CAPITULO 3.
EL NIVEL CELULAR DE ORGANIZACIÓN.
ORNELAS.
La célula es la unidad básica funcional y estructural mas pequeña de los organismos vivos. Se compone de partes características, cuyo trabajo esta coordinado de tal manera que cada tipo de célula lleva a cabo una función estructural o bioquímica única. La citología es la ciencia que estudia la estructura celular, en tanto que la cito fisiología se aboca al estudio de las funciones que realizan estas unidades. Conforme el estudiante conozca las diferentes partes de la célula y las relaciones entre ellas, aprenderá que su función y estructura son interdependientes e inseparables. Las células realizan numerosas reacciones químicas para dar origen al proceso vital que se lleva a cabo de manera compartí mentalizada; es decir, estructuras especializadas dentro de la célula efectúan reacciones químicas aisladas, las cuales están coordinadas unas con otras para mantener con vida tanto la célula como los tejidos, los órganos, los sistemas y todo el organismo.
En términos muy generales, las células efectúan diversas funciones básicas. Por ejemplo, regulan el flujo de entrada (influjo) y de salida de los materiales a fin de asegurar las condiciones optimas para el proceso vital prevaleciente dentro de ellas. Asimismo, utilizan su información genética (ADN) para guiar la síntesis de la mayoría de sus componentes y dirigir gran parte de sus actividades químicas. Entre esas actividades esta la generación de ATP, por el desdoblamiento de los nutrientes, la síntesis molecular, la transportación de las moléculas dentro y entre las células, la remoción de los desechos y el movimiento parcial o incluso de toda la célula.
DESCRIPCIÓN GENERAL DE LA CÉLULA
OBJETIVO
Conocer y describir las tres partes principales de la célula.
En la figura 0.3.1, se presenta el esquema general de una célula que en realidad corresponde a una integración grafica de los diferentes tipos de células del cuerpo humano. Si bien estas unidades básicas tienen muchas de las característica; que aparecen en la figura, no todas ellas poseen este conjunto de componentes. Para facilitar el estudio es posible dividir las células en tres partes principales: membrana plasmática, citoplasma y núcleo.
• La membrana plasmática conforma la superficie extera de la célula, que separa su ambiente interno del externo.
Constituye una barrera selectiva que organiza el flujo de materiales que entran y salen de la célula a fin de establecer y mantener un ambiente apropiado para las actividades que lleva a cabo. La membrana plasmática también desempleeña una función clave en la comunicación entre las diferentes células, así como entre estas y su ambiente externo.
• El citoplasma comprende todo el contenido celular entre la membrana plasmática y el núcleo. Este compartimiento se puede dividir en dos componentes: cito sol y organeros.
El cito sol consiste en la porción liquida del citoplasma que esta formada principalmente por agua con partículas disueltas y suspendidas.
Los organeros son estructuras celulares altamente organizadas, cada una de las cuales tiene forma y función especificas. Entre los ejemplos podemos citar cito esqueleto, ribosomas, retículo endoplasmico, complejo de Golgi, lisosomas, peroxisoma y mitocondria.
El núcleo, si bien desde el punto de vista técnico es un organelo, se estudia aparte en virtud de sus numerosas y di-versas funciones. En su interior se encuentran los genes, que controlan la estructura de la célula y la mayoría de las actividades que realiza.
LA MEMBRANA PLASMÁTICA
OBJETIVO
• Describir la estructura y funciones de la membrana plasmática.
La membrana plasmática constituye una barrera flexible y fuerte que envuelve y contiene el citoplasma o matriz de la célula. El modelo de mosaico del liquido describe su estructura. La disposición molecular de la membrana es similar a un mar de lípidos en movimiento constante, que contiene muchas proteínas diferentes (fig. 0.3.2), las cuales pueden flotar libremente como icebergs, estar ancladas en ubicaciones especificas o moverse a través del mar de lípidos. Estos actúan como barreras que permiten la entrada o salida de sustancias polares, mientras que algunas proteínas de la membrana plasmática funcionan como "porteros" de la célula que regulan el trafico hacia dentro y hacia área de los iones y las moléculas polarizadas. La membrana plasmática de las células típicas del cuerpo esta compuesta por una mezcla aproximada de proteínas y lípidos en igual proporción, los cuales se mantienen juntos gracias a fuerzas no covalentes, como los enlaces de hidrogeno. Sin embargo, dado que las proteínas de la membrana son mas grandes y voluminosas que los lípidos, hay aproximadamente 50 moléculas de estos por cada proteína.
La bicapa lipidica
El marco estructural básico de la membrana plasmática es la bicapa de lípidos (o lipidica), dos capas confrontadas que están constituidas por tres tipos de moléculas de lípidos fosfolipidos, colesterol y glucolipido (vease la fig. 0.3.2). Los fosfolipidos, que contienen grupos o radicales fosfato, integran 75% de la membrana. En cantidades mas pequeñas se halla el colesterol, un esteroide al que esta unido un radical -OH (alcohol), así como diversos glucolipidos enlazados a grupos de hidratos de carbono (azucares).
La disposición de la bicapa se mantiene porque los lípidos son moléculas amfipaticas, es decir, tienen partes de carácter polar y apolar. En los fosfolipidos (vease la figura 0.2.18) la parte polar corresponde al fosfato, que es hidrófilo. Las partes no polares son dos caudas largas de ácido graso, formadas por cadenas hidrofobas de hidrocarburos. Debido a que hay afinidad entre iguales, en la bicapa las moléculas fosfolipidas orientan sus cabezas polares hacia afuera, es decir, uno de sus lados se proyecta hacia el entorno acuoso citosol en la cara interna y liquido extracelular hacia la externa. Los ácidos grasos hidrofobos extienden sus caudas en ambas mitades de un punto de la bicapa hacia el centro de la membrana, con lo que conforman en la zona interna una región apolar e hidrófoba.
Los glucolipidos constituyen 5% de la membrana lipidica; sus grupos carbohidratos forman una cabeza polar, mientras que sus caudas de ácidos grasos carecen de polaridad. Se encuentran solo en la capa de la membrana que co-munica con la fase del liquido extracelular y, por ello, se dice que la bicapa es asimétrica; dicho en otras palabras, sus dos lados son diferentes. De la membrana plasmática, 20% restante de los lipidos esta compuesto por moléculas de colesterol débilmente anfipaticas (vease la fig. 0.2.19), las cuales se hallan interpuestas entre los otros lípidos de ambas capas. El diminuto grupo unido al radical -OH constituye la única región polar del colesterol y forma enlaces de hidrogeno con las cabezas polares de los fosfolipidos y los glucolipidos. Los anillos de esteroides rígidos y las caudas de hidrocarbono del colesterol, que no son de carácter polar, se hallan fijas entre" las colas de ácido graso de los fosfolipidos y los glucolipidos.
De manera característica la membrana plasmática tiene mas colesterol que la membrana de los organelos.
Disposicion de las proteinas en la membrana
Estas proteínas se dividen en dos categorías integras y periféricas según se hallen insertas o no en la membrana (fig. 0.3.2). Las proteínas integras se extienden dentro o a través de la bicapa entre las caudas de los ácidos grasos y solo se pueden extirpar por métodos que alteran la estructura membranosa. Por otra parte, las proteínas periféricas se relacionan con lípidos o con las proteínas integras que la membrana tiene en sus lados interno o externo, y se pueden extraer por métodos que no alteran su integridad. Al igual que los lípidos, las proteínas de la membrana integral son amfipaticas; sus regiones hidrófilas se proyectan ya sea hacia la fase del liquido extracelular o hacia el citosol, y sus zonas hidrofobas se extienden entre las caudas de ácidos grasos. Las proteínas integras que pasan a través de la bicapa entera y sobresalen en ambos lados de la membrana reciben el nombre de proteínas transmembranosas. Con frecuencia la porción de una proteína que entra en la región apolar de la membrana consiste en una helice alfa la cual esta formada por aminoácidos que constan principalmente de cadenas laterales hidrófobas apolares. En algunos casos la cadena de aminoácidos de la proteína oscila de un lado y otro de la membrana. Los segmentos de la helice alfa hidrófoba que atraviesan de un lado a otro dicha membrana se alternan con las porciones polares hidrófilas que se proyectan hacia el liquido en ambos lados. Aunque muchas de las proteínas integras pueden flotar lateralmente en la bicapa lipida, cada proteína individual tiene una orientación con respecto a las caras interna y externa de la membrana y de esta manera contribuye a que esta sea asimétrica.
