígdalas están dispuestas estratégicamente para participar en respuestas inmunitarias contra sustancias extrañas inhaladas o ingeridas. La amígdala faríngea o tejido adenoide esta incluido en la pared posterior de la nasofaringe. Las dos amígdalas palatinas se localizan en la parte posterior de la boca, una en cada lado, y son las que habitualmente se extraen en la amigdalectomía. El par de amígdalas linguales, situadas en la base de la lengua, también se extirpan a veces en dicha intervención quirúrgica.
ANATOMÍA DEL DESARROLLO
DEL SISTEMA LINFÁTICO
El sistema linfático empieza a formarse hacia el final de la quinta semana de vida embrionaria. Los vasos linfáticos se desarrollan a partir de sacos linfáticos que surgen de las venas en desarrollo, las cuales se derivan del mesodermo.
Los primeros sacos linfáticos que aparecen son el par de sacos linfáticos yugulares, en la unión de las venas yugular interna y subclavia (fig. 22.8). A partir de dichos sacos, los plexos capilares se diseminan al tórax, a las extremidades superiores, el cuello y la cabeza. Algunos plexos crecen y forman vasos linfáticos en las regiones correspondientes. Cada saco linfático yugular conserva al menos una conexión con la vena yugular respectiva, y el izquierdo se transforma en la porción superior del conducto torácico o linfático izquierdo.
El siguiente en aparecer es el saco linfático retroperitoneal impar, en la raíz del mesenterio intestinal. Se desarrolla a partir de la vena cava primitiva (común) y las venas mesonéfricas (del riñón primitivo). Los plexos de capilares y vasos linfáticos se diseminan a partir del saco mencionado a las vísceras abdominales y el diafragma. El saco establece conexiones con la cisterna del quilo; pero pierde las que lo unen con las venas cercanas.
Casi al mismo tiempo que el linfático retroperitoneal, se desarrolla otro saco, la cisterna del quilo, sobre la pared abdominal posterior, en plano infradiafragmático. Es el origen de la porción inferior del conducto torácico y de la cisterna del quilo, en la base de dicho conducto. Al igual que el saco retroperitoneal, la cisterna del quilo pierde sus conexiones con las venas circundantes.
El último de los sacos linfáticos es el par de sacos linfáticos posteriores, que se desarrollan a partir de las venas iliacas, y producen los plexos capilares y vasos linfáticos de la pared abdominal, pelvis y extremidades inferiores. Se unen a la cisterna del quilo y pierden su conexión con las venas adyacentes.
Con la salvedad de la parte anterior del saco que es origen de la cisterna del quilo, todos los sacos linfáticos experimentan la invasión de células mesenquimatosas y se convierten en grupos de ganglios linfáticos.
El bazo se desarrolla a partir de células mesenquimatosas, entre capas del mesenterio dorsal del estómago. El timo se origina en una protuberancia de la tercera bolsa faríngea (fig. 18.22a).
RESISTENCIA INESPECÍFICA
A ENFERMEDADES
Aunque son varios los mecanismos que contribuyen a dicha resistencia, todos tienen algo en común: brindan protección inmediata contra una amplia gama de microorganismos patógenos y sustancias extrañas. Como lo indica su nombre, tal resistencia carece de respuestas específicas contra invasores también específicos, en vez de lo cual su mecanismo de protección funciona siempre de la misma manera, sin importar el tipo de invasor. Entre dichos mecanismos, se incluyen las barreras físicas y químicas externas que forman la piel y las mucosas, así como varios tipos de defensas inespecíficas internas, entre ellas las proteínas antimicrobianas, células asesinas naturales y fagocitos, inflamación y fiebre.
Primera línea de defensa: piel y mucosas
La piel y las mucosas son la primera línea de defensa del cuerpo humano contra los microbios patógenos. Ambas son barreras físicas y químicas que obstaculizan la penetración en el organismo de dichos microbios y las sustancias extrañas, con las consecuentes enfermedades.
Con sus numerosas capas de células queratinizadas estrechamente apiñadas, la capa epitelial externa de la piel o epidermis, constituye una barrera física excelente contra la entrada de microbios (fig. 5.1). Además, la esfacelación (desprendimiento) periódica de las células epidérmicas ayuda a eliminar bacterias de la superficie cutánea. Las bacterias rara vez penetran en la superficie intacta de la epidermis sana. Sin embargo, la interrupción de la comunidad de la piel por cortaduras, quemaduras o punciones hacen posible que los microorganismos patógenos atraviesen la epidermis e invadan los tejidos adyacentes o circulen con la sangre a otras partes del cuerpo.
La capa epitelial de mucosas, que reviste el interior de cavidades corporales, secreta el líquido llamado moco, que lubrica y humecta la superficie de tales cavidades. Dada su consistencia levemente viscosa, el moco atrapa numerosos microbios y sustancias extrañas. La mucosa de la nariz tiene pelos recubiertos de moco, también para atrapar y filtrar microbios, polvo y contaminantes del aire inhalado. La de las vías respiratorias superiores posee cilios, los cuales son proyecciones microscópicas, parecidas a pelos, de la superficie de las células epiteliales. La acción ondulante de los cilios impulsa el polvo y los microbios inhalados, que están atrapados en el moco, hacia la garganta. La tos y los estornudos aceleran el movimiento del moco.
Otros líquidos que producen diversos órganos también ayudan a proteger la superficie epitelial de la piel y las mucosas. Por ejemplo, el aparato lagrimal de los ojos (fig. 16.4) produce y secreta lagrimas en respuesta a agentes irritantes. El parpadeo distribuye las lágrimas sobre la superficie del globo ocular y la acción de lavado constante de las lágrimas ayuda a diluir microbios y evitar que se asienten en la superficie ocular.
La saliva, producida por las glándulas salivales, barre los microbios de la superficie de los dientes y la mucosa de la boca, de forma similar a la acción de lágrimas en los ojos. El flujo de saliva reduce la invasión de microorganismos en dicha zona.
La limpieza de la uretra con el flujo de orina retarda la colonización microbiana del aparato urinario. De igual modo, las secreciones vaginales desplazan los microbios hacia fuera del cuerpo femenino. La defecación y el vomito son otros procesos útiles para expulsar microorganismos. Por ejemplo, en respuesta a toxinas microbianas que irritan el revestimiento de la porción inferior del tubo digestivo, se contrae con fuerza el músculo liso de ésta y la diarrea consecuente expulsa rápidamente gran parte de los microorganismos.
Ciertas sustancias también contribuyen al alto grado de resistencia de la piel y las mucosas contra la invasión microbiana. Las glándulas sebáceas cutáneas secretan una sustancia aceitosa, el sebo, que forma una película protectora en la superficie; sus ácidos grasos insaturados inhiben el crecimiento de ciertos hongos y bacterias patógenas. La acidez de la piel (pH 3 a 5) se debe en parte a la secreción de ácidos grasos y láctico. El sudor arrastra microbios de la superficie de la piel, además de que contiene la lisozima, enzima que desdobla la pared celular de ciertas bacterias. Dicha enzima también se encuentra en lágrimas, saliva, secreciones nasales y líquidos corporales, y en todos ellos tiene actividad antimicrobiana. El jugo gástrico, que produce las glándulas del estómago, es una mezcla de ácido clorhídrico, enzimas y moco. Su acidez considerable (pH 1.2 a 3.0) destruye muchas bacterias y gran parte de las toxinas bacterianas. Por último, las secreciones vaginales también son levemente ácidas, lo cual inhibe la proliferación bacteriana.
Segunda línea de defensa: defensas internas
Cuando los patógenos logran atravesar las barreras mecánicas y químicas impuestas por la piel y las mucosas, se enfrentan a una segunda línea de defensa: las proteínas antimicrobianas internas, fagocitos, células NK, inflamación y fiebre.
Proteínas antimicrobianas
La sangre y el líquido intersticial contienen tres clases de proteínas antimicrobianas principales que inhiben el crecimiento de los microorganismos: interferones, complemento y transferrinas.
1. Los linfocitos, macrófagos y fibroblastos infectados por virus producen proteínas llamadas interferones o INF. Una vez que las células infectadas liberan los INF, éstos difunden hacia las células vecinas no infectadas, donde inducen la síntesis de proteínas antivirales que interfieren con la replicación de los virus. Aunque los INF no evitan la infección de las células por los virus, sí inhiben la replicación de éstos. Los virus causan enfermedad sólo si son capaces de replicarse en el interior celular. Así, los INF constituyen un importante mecanismo de defensa contra las infecciones por diversos virus. Las tres clases de INF que existen son: inteferón alfa, interferón beta e interferón gamma.
2. Tanto en el plasma sanguíneo como en la membrana plasmática celular, se encuentra un grupo de proteínas inactivas en condiciones normales, que conforman el sistema del complemento.
Cuando estas proteínas se activan, "complementan" o aceleran ciertas reacciones inmunes. El sistema del complementó provoca la citólisis (destrucción) de los microbios, promueve la fagocitosis y contribuye a la respuesta inflamatoria.
3. Las transferrinas son proteínas fijadoras de hierro que inhiben el crecimiento de ciertas bacterias al disminuir la cantidad de hierro disponible para su crecimiento normal.
Fagocitos y células citolíticas naturales (NK)
Cuando los microorganismos atraviesan la piel y las mucosas o sortean las proteínas antimicrobianas presentes en el plasma sanguíneo, el siguiente mecanismo inespecífico de defensa esta constituido por los fagocitos y las células natural killer. Cerca del 5 al 10% de los linfocitos sanguíneos corresponden a células NK. También se hallan presentes en el bazo, ganglios linfáticos y médula ósea roja. Las células NK carecen de las moléculas de membrana que identifica a los linfocitos B y T, pero son capaces de destruir una amplia variedad de células infectadas y ciertas células tumorales. Las células NK atacan a cualquier célula del organismo que exprese en su membrana proteínas anómalas extrañas.
La unión de las células NK a una célula diana, como puede ser una célula infectada, induce la liberación por parte de las células citolíticas naturales, de gránulos cargados de sustancias tóxicas. Algunos gránulos contienen una proteína llamada perforina que se inserta en la membrana plasmática de la célula diana, creando canales (perforaciones) en la membrana. Como resultado, el líquido extracelular ingresa en la célula diana haciendo que ésta se hinche y estalle en un proceso conocido como citólisis. Otros gránulos liberan granzimas, proteínas que actúan como enzimas proteolíticas digestivas que inducen la apoptosis o autodestrucción de la célula diana. Este proceso destruye a las células infectadas, pero no a los microorganismos presentes en su interior; los microorganismos liberados al exterior celular intactos o no, son destruidos por los fagocitos.
Los fagocitos (fago, de phagéin, comer) son células especializadas que llevan a cabo el proceso de fagocitosis, la ingestión de microorganismos u otras partículas, como restos celulares (véase fig. 3-11).
Las principales células fagocíticas son los neutrófilos y macrófagos. Cuando se produce una infección, los neutrófilos y macrófagos migran hacia el área infectada. Durante la migración, los monocitos incrementan su tamaño y se transforman en fagocitos macrófagos activos llamados macrófagos circulantes.
Otros macrófagos, los macrófagos estables, permanecen en tejidos específicos. Entre ellos podemos señalar a los histiocitos (macrófagos del tejido conectivo), células estrelladas reticuloendoteliales, (células de Kupffer) en el hígado, macrófagos alveolares en el pulmón, microglia en el sistema nervioso y macrófagos titulares en el bazo. Además de ser uno de los mecanismos de la defensa innata la fagocitosis cumple un importante papel en la inmunidad ( resistencia específica), como se verá más adelante en este capitulo.
La fagocitosis se lleva a cabo en cinco etapas sucesivas: quimiotaxis, adhesión, ingestión, digestión y destrucción (fig. 22-9).
Quimiotaxis. La fagocitosis se inicia con la quimiotaxis, el movimiento estimulado químicamente de los fagocitos hacia el sitio de lesión. Las sustancias químicas que atraen a los fagocitos pueden provenir de los microorganismos invasores, los glóbulos blancos, las células dañadas de los tejidos o las proteínas activadas del complemento.
Adhesión. La fijación de los fagocitos a los microorganismos o a otras sustancias extrañas se denomina adhesión. La unión de proteínas del complemento a los patógenos facilita esta adhesión.
Ingestión. Luego de la adhesión, la membrana plasmática de los fagocitos emite prolongaciones llamadas seudópodos, los cuales se fusionan, englobando a los microorganismos en una estructura especializada, el fagosoma.
Digestión. El fagosoma ingresa en el citoplasma celular fusionandose con los lisosomas que allí se encuentran y forma una estructura única, de mayor tamaño, el fagolisosoma. El lisosoma aporta lisozima, que rompe las paredes microbianas, y otras enzimas digestivas que degradan hidratos de carbono, proteínas, lípidos y ácidos nucleicos. El fagocito también produce sustancias oxidantes letales como el anión superóxido, el anión hipoclorito y el peróxido de hidrógeno en un proceso denominado estallido oxidativo.
destrucción. El ataque químico mediado por la lisozima, las enzimas digestivas y oxidantes en el interior del fagolisosoma rápidamente destruye varios tipos de patógenos. Cualquier sustancia que no pueda degradarse permanece en la célula, en estructuras denominadas cuerpos residuales.
EVASIÓN MICROBIANA DE LA FAGOCITOSIS
Algunos microorganismos, como las bacterias productoras de neumonía, presentan una estructura extracelular, la cápsula, que evita la adhesión. Esto dificulta físicamente su ingestión por parte de los fagocitos. Otros microorganismos, como las bacterias secretoras de toxinas que provocan un tipo de intoxicación alimentaria, pueden ser digeridas por los fagocitos pero no destruidas; en lugar de esto, las toxinas producidas por estas bacterias (leucocidinas) pueden provocar la destrucción del fagocito al estimular la liberación de sus enzimas lisosómicas al citoplasma. Incluso otros microorganismos, como la bacteria de la tuberculosis, inhibe la fusión del fagosoma con el lisosoma evitando de esta manera la exposición de la bacteria a las enzimas lisosómicas. Estas bacterias, aparentemente, pueden también hacer uso de las sustancias químicas presentes en sus paredes celulares para contrarrestar los efectos de los oxidantes producidos por los fagocitos. La multiplicación microbiana subsiguiente en el interior del fagosoma puede producir la eventual destrucción del fagocito. .