Muchas proteínas integras son glucoproteínas con grupos de carbohidratos unidos a los extremos que se extienden hasta la fase del liquido extracelular. Los carbohidratos pueden ser cortos, rectos o en cadenas ramificadas compuestas por dos a 60 monosacáridos. Las porciones carbohidratos de los glucolipidos y las glucoproteinas forman una extensa cubierta de azúcar denominada glucocaliz, que desempefia varias funciones importantes y cuya composición actúa como una forma de identidad molecular que permite que las celúlas se reconozcan unas a otras. Por ejemplo, una de las bases de la respuesta inmunitaria que ayuda al cuerpo a destruir los microorganismos invasores es la capacidad de los leucocitos (células blancas) para detectar un glucocaliz extrafio. Ademas, esta cubierta hace posible que en ciertos tejidos las células se adhieran unas a otras con lo que evita que sean digeridas por las enzimas del liquido extracelular. Las propiedades hidrófilas del glucocaliz atraen una película de liquido a la superficie de muchas células, una propiedad que tiene una funcion doble: hace resbalosos a los eritrocitos mientras fluyen a través de los vasos sanguíneos estrechos y evita que las células que revisten las vías respiratorias y el tracto gastrointestinal se sequen.
Funciones de las proteínas de la membrana
En general, los diferentes tipos de lípidos varían solo ligeramente de una membrana a otra. Mas ahi, las membranas de las distintas células y de los diferentes organelos singularizan notablemente diferentes clases de proteínas, las cuales a su vez determinan muchas de las funciones que pueden realizar las membranas plasmáticas. En la figura 0.3.3 se ilustran varias de estas funciones. Algunas proteínas transmembranosas forman canales que tienen un poro o agujero a través del cual pueden entrar o salir de la célula sustancias especificas como los iones potasio (K ). La mayoría de dichos canales son selectivos, ya que solo permiten el paso de un ion determinado. Otras proteínas de la membrana actúan como transportadores; es decir se unen a una sustancia polar en un lado de la membrana y luego cambian su forma para llevarla al otro lado donde la liberan. Algunas proteínas integras se denominan receptores los cuales sirven como sitios de reconocimiento celular. Estas proteínas identifican y enlazan una molécula especifica (p. ej., la hormona insulina o un nutriente como la glucosa) que resulta importante para algunas funciones celulares. Se denomina ligando una molécula especifica que se une a un receptor. Algunas proteínas integras y periféricas son enzimas. Las membranas de las glucoproteinas y los glucolipidos con frecuencia se llaman marcadores de identidad celular, los cuales hacen posible que una célula reconozca a otras de la misma clase durante la formación de un tejido o identifiquen y respondan a células extrañas potencialmente peligrosas. Entre los marcadores de identidad celular están los del tipo sanguíneo (ABO). Cuando se transfunde sangre, su tipo debe ser compatible con el de la persona' que la recibe. Las proteínas periféricas e integrales pueden servir como ligadores, que fijan las proteínas en la membrana plasmática de las células vecinas, una a otra, o a los filamentos dentro y fuera de la célula.
Fluidez de la membrana
Las membranas son estructuras liquidas, mas que aceitosas, ya que así la mayor parte de sus lípidos y muchas de sus proteínas, pueden girar fácilmente y moverse hacia los lados en el interior de la mitad de la bicapa donde se localizan. Las moléculas lapidas vecinas intercambian lugares aproximadamente 10 millones de veces por segundo y pueden vagar alrededor de la célula en pocos minutos. La fluidez de la membrana de-pende del numero de enlaces dobles en las caudas de ácido graso de los lípidos que constituyen la bicapa y de la cantidad de colesterol presente. Cada enlace doble es un eslabón en la cauda de ácido graso (vease la fig. 0.2.18), el cual evita que las moléculas lipidicas formen un empaque rígido en la membrana. La fluidez de los lípidos permite que autosellen esta cubierta si es desgarrada o punzada. Cuando se introduce una aguja a través de la membrana plasmática y luego se extrae, el sitio de perforación cierra espontáneamente y la célula no revienta. Esta propiedad hace posible que los genetistas fertilicen un óvulo al inyectar el espermatozoide con una jeringa diminuta o reemplacen el núcleo de una celúla en experimentos de clonación, como los realizados para crear la clona de la oveja llamada Dolly.
No obstante la gran movilidad de lípidos y proteínas de la membrana en su mitad de la bicapa, cambian de manera aleatoria de una mitad a otra. Esto se debe a que las parte; hidronlas pasan con gran dificultad a través del centro hidrófobo de la membrana. Dado que estos cambios son poco frecuentes, las mitades de la membrana de la bicapa permanecen asimétricas la mitad externa contiene algunos lípidos y proteínas que son estructural y funcionalmente distintos de los que están en la mitad interna.
En virtud de la manera en que el colesterol forma enlaces o puentes de hidrogeno con las cabezas de fosfolipidos y glucolipidos adjuntos y llena los espacios entre las caudas dobladas de ácido graso, confiere mayor fuerza a los lípidos de la bicapa, pero menor fluidez a la temperatura normal del cuerpo. Vale la pena hacer notar que a bajas temperaturas el colesterol tiene efecto opuesto -incrementa la fluidez de la membrana, lo que resulta importante para los animales de sangre fría (mas no para los humanos, que son de sangre caliente). Cuando se acumulan cantidades excesivas de colesterol en las membranas plasmáticas estas se tornan rígidas y menos flexibles. Una consecuencia de lo antes expuesto es la aterosclerosis, comúnmente conocida como "endurecimiento de las arterias", o acumulación de colesterol en las membranas plasmáticas de las células que revisten los vasos sanguíneos, un problema característico de esta enfermedad.
Permeabilidad de la membrana
Una membrana es permeable para ciertas cosas que pueden pasar a través de ella e impermeable para otras. Aunque las membranas plasmáticas no son completamente impermeables a todas las sustancias, permiten que algunas de ellas las atraviesen más rápidamente que otras. Esta propiedad se denomina permeabilidad selectiva.
La porción lipidia de la bicapa de la membrana es permeable a la mayoría de las moleculas apolares y sin carga. Como las de oxigeno, dióxido de carbono y esteroides, perc es impermeable a los iones y a las moléculas polares con carga, como la glucosa. Asi mismo tiene permeabilidad al agua. Una propiedad sorprendente, ya que la molécula de H2O es polar. Incluso las membranas fabricadas artificialmente tienen permeabilidad al agua, que se cree pasa a través de la bicapa lipidica de la siguiente manera: cuando las colas de ácido graso de la membrana de fosfolipidos y glucolipidos se mueven de manera aleatoria, dejan pequeños espacios por breves instantes en el ambiente hidrófobo de la membrana. Las moléculas de agua son lo suficientemente pequeñas para moverse de un espacio a otro hasta cruzarla.
Las proteínas que funcionan como canales y transportadores hacen que la membrana plasmática sea permeable a una variedad de pequeñas sustancias de tamafios pequeño y mediano polares y cargadas (entre ellas iones) que no pueden cruzar la bicapa lipidica. Esas proteínas son muy selectivas cada una ayuda solo a una molécula o ion especifico a cruzar la membrana. Las macromoléculas, como las proteínas, no tienen la capacidad de atravesarla membrana excepto por transporte vesicular (que se estudia mas adelante en este capitulo).
Paso por gradientes a través de la membrana plasmática
En virtud de que la membrana plasmática de las células posee permeabilidad selectiva, admite unas sustancias en el citosol mientras excluye otras. Esta propiedad hace posible que la célula viva mantenga grandes o pequeñas concentraciones de ciertas sustancias selectas ya sea en el citosol o en el liquido extracelular. La diferencia en la cantidad de un quimico entre un lado y otro de la membrana plasmática se denomina gradiente de concentración. En el liquido extracelular hay muchos iones y moléculas en mayor cantidad que en el citosol, mientras que con otros sucede lo contrario. Por ejemplo, las moléculas de oxigeno y los iones sodio (Na+) son mas abundantes en el liquido extracelular que en el citosol, en tanto que sucede lo opuesto con las de dióxido de carbono y los iones potasio (K+) (fig. 0.3.4a). La membrana plasmática también da origen a una diferencia en la distribución de iones con carga positiva y negativa entre sus dos lados; esto es lo que se denomina gradiente eléctrico, también llamado potencial de membrana. De manera característica, la superficie interna de la membrana tiene una carga negativa y la externa es mas positiva (fig. 0.3.4a).
Como se vera un poco mas adelante, tanto el mantenimiento de la concentración como los gradientes eléctricos son importantes para la vida de la célula, porque ayudan a movilizar sustancias a través de la membrana plasmática. En muchos casos, una molécula cruza la membrana hacia el sitio donde su gradiente de concentración es mas bajo. Dicho en otras palabras, una sustancia se mueve "colina abajo" desde el punto donde su concentración es mas alta hasta el lugar donde es mas bella. De manera similar un ion con carga positiva tendera a moverse en dirección del área con carga negativa. Dado que los iones reciben influencia tanto del gradiente de concentración como del gradiente eléctrico, el efecto combinado recibe el nombre de gradiente electromecánico.
TRANSPORTE A TRAVÉS DE LA MEMBRANA PLASMÁTICA
OBJETIVO
• Describir el proceso por el cual se lleva a cabo el transporte de sustancias a traves de la membrana plasmática.
El transporte de materiales a través de la membrana plasmática resulta esencial para la vida de la célula. Se requiere que ciertas sustancias entren en ella para que contribuyan a las reacciones metabólicas; otras deben salir, porque son productos elaborados por la célula que deben ser exportados o materiales de desecho. Como se ha expuesto algunas sustancias pueden cruzar la porción lipidica de la bicapa, mientras que otras usan canales o transportadores de las proteínas de la membrana plasmática.