Inflamación
La inflamación es una respuesta defensiva, inespecífica, del organismo ante una lesión tisular. Entre las causas que pueden producir inflamación podemos mencionar a los gérmenes patógenos, las abrasiones, la irritación química, las deformaciones o trastornos celulares y las temperaturas extremas. Los cuatro signos y síntomas que caracterizan al proceso inflamatorio son: rubor, dolor, calor y tumefacción (edema o hinchazón.) La inflamación puede también producir la pérdida de la función del área afectada (por ejemplo la pérdida de la sensibilidad), dependiendo del sitio y de la extensión de la lesión. La inflamación es un intento de eliminar a los microorganismos, toxinas o sustancias extrañas presentes en el sitio de lesión, impedir la diseminación hacia otros tejidos y preparar al sitio de lesión para el proceso de reparación tisular en un intento de restaurar a homeostasis del tejido afectado.
Debido a que la inflamación es uno de los mecanismos de respuesta inespecífica, la respuesta de los tejidos a un corte es similar a la respuesta que se dispara ante una quemadura, radiaciones o invasiones virales o bacterianas. En cada caso, la repuesta inflamatoria consiste en tres pasos básicos: 1) vasodilatación e incremento de la permeabilidad vascular, 2) migración (movilización) de los fagocitos presentes en la circulación y el líquido intersticial y, en última instancia, 3) la reparación tisular.
VASODILATACIÓN Y AUMENTO DE LA PERMEABILIDAD VASCULAR
Los dos cambios inmediatos que se producen en los vasos sanguíneos de la región lesionada son: vasodilatación (aumento del diámetro) de las arterias y aumento de la permeabilidad capilar (fig. 22-10). El aumento de la permeabilidad implica que las sustancias que normalmente están retenidas en la circulación sanguínea puedan salir de los vasos sanguíneos. La vasodilatación incrementa el flujo de sangre hacia el área de la lesión y el aumento en la permeabilidad posibilita que las proteínas de defensa, como los anticuerpos y los factores de la coagulación, salgan del torrente circulatorio y se dirijan al sitio lesionado. El aumento de la permeabilidad también permite remover toxinas microbianas y células muertas.
Entre las sustancias que contribuyen al proceso de la vasodilatación, aumento de la permeabilidad y otros aspectos de la respuesta inflamatoria, se encuentran los siguientes:
.Histamina. En respuesta a la lesión, los mastocitos del tejido conectivo, los basófilos y las plaquetas de la sangre liberan histamina. Los neutrófilos y macrófagos que son atraídos al sitio de lesión, también estimulan la liberación de histamina la cual provoca vasodilatación y aumento de la permeabilidad vascular.
Cininas. Estos polipéptidos se forman en el plasma sanguíneo a partir de precursores inactivos, los cininógenos. Inducen vasodilatación, aumento de la permeabilidad vascular y sirven como agentes quimiotácticos para los fagocitos. Un ejemplo de cinina lo constituye la bradiquinina.
Prostaglandinas (PG). Estos lípidos, en especial los pertenecientes al grupo E, son liberados por las células dañadas potenciando los efectos que la histamina y las cininas ejercen sobre la función endotelial. También estimulan la migración de los fagocitos a través de las paredes capilares.
Leucotrienos (LT). Los LT son producidos por los mastocitos y basófilos, y generan aumento de la permeabilidad vascular, a su vez, participan en la adhesión de los fagocitos a los patógenos y actúan como agentes quimiotácticos de los fagocitos.
Complemento. Los diferentes componentes del sistema del complemento estimulan la liberación de histamina, atraen neutrófilos por quimiotaxis y promueven la fagocitosis; algunos componentes pueden, también, mediar la destrucción bacteriana.
La dilatación arteriolar y el aumento de la permeabilidad capilar producen tres de los síntomas característicos de la inflamación, calor, rubor (eritema) y tumefacción (edema.). El calor y el rubor resultan de la gran cantidad de sangre acumulada en el área lesionada. Como consecuencia de la elevación de la temperatura, las reacciones metabólicas se realizan más rápidamente, liberando calor adicional. El edema se produce como resultado del aumento de la permeabilidad vascular, lo cual permite que una mayor cantidad de líquido se movilice desde el plasma sanguíneo hacia los espacios titulares.
El dolor es un síntoma central de la inflamación. Es el resultado de la lesión de las neuronas y de las sustancias químicas tóxicas liberadas por los microbios. Las cininas afectan algunos de los terminales nerviosos produciendo gran parte del dolor asociado a la inflamación. Las prostaglandinas intensifican y dolor y prolongan el dolor asociado a la inflamación. También puede atribuirse el dolor al aumento de presión tisular provocado por el edema.
El incremento de la permeabilidad capilar deja escapar factores de la coagulación hacia los tejidos. La cascada de la coagulación se pone en marcha, y el fibrinógeno, en última instancia se transforma en una malla insoluble y gruesa de fibrina, que contiene y atrapa a los microorganismos invasores, evitando su diseminación.
MIGRACIÓN DE LOS FAGOCITOS Dentro de la primera hora luego del inicio del proceso inflamatorio, los fagocitos aparecen en escena. A medida que se acumulan grandes cantidades de sangre, los neutrófilos comienzan a adherirse a la superficie interna del endotelio vascular (fig.22.10). Luego, los neutrófilos inician su migración a través de las paredes capilares para alcanzar el sitio de lesión. Este proceso llamado migración, depende de la quimiotaxis. Los neutrófilos intentan llevar a cabo la destrucción de los microorganismos invasores por medio de la fagocitosis. La médula ósea roja produce y libera células adicionales, asegurando un aflujo constante de neutrófilos hacia el área dañada. Este incremento en el número de glóbulos (Ieucocitos) se conoce como leucocitosis.
Si bien los neutrófilos son las células que predominan en las etapas iniciales de un proceso infeccioso, éstos mueren rápidamente. A medida que la respuesta inmunitaria progresa, los monocitos siguen a los neutrófilos en su camino hacia el área afectada. Una vez en el tejido, los monolitos se diferencian a macrófagos circulantes y aumentan la actividad fagocítica de los macrófagos fijos presentes en el lugar. Fieles a su nombre, los macrófagos son fagocitos de mayor potencia que los neutrófilos. Son lo suficientemente grandes como para englobar restos tisulares lesionados, neutrófilos viejos y microorganismos invasores.
Finalmente, los macrófagos también mueren. En pocos días se forma un conjunto de fagocitos muertos y tejido dañado; esta colección de células muertas y líquido se denomina pus. La formación de pus se observa en la mayoría de los procesos inflamatorios y habitualmente continúa hasta que la infección remite. En ocasiones, el pus alcanza la superficie corporal o drena en alguna cavidad interna y se disemina; otras veces persiste aun después de la finalización del proceso infeccioso. En estos casos, la secreción purulenta se elimina en forma gradual, reabsorbiéndose en el transcurso de días.
ABSCESOS Y ÚLCERAS
Si no es posible eliminar el pus del área inflamada, se genera un absceso, una acumulación excesiva de pus en un área limitada. Como ejemplos comunes podemos citar a los granos y a los forúnculos. Cuando el tejido inflamado superficial se desprende de la superficie de un órgano o tejido, el resultado es la formación de una herida abierta llamada úlcera. Las personas que tienen trastornos circulatorios, por ejemplo las personas diabéticas con aterosclerosis avanzada, presentan mayor susceptibilidad a sufrir u1ceraciones del tejido de sus miembros inferiores, en especial las piernas. Estas ulceraciones se denominan úlceras de estasis, y se generan debido a la escasa oxigenación y aporte de nutrientes a los tejidos que los hace más vulnerables incluso a las lesiones y a las infecciones más leves.
FIEBRE
La fiebre es la temperatura corporal anormalmente elevada, producto de la reprogramación del termostato hipotalámico. Por lo general, tiene lugar durante los procesos infecciosos e inflamatorios. Muchas toxinas bacterianas tienen la capacidad de elevar la temperatura a veces la temperatura corporal, a veces provocando la liberación de citocinas productoras de fiebre como la interleucina (ILl) de los macrófagos. La elevación de la temperatura corporal potencia la acción de los interferones, inhibe el crecimiento de algunos microbios e incrementa la velocidad de las reacciones que contribuyen a la reparación de los tejidos.
En el cuadro 22.1 se enumeran los componentes de la respuesta inespecífica.
RESISTENCIA ESPECíFICA: INMUNIDAD
La capacidad del organismo de defenderse de agentes invasores específicos, como las bacterias, las toxinas, los virus y los tejidos extraños, se denomina resistencia específica o inmunidad. Las sustancias que se reconocen como extrañas, capaces de iniciar una respuesta inmunitaria, se denominará antígenos (Ag). Hay dos propiedades que diferencian a la inmunidad de la respuesta inespecífica: 1) especificidad para una molécula extraña en particular (antígenos), lo cual incluye, a su vez, distinguir las moléculas propias del organismo de las ajenas, y 2) memoria para la mayoría de las moléculas con que tomaron contacto, de manera tal que ante un segundo encuentro se pongan en marcha respuestas más rápidas y de mayor intensidad. La rama de la ciencia que se encarga del estudio de las respuestas del organismo ante la presencia de antígenos se llama inmunología (inmuno, de inmunis, libre, exento, y logía, de lógos, estudio). El sistema inmunitario está compuesto por aquellas células y tejidos que se encargan de llevar a cabo la respuesta inmunológica.
Maduración de las células T y de las células B
Las células que desarrollan inmunocompetencia, capacidad de llevar a cabo respuestas inmunológicas al ser estimuladas de forma adecuada, son linfocitos llamados células B y células T. Ambas se originan en los órganos linfáticos primarios (médula ósea roja y timo) a partir de células madre pluripotenciales (sytem cells) están presentes en la médula ósea roja (véase fig.19-3). Las células B completan su maduración en la médula ósea roja, proceso que continúa toda la vida. Las células T se originan de células pre- T que migran desde la médula ósea roja hacia el timo, donde maduran (fig. 22-11). La mayoría de las células T se generan antes de la pubertad, pero continúan su maduración y dejan el timo durante la vida.
Antes que las células T salgan del timo o que las células B dejen la médula ósea, comienzan a sintetizar gran cantidad de proteínas específicas que se insertan en sus membranas plasmáticas. Algunas de estas proteínas funcionan como receptores antigénicos, moléculas capaces de reconocer antígenos específicos (fig. 22-11). Las células T salen del timo como células CD4 o CD8, lo cual significa que además de sus receptores antigénicos, su membrana plasmática presenta proteínas llamadas CD4 o CD8. Como se verá más adelante, en este capitulo, estos dos tipos de células T tiene funciones muy diferentes.
Tipos de respuesta inmunitaria
La inmunidad consiste en dos clases de respuestas muy relacionadas, ambas producto de la presencia de antígenos. En la respuesta inmunitaria mediada por células (inmunidad celular), CD8 proliferan a células T citotóxicas que atacan directamente al antígeno invasor. En la respuesta inmunitaria mediada por anticuerpos (inmunidad humoral), las células B se diferencian a células plasmáticas (plasmocitos), las cuales sintetizan y secretan un tipo de proteínas específicas llamadas anticuerpos (Ac) o inmunoglobulinas. Un determinado anticuerpo puede unirse e inactivar a un antígeno específico. La mayoría de las células T CD4 se transfoman en células colaboradoras (T helpers) que asisten tanto en la respuesta inmunitaria celular como en la respuesta inmunológica mediada por anticuerpos.
La inmunidad mediada por células es particularmente efectiva contra: 1) patógenos intracelulares, entre los que se incluyen los virus, bacterias u hongos que se encuentran en el interior de las células; 2) algunas células tumorales; y 3) tejidos extraños provenientes de trasplantes. De esta manera, la inmunidad celular siempre involucra el ataque de células contra células. En la inmunidad mediada por anticuerpos, la respuesta se produce contra: 1) antígenos presentes en los líquidos corporales y 2) patógenos extracelulares, entre los que se encuentra cualquier virus, bacterias u hongos localizados fuera de las células. Dependiendo de su localización, un patógeno en particular puede generar ambos tipos de respuesta inmunitaria.
Antígenos y receptores antigénicos
Los antígenos presentan dos grandes características: inmunogenicidad y reactividad. La inmunogenicidad es la capacidad de producir una respuesta inmunitaria estimulando la producción de anticuerpos específicos, la proliferación de células T específicas o ambas. El término antígeno deriva de su función como generadora de anticuerpos. Reactividad es la capacidad que tienen los antígenos de reaccionar específicamente con los anticuerpos o células que estos estimularon. Estrictamente hablando, los inmunólogos definen a los antígenos como aquellas sustancias que presentan reactividad y las sustancias que presentan tanto inmunogenicidad como reactividad son consideradas antígenos completos (inmunógenos). Sin embargo, el término antígeno se suele aplicar a sustancias inmunogénas y reactivas, y éste será el criterio de utilización del termino.
Los microbios enteros o parte de ellos pueden actuar como antígenos. Los componentes químicos de las estructuras bacterianas como los flagelos, la cápsula y las paredes celulares son antigénicas así como también lo son las toxinas bacterianas. Como ejemplo de sustancias no microbianas que funcionan como antígenos podemos citar a los componentes del polen, la clara de huevo, células sanguíneas incompatibles y tejidos u órganos transplantados. La gran variedad de antígenos presentes en el medio ambiente provee una miríada de posibilidades para que se desencadenen respuestas inmunitarias. Es habitual que sólo pequeñas porciones de moléculas antigénicas de mayor tamaño actúen como disparadores de las respuestas inmunitarias. Estas pequeñas porciones se denominan epítopos, o determinantes antigénicos (fig. 22-12). La mayoría de los antígenos presenta varios epítopos, cada uno de los cuales induce la producción de un anticuerpo en particular o activa una célula T específica.
Los antígenos que sobreviven a los mecanismos de defensa inespecíficos, en general siguen uno de los tres caminos posibles dentro de los tejidos linfáticos: 1) la mayoría de los antígenos que ingresan al torrente sanguíneo (por ejemplo, a través de algún vaso sanguíneo lesionado) quedan atrapados a medida que circulan por el bazo, 2) aquellos antígenos que penetran la piel ingresan a los vasos linfáticos y quedan alojados en los ganglios linfáticos y 3) los antígenos que atraviesan las membranas mucosas quedan atrapados en el tejido linfático asociado a las mucosas (MALT).
Naturaleza química de los antígenos
Los antígenos son moléculas grandes y complejas. Por lo general son proteínas. Sin embargo también pueden actuar como antígenos los ácidos nucleicos, lipoproteínas, glucoproteínas y ciertos polisacáridos de gran tamaño. Las células T sólo responden a antígenos proteicos; las células B, en cambio, responden a antígenos proteicos, ciertos lípidos, hidrato s de carbono y ácidos nucleicos. Los antígenos completos tienen por lo general elevado peso molecular, de 10 000 o más daltons, aunque moléculas grandes compuestas de unidades repetitivas simples por ejemplo, la celulosa y muchos plásticos en general no son antigénicas. De esta forma, los materiales plásticos pueden utilizarse en válvulas cardiacas artificiales y prótesis articulares.
Una sustancia de menor peso, que presenta reactividad pero carece de inmunogenicidad, se denomina hapteno (= captar). Un hapteno puede estimular la respuesta inmunitaria sólo si se presenta unido a una molécula transportadora de mayor tamaño. Un ejemplo es la pequeña toxina lipídica de la hiedra venenosa, la cual inicia una respuesta inmunitaria una vez que se une a alguna proteína del organismo. De igual manera, algunos fármacos, como la penicilina, pueden combinarse con proteínas del cuerpo para formar complejos inmunes. La respuesta inmune que se pone en marcha ante la presencia de haptenos es responsable de algunas reacciones alérgicas hacia ciertos fármacos y otras sustancias que se hallan presentes en el ambiente.
Como regla, los antígenos son sustancias extrañas; por lo general no son partículas que formen parte de los tejidos corporales. Sin embargo, alguna veces el sistema inmunitaria falla en la distinción entre "amigos" (propio) y "enemigos" (ajeno). El resultado es un trastorno autoinmune en el cual las moléculas propias o las células se reconocen como extrañas y, por lo tanto, son destruidas como tales.
Diversidad de los receptores antigénicos
Una característica sorprendente que presenta el sistema inmunitario es la capacidad de reconocer y ligar hasta mil millones (109) de epítopos diferentes. Aún antes de que se produzca el ingreso de un antígeno en particular, las células T y células B capaces de reconocer dicho antígeno y responder ante él están preparadas y esperando. Las células del sistema inmunitario pueden incluso reconocer moléculas generadas artificialmente que no existen en la naturaleza. Esta capacidad de reconocer tantos epítopos se basa en una igualmente amplia variedad de receptores antigénicos presentes. Dado que las células humanas contienen sólo alrededor' de 35 000 genes, ¿cómo es posible que se generen mil millones o más de receptores antigénicos?
La respuesta a este dilema resultó ser simple en cuanto a su concepto. La variedad de receptores de las células B y las células T es el resultado de la mezcla y del re acomodamiento de unos cientos de versiones de una gran cantidad de segmentos génicos. Este proceso se denomina recombinación genética. Los segmentos génicos se organizan en diferentes combinaciones a medida que los linfocitos se desarrollan partir de las células madre en la médula ósea roja y el timo. Esta situación es similar a mezclar un mazo de 52 naipes y repartir luego tres cartas. Si esto se repite una y otra vez, se generarían muchas más de 52 combinaciones diferentes de tres cartas. Como resultado de la recombinación genética, cada célula B o T tiene un único juego de segmentos génicos que codifican para un receptor antigénico único. Luego de la transcripción y de la traslación, las moléculas del receptor se incorporan a la membrana plasmática.
Antígenos del complejo mayor de histocompatibilidad
En la membrana plasmática de las células del organismo se encuentran "antígenos propios", los antígenos del complejo mayor de histocompatibilidad (CMH.) Estas glucoproteínas de membrana también se conocen con el nombre de antígenos leucocitaris , (human leukocyte antigens, HLA) debido a que se identificaron por primera vez sobre los glóbulos blancos. A menos que una persona tenga un hermano gemelo idéntico, los antígenos del CMH son únicos. De miles a cientos de miles de moléculas del CMH están presentes en la superficie de cada una de las células del cuerpo a excepción de los glóbulos rojos. Aunque los antígenos del CMH son los responsables del rechazo de tejidos cuando se trasplantan de una persona a otra, su función normal es la de asistir a las células T en el reconocimiento de antígenos extraños, no propios. Tal reconocimiento es un importante primer paso en cualquier respuesta inmunitaria.
Existen dos tipos de antígenos del complejo mayor de histocompatibilidad, clase I y clase II. Las moléculas del CMH clase I, (CMH-I) se expresan en las membranas plasmáticas de todas las células, excepto en los glóbulos rojos. Las moléculas del CMH clase II (CMH-II) están presentes en la membrana de las célula doras de antígenos (se describe en la próxima sección).
Vías de procesamiento antigénico
Para que se lleve a cabo una respuesta inmunitaria, las células B y las células T deben reconocer la presencia de antígenos extraños. Las células B reconocen y se unen a los antígenos que están presentes en la linfa, líquido intersticial o plasma sanguíneo. Las células T reconocen fragmentos de proteínas antigénicas que se presentan y procesan de una cierta manera. En el procesamiento antigénico, las proteínas antigénicas se fragmentan en péptidos de menor tamaño que luego se asocian a las moléculas del CMH. A continuación, el complejo antigeno CMH se inserta en la membrana plasmática de una célula del órganismo. La inserción de este complejo se denomina presentación antigénica. Cuando el fragmento peptídico proviene de una proteína propia, las células T ignoran al complejo antígenoCMH. Sin embargo si el fragmento peptídico proviene de una proteína extraña, las células T reconocen al complejo antígeno CMH como extraño, y se pone en marcha la respuesta inmunitaria. El procesamiento y la presentación antigénica se lleva a cabo de dos maneras, dependiendo de si el antigeno se localiza en el interior o en el exterior de las células del organismo.
Procesamiento de antígenos exógenos
Los antígenos foráneos que se encuentran en el líquido extracelular se conocen como antígenos exógenos. Éstos comprenden bacterias. Y toxinas bacterianas, gusanos, parásitos, polvos y polen inhalado y virus que aún no infectaron a las células, Una clase especial de células, las células presentadoras de antígenos (CPA), procesan y presentan a los antígenos exógenos. Las CPA comprenden a las células dendríticas, macrófagos, y células B. Se localizan estratégicamente en aquellos sitios donde es más probable que los antigénos vulneren los mecanismos de defensa inespecífica permitiendo su ingreso al organismo, como la epidermis y dermis de la piel (las células de Langerhans constituyen un tipo de célula dendrítica); las mucosas que recubren el tracto respiratorio, digestivo, urinario y reproductor y los ganglios linfáticos. Después del procesamiento y de la presentación antigénica, las CPA migran desde los tejidos linfáticos por vía linfática, hacia los ganglios linfáticos.
El procesamiento y la presentación antigénica se lleva a cabo en distintos pasos, que ocurren de la siguiente manera (fig. 22-13).
1.- Internalización de los antígeno. La internalización de los antigénos en las células presentadoras de antígenos se realiza por fagocitosis o endocitosis. Se lleva a cabo en casi cualquier sector del organismo donde los intrusos, como los microbios, traspasaron los mecanismos de defensa inespecíficos.
2.- Digestión de los antígenos en fragmentos de péptidos. En el interior del fagosoma o del endosoma, las enzimas digestivas rompen las grandes proteínas antigénicas y forman pequeños fragmentos peptídicos.
3.- Síntesis de moléculas del CMH-Il. Al mismo tiempo, las CPA sintetizan moléculas del CMH-II y las empaquetan en vesículas.
4.- Fusión de vesículas. Las vesículas que contienen los fragmentos de los péptidos antigénicos y las vesículas con las moléculas del CMH-ll se unen y se fusionan.
5.- Unión de los fragmentos peptídicos a las moléculas del CMH-II. Luego de la fusión de ambos tipos de vesículas, los fragmentos de los péptidos antigénicos se asocian a las moléculas del CMH-II.
6.-Inserción del complejo antígeno CMH-Il en la membrana plasmática. La vesícula que contiene los complejos antígenoCMH-II sufre exocitosis. Como resultado de ellos, los complejos antígeno CMH-II se insertan en la membrana plasmática.
Una vez realizado el procesamiento antigénico, las células presentadoras de antígenos migran a los tejidos linfáticos para presentar los antígenos a las células T. En los tejidos linfáticos, un pequeño número de células T, con receptores antigénicos cuya forma es complementaria de los antígenos presentados, reconoce y se une a los complejos antígenoCMH-II iniciando la respuesta inmunitaria mediada por células o la respuesta mediada por anticuerpos. La presentación de antígenos exógenos junto a las moléculas del CMH-II, mediada por las células presentadoras de antígenos, permite informar a las células T que hay un intruso en el cuerpo, y que debe ponerse en marcha una acción defensiva. .
Procesamiento de antígenos endógenos
Los antígenos extraños que se sintetizan en el interior de las células del cuerpo se denominan antígenos endógenos. Tales antígenos pueden ser proteínas virales generadas una vez que los virus infectan a las células utilizando la maquinaria metabólica (biosintética) de la célula huésped o proteínas anormales sintetizadas por células tumorales. Los fragmentos de los antígenos endógenos se asocian a moléculas del complejo mayor de histocompatibilidad, en el interior de la célula infectada. El resultado es la formación de complejos fragmento antigénico endógeno CMH-I, los cuales se movilizan hacia la membrana plasmática, donde se expresan en la superficie celular. La participación de antígenos endógenos unidos a moléculas del CMH-señaliza la presencia de células infectadas que necesitan ayuda.
Citocinas
Las citocinas son pequeñas hormonas proteicas encargadas de estimular o inhibir varias funciones celulares normales, como el crecimiento y la diferenciación celular. Los linfocitos y las células presentadoras de antígenos secretan citocinas, como también lo hacen los fibroblastos, las células endoteliales, los monocitos, los hepatocitos y las células renales. Algunas citocinas estimulan la proliferación de las células progenitoras presentes en la médula ósea roja. Otras regulan las actividades de las células involucradas en los mecanismos de defensa inespecíficos o en la respuesta inmunitaria, tal como se describe en el cuadro 22-2.
TERAPIA CON CITOCINAS
La terapia con citocinas implica la utilización de estas sustancias en el tratamiento de diferentes patologías. Los interferones fueron las primeras citocinas que demostraron tener efectos limitados contra algunos tipos de cánceres humanos. El interferón alfa (Intron A) está aprobado en los Estados Unidos para el tratamiento del sarcoma de Kaposi, un tipo de cáncer muy frecuente en pacientes infectados con HIV, el virus causante del SIDA. También está aprobado el uso del interferón alfa en el tratamiento del herpes genital causado por el herpes virus; el tratamiento de la hepatitis B y C, producidas por los virus de la hepatitis B y C, respectivamente; y el tratamiento de las leucemias de células pilosas. Una forma de interferón beta (Betaseron) disminuye la progresión de la esclerosis múltiple (EM) así como la frecuencia y gravedad de sus ataques. De las interleucinas, la más ampliamente utilizada en la terapia contra el cáncer es la interleucina 2. Si bien este tratamiento es efectivo pues causar la regresión del tumor en algunos pacientes, también puede ser muy tóxico. Entre los efectos adversos pueden citarse la fiebre levada, debilidad marcada y dificultad respiratoria debido a endema pulmonar e hipotensión que puede culminar en shock.
Activación, proliferación y diferenciación de las células T
En un momento determinado, las células T se encuentran en estado inactivo. Como se ha visto en la sección anterior, los receptores antigénicos presentes en la superficie de las células T, llamados receptores de células T (RCT, T-cell receptors), reconocen y se unen específicamente a los fragmentos antigénicos extraños que son presentados como complejos antígeno CMH. Hay millones de células T diferentes; cada una tiene un RCT único que reconoce un complejo antigéno CMH específico. Cuando un antígeno ingresa al organismo,solo un bajo número de células T expresan los RCT que son capaces de reconocer y ligarse al antígeno. El reconocimiento antigénico también involucra otras proteínas de superficie presentes en las células T, las proteínas CD4 o CD8. Estas proteínas interactúan con los antigenos del CMH y ayudan a mantener la unión entre RCT-CMH. Por esta razón se denominan correceptores. El reconocimiento antigénico por medio de los RCT junto con las proteínas CD4 o CD8 es la primera señal en la activación de las células T.
Una célula T se activa sólo si se une al antígeno extraño, y al mismo tiempo recibe una segunda señal, en un proceso denominado coestimulación. Existen más de 20 coestimuladores, algunos de los cuales son citocinas, como la interleucina 2. Otro de los coestimuladores está constituido por un par de proteínas de membrana, una situada en la superficie de la célula T y la segunda en la membrana de la célula presentadora de antígenos, lo que permite que ambas células se adhieran una a otra por más tiempo.
La necesidad de dos señales se asemeja un poco al arranque y manejote un auto: cuando se introduce la llave correcta (antígeno) en el contacto (RCT) y se gira, el auto avanza (reconocimiento del antígeno específico), pero no lo hace hasta que la caja de cambios sea puesta en primera marcha (coestimulación). La necesidad de la coestimulación puede impedir que la respuesta inmunológica se ponga en marcha accidentalmente. Diversos coestimuladores pueden afectar la función de las células T de maneras diferentes, de la misma manera que poner reversa en un auto tiene un efecto diferente que poner primera. Más aún, el reconocimiento (unión del antígeno al receptor) sin la coestimulación lleva a un prolongado estado de inactividad llamado energía, tanto en células B como en células T. La energía es más o menos como ¡dejar el auto en punto muerto (neutral) con el motor encendido hasta que se consuma todo el combustible!