Las sustancias que cruzan las membranas celulares por el proceso de transportación se clasifican conforme a dos criterios: 1) su movimiento es mediado o no; 2) son activas o pasivas. En el transporte mediado, los materiales pasan a través de la membrana con ayuda de una proteína transportadora; mientras que en el transporte no mediado, no participan lo; transportadores. En el transporte pasivo, una sustancia se mueve cuesta abajo y cruza la membrana gracias a su gradiente de concentración y solo utiliza su energía cinética, en tanto que la energía celular de transporte activo, por lo regular en forma de ATP, se emplea para llevar el material en sentido opuesto a su gradiente de concentración.
Las células vivas utilizan tres procesos de transporte (fig.0.3.5a). Dos de ellos son directos (no mediados): difusión a través de la bicapa lipidica o por un canal. En un proceso pasivo, la difusión facilitada requiere un mediador. Entre los de transportación activa, que son mediados, es posible distinguir varios tipos generales de transportadores (fig.0.3.5b).Los unitransportadores trasladan una sola sustancia a través de la membrana. Los bitransportadores llevan simultáneamente dos; además son cotransportadores, es decir, mueven ambas sustancias en la misma dirección o contratransportadores por que movilizan las dos sustancias en direcciones opuestas por la membrana.
Otro proceso por el cual los materiales pueden entrar o salir de la célula es el que involucra la formación de las vesículas que rodean la membrana. En el transporte vesicular (veanse las figuras 0.3.13 y 0.3.15) se desprenden diminutas vesículas de la membrana para introducir materiales a la célula, un fenómeno llamado endocitosis, o se fusionan con la membrana plasmática para liberar los materiales en la célula, lo que se denomina exocitosis. Las partículas grandes como una bacteria entera, los eritrocitos y las macromoléculas, como los polisacáridos y las proteínas, pueden entrar y salir de las células por este tipo de transporte.
Fundamentos de la difusión
Para comprender por que se diseminan los materiales a través de la membrana, es preciso entender como se difunden en un liquido. Se denomina difusión a la mezcla aleatoria de partículas que ocurre en una solución como resultado de la energía cinética de dichas partículas. Tanto los solutos, las sustancias disueltas, como los solventes, líquidos donde se disuelven, entran en difusión. Si la cantidad de una molécula en particular es alta en una zona de la solución y baja en otra, será mayor el desplazamiento del sitio donde es mas abundante hacia el lugar donde es menor su cantidad. La diferencia del paso en ambas direcciones recibe el nombre de difusión neta. Las sustancias correspondientes a la difusión neta se mueven de la zona de alta concentración a la de baja, y por ello se dice que se desplazan hacia abajo o hacia el área donde su gradiente de concentración es bajo. Se logra el equilibrio cuando las moléculas finalmente se distribuyen de manera homogénea en el liquido. Una vez en equilibrio, las moléculas prosiguen su desplazamiento aleatorio en virtud de su energía cinética, pero ya no modifican su concentración.
Como ejemplo de lo anterior podemos citar lo que sucede cuando en un recipiente de vidrio lleno de agua se pone un tinte (fig. 0.3.6). Cerca del lugar donde este se vierte, el color es intenso porque ahí es mas alta su concentración, pero conforme es mayor la distancia el tono se torna mas y mas claro, a medida que es menos concentrado. Tiempo después, cuando se alcanza el equilibrio, la solución de agua con el tinte alcanza un color uniforme. Las moléculas del tinte lo-gran su difusión neta al diseminarse desde el punto donde su concentración es mas alto, hasta que se encuentran mezcladas de manera uniforme.
En el ejemplo anterior, no participa membrana alguna, aunque cuando hay una de por medio, las sustancias a las que es permeable también pueden difundirse a través de ella. La velocidad de difusión de las moléculas por la membrana plasmática depende de varios factores.
1. Gradiente de concentración. Mientras mayor sea la diferencia entre ambos lados de la membrana, mayor será la velocidad de difusión. Cuando las partículas se encuentran diseminadas, los factores que determinan su velocidad de difusión a través de la membrana son el diferencial de concentración y los gradientes eléctricos (el gradiente electroquímico).
2. Temperatura. A mayor temperatura, mas rápida es la velocidad de difusión. En una persona con fiebre, ocurre mas pronto todo el proceso de difusión corporal.
3. Tamaño o masa de la sustancia que se difunde. Mientras mas grande resulte el tamaño o la masa de la partícula es menor su velocidad de difusión. Las moléculas de menor peso molecular se difunden con mayor rapidez que las de mayor peso.
4. Área superficial. Cuanto mas grande es el área superficial de la membrana disponible para la difusión, mayor será la velocidad de este proceso. Por ejemplo, las células de los pulmones tienen un área mas grande para la propagación del oxigeno del aire en la sangre. En algunas enfermedades, como el enfisema, se reduce el área superficial, lo que hace mas lenta la velocidad de difusión y reduce el ritmo respiratorio.
5. Distancia de difusión. Mientras mas grande sea la distancia en que ocurre, mas tiempo se requerida para la difusión. Este fenómeno a través de la membrana plasmática solo toma una fracción de segundo porque es muy delgada.
A continuación se estudia la forma en que se lleva a cabo el transporte en la membrana al considerar como la cruza un solvente acuoso; posteriormente se analizan las distintas categorías de solutos.
Osmosis
La osmosis es el paso de un. solvente a través de una membrana con permeabilidad selectiva. En los sistemas vivos, el agua constituye el solvente que cruza por osmosis la membrana plasmática desde un área de mayor concentración a otra donde esta resulta menor. Otra manera de entender esta idea consiste en considerar la concentración de solutos o sustancias disueltas: en la osmosis el agua se mueve a través de la membrana con permeabilidad selectiva desde el área con menor concentración de solutos a una con mayor. Las moléculas de este liquido penetran esta barrera de dos maneras por difusión a través de la bicapalipidica, como antes se menciono, y a través de acuaporinas, proteínas transmembranas que funcionan como canales de agua.
La osmosis ocurre solo cuando una membrana es permeable al agua, pero no a ciertos solutos. Es factible entender el fenómeno de la osmosis mediante un experimento sencillo. Considérese un tubo de vidrio en forma de "U", en el cual una membrana con permeabilidad selectiva separa los brazos izquierdo y derecho (fig. 0.3.7a). Se vierte cierto volumen de agua pura en la parte izquierda y, en la derecha, una cantidad igual de solución, que contiene un soluto al cual es impermeable la membrana. Dado que la concentración de agua es mas alta en el brazo izquierdo que en el derecho, el movimiento neto de las moléculas de agua osmosis ocurre en sentido de mayor a menor. A la vez la membrana evita la difusión de los solutos del lado derecho al izquierdo.
Como resultado, disminuye el volumen de agua del lado derecho y aumenta la solución en el opuesto (fig. 0.3.7b).
El estudiante puede suponer que el proceso de osmosis seguirá hasta que no quede agua en el brazo izquierdo, pero no sucede así. En este experimento, cuanto mas alta sea la columna de solución en el lado derecho, mayor presión ejercerá sobre la cara correspondiente de la membrana; este fenómeno recibe el nombre de presión hidrostática, la cual fuerza a las moléculas de agua a moverse de regreso al brazo izquierdo. El equilibrio se alcanza cuando la cantidad de moléculas de H2O que se mueve de derecha a izquierda, en virtud de la presión hidrostática, es igual a la impulsada en sentido opuesto por la osmosis (fig 0.3.7b). Una solución con partículas disueltas que no pueden cruzar la membrana ejerce una fuerza denominada presion osmótica, la cual es proporcional a la concentración de partículas disueltas que no pueden atravesar la membrana (mientras mayor sea la concentración de solutos, mas alta será la presión osmótica.
Considere el lector lo que sucedería si se usara un pistón para aplicar mas presion sobre el liquido del brazo derecho. Con suficiente presion, seria factible restaurar el volumen inicial, y la concentración de solutos en este lado seria la misma que al empezar el experimento (fig. 0.3.7c). La presion necesaria para reestablecer el estado inicial debe ser igual a la presion osmótica, por lo cual cabe definir este concepto de otra manera: es la presion que se requiere para detener el movimiento del agua hacia una solución que contiene solutos cuando los dos se encuentran separados por una membrana que solo es permeable al agua. Adviértase que la presion osmótica de una solución no produce movimiento de H2O durante la osmosis. En vez de ello, constituye la presion que evita dicho movimiento.
La tonicidad de una solución influye en el volumen y la forma de las células corporales al ocasionar la osmosis del agua que entra y sale de ellas. Normalmente, la presion osmótica del citosol es igual a la de la fase intersticial entre las células. Como dicha presion es igual en ambos lados de la membrana plasmática (que tiene permeabilidad selectiva), el volumen celular permanece relativamente constante.