Una vez que la célula T recibió estas dos señales (reconocimiento antigénico y coestimulación), se activa. Una célula T activada aumenta de tamaño, comienza a proliferar (se divide varias veces) y se diferencia (forma células más especializadas.) El resultado es la formación de una población de células idénticas que se denomina clon, el cual puede reconocer el mismo antígeno específico. Antes de la primera exposición a un antígeno dado, sólo unas pocas células T pueden ser capaces de reconocerlo, pero una vez que la respuesta inmunitaria ha comenzado, hay miles de células T capaces de reconocer al antígeno en cuestión. La activación, proliferación y diferenciación de las células T es un proceso que tiene lugar en los tejidos y órganos linfáticos secundarios. Es muy probable que la inflamación de las amígdalas o de los ganglios linfáticos del cuello esté causada por la proliferación de los linfocitos involucrados en la respuesta inmunitaria.
Tipos de células T
Los tres tipos principales de células T diferenciadas son las células T colaboradoras (helper), las células T citotóxicas y las células T de memoria.
Células T colaboradoras (helper)
La mayoría de las células T que presentan la proteína CD4 en su superficie, se diferencian a células T helper, también conocidas como células T CD4. Las células T helper inactivas (en reposo) reconocen fragmentos de antígenos exógenos en asociación a moléculas del complejo mayor de histocompatibilidad clase II (CMH-II) en la superficie de una CPA (fig. 22-14a). Con ayuda de la proteína CD4, la célula T helper y la CPA interactúan una con la otra (reconocimiento antigénico), se lleva a cabo la coestimulación y la célula T helper se activa.
Horas después de que se produjo la coestimulación, las células T helper activadas comienzan a secretar varias citocinas (véase cuadro 22-2). Diferentes subgrupos de células T helper se especializan en la producción de tipos particulares de citocinas. Una citocina de gran importancia, que es producida por las células T helper, es la interleucina2 (IL-2), la cual es necesaria para la casi totalidad de las respuestas inmunitarias además de ser el disparador primordial en la proliferación de las células T. La IL-2 actúa como un coestimulador para las células T helper o células T citotóxicas que se encuentran en reposo, y potencia y acelera la activación y proliferación de las células T, células B y células NK.
Algunas de las acciones que ejerce la interleucina-2 constituyen un ejemplo de sistema de retroalimentación positiva beneficiosa. Como se mencionó, la activación de las células T helper las estimula a iniciar la secreción de IL-2, la cual después actúa de manera autocrina uniéndose a los receptores para IL-2, presentes en la membrana plasmática de las células que la secretaron inicialmente. Uno de los efectos es la estimulación de la división celular. A medida que las células T colaboradoras proliferan, tiene luego un mecanismo de retroalimentación positiva ya que se secretan mayores cantidades de IL-2, que inducen un mayor incremento en la tasa de proliferación celular. La IL-2 también ejerce sus efectos en forma paracrina al unirse a los receptores para IL-2 de células T colaboradoras vecinas, células T citotóxicas o células B. Si cualquiera de estas células ya reconoció algún antígeno, la IL-2 actúa como señal coestimuladora.
Células T citotóxicas
Las células T que presentan la molécula CD8 en su estructura, se transforman en células T citotóxicas, también llamadas células CD8. Las células T citotóxicas reconocen antígenos extraños en combinación con el complejo mayor de histocompatibilidad de clase I (CMH-I) sobre las superficie de: 1) células del organismo infectadas por microbios, 2) algunas células tumorales y 3) células provenientes de trasplantes de tejidos (fig. 22-14b). El reconocimiento implica la unión del RCT y la proteína CD8 al CMH-I. Seguido al reconocimiento antigénico, se lleva a cabo la coestimulación. Para que la célula T citotóxica se active, se necesita la coestimulación provista por la IL-2 o alguna otra citocina producida por las células T colaboradoras. (Recuerde que las células T helper requieren para su activación la presencia del complejo antígeno-CMH-II así máxima activación de las células T citotóxicas implica la presentación de antígenos asociados tanto a moléculas del CMH-I como del CMH-II.
Células T de memoria
Las células T restantes de la proliferación clonal llevada a cabo por la respuesta de inmunidad celular se denominan células T de memoria. Cuando un patógeno reingresa en el organismo llevando el mismo antígeno que provocó la respuesta inmunitaria por primera vez, miles de células de memoria se encuentran disponibles para comenzar una reacción mucho más veloz que la producida durante la primera invasión. La segunda respuesta es, por lo general, tan rápida y tan intensa que los patógenos son destruidos antes de que pueda aparecer algún signo o síntoma.
Eliminación de invasores
Las células T citotóxicas, en la respuesta inmunológica mediada por células, se comportan como verdaderos soldados que marchan hacia el frente de batalla cuando se hacen presentes los invasores. Éstas abandonan los tejidos y órganos linfáticos secundarios y migran en busca de las células diana infectadas, células cancerosas, y células provenientes de trasplantes, para su destrucción (fig, 22-15). Las células T citotóxicas reconocen y se unen a las células diana. Luego, las células T citotóxicas asistan el "golpe letal" que las elimina.
Las células T citotóxicas se encargan de la eliminación de cualquier célula del organismo que se encuentre infectada, de manera muy similar a como lo hacen las células NK. La diferencia principal entre ambas es que las células T citotóxicas tienen receptores específicos para un microorganismo en particular, de manera tal que son capaces de eliminar sólo a aquellas células diana infectadas con un tipo en particular de microorganismos; por su parte, las células NK son capaces de destruir una amplia variedad de células del organismo infectadas por microbios. Las células T citotóxicas tienen dos mecanismos principales por los cuales eliminan a las células infectadas.
1. Las células T citotóxicas, a través de sus receptores de superficie, reconocen y unen a las células diana que presentan antígenos microbianos en su superficie. Las células T citotóxicas, entonces, secretan granzimas, enzimas proteolíticas que disparan el mecanismo de apoptosis (fig. 22-15a). Una vez que la célula infectada se destruye, los microorganismos liberados son eliminados por los fagocitos.
2. De manera alternativa, las células T citotóxicas se unen a las células infectadas y liberan dos proteínas desde sus gránulos: perforina y granulisina. La perforina se inserta en la membrana plasmática de la célula diana creando canales en la membrana (fig. 22-15b). Como resultado de ello, el líquido extracelular fluye hacia el interior de la célula diana produciéndose la citólisis (estallido celular) de aquella. Otros gránulos de las células T citotóxica liberan granulisina, la cual ingresa a la célula través de canales y provoca la destrucción de los microorganismos al perforar sus membranas. Las células T citotóxicas también pueden destruir a las células infectadas a partir de la liberación de una molécula tóxica, la linfotoxina, regulada por la activación enzimática de la célula diana. Estas enzimas provocan la fragmentación del ADN de la célula infectada lo que la lleva a su muerte. Además, las células T citotóxicas secretan interferón gamma, que atrae a las células fagocíticas activadas y factor inhibidor de la migración de macrófagos, lo cual evita la migración de los fagocitos desde el sitio de infección. Después de separarse de la célula diana, las células T citotóxicas pueden ir en busca de otras células diana para destruirlas.
Vigilancia inmunitaria
Cuando una célula normal se transforma en una célula cancerosa, a menudo expone moléculas nuevas en su superficie denominadas antígenos tumorales. Estas moléculas raramente, si es que lo hacen alguna vez, se expresan en la superficie de las células normales. Si el sistema inmunológico reconoce un antígeno tumoral como ajeno (extraño), puede entonces, destruir a aquellas células cancerosas portadoras de dicho antígeno. Tal respuesta inmunológica se conoce como vigilancia inmunitaria, y es llevada a cabo por los linfocitos T citotóxicos, macrófagos y células NK. La vigilancia inmunitaria es más eficaz en la eliminación de células tumorales transformadas por acción de virus oncogénicos (productores de cáncer). Es por esta razón que los pacientes receptores de trasplantes, bajo tratamiento con fármacos inmunosupresores para impedir el rechazo de trasplantes, presentan mayor incidencia de cánceres inducidos por virus. El riesgo de padecer cánceres de otros tipos no se ve incrementado.
Rechazo de injertos y tipificación de tejidos
El trasplante de órganos se describe como el reemplazo de un órgano lesionado o enfermo, tal como corazón. hígado, riñones, pulmones o páncreas, con órganos provenientes (donados) por otro individuo. Frecuentemente, el sistema inmunitario reconoce a las proteínas en el Órgano trasplantado como extrañas y de esta forma se ponen en marcha las respuestas inmunitarias celular y humoral contra dichas proteínas. Este fenómeno se conoce como rechazo de injertos.
El éxito de un trasplante de tejidos u órganos depende de la histocompatibilidad (la compatibilidad tisular entre el donante y el receptor). Cuanto más similares son los antígenos del CMH, mayor es la histocompatibilidad, y por lo tanto, menor es la posibilidad de rechazo del trasplante. La tipificación tisular (estudio de histocompatibilidad) se realiza antes de cualquier trasplante de órganos. En los Estados Unidos, se cuenta con un registro nacional computarizado que ayuda a los médicos en la selección de los candidatos para recibir un transplante mas histocompatibles y mas urgentes cada vez que se dispone de un donante. Cuanta más concordancia haya entre las proteínas del complejo mayor de histocompatibilidad del donante receptor, más débil será la respuesta de rechazo de injertos.
Para reducir el riesgo de rechazo de injertos, los receptores de trasplantes de órganos reciben tratamiento con fármacos inmunodepresores. Una de las drogas que se utiliza es la ciclosporina, un derivado de un hongo, que inhibe la secreción de interleucina-2 por parte de las células T helper, pero que ejerce sólo efectos mínimos sobre los linfocitos B. Así se reduce el riesgo de rechazo mientras se mantiene la resistencia a algunas enfermedades.
INMUNIDAD MEDIADA POR ANTICUERPOS
En nuestro organismo no sólo existen millones de células T diferentes sino también millones de células B diferentes, cada una de ellas capaz de responder frente a un antígeno específico. Las células
T citotóxicas abandonan los tejidos (linfáticos en busca de antigénos extraños para su destrucción, pero las células B permanecen en su lugar. Ante la presencia de un antígeno específico, las células B específicas que se encuentran en los ganglios linfáticos, bazo y tejido linfático asociado a las mucosas se activan. Se diferencian a células plasmáticas, las cuales secretan anticuerpos específicos que a su vez circulan por la linfa y por el torrente sanguíneo el plasma hará llegar al sitio de invasión.
Activación, proliferación y diferenciación de células B
Durante la activación de las células B, los antígenos se unen al receptor de las células B (RCB) (fig. 22-16). Estas proteínas integrales de transmembrana son químicamente similares a los anticuerpos que finalmente secretan las células plasmáticas. Aunque las células B pueden responder a un antígeno sin procesar presentes en la linfa o en el líquido intersticial, su respuesta es mucho más intensa cuando ellas procesan los antígenos. El procesamiento antigénico que se lleva a cabo en las células B ocurre de la siguiente manera: el antígeno ingresa en la célula B, se separa en fragmentos peptídicos que se combinan con los antígenos propios del CMH-Il, que se translocan hacia la membrana del linfocito B. Las células T helper reconocen a los complejos antígenos-CMH-Il y proveen la coestimulación necesaria para la proliferación y diferenciación de las células B. Las células T helper producen interleucina-2 y otras citocinas que actúan como moléculas coestimuladoras en la activación de las células B. Las células T, asimismo, producen interleucina-4 e interleucina-6, que facilitan la proliferación de las células B, su diferenciación en células plasmáticas y la secreción de anticuerpos por parte de éstas.
Algunas de las células B activadas aumentan de tamaño, se dividen y diferencian en un clon de células plasmáticas (plasmocitos) productoras de anticuerpos. Unos días después de la exposición al antígeno, las células plasmáticas comienzan a secretar cientos de millones de anticuerpos por día por un lapso de aproximadamente 4 o 5 días, hasta que las células plasmáticas mueren. La mayoría de los anticuerpos viajan a través de la linfa y de la sangre hacia el sitio de invasión. Algunas de las células B activadas no se diferencian a plasmocitos, sino que permanecen como células B de memoria, las cuales se encuentran preparadas para responder más rápidamente y con mayor intensidad, en el caso de que el mismo antígeno reaparezca en el futuro.
Los diferentes antígenos estimulan diferentes células B, las cuales se transforman en células plasmáticas y sus respectivas células B de memoria. Todas las células B de un mismo clon secretan sólo un tipo específico de anticuerpos, los cuales son idénticos al receptor antigénico expresado por la célula B que inicialmente respondió al antígeno. Cada antígeno específico sólo activa a aquellas células B que están predestinadas (por la combinación de segmentos génicos que presenta) a secretar anticuerpos específicos para dicho antígeno. Los anticuerpos producidos por un clon de células plasmáticas ingresan a la circulación, donde forman complejos antígeno anticuerpo con el antígeno que inició su producción.
Anticuerpos
Un anticuerpo (Ac) es capaz de combinarse específicamente con el epítopo del antígeno que estimuló su producción. La estructura de los anticuerpos concuerda con su antígeno, de la misma forma que lo hace una cerradura con su llave. En teoría, las células plasmáticas son capaces de secretar tantos anticuerpos diferentes como receptores de células B diferentes haya, debido a que la misma recombinación de segmentos génicos codifica tanto para los RCB como para los anticuerpos que finalmente secretan las células plasmáticas.
Estructura de los anticuerpos
Los anticuerpos pertenecen a un grupo de glucoproteínas llamadas globulinas y por esta razón se conocen también como inmunoglobulinas (Ig). La mayoría de los anticuerpos contienen cuatro cadenas polipeptídicas (fig. 22-17). Dos de estas cadenas son idénticas entre sí y se denominan cadenas pesadas (H); cada una consiste en una cadena de 450 aminoácidos. Cada cadena polipeptídica presenta ramificaciones constituidas por pequeñas cadenas de hidratos de carbono. Las otras dos cadenas polipeptídicas, también idénticas entre sí, se denominan cadenas livianas (L), y están formadas por unos 220 aminoácidos cada una. Un puente disulfuro (S-S) une cada cadena liviana con una cadena pesada. A su vez, dos puentes disulfuro mantienen unida la porción central de ambas cadenas pesadas; esta región de los anticuerpos que presenta gran flexibilidad, es la llamada región bisagra. Debido a que los "brazos" de los anticuerpos pueden moverse a medida que la región bisagra se dobla, éstos puede presentar forma de T (fig. 22-17a) o forma de Y (fig. 2217b). Más allá de la región bisagra, una parte de las cadenas pesadas forma la región constante de la inmunoglobulina.