Cualquier solución en la cual una célula (por ejemplo, los eritrocitos [RBC]) mantiene su forma y volumen normales se denomina isotónica (fig. 0.3.8a). Es decir, se trata de un liquido en el cual la concentración de solutos que no pueden cruzar la membrana plasmática es la misma en ambos lados. En circunstancias normales, una solución al 0.9% de NaCl (cloruro de sodio o sal de mesa común) llamada solución salina normal es isotónica para los eritrocitos. Si bien la membrana plasmática de la células rojas permite el transito del agua en ambos sentidos se comporta como si fuera impermeable a los solutos de Na y CT. Cualquier ion de estos elementos que entra en la célula por medio de los canales o de transportadores es devuelto de inmediato, por el transporte activo y otros medios. Por ende, se considera que estos iones se comportan como si no pudieran penetrar la membrana. Cuando se bafla un eritrocito en solución salina al 9% las moléculas de agua entran y salen a la misma velocidad, lo que hace posible que estas células mantengan su forma y volumen inalterados.
El resultado es diferente si el eritrocito se pone en una solución que tiene menor concentración de solutos que el citosol intracelular. Este liquido se denomina solución hipotonica. (fig. 0.3.8b). En este caso las moléculas de agua entran en la célula mas rápido de lo que salen y ocasionan que esta se hinche y finalmente estalle. La ruptura de un eritrocito por esta causa recibe el nombre de hemólisis. El agua pura es fuertemente hipotónica.
Una solución hipertónica posee una concentración mas elevada de solutos que el citosol dentro de un eritrocito (fig. 0.3.8c). Una solución de NaCl al 2% constituye un ejemplo de un liquido hipertónico. Lo cual significa que las moléculas de agua se salen de la célula con mayor rapidez de la que entran, lo que ocasiona que esta se contraiga. Este fenómeno se denomina cremación. Los eritrocitos y otras células del cuerpo pueden ser dañadas o destruidas si se exponen a líquidos hipotónicos e hipertónicos. Por esta razón la mayoría de las soluciones intravenosos (SI) que se infunden en la sangre de pacientes son isotónicas.
Difusión a través de la bicapa lipidica
Las moléculas hidrófobas apolares se difunden a través de la bicapa de la membrana plasmática dentro y fuera de la célula. Dichas moléculas comprenden oxigeno, dióxido de carbono, gases de nitrógeno, ácidos grasos, esteroides y vitaminas liposolubles (A, E, D y K); alcoholes y amoniacos. En virtud de que la membrana plasmática es muy permeable a todas estas sustancias, ellas no contribuyen a la presion osmótica ni a la tonicidad de los líquidos corporales. La difusión a través de la bicapa lipidica es importante para el movimiento del oxigeno y el dióxido de carbono entre la sangre y las células corporales y entre la sangre y el aire dentro de los pulmones durante la respiración. También constituye una ruta para la absorción de algunos nutrientes y la excreción de los desechos de las células, actividades que contribuyen a la homeostasis.
Difusión a través de los canales de la membrana
La mayor parte de los conductos de la membrana son canales ionicos, que permiten el paso de pequeños iones inorgánicos, los cuales son demasiado hidrófilos para penetrar el interior apolar de la membrana plasmática. Cada ion solo puede atravesar la membrana por los sitios donde hay canales específicos que permiten su paso. En las membranas plasmaticas típicas, el mayor numero de canales ionicos corresponde a los que son selectivos para K+ (iones potasio) o Cl- (de cloro); hay menos disponibles para Na+ (de sodio) o Ca2+ (de calcio). La difusión de solutos a través de los canales generalmente es mas lenta que la que se efectúa por la bicapa lipidica, porque los canales ocupan un área superficial total menor en la membrana. No obstante, la difusión a través de los canales es un proceso muy rápido: mas de un millón de iones potasio pueden pasar a través de los canales de K+ ¡en un segundo!
Muchos canales ionicos están abiertos de manera permanente, sin embargo otros tienen compuertas una parte del canal de proteína actiua como "puerta", al cambiar su forma de una manera que abre el poro y otra que lo cierra (fig. 0.3.9). Algunas compuertas alternan de manera aleatoria las posiciones abierta y cerrada, en tanto que otras se encuentran reguladas por cambios eléctricos o químicos dentro y fuera de la célula. Cuando las puertas se abren los iones salen o entran en dirección del sitio donde es menor su gradiente electroquímico.
Las membranas plasmáticas de los diversos tipos de células pueden poseer menos o mas canales iónicos y, por lo tanto, tener diferentes permeabilidades a los distintos iones. El numero total de estos conductos, que hay para un soluto especifico, también se encuentra regulado por señales dentro y fuera de la célula.
Difusión facilitada
Varios solutos, demasiado polares o que poseen cargas muy elevadas para atravesar la bicapa y que también resultan muy grandes para cruzar la membrana por los canales, pasan esta barrera por difusión facilitada. Originalmente se pensó que este proceso se llevaba a cabo por medio de una proteína portadora que se una a un soluto y atravesaba con el la membrana. No obstante, el nombre es poco adecuado porque el proceso no corresponde al de difusión en el sentido estricto del término. En vez de ello, un soluto se une a un transportador especifico en un lado de la membrana y es liberado en el lado opuesto; después el transportador cambia su forma. Por este motivo algunos investigadores abogan porque se cambie el nombre del proceso por el de transporte facilitado. Sin embargo, al igual que en la difusión, el resultado neto consiste en el movimiento de la zona de mayor concentración a donde esta es menor. Lo anterior ocurre porque con mayor frecuencia el soluto se une al transportador en el lado de la membrana con mayor concentración antes de ser trasladado al sitio donde es menor. Una vez que se llega al equilibrio entre ambos lados, las moléculas disueltas se unen al transportador en el citosol y se desplazan hacia el liquido extracelular con la misma rapidez con que se enlazan al transportador en la fase fuera de la célula para entrar al citosol. Por ende, la velocidad de la difusión facilitada se halla determinada por el gradiente de concentración en sentido descendente entre los dos lados de la membrana. La cantidad de transportadores disponibles para la difusión facilitada en la membrana plasmática tiene un limite superior llamado transporte máximo, que es la mayor velocidad a la cual puede ocurrir la difusión facilitada. Una vez que todos los transportadores se encuentran ocupados se llega al transporte máximo, y el mayor incremento en el gradiente de concentración no ocasiona que la difusión facilitada se lleve a cabo con mayor rapidez. Por lo tanto este proceso muestra saturación.
Entre los solutos que pasan a través de la membrana por difusión facilitada están la glucosa, la galactosa, la urea, la fructosa y algunas vitaminas. La glucosa entra en muchas células corporales de la siguiente manera (fig. 0. 3.10):
1-.La molécula de glucosa se une a una proteína transportadora denominada GluT en la superficie extracelular de la membrana.
2-.Glut inicia el proceso por el cual modifica su conformación.
3-.Glut libera la glucosa en el otro lado de la membrana.
Una vez que la glucosa entra en la célula por difusión facilitada, una enzima llamada cinasa se enlaza a un grupo fosfato para producir un molécula distinta (conocida como glucosa-6-fosfato). Esta reacción hace que la glucosa se mantenga en niveles bajos de concentración. intracelular, por lo que su gradiente siempre favorece la difusi6n facilitada de esta molécula hacia dentro, y no hacia afuera de las células.
Con frecuencia se regula la permeabilidad selectiva de la membrana plasmática para lograr la homeostasis. Por ejemplo, la hormona insulina promueve la inserción de muchas copias de un tipo determinado de transportador de glucosa dentro de la membrana plasmática de ciertas células. Así el efecto de la hormona insulina consiste en incrementar el transporte máximo por difusión facilitada de la glucosa. Al haber mas transportadores disponibles las células del cuerpo pueden tomar con mayor rapidez la glucosa de la sangre.
Transporte activo
Algunos solutos cargados o polares que deben entrar o salir de las células no pueden cruzar la membrana plasmática por ninguna de las formas de transporte pasivo, porque necesitan moverse en sentido ascendente en contra de su gradiente de concentración. Dichas sustancias solo pueden cruzar la membrana por transporte activo, el cual es mediado por un proceso en el que las proteínas transportadoras requieren energía para realizar su movimiento "cuesta arriba". Para ello se utilizan dos fuentes de energía: 1) la que proviene de la hidrólisis del ATP, considerada la fuente de transporte activo primario, y 2) la energía almacenada en un gradiente de concentración ionica constituye la fuente de transporte activo secundario. Al igual que la difusion facilitada, el proceso de transporte activo tiene capacidad máxima y un punto de sa-turacion. Los solutos que se transportan de manera selectiva a través de la membrana plasmática comprenden diversos iones como Na+, K+, Ca2+, I-, y Cl-; aminoacidos y monosacaridos. (Debe recordarse que algunas de estas sustancias también cruzan la membrana por difusión o difusión facilitada cuando la célula dispone de canales y transportadores adecuados.)