Dentro de las cadenas H y L se diferencian dos regiones. Los extremos de las cadenas H y L, denominados regiones variables (V), constituyen los sitios de reconocimiento antigénico. La región variable, la cual es diferente para cada clase de anticuerpo, es la porción del anticuerpo que se encarga de reconocer y unir específicamente a un tipo en particular de antígeno. Debido a que la mayoría de los anticuerpos presentan dos sitios de unión para antígenos, se dice que son bivalentes. La flexibilidad de la región bisagra de los anticuerpos permite que éstos se unan en forma simultánea a dos epítopos distanciados entre sí, por ejemplo, sobre la superficie de un microorganismo.
El resto de las cadenas H y L, llamadas regiones constantes (C), es prácticamente la misma para todos los anticuerpos de la misma clase y es la responsable de la reacción antígeno anticuerpo que se lleva a cabo. Sin embargo, la región constante de la cadena H difiere de una clase de anticuerpo a otra, y es su estructura la que sirve como base para la distinción de cinco clases de anticuerpos diferentes, los cuales se designan IgG. 19A. IgM. IgD e IgE. Cada clase de anticuerpo presenta una estructura química distintiva y un papel biológico específico. Debido a que aparecen primero y presentan una vida media relativamente corta, los anticuerpos de tipo IgM son indicadores de infección reciente. En un paciente enfermo la presencia de un determinado patógeno puede establecerse mediante la detección de anticuerpos IgM específicos para un microorganismo en particular. La resistencia a las infecciones del feto y de los recién nacidos se debe principalmente a los anticuerpos IgG maternos, los cuales son capaces de atravesar la placenta antes del nacimiento, y a la IgA que se secreta por leche materna después del nacimiento. En el cuadro 22-3 se resume las estructuras y funciones de las cinco clases de anticuerpos.
Acciones de los anticuerpos
Las cinco clases de anticuerpos difieren en cuanto a sus funciones; sin embargo, todos ellos actúan inactivando, de alguna manera, a los antígenos. Entre las acciones de los anticuerpos podemos citar las siguientes:
.Neutralización de los antígenos. La reacción entre un antígeno y un anticuerpo bloquea o neutraliza algunas toxinas bacterianas y evita la adhesión de algunos virus a las células diana.
Inmovilización bacteriana. Ya que los anticuerpos se generan por la presencia de antígenos presentes en los cilios o flagelos de las bacterias móviles, la reacción antígeno anticuerpo puede
causar en la bacteria la pérdida de la motilidad, lo que limitaría su diseminación hacia los tejidos vecinos.
Aglutinación y precipitación de antígenos. Debido a que los anticuerpos presentan dos o más sitios de unión para los antígenos, la reacción antígeno anticuerpo puede realizarse en forma cruzada entre distintos patógenos, produciendo la aglutinación (agrupamiento) de éstos. Las células fagocíticas digieren a los microbios aglutinados con más rapidez. De la misma forma, los antígenos solubles secretados pueden precipitar al reaccionar en forma cruzada con los anticuerpos, y son más fácilmente fagocitados.
Activación del complemento. Los complejos antígeno-anticuerpo ponen en marcha la activación de la vía clásica del sistema del complemento (se examinará en breve.)
Facilitación de la fagocitosis. El eje de los anticuerpos actúa como una "bandera" que atrae a los fagocitos, una vez que los antígenos se unieron a la región variable de los anticuerpos. Éstos facilitan la actividad de los fagocitos debido a que producen aglutinación y precipitación de los antígenos, activación del sistema del complemento y mediante la cobertura de los microorganismos con anticuerpos (opsonización), lo que los hace más susceptibles a la fagocitosis.
Anticuerpos monoclonales
Los anticuerpos producidos por las células plasmáticas contra un antígeno dado pueden obtenerse de la sangre de un individuo. Sin embargo y dado que un antígeno típico presenta diversos epítopos, son varios los clones de células plasmáticas que producen anticuerpos contra dicho antígeno. Si pudiera aislarse una célula plasmática e inducirla a proliferar para formar un clon de células idénticas, entonces se podría producir gran cantidad de anticuerpos idénticos. Desafortunadamente es muy difícil cultivar linfocitos y células plasmáticas, obstáculo que fue vencido por los científicos mediante la fusión de las células B con células tumorales, las cuales crecen con facilidad y proliferan en forma indefinida. El resultado de esta hibridación celular se conoce como hibridoma. Los hibridomas son fuentes duraderas de grandes cantidades de anticuerpos puros e idénticos denominados anticuerpos monoclonales (AcM). debido a que provienen de un único clon de células plasmáticas idénticas. Uno de los usos clínicos de los anticuerpos monoclonales es la medición de los niveles de un fármaco en la sangre de un paciente. Otros usos incluyen el diagnóstico de faringitis estreptocócica (test rápido estreptocócico o strep-test), embarazos. alergias y enfermedades tales como la hepatitis, rabia y algunas de transmisión sexual. Los AcM también se utilizaron para la detección de cánceres en las etapas más tempranas, así como también para determinar la extensión de las metástasis. Son igualmente útiles en la preparación de vacunas para contrarrestar los rechazos asociados a trasplantes, para el tratamiento de enfermedades autoinmunitarias y, tal vez, para el tratamiento del SIDA. .
Papel del sistema del complemento en la inmunidad
El sistema del complemento es un sistema defensivo, compuesto por más de 30 proteínas diferentes sintetizadas en el hígado, que se encuentran circulando por el plasma sanguíneo y por los tejidos del organismo. En conjunto, las proteínas del sistema del complemento destruyen a los microorganismos activando la fagocitosis, citólisis e inflamación: asimismo previenen el daño excesivo a los tejidos del organismo.
La mayoría de las proteínas del complemento se designan con una letra mayúscula C, numeradas desde Cl hasta C9, nombradas en el orden en que fueron descubiertas. Las proteínas C l-C9 del complemento se encuentran inactivas; mediante la división enzimática, se generan los fragmentos activos de ellas, que se indican con las letras minúsculas a y b. Por ejemplo, la proteína inactiva C3 del complemento se divide y genera los fragmentos activados C3a y C3b. Son los fragmentos activados los que llevan a cabo las funciones destructivas de las proteínas C1-C9 del complemento. Otras proteínas del complemento son los factores B. D Y P (properdina).
Las proteínas del complemento actúan en forma de cascada: una reacción desencadena otra reacción, la cual inicia otra reacción y así sucesivamente. Con cada reacción se generan cada vez más productos, de manera tal que el efecto neto es amplificado muchas veces.
La activación del sistema del complemento se puede producir por tres vías diferentes (que se describirán en breve), las cuales activan al componente C3. Una vez que se activa C3, se inicia una cascada de reacciones que inducen la fagocitosis, la citólisis y la inflamación de la siguiente manera (Fig. 22-18):
1.- El C3 inactivo se divide y forma los fragmentos activados C3a y C3b.
2.- El fragmento C3b se adhiere a la superficie de los microorganismos, mientras que los receptores de los fagocitos se unen al C3b. De esta manera, C3b facilita la fagocitosis, cubriendo a los microorganismos, proceso que se conoce como opsonización. La opsonización promueve la adhesión de los fagocitos a los microbios.
3.- El fragmento C3b también inicia una serie de reacciones que provocan citólisis. Primero, C3b divide a C5. El fragmento C5b se une, entonces, a C6 y C7, los cuales se unen a la membrana plasmática del microorganismo invasor. Después, C8 y varias moléculas de C9 se unen a las demás proteínas del complemento, las cuales en conjunto forman un complejo de ataque de membrana, con forma de cilindro, que se inserta en la membrana plasmática.
4.- El complejo de ataque de membrana crea canales en la membrana plasmática, lo que produce citólisis, el estallido de la célula microbiana causado por el ingreso de grandes cantidades de líquido extracelular a través de los canales.
5.- C3a y C5a se unen a los mastocitos, provocando la liberación, histamina por parte de éstos, lo cual genera el aumento de la permeabilidad vascular durante la inflamación. C5a también atrae a los fagocitos hacia el sitio de inflamación (quimiotaxis).
C3 puede activarse de tres formas diferentes: 1) La vía clásica se inicia cuando los anticuerpos se unen a los antígenos (microorganismos). El complejo antígeno-anticuerpo formado se une a C1, activándolo. Por último, se activa C3 y el fragmento C3b induce la fagocitosis, citólisis e inflamación. 2) La vía alterna no implica presencia de anticuerpos. Esta se inicia por la interacción entre complejos de lípidos-hidratos de carbono sobre la superficie de los microorganismos y los factores del complemento B, D y P. Esta interacción activa a C3. 3) En la vía de las lectinas, los macrófagos que digieren a los microorganismos liberan sustancias químicas que inducen en el hígado la secreción de proteínas denominadas lectinas.
Estas se unen a los hidratos de carbono presentes en la superficie microbiana, que en última instancia provoca la activación de C3.
Una vez que el sistema del complemento se activa, las proteínas plasmáticas y las proteínas que se encuentran en las células del organismo, como las células sanguíneas, degradan al C3b activado. En sentido, la capacidad destructiva del sistema del complemento disminuye rápidamente, de manera tal de minimizar todo daño posible que pudiera ser causado a los tejidos.
Memoria inmunológica
El sello de las respuestas inmunitarias es la memoria inmunológica para antígenos específicos que iniciaron una respuesta inmunitaria en el pasado. La memoria inmunológica se debe a la presencia, durante un período prolongado de tiempo, de anticuerpos y linfocitos de vida larga, que emergen durante la proliferación y diferenciación de las células B y T, ante el contacto con el antígeno.
Las respuestas inmunitarias, ya sean mediadas por células o por anticuerpos, se desarrollan más rápidamente y con mayor intensidad después de la segunda exposición u otra posterior con un antígeno dado, que en la exposición primaria. En un principio, sólo unas pocas células tienen la especificidad suficiente como para responder, de manera tal que la respuesta inmunitaria se desarrolla en un lapso de varios días, hasta que alcanza su máxima intensidad. Debido a que miles de células de memoria se generan luego del encuentro primario con un antígeno, la próxima vez que el antígeno se hace presente, aquellas proliferan y se diferencian a células plasmáticas o células T citotóxicas en tan sólo horas.
Una forma de medir la memoria inmunológica es mediante el título de anticuerpos, la concentración sérica de anticuerpos. Después del contacto inicial con un antígeno, durante varios días no se detectan anticuerpos. Posteriormente se detecta un leve incremento en el título de anticuerpos, IgM primero e IgG después, seguido por disminución (fig. 22-19). Esto se conoce como respuesta primaria.
Las células de memoria pueden permanecer durante décadas.
Cada nuevo encuentro con el mismo antígeno produce rápida proliferación de células de memoria. Después de varios encuentros, el título de anticuerpos asciende a valores mucho mayores que los alcanzados durante la respuesta primaria, constituidos principalmente por anticuerpos de tipo IgG. Esta respuesta, más acelerada e intensa, se denomina respuesta secundaria. Los anticuerpos producidos durante la respuesta secundaria presentan mucha mayor afinidad por el antígeno que aquellos producidos en la respuesta primaria y son, por lo tanto, más eficaces a la hora de la neutralización del antígeno.
Las respuestas primarias y secundarias se desarrollan en el contexto de una infección microbiana. Cuando una persona se recupera de una infección sin haber tomado fármacos antimicrobianos, es debido a la respuesta inmunitaria primaria. Si el mismo microorganismo nos infectase otra vez, la respuesta secundaria podría ser tan rápida que los microorganismos serían destruidos aun antes de que aparezca algún signo o síntoma clínico de la infección.
La memoria inmunológica constituye la base de la inmunización mediante vacunas contra ciertas enfermedades (por ejemplo, la poliomielitis). Cuando uno recibe una vacuna, la cual puede estar elaborada con microbios atenuados (debilitados) o muertos, o en subunidades de ellos, nuestras células B y T se activan. Así, en caso de encontramos con el patógeno vivo contra el cual fuimos vacunados, nuestro cuerpo iniciará la respuesta secundaria. El cuadro 22-4 muestra los encuentros antigénicos que son capaces de proveer inmunidad adquirida, tanto natural como artificial.
AUTORRECONOCIMIENTO Y AUTOTOLERANCIA
Para el funcionamiento adecuado, nuestras células T deben presentar dos características: 1) ser capaces de reconocer nuestras propias proteínas del complejo mayor de histocompatibilidad (CMH), proceso conocido como autorreconocimiento y 2) carácter de reactividad frente a fragmentos peptídicos de nuestras propias proteínas, condición conocida como autotolerancia (fig.22-20). Las células B también presentan autotolerancia. La pérdida de esta tolerancia lleva al desarrollo de las enfermedades autoinmunitarias.
Las células pre-T, en el timo, desarrollan la capacidad de autorreconocimiento a través de la vía de la selección positiva (fig. 22-20a). En este proceso, algunas de las células pre- T expresan receptores de células T (RCT) que interaccionan con moléculas propias del CMH presentes sobre las células epiteliales de la corteza tímica. Debido a esta interacción, las células T son capaces de reconocer partes de algún complejo antígeno CMH. Estas celulas T inmaduras fracasan en el reconocimiento de las moléculas propias del CMH sobre el epitelio de la corteza tímica y son las que mueren por apoptosis.
El desarrollo de la autotolerancia se lleva a cabo mediante un proceso de eliminación, llamado selección negativa en la cual las células T interactúan con las células dendrítícas localizadas en la unión entre la corteza y la médula del timo. A través de este proceso las células T que presentan receptores capaces de reconocer fragmentos peptídicos u otros antígenos propios, o sea, fragmentos moleculares presentes normalmente en el plasma, se eliminan o se inactivan (fig. 22-20a). Las células T seleccionadas para sobrevivir no responden a los autoantígenos, que no son más que los fragmentos moléculas que están presentes en el organismo en condiciones normales. La selección negativa se realiza por mecanismos de delección y anergia. En la delección, las células T autorreactivas mueren por apoptosis; en la anergia, en cambio, permanecen viables, pero incapaces de responder a la estimulación antigénica. Sólo el 1 a 5% de las células T inmaduras presentes en el timo reciben las señales apropiadas para evitar la apoptosis durante la selección positiva, así como también durante la selección negativa, emergiendo, de esta manera, como células T maduras e inmunocompetentes.