Transporte activo primario
En el transporte activo primario, la energía derivada de la hidrólisis del ATP cambia la forma de la proteína transportadora, la cual bombea una sustancia a través de la membrana plasmática en contra de su gradiente de concentración. Mas aun, las proteínas transportadoras que realizan este traslado con frecuencia son llamadas bombas. Una célula típica gasta 40% del ATP que genera en el transporte activo primario. Los medicamentos que interrumpen la producción de ATP —por ejemplo el veneno cianuro son letales porque detienen el transporte activo de las células a través del cuerpo.
El mecanismo de transporte activo primario de mayor prevalencia expulsa los iones sodio (Na+) de la célula e introduce los de potasio (K+). Este transportador se denomina bomba de Na+/K+ o, de manera mas sencilla, bomba de sodio, en virtud de los iones específicos que mueve. Todas las células poseen cientos de estas bombas en su membrana plasmática. Dado que una porcion de la bomba de Na+ /K+ actúa como una enzima que hidroliza el ATP (un ATPasa), tambien se le llama bomba de Na+/K+ ATPasa. Una de sus funciones consiste en mantener baja la concentración de iones sodio en el citosol al bombearlos al liquido extracelular en contra del gradiente de concentración de Na+. De manera coincidente la bomba de sodio moviliza los iones potasio contra el gradiente de concentración de este elemento. Como K+ y Na+ se filtran lentamente de regreso a través de la membrana plasmática hacia el sitio donde sus gradientes electromecánicos son menores por transporte pasivo o transporte activo secundario la bomba de sodio debe actuar ininterrumpidamente a fin de mantener baja la concentración de los iones Na y altos los niveles de los iones potasio en el citosol. En la figura 0-3.11 se expone gráficamente esta explicación sobre la bomba de sodio.
1-En el citosol tres iones sodio (Na+) se unen a la proteína bomba.
2-Una vez que se produce el enlace de Na desencadena la hidrólisis del ATP en ADP, una reacción que también une un grupo fosfato a la proteína bomba, la cual modifica su forma a fin de liberar los tres Na en el liquido extracelular. Ahora la forma de la proteína bomba propicia el enlace de los iones potasio en este liquido.
3-La unión de los iones potasio hace que la bomba libere el grupo fosfato, lo cual de nueva cuenta hace que cambie su forma.
4-Conforme la bomba recupera su forma original dos K+ son liberados en el citosol. En este punto, la bomba se encuentra lista, una vez más, para enlazarse a Na y repetir el ciclo.
Además de mantener bajo el nivel extracelular de Na+ y alto el de K+, se requiere la operación continua de la bomba para mantener el volumen normal de la celula. Es necesario recordar que la presion osmótica de una solución es proporcional a la concentración de sus partículas de solutos que no pueden penetrar la membrana. Dado que los iones sodio se difunden en la célula o entran por medio del transporte activo secundario, de inmediato son bombeadas hacia afuera, y es como si nunca hubieran entrado. En efecto, los iones sodio se comportan como si no penetraran la membrana. Por lo tanto, contribuyen de manera importante a la presion osmótica del liquido extracelular. Una condición similar tiene vigencia para los iones potasio, que aportan la mitad de la presion osmótica en el citosol. Al ayudar a mantener la presion osmótica en ambos lados de la membrana plasmática, la bomba de sodio asegura que la célula nunca se contraiga ni hinche por el movimiento del agua que sale o entra por osmosis.
Transporte activo secundario
En el transporte activo secundario la energía almacenada del gradiente de concentración del ion sodio o de hidrogeno se usa para conducir otra sustancia a través de la membrana en contra de su propio gradiente de concentración. Dado que el transporte activo primario establece el gradiente de Na+ o H+ , el secundario utiliza de manera indirecta la energía obtenida de la hidrólisis del adenosintrifosfato.
La bomba de sodio mantiene alto el gradiente de concentración de Na+ a través de la membrana plasmática. Como resultado de ello, los iones sodio tienen almacenada energía potencial, como sucede con el agua contenida por un dique. De conformidad con esto, si existe una ruta para que los Na+ se filtren de regreso, parte de la energía almacenada puede ser convertida en energía cinética (energía de movimiento) y ser usada para el transporte de otras sustancias en contra de su gradiente de concentración. En esencia, las proteínas del transporte activo secundario aprovechan la energía del gradiente de concentración de Na+ al proporcionar vías de acceso faci-les para que los iones sodio se filtren en las células. La proteína funciona como un transportador doble al unirse simultanea-mente a Na y otra sustancia, entonces cambia su forma para que ambas sustancias atraviesen la membrana al mismo tiempo.
Las membranas plasmáticas contienen varios contratransportadores y contra transportadores que son energizados por el gradiente de Na+ (fig. 0.3.12a). En casi todas las células del cuerpo, la concentración de iones calcio (Ca2+ ) es baja en el citosol porque los contra transportadores de Na+ /Ca2+ permiten que los iones sodio entren en la celula en tanto expulsan los de calcio. Del mismo modo, los contra transportadores de Na+/H+ ayudan a regular el pH del citosol (concentración de H ) al aprovechar el gradiente de Na para expeler el exceso del ion hidrogeno. Por otra parte, la glucosa y los aminoácidos de la dieta son introducidos dentro de las células que recubren el intestino delgado por los cotransportadores de Na+-glucosa y Na aminoácido (fig. 0.3.12b). En cada caso los iones sodio se mueven hacia el lado menor del gradiente de concentración, mientras que los solutos acoplados se desplazan en sentido ascendente contra su gradiente de concentración. Debe recordarse que todos esos cotransportadores y contratransportadores pueden realizar su trabajo porque las bombas de sodio mantienen baja la concentración de Na+ en el citosol.
APLICACIÓN CLÍNICA
Digitalis
Debido a que refuerza los latidos cardiacos, a me-nudo se administra digitalis a los padentes con insuficiencia cardiaca, un trastorno en el que se debilita el bombeo del corazón. El digitalis ejerce su efecto al reducir el bombeo del sodio, lo que permite que se acumulen mas Na+ dentro del músculo cardiaco. Como resultado de lo anterior, una pequeña concentración de Na+ cruza la membrana, con lo cual disminuyen los contratransportadores de Na+/Ca2+. De esta manera permanecen mas iones calcio dentro de la célula del músculo cardiaco. El ligero incremento en el nivel de Ca2+ en el citosol de estas células incrementa la fuerza de sus contracciones y por ende refuerza los latidos del corazón. Como ilustra este ejemplo, el equilibrio entre las concentraciones de Na+ y Ca2+ en el citosol y en el liquido extracelular es crucial para el funcionamiento normal de las células musculares.
Transporte vesicular
Una vesícula es un pequeño saco esférico que brota en la membrana y luego se desprende de ella. La permeabilidad de la cubierta vesicular es selectiva y contiene pequeñas cantidades de liquido mas partículas disueltas o suspendidas. Las vesículas transportan sustancias de una estructura u otra dentro de las células, las toman del liquido extracelular y las liberan en el intracelular. Mas adelante en este capitulo se explica como se lleva a cabo el transporte entre las estructuras de la célula, una vez que el estudiante comprenda mejor la anatomía de esta unidad vital. Por el momento concentraremos nuestra atención en los dos principales tipos de transporte vesicular entre una célula y el liquido extracelular: 1) endocitosis, en el cual los materiales se mueven dentro de la célula en una vesícula formada a partir de la membrana plasmática, y 2) exocitosis en el que los materiales se mueven fuera de la célula por fusión de las vesículas con la membrana plasmática. Tanto la endocitosis como la exocitosis requieren la energía proporcionada por ATP.
Endocitosis
En la endocitosis el material que se introduce en la célula esta rodeado por un área de la membrana plasmática, que se desprende dentro de la célula para formar una vesícula, la cual contiene los materiales ingeridos. En esta sección se consideran tres tipos de endocitosis: mediada por receptores, pinocitosis y fagocitosis.
Endocitosis mediada por receptores. Se trata de un proceso altamente selectivo por el cual la célula se une a ciertos ligandos. La vesícula se forma luego de que una proteína receptora de la membrana plasmática reconoce y se enlaza a un ligando especifico en el liquido extracelular. Mediante este mecanismo las células captan transferían (una proteína que transporta hierro en la sangre), algunas vitaminas, lipoproteínas de baja densidad (LBD), anticuerpos, ciertas hormonas y otras macromoléculas o partículas especificas. La endocitosis mediada por receptores se efectua de la siguiente manera (fig.0.3.13).
1 Fijación. En el lado extracelular de la membrana plasmática, un ligando se une a un receptor especifico para formar un complejo ligando receptor. Los receptores son proteínas integras de la membrana que se encuentran concentradas en regiones particulares de esta, denominadas poros recubiertos de datrina, en virtud de que la membrana esta recubierta en su lado citoplasmático por esta proteína periférica. La interacción de la clatrina con el complejo ligando-receptor ocasiona que la membrana se invagine (se pliegue hacia dentro).
2 Formación de la vesícula. Los bordes de la membrana alrededor de la zona de revestimiento de clatrina se fusionan y cierran una porción de la primera, de lo que resulta la vesícula recubierta de clatrina que contiene el complejo ligando-receptor.