Una vez que las células T emergen del timo, todavía pueden llegar a encontrar una proteína propia que no les sea familiar; en esos casos también se pueden volver anérgicas si no está presente el coestimulador necesario (fig. 22-20b). La deleción de las células T autorreactivas también puede ocurrir luego de que abandonaron el timo.
De la misma forma, las células B generan tolerancia a través de la deleción y anergia (fig. 22-20c). A medida que las células B se desarrollan en la médula ósea, aquellas que presentan receptores antigénicos capaces de reconocer antígenos propios comunes (como lo son las moléculas del CMH o los antígenos de los grupos, sanguíneos) son eliminadas. Sin embargo, una vez que las células B se liberan al torrente sanguíneo, la anergia parece el mecanismo principal responsable de impedir que las células B respondan ante la presencia de proteínas propias. Cuando las células B se encuentran con un antígeno que no está asociado a una célula presentadora de antígenos, por lo general se encuentra ausente la señal coestimuladora. En este caso, la célula B es propensa a volverse anérgica (inactivada) más que activada.
El cuadro 22-5 resume las funciones de las células que participan en la respuesta inmunológica.
INMUNOLOGÍA DEL CÁNCER
A pesar de que el sistema inmunológico es capaz de responder ante la presencia de células cancerosas, en general, la protección que brinda es inadecuada, lo cual se pone de manifiesto en la cantidad de personas que mueren de cáncer cada año. En los últimos 25 años, la gran mayoría de las investigaciones realizadas se enfocaron en la inmunología del cáncer, el estudio de las formas en que pueden utilizarse las respuestas inmunológicas para la detección, control y tratamiento del cáncer. Por ejemplo, algunos tumores de colon liberan antígeno carcinoembrionario (ACE) hacia la sangre y las células cancerosas prostáticas liberan antígeno prostático específico (APE). La detección de antígenos en el torrente sanguíneo no se considera diagnóstico definitivo de cáncer; ello se debe a que en otras patologías, no cancerosas, también se puede registrar la liberación de ambos antígenos. Sin embargo la detección sérica de altos niveles de antígenos relacionados con cáncer, por lo general indica la presencia de un tumor maligno.
Encontrar la manera de inducir una respuesta potente, por parte de nuestro sistema inmunitario contra las células cancerosas, ha sido difícil. Se probaron varias técnicas diferentes, todas ellas con muy poco éxito. Una de las metodologías propuestas consiste en obtener linfocitos inactivados de una muestra de sangre luego cultivarlos con inlerleucina-2, El resultado es la formación de células asesinas activadas por linfocinas, las cuales se transfunden nuevamente al paciente. A pesar de que las células LAK lograron mejoras notables en algunos