3 Eliminación de la cubierta. Casi inmediatamente después de que se ha formado, la vesícula pierde su cubierta de clatrina y queda sin recubrimiento (vesícula no recubierta).
4 Fusión con el endosoma primario. Varias vesículas no recubiertas se fusionan con un endosoma primario, una vesícula que hace las veces de compartimiento. Dentro de cada endosoma primario, los ligandos se separan de sus receptores. Finalmente, el contenido de este endosoma se recicla en el plasma o se transporta a un lisosoma, que es un organelo donde ocurre la descomposición.
5 Reciclado de receptores. Muchos receptores dentro del endosoma primario entran en una vesícula transportadora, la cual se desprende del endosoma primario y devuelve los receptores a la membrana plasmática. También se reintegra la clatrina a la superficie interna de dicha membrana.
6 Descomposición en los lisosomas. Los ligandos (y sus receptores) destinados a su descomposición en los lisosomas, son transportados por una o mas vesículas transportadoras desde el endosoma primario al endosoma secundario. Este ultimo forma otras vesículas transportadoras que se fusionan con los lisosomas. Dentro de estos se encuentran las enzimas digestivas que desdoblan una variedad de moléculas grandes, como las proteínas, los polisacaridos, los lípidos y los ácidos nucleicos.
7 Transcitosis. En algunos casos, los ligandos y los receptores entran en transcitosis, un proceso por el cual son trasladados a través de la célula dentro de las vesículas transportadoras que realizan la exocitosis en el lado opuesto. Conforme las vesículas transportadoras se fusionan con la membrana plasmática vierten su contenido en el liquido extracelular. La transcitosis se lleva a cabo con mayor frecuencia a través de las células endoteliales que recubren los vasos sanguíneos y constituye un medio para movilizar los materiales entre el plasma sanguíneo y el liquido intersticial.
APLICACIÓN CLÍNICA
Virus y endocitosis mediada por receptores
La endocitosis mediada por receptores normalmente permite importar los materiales necesarios, sin embargo algunos virus aprovechan este mecanismo para entrar e infectar las células. Por ejemplo, el virus de la inmunodeficiencia humana (VIH), causante del síndrome de inmunodeficiencia adquirida (sida), entra en las células al unirse a un receptor, llamado CD4, de la membrana plasmática. Este receptor forma parte de la superficie de algunas células, como los leucocitos denominados células T cooperadoras. Después de que se enlaza a CD4, el VIH entra en la célula mediante la endocitosis mediada por un receptor.
Fagocitosis. La fagocitosis es una forma de endocitosis por la cual grandes partículas sólidas como bacterias enteras c virus son captados por la célula (fig. 0.3.14). Comienza cuando la partícula se une a un receptor de la membrana plasmática lo que provoca que la célula extienda proyecciones de su membrana y citoplasma denominadas seudopodes. Los seudopodos rodean la partícula fuera de la célula y la membrana se fusiona para formar una vesícula llamada fagosoma que entra en el citoplasma. El fagosoma se une con uno c mas lisosomas y el material ingerido es desdoblado por la; enzimas lisosomales. En la mayoría de los casos cualquier material no digerido que entra en el fagosoma, el cual ahore recibe el nombre de cuerpo residual, es llevado de regreso por exocitosis al liquido extracelular donde es liberado.
La fagocitosis solo ocurre en ciertas células del cuerpo denominadas fagocitos, que son una clase de células que engullen y destruyen bacterias y otras sustancias extrañas. Los fagocitos comprenden ciertas clases de leucocitos y macrófagos, que se hallan presentes en la mayoría de los tejidos. El proceso de fagocitosis es un mecanismo vital de defensa que ayuda a proteger al cuerpo de las enfermedades.
Pinocitosis. Este término designa una forma de endocitosis que consiste en la captación no selectiva de diminutas gotas de liquido extracelular (fig. 0.3.15). No participan proteínas receptoras; todos los solutos disueltos en el liquido extracelular son elevados dentro de la célula. En la pinocitosis la membrana plasmática forma una vesícula pinota que contiene una gota de liquido extracelular; luego esta se desprende de la membrana y entra en el citosol. Una vez en el interior de la célula se fusiona con uno o mas lisosomas, donde las enzimas desdoblan los materiales engullidos. Al igual que en la fagocitosis, los materiales no digeridos se acumulan en un cuerpo residual. En la mayoría de los casos este se fusiona con la membrana plasmática y expele su contenido fuera de la célula por exocitosis; la mayoría de las células del cuerpo realizan la pinocitosis.
Exocitosis
Según se ilustra en las figuras 0.3.13 a 0.3.15, la exocitosis comprende el movimiento de los materiales fuera de la célula, en vesículas que se fusionan con la membrana plasmática. El material expulsado puede ser algún producto de desecho o material útil secretado. Todas las células llevan a cabo el proceso de exocitosis, pero resulta especialmente importante en dos tipos de células: las nerviosas, que liberan sustancias llamadas neurotransmisores, y las secretoras, que producen o secretan enzimas digestivas u hormonas. Durante la exocitosis en las células secretoras, las vesículas en-vueltas por la membrana, denominadas vesículas secretoras, se forman dentro de la célula, se fusionan con la membrana plasmática y liberan su contenido en el liquido extracelular.
Las partes de la membrana plasmática que se pierden durante la endocitosis son recuperadas y recicladas por la exocitosis. El equilibrio entre ambos procesos mantiene el área superficial de la membrana plasmática relativamente constante. El intercambio membranoso resulta muy extenso en ciertas células. Por ejemplo, las secretoras del páncreas reciclan una cantidad de membrana plasmática igual a la superficie entera de la célula cada 90 minutos.
En el cuadro 0.3.1 se resumen los procesos mediante los cuales son transportados los materiales dentro y fuera de las células.
CITOPLASMA
OBJETIVO
• Describir la estructura y función del citoplasma, citosol y organelos.
El citoplasma se puede dividir en dos componentes: 1) citosol y 2) organelos.
Citosol
El citosol es la parte liquida que circunda a los organelos (vease la fig. 0.3.1) y constituye aproximadamente el 55% del volumen total de la célula. Si bien varia en su composición y consistencia de una parte a otra de la célula, contiene de 75 a 90% de agua mas componentes disueltos y suspendidos. Entre ellos hay varios iones, glucosa, aminoácidos, ácidos grasos, proteínas y lípidos, ATP y productos de desecho. También están presentes varias moléculas organicas que se agregan en masas y son almacenadas. Dichas moléculas pueden aparecer y desaparecer varias veces a lo largo de la vida de las células. Entre los ejemplos es posible mencionar gotitas de lipidos, que contienen trigliceiidos y cúmulos de moléculas de glucogeno, denominadas granulos de glucogeno (vease la fig.0. 3.1).
El citosol es el sitio donde se efectúan muchias reacciones químicas que son necesarias para la vida celular. Por ejemplo, las enzimas en el citosol catalizan numerosas reacciones químicas; como resultado de ellas, se produce energía que es capturada y aprovechada para las actividades de las células. Mas aun, esas reacciones proporcionan los bloques de construcción para mantener la estructura, función y crecimiento celulares.
Organelos
Como se expuso previamente, los organelos son estructuras especializadas que tienen formas características y realizan funciones específicas en el crecimiento, mantenimiento y reproducción de las células. Muchas reacciones químicas ocurren en una célula simultáneamente, sin embargo hay poca interferencia entre los distintos tipos de reacciones porque ocurren en diferentes organelos. Cada tipo de estos tiene un conjunto de enzimas características propias que desempeñan reacciones especificas, y cada uno se encuentra en un compartimiento funcional, donde ocurren procesos fisiológicos particulares. No obstante, los organelos con frecuencia colaboran entre ellos para mantener la homeostasis.
Los diversos tipos de organelos difieren en las distintas celulas, según la función que esta tenga encomendada. Los organelos no membranosos carecen de esta cubierta por lo que se hallan en contacto directo con el citosol. Entre los ejemplos de ellos podemos citar el citoesqueleto, centrosomas, cilios, flagelos y ribosomas. Por otra parte, los organelos membranosos están envueltos por una o dos bicapas lipídicas (semejantes a la membrana plasmática) que separan su interior del citosol. Ejemplos de los organelos membranosos son el retículo endoplasmico, el complejo de Golgi, la mitocondria, los lisosomas y peroxisomas. Aunque el núcleo es un organelo membranoso se estudia aparte, dado que tiene especial importancia en el control de la vida celular.
El citoesqueleto
El citoesqueleto constituye una red de distintas clases de filamentos proteinicos que se extienden por el citosol (fig. 0.3.16). El citoesqueleto proporciona el marco estructural de la célula y sirve como un andamio que ayuda a determinar la forma de una célula y a organizar su contenido. También controla los movimientos celulares, entre ellos el transporte interno de organelos y algunas sustancias químicas, el movimiento de los cromosomas durante la división celular y el de toda la unidad como fagocito. Aunque su nombre conlleva la noción de rigidez, constantemente se reorganiza conforme la célula se mueve y cambia de forma, como en la división celular. El citoesqueleto se halla integrado por tres filamentos. En orden creciente, según su diámetro, estas estructuras son microfilamentos, filamentos intermedios y microtubulos.