pocos casos, la mayor parte de los pacientes sufrió diversas complicaciones. En otro método, los linfocitos provienen de una pequeña muestra de biopsia de un tumor y luego se cultivan en un medio con interleucina-2. Una vez que proliferan en cultivo, estos linfocitos infiltrantes de tumores (tumor-infiltra/ing lymphocytes, T/L) se inyectan nuevamente al paciente. Cerca de la cuarta parte de los pacientes con melanomas malignos o carcinoma de células renales que recibieron terapia con TIL mostraron importante mejoría. La gran cantidad de estudios que actualmente se están llevando a cabo, dan razones para esperar que los métodos basados en la inmunidad, a la postre, conduzcan a la cura del cáncer. .
ESTRÉS E INMUNIDAD
El campo de la psiconeuroinmunología (PNI) estudia las vías de comunicación existentes entre el sistema nervioso, el sistema endocrino y el sistema inmunitario. Las investigaciones acerca de la PNI parecen justificar lo que ya muchos observaron: nuestros pensamientos, sentimientos, estados de ánimo y creencias, influyen tanto sobre el nivel de nuestra salud como también el curso de la enfermedad. Por ejemplo, el cortisol, hormona secretada por la corteza suprarrenal, asociada a la respuesta de estrés, ejerce acciones inhibitorias sobre nuestro sistema inmunitaria.
Si se quiere observar la relación existente entre nuestro estilo de vida y nuestra función inmunitaria, examínese, entonces, el ámbito de una universidad. A medida que progresa el semestre y las cargas de trabajo se van incrementando, aumentan las consultas de los estudiantes en los servicios de atención de salud estudiantil. Cuando el trabajo y el estrés se acumulan, los hábitos se modifican. Mucha gente fuma o consume mayor cantidad de alcohol cuando están estresados, hábitos que deterioran la función inmunitaria. Ante una situación de estrés, las personas no llevan una alimentación adecuada ni realizan ejercicios en forma regular, dos hábitos que mejoran el funcionamiento del sistema inmunitario. Aquellas personas que resisten el impacto negativo del estrés sobre la salud son más propensas a experimentar un sentido de control en el futuro, a comprometerse laboral mente, a tener expectativas positivas acerca de uno mismo y a sentirse contenidos por su grupo social. Para incrementar esta resistencia al estrés, se debe cultivar una mirada positiva, involucrarse en el trabajo y crear buenas relaciones con los demás.
Un período de sueño y de relajación adecuados son de es importancia para un sistema inmunitario sano. Pero cuando las horas del día no son suficientes a veces prima la tentación a robarle horas a la noche. De esta manera. el acortar el período del sueño brindaría unas horas más de tiempo productivo, a corlo plazo; sin embargo; a largo plazo el efecto puede ser el contrario, especialmente si se contraen enfermedades que obligan a perder varios días de clases, y también porque entorpece nuestra concentración y bloquea la creatividad.