Microfilamentos. Son los elementos mas delgados del citoesqueleto. La mayor parte de ellos están formados por la proteína actina y se encuentran concentrados en la periferia de la célula {vease fig. 0.3.16a y b). Desempeñan dos funciones generales: movimiento y apoyo mecánico. En lo que ante al movimiento, los microfilamentos participan en la contracción muscular, división y locomoción de la célula, como sucede durante la migradon de las células embrionicas durante el desarrollo, la invasión de tejidos por los leucocitos que combaten las infecciones o la migración de las células cutáneas en la curación espontánea de una herida.
Los microfilamentos proporcionan mucho del apoyo mecánico que le confiere fuerza y forma a las células. Anclan el citoesqueleto a las proteínas integras en la membrana plasmática. Asimismo proporcionan apoyo mecánico para las extensiones celulares denominadas micro vellosidades, que son proyecciones microscópicas de la membrana plasmática. Dentro de las micro vellosidades se encuentra un conjunto de microfilamentos paralelos que sirven de apoyo y las adhieren al citoesqueleto (vease fig. 0.3.16a ).
Dichas micro vellosidades son abundantes en las superficies de las células que participan en la absorción, como las epiteliales que revisten el intestino delgado. Algunas de ellas se extienden hasta la membrana plasmática y ayudan a que las células se mantengan unidas entre si o al material extracelular.
Filamentos intermedios. Como su nombre lo sugiere, los filamentos intermedios son un poco mas gruesos que los microfilamentos, pero mas linos que los microtubulos (vease fig. 0.3.1603_16.jpg). Pueden estar formados por distintas clases de proteínas; que son excepcionalmente resistentes. Se encuentran en partes de la celula sujetas a tensión mecánica y también pueden anclar organelos, entre ellos al núcleo (vease fig. 0.3.16a).
Microtubulos. Son los componentes mas grandes del citoesqueleto, constituyen tubos largos huecos y sin ramificaciones, formados principalmente por una proteína denominada tubulina. Un organelo llamado centrosoma (que se describe en el siguiente párrafo) sirve como sitio de inicio para el ensamble de los microtubulos, los cuales crecen desde el centromero hasta la periferia de la célula (vease la fig. 0.3.16a ). Los microtubulos ayudan a determinar la forma y función de la célula en el transporte intracelular de los organelos, como las vesículas secretoras y la migración de los cromosomas durante la division celular. También participan en el movimiento de las proyecciones celulares especializadas como son los cuerpos ciliares y flagelares. Las proteínas motoras denominadas cinesinas y dineinas se encargan de energizar los movimientos en los cuales participan los microtubulos. Estas proteínas actúan como motores en miniatura que impulsan sustancias y organelos a lo largo de los microtubulos, como una locomotora tira o empuja los carros de ferrocarril.
Centrosoma
El centrosoma, que se ubica cerca del núcleo, consta de dos componentes: el área pericentriolar y los centríolos (fig.03.17a). El área pericentriolar constituye una región del citosol compuesta por una densa red de pequeñas fibras de proteína. Esta zona es un centro de organización para el huso mitótico, que desempeña un papel básico en la división celular y en la formación de microtubulos en las células indivisibles. Dentro del área pericentriolar hay un par de estructuras cilíndricas llamada centríolos, cada una de las cuales esta compuesta por nueve racimos o haces de tres microtubulos (tripletes) dispuestos en forma circular, una distribución denominada disposición 9+0 (fig. 0.3.17b). El 9 alude a los nueve haces de microtubulos y el 0 a la falta de microtubulos en el centro. El eje largo de un centríolo se halla en un . Angulo recto con respecto al mismo eje del otro (fig. 0.3.17c). Los centríolos participan en la formación o regeneración de cilios y flagelos.
Cilios y flagelos
Los microtubulos son los componentes funcionales y estructurales dominantes en cilios y flagelos; constituyen proyecciones móviles de la superficie celular. En el cuerpo humano las células que se hallan firmemente ancladas en su lugar usan los cilios para mover los líquidos por su superficie; las células móviles, como los espermatozoides, usan los flagelos para impulsarse en un medio liquido.
Los cilios son numerosos filamentos cortos que se ex-tienden desde la superficie celular (vease fig. 0.3.1). Cada uno (de forma singular) contiene un centro de microtubulos en-vueltos por la membrana plasmática (fig.0. 3.18a). Los microtubulos están dispuestos de tal manera que un par en el centro se encuentra rodeado por nueve haces de dos microtubulos (dobletes), una disposición denominada 9+2. Cada cilio se halla fijo a un cuerpo basal justo debajo de la membrana plasmática. Un cuerpo basal es similar en estructura a un centríolo. En efecto, se considera que los cuerpos básales y los centríolos constituyen dos diferentes manifestaciones funcionales de la misma estructura. La función de estos cuerpos consiste en iniciar el ensamblaje de cilios y flagelos. Un cilio despliega un patrón semejante a un remo, el cual es relativamente rígido durante el golpe de impulso, pero relativamente flexible en el de recuperación (fig. 0. 3.18c).El movimiento coordinado de numerosos cilios en la superficie de una célula asegura el movimiento constante del liquido a lo largo de la superficie celular. Por ejemplo, muchas células del tracto respiratorio poseen cientos de cilios que les ayudan a barrer lejos de los pulmones las partículas extrañas atrapadas en el moco. La nicotina de los cigarrillos tiende a paralizar este movimiento. Por esta razón, la tos de los fumadores con frecuencia remueve las partículas extrañas de los conductos respiratorios. Las células que revisten las trompas uterinas (falopianas) también son cilios que barren los óvulos hacia el útero.
Los flagelos son similares en estructura a los cilios pero mucho mas largos. Con frecuencia mueven una célula entera. Un flagelo genera movimiento hacia adelante a lo largo de su eje mediante un culebreo rápido conforme a un patrón oscilante (fig. 0.3.18c). El uúnico ejemplo de un flagelo en el cuerpo humano son los espermatozoides caudados, que impulsan al esperma hacia su encuentro con el óvulo.
Ribosomas
Son sitios donde se sintetizan proteínas. Estos diminutos organelos constituyen haces de cadenas ARN ribosómico (ARNr) y muchas proteínas también ribosómicas. Los ribosomas reciben este nombre porque tienen un alto contenido de ácido ribonudeico. Desde el punto de vista estructural, constan de dos subunidades, una aproximadamente de la mitad del tamaño de la otra (fig. 0.3.19). Ambas subunidades están separadas del nucleolo, un cuerpo esférico dentro del núcleo. Una vez que son producidas salen del núcleo y se unen al citosol.
Algunos ribosomas llamados libres no se hallan unidos a ninguna estructura en el citoplasma. Ante todo, sintetizan las proteínas que se usan en el interior de la célula. Otros, denominados ribosomas unidos a la membrana, están adheridos a la membrana nuclear y a una membrana con muchos pliegues llamada retículo endoplasmico. Los ribosomas sintetizan proteínas destinadas a su inserción en la membrana plasmática o para su exportación fuera de la célula. Asimismo, los ribosomas se encuentran dentro de la mitocondria donde se encargan de sintetizar las proteínas mitocondriales. En ocasiones de 10 a 20 ribosomas se conjuntan para formar una estructura semejante a una soga llamada polirribosoma.
Retículo endoplasmico
El retículo endoplasmico o RE es una red de membranas que forman sacos aplanados o túbulos denominados cisternas (cavidades) (fig. 0.3.20). Se extiende de la membrana que envuelve al núcleo (cubierta nuclear), a la cual esta conecta-do, por todo el citoplasma. El RE es tan extenso que constituye mas de la mitad de las superficies membranosas dentro del citoplasma de la mayoría de las células.
Las células contienen dos distintas formas de RE (interrelacionadas), que difieren en estructura y función. La membrana del retículo endoplasmico rugoso es una continuación de la membrana nuclear y habitualmente se pliega en una serie de sacos aplanados. La superficie externa de la membrana rugosa del RE esta unida a los ribosomas, los sitios donde se sintetizan las proteínas, las cuales al ser sintetizadas entran en las cisternas para su procesamiento y distribución. En algunos casos las enzimas dentro de las cisternas fijan las proteínas a los carbohidratos para formar glucoproteinas. En otros, unen proteínas con fosfolipidos también sintetizados por el retículo endoplasmico rugoso. Estas moléculas pueden ser incorporadas a las membranas de los organelos o a la membrana plasmática. Por ende, puede afirmarse que el RE es la fabrica donde se sintetizan las proteínas secretadas y las moléculas de la membrana.