Aún si se destina el tiempo para dormir ocho horas por día el estrés es capaz de producir insomnio. Si por la noche no logramos conciliar el sueño, es hora de mejorar nuestra forma de lidiar estrés y nuestras habilidades para la relajación Hay que asegurarse de dejar las preocupaciones de lado antes de ir a dormir. .
EL ENVEJECIMIENTO Y EL SISTEMA INMUNITARIO
Con el pasar de los años, la gran mayoría de las personas se hacen más susceptibles a cualquier tipo de infecciones y procesos malignos. Su respuesta a las vacunas disminuye y tienden a producir más anticuerpos (anticuerpos contra moléculas propias del organismo.). Además, el nivel de funcionamiento del sistema inmunitario se ve reducido. Por ejemplo, las células T se vuelven menos reactivas ante la presencia de antígenos y también menos células T responden a las infecciones. Esto podría ser el resultado de la atrofia del timo relacionada con la edad o por la menor producción de hormonas tímicas. Debido a que la población de las células T disminuye con la edad, las células B también se hacen menos reactivas. En consecuencia, los niveles de anticuerpos no se incrementan tan rápidamente en respuesta a la presencia de antígenos, lo cual aumenta la susceptibilidad a diversas infecciones. Por esta razón clave a las personas de edad avanzada se les "recomienda la aplicación anual de la vacuna contra la influenza (gripe).
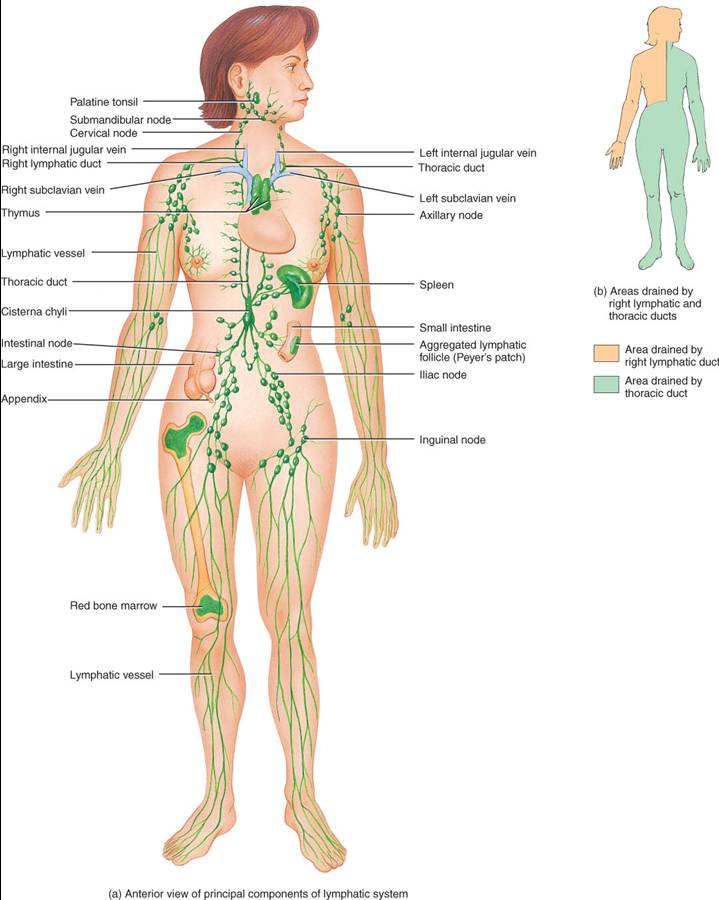{kind=link}
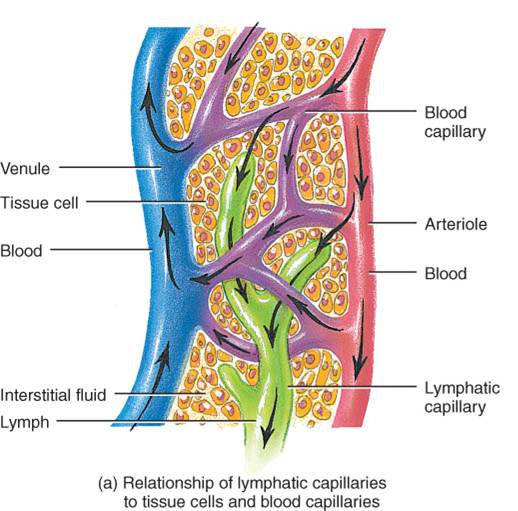{kind=link}
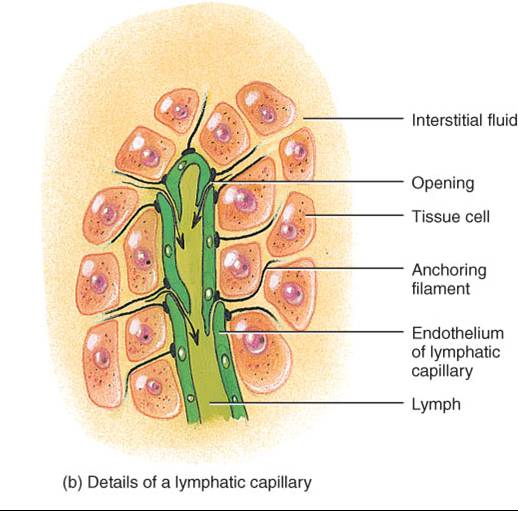{kind=link}
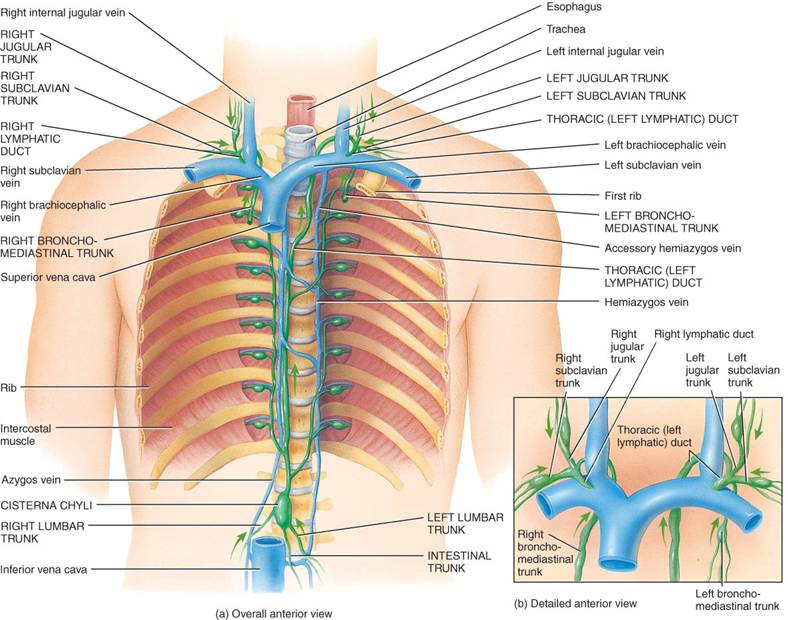{kind=link}
{kind=link}
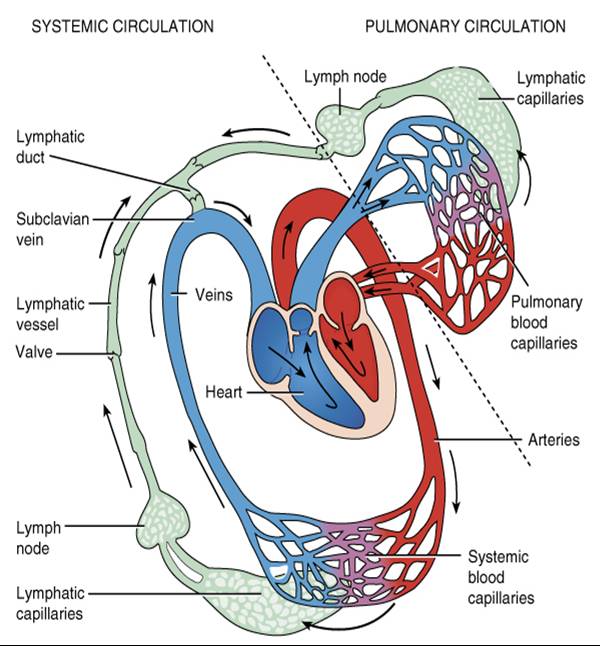{kind=link}
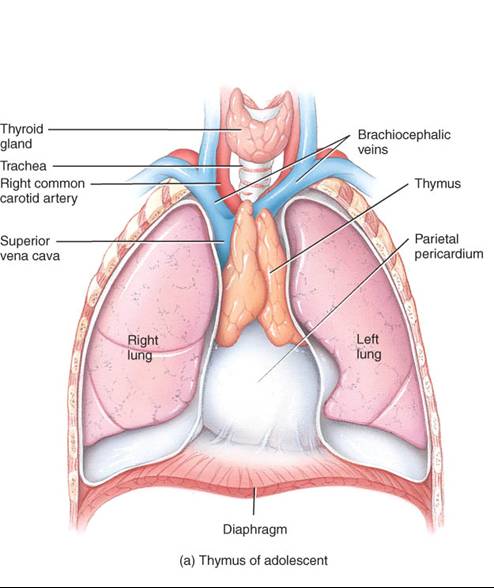{kind=link}
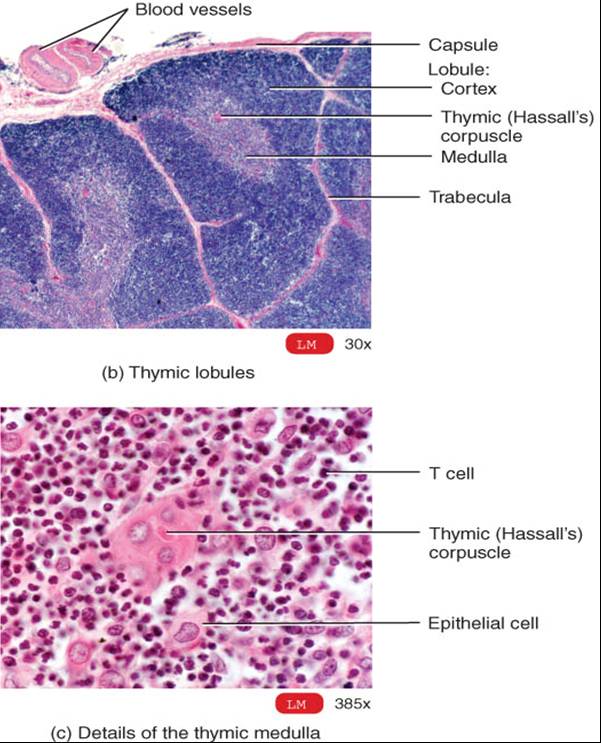{kind=link}
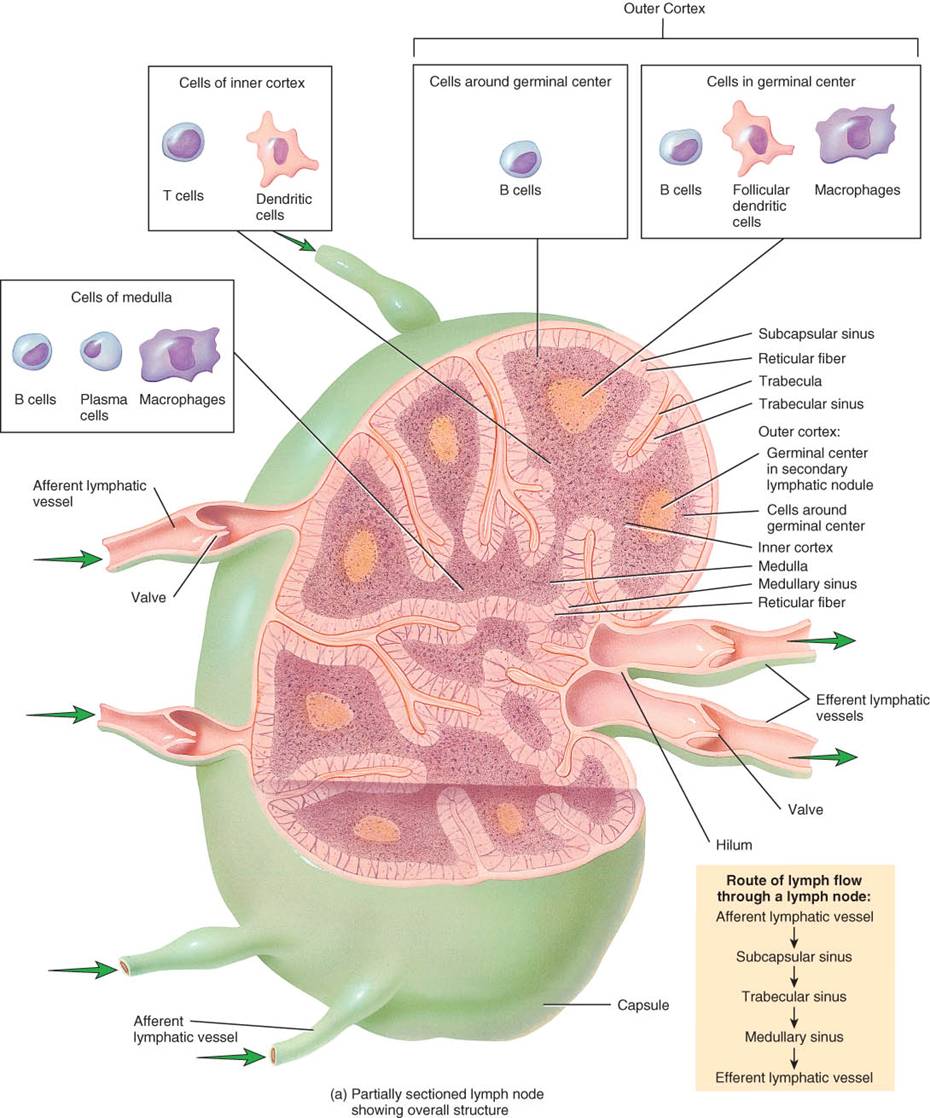{kind=link}
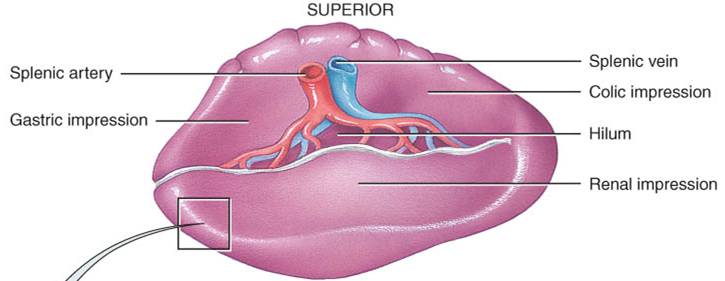{kind=link}
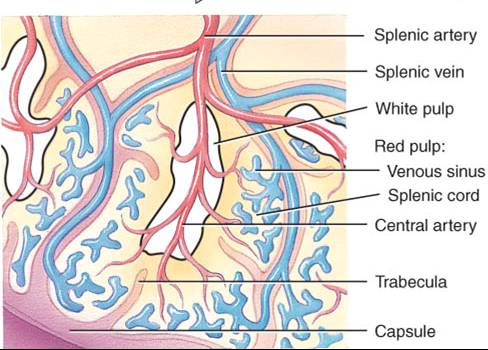{kind=link}
{kind=link}
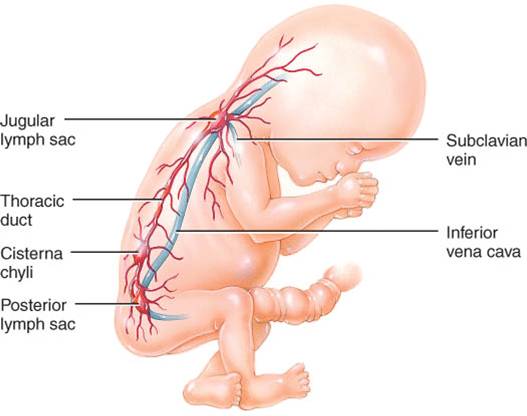{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
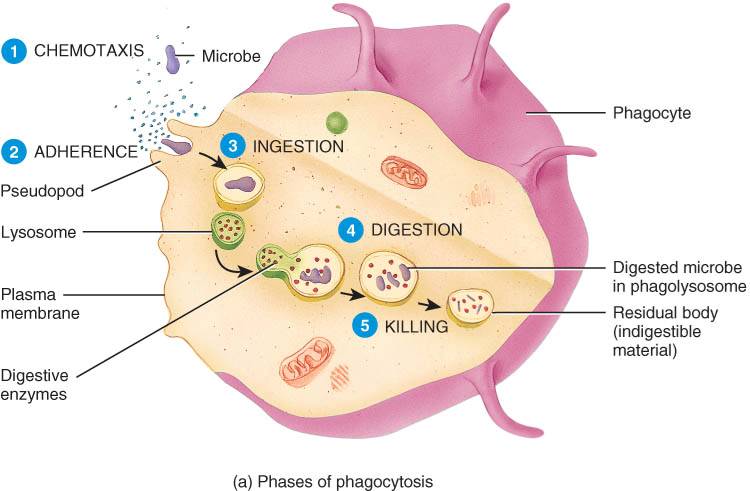{kind=link}
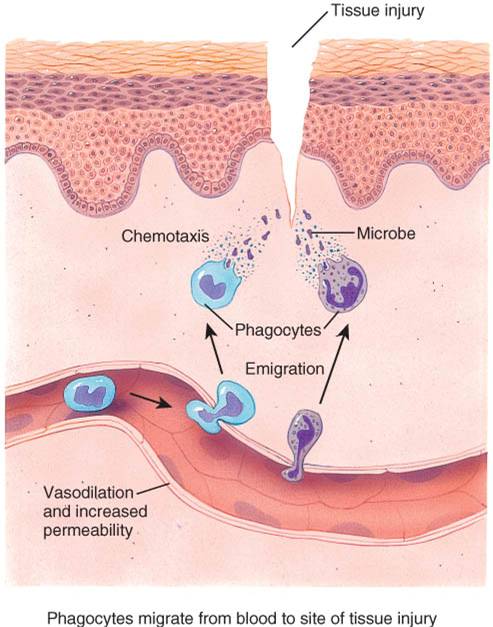{kind=link}
{kind=link}
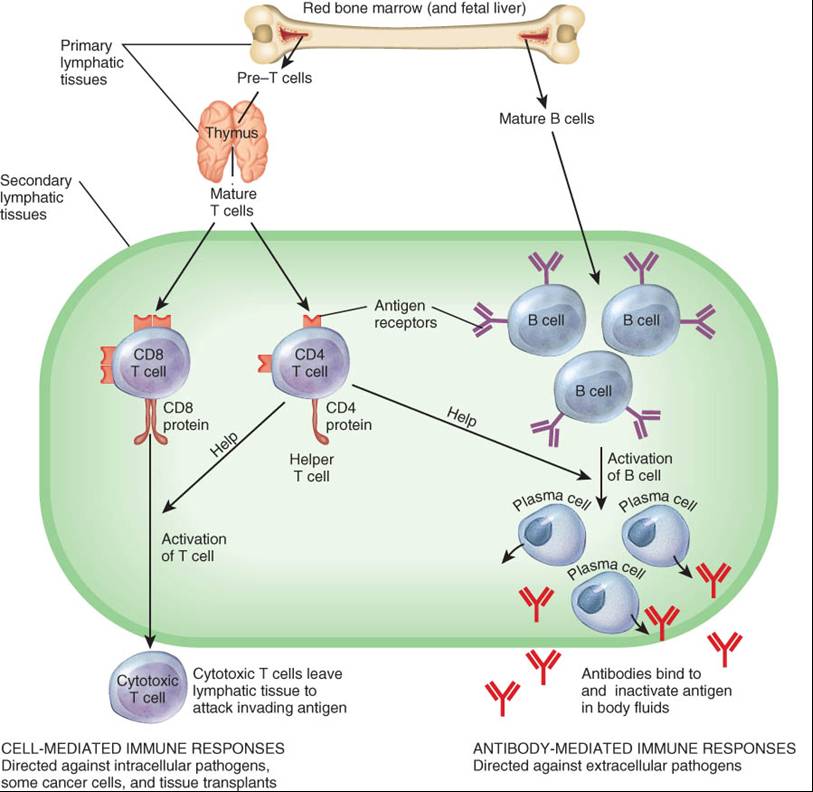{kind=link}
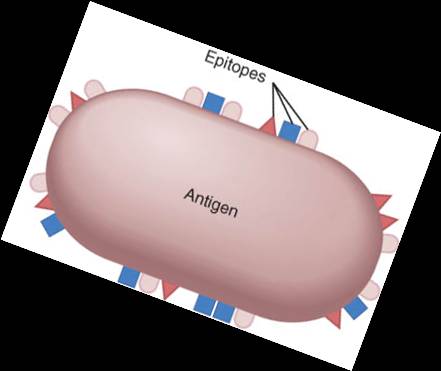{kind=link}
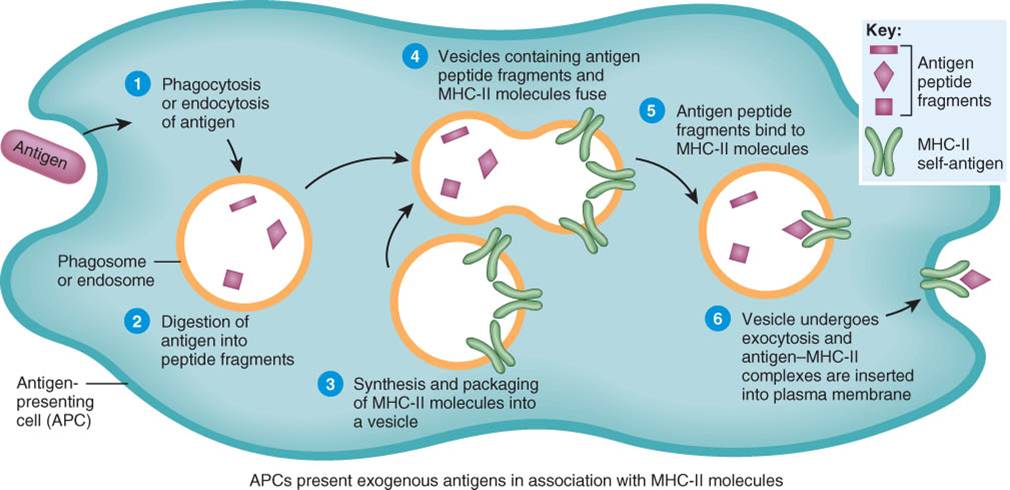{kind=link}
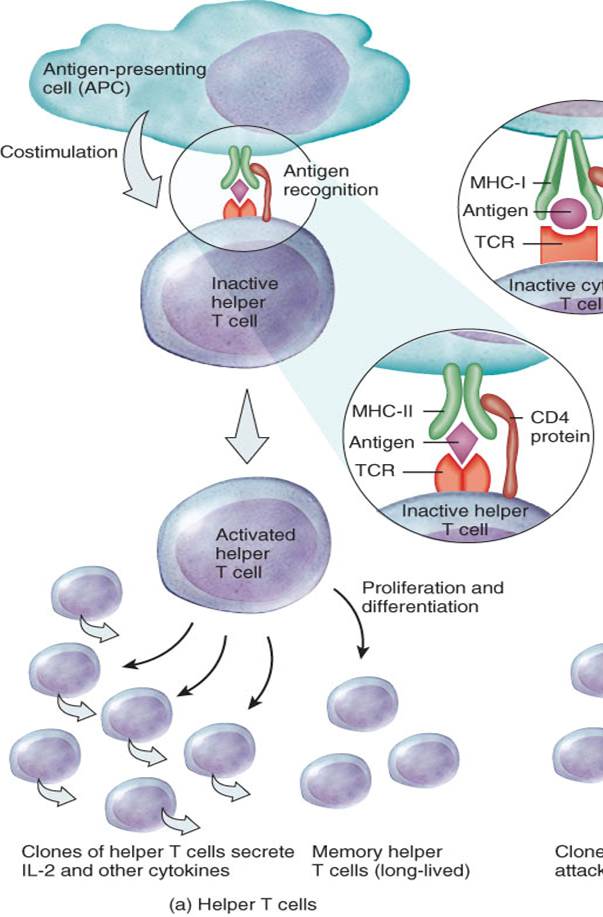{kind=link}
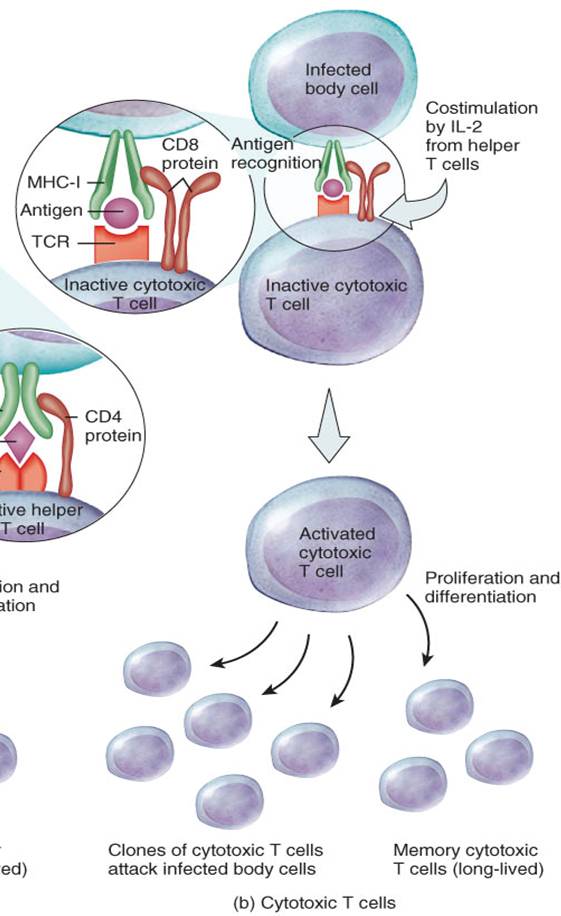{kind=link}
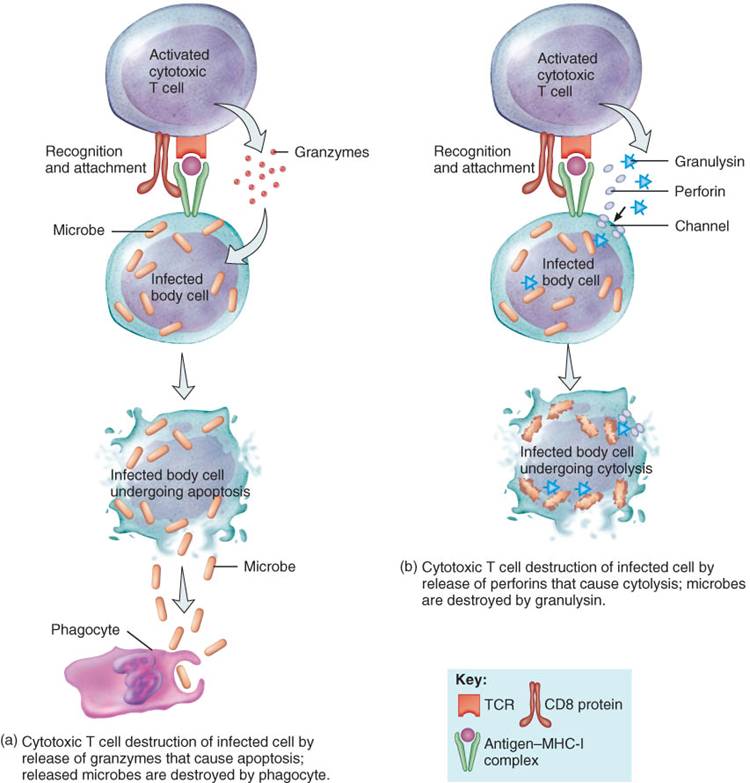{kind=link}
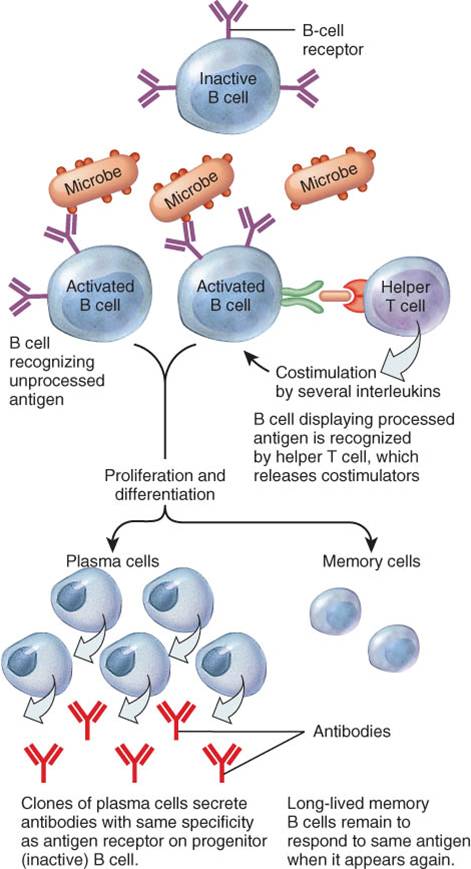{kind=link}
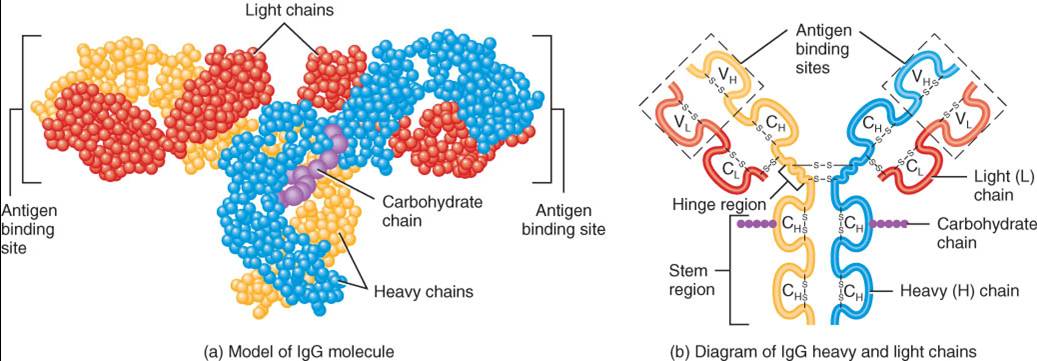{kind=link}
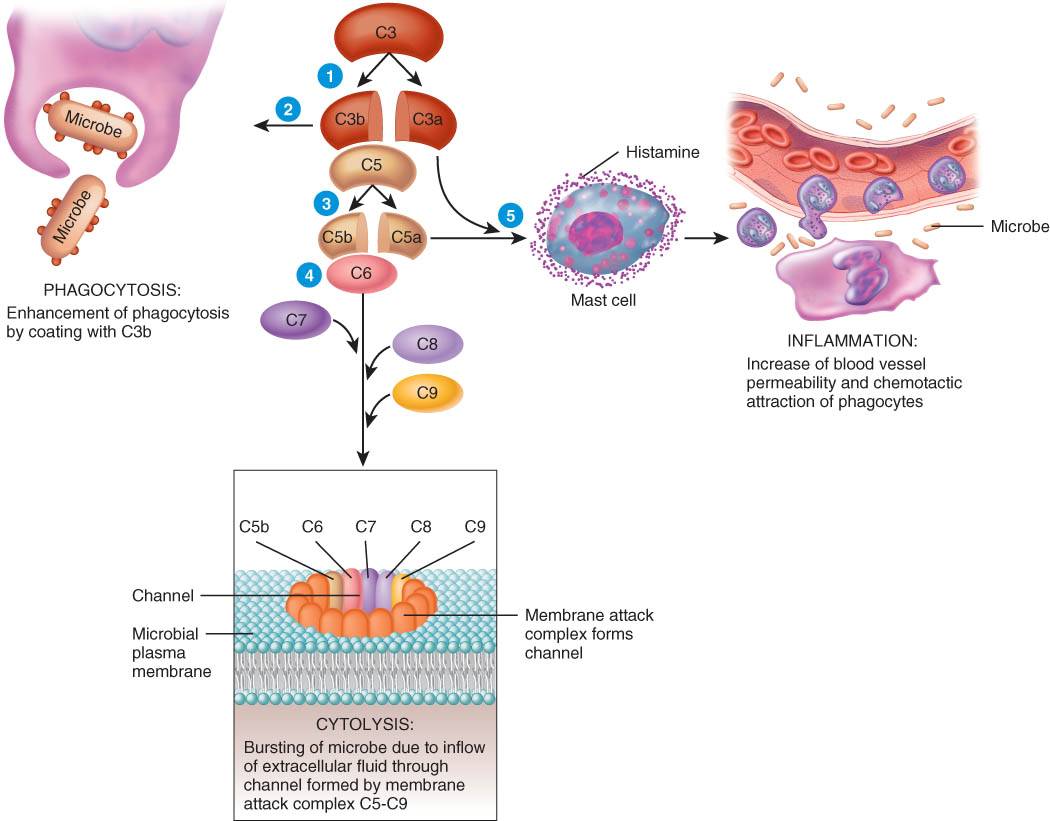{kind=link}
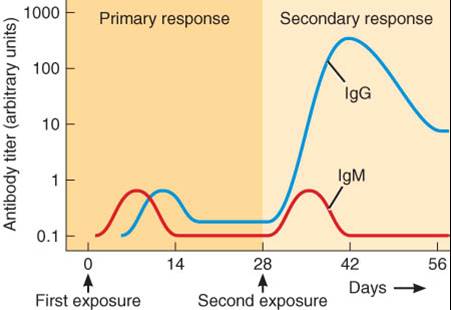{kind=link}
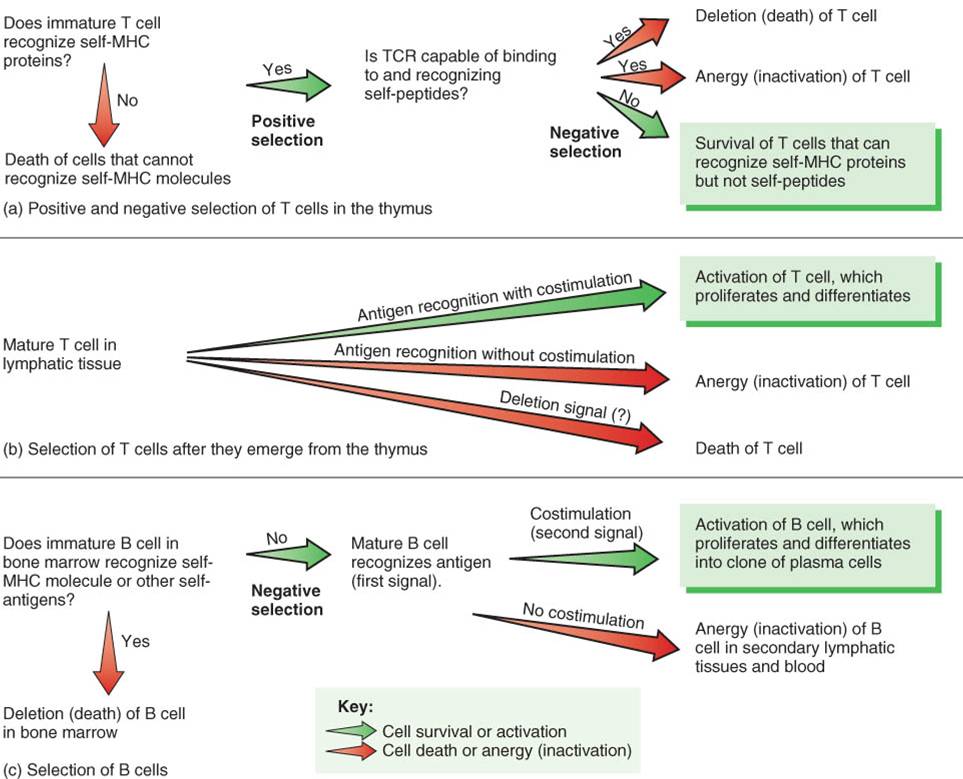{kind=link}