El retículo endoplasmico liso se extiende desde la membrana rugosa del RE para formar una red de túbulos membranosos (vease fig. 0.3.20). A diferencia del retículo endoplasmico rugoso, el liso no tiene ribosomas en la superficie externa de su membrana. Sin embargo contiene enzimas únicas que le confieren mayor diversidad funcional que la del RE liso. Aunque no sintetiza proteínas si lo hace con los fosfolipidos, como el RE rugoso. Además, sintetiza grasas y esteroides, como estrógenos y testosterona. En las células hepáticas, las enzimas del RE liso contribuyen a la liberación de glucosa en el torrente sanguíneo e inactiva o desintoxica medicamentos y otras sustancias potencialmente peligrosas, por ejemplo el alcohol (etilo). En las células musculares, los iones calcio liberados del retículo sarcoplasmico, una forma de RE liso, activan los procesos de contracción.
Complejo de Golgi
La mayoría de las proteínas sintetizadas por los ribosomas que se unen al RE rugoso finalmente son transportadas a otras regiones de la célula. El primer paso en la vía de transporte es un organelo llamado complejo de Golgi, que esta formado por tres a 20 sacos membranosos aplanados con bordes prominentes denominados cisternas (fig.0. 3.21). Estas con frecuencia son curvadas, lo que les confiere la forma de una copa. La mayor parte de las células solo tienen un complejo de Golgi, aunque algunas poseen muchos. Dicho complejo es mas extenso en las células que secretan proteínas en el liquido extracelular. Este hecho es clave para la función del organelo en la célula.
Las cisternas en los extremos opuestos del complejo de Golgi difieren entre si de tamaño, forma, contenido, actividad enzimática y cantidad de vesículas (como se describe en los párrafos siguientes). La entrada convexa o cara si se forma por sisternas que se encuentran pegadas a la membrana rugosa. La salida cóncava o cara trans esta integrada por cisternas contiguas a la membrana plasmática. Las cisternas entre las caras de entrada y de salida reciben el nombre de cisternas intermedias.
La entrada o cara cis, las cisternas intermedias y la cara de salida del complejo de Golgi contienen enzimas diferentes que les permiten a cada una modificar, ordenar y empacar las proteínas para que puedan ser transportadas a su destine Por ejemplo, la cara de entrada recibe y modifica las proteínas que produce el retículo endoplasmico rugoso. Las cisternas intermedias apiaden azucares a las proteínas y lípidos para formar glucoproteinas. La cara de salida modifica adicional-mente las moléculas y luego las clasifica y empaca para que sean transportadas a sus destinos finales.
Las proteínas llegan y pasan a través del complejo de Golgi mediante una serie de intercambios entre las vesículas (fig. 0. 3.22).
1-Las proteínas sintetizadas por los ribosomas del retículo endoplasmico rugoso (RE) se encuentran rodeadas por una porción de membrana RE, que tarde o temprano brota de la superficie membranosa para formar una vesícula de transporte.
2-Las vesículas de transporte se desplazan hacia el cis de entrada del complejo de Golgi.
3-Tales vesículas se fusionan con la cara de entrada del complejo de Golgi y liberan las proteínas dentro de la cisterna.
4-Las proteínas son modificadas y se desplazan de la cis de entrada a una de las cisternas intermedias por medio de las vesículas de transferencia que brotan en los bordes de la cisterna. Las enzimas de las cisternas intermedias modifican las proteínas para formar glucoproteinas, glucolipidos y lipoproteínas.
5-Los productos de las cisternas intermedias se mueven por medio de las vesículas de transferencia hacia el interior de la cara de salida.
6-Dentro de la cara de salida, se modifican aun mas los productos y luego son clasificados y empacados.
7- Algunas de las proteínas procesadas dejan la cara de salida en las vesículas secretoras, las cuales las llevan a la membrana plasmática para que se incorporen a ella, donde son descargadas por exocitosis en el liquido extracelular.
8-Otras proteínas procesadas dejan la cara de salida en vesículas que vierten su contenido en la membrana plasmática para que se incorporen a esta. Al hacer esto, el complejo de Golgi añade nuevos segmentos de membrana plasmática conforme se pierden los segmentos existentes y modifica el numero y distribución de las moléculas de dicha membrana.
9-Finalmente, algunas de las proteínas procesadas dejan la cara de salida dentro de vesículas que reciben el nombre de vesículas de almacenaje. La principal vesícula de almacenaje es un lisosoma, cuya estructura y funciones se describen mas adelante.
APLICACIÓN CLÍNICA
Fibrosis quística J En algunas ocasiones la defidencia en la trayectoria
de una molécula ocasiona trastornos, como en el caso de la fibrosis quística, una enfermedad mortal hereditaria que afecta varios sistemas corporales. La proteína defectuosa producida por el gen mutado de la fibrosis quística no logra llegar a la membrana plasmática donde debería insertarse para ayudar a bombear iones cloro (CT) fuera de las células. Evidente-mente las proteínas que deberían ser bombeadas se fijan en el retículo endoplasmico o en el complejo de Golgi y nunca llegan a su destino correcto. El resultado es un desequilibrio en el transporte de liquido y iones a través de la membrana plasmática, lo que causa la formación de moco fuera de ciertos tipos de células. El moco acumulado obstaculiza las vías respiratorias de los pulmones, lo que a su vez ocasiona dificultad para respirar y evita que el páncreas secrete de manera adecuada enzimas digestivas, lo que causa trastornos digestivos.
Lisosomas
Los lisosomas son membranas envueltas en vesículas que se forman en el complejo de Golgi (fig. 0.3.23). Dentro de ellas hay hasta 40 clases de poderosas enzimas digestivas o hidroliticas que tienen la capacidad de descomponer una amplia variedad de moléculas. Las enzimas lisosomicas trabajan mejor en pH ácido y la única membrana lisosomica incluye bombas de transporte activo que mueven iones hidrogeno (H ) en los lisosomas. Por ende, el interior de estos tiene un pH de 5, que es 100 veces mas ácido que el pH del citosolico el cual es de 7. La membrana lisosomica tambien permite que los productos finales de la digestión, como los azucares y aminoácidos sean transportados dentro del citosol.
Así mismo, las enzimas lisosomicas son recicladas por la propia estructura de la célula. Un lisosoma puede engullir otro organelo, digerirlo y regresar sus componentes al citosol para que vuelvan a ser usados. De esta manera hay un reemplazo continuo de los organelos viejos. El proceso por el cual los organelos desechados son digeridos recibe el nombre de autofagia. Por ejemplo, una célula del hígado humano recicla aproximadamente la mitad de su contenido citoplasmico cada semana. En el proceso de autofagia, los organelos que serán digeridos son envueltos por una membrana derivada de retículo endoplasmico denominada autofagosoma; entonces la vesícula se fusiona con un lisosoma. Asimismo las enzimas lisosomicas pueden destruir a su propia célula mediante un proceso llamado autolisis, que ocurre en algunos trastornos patológicos; ellas también son responsables del deterioro de los tejidos que ocurre inmediatamente después de la muerte.
Aunque la mayoría de los procesos digestivos en los que participan los lisosomas ocurren en el interior de la célula, hay algunos casos en que las enzimas operan en la digestión extracelular. Un ejemplo es la liberación de las enzimas lisosomicas durante la fertilización. La cabeza de la célula espermatica suelta enzimas lisosomicas que ayudan al esperma a penetrar el óvulo.
APLICACIÓN CLÍNICA
Enfermedad de Tay Sachs
Algunos trastornos son causados por fallas lisosomicas. Por ejemplo, la enfermedad de Tay Sachs, que por lo común afecta a niños de Europa oriental descendientes de Ashkenazis, se trata de un trastorno hereditario que se caracteriza por la falta de una sola enzima Lisosomica, que normalmente desdobla o descompone una membrana gluco Lipida Llamada gangliosido GM2, que tiene especial prevalencia en las células nerviosas. Conforme los gangliosidos GM2 se acumulan, la función de Las células nerviosas se hace menos eficiente. Los niños con este problema de manera característica experimentan convulsiones y rigidez muscular. Gradualmente se quedan riegos, dementes, pierden La coordinación y finalmente mueren antes de los cinco años de edad. Se ha logrado identificar el gen de La enfermedad de Tay Sachs y una prueba cromosomica puede detectar si un adulto es portador del gen defectuoso.
Peroxisomas
Otro grupo de organelos similares en estructura a los lisosomas, pero mas pequeños, son los llamados peroxisomas [ver fig. 03.1). Aunque alguna vez se pensó que estos se formaban por brotes del retículo endoplasmico, en la actualidad por lo regular existe acuerdo en que se forman por la división de peroxisomas preexistentes.
Los peroxisomas contienen una o mas enzimas que pueden oxidar (remover los átomos de hidrogeno) varias sustancias orgánicas. Por ejemplo, sustancias como los aminoácidos y ácidos grasos son oxidados en los peroxisomas como parte del metabolismo normal. Además, las enzimas de los peroxisomas oxidan las sustancias toxicas como el alcohol. Un producto intermedio de la oxidación es el peroxido de hidrogeno (H2O2), un compuesto potencialmente toxico. Sin embargo, los peroxisomas también contienen una enzima llamada catalasa, que descompone el H2O2 En virtud de que la generación y degradación del peroxido de hidrogeno ocurre dentro del propio organelo, los peroxisomas protegen otras partes de la célula de los efectos nocivos del H2O2.
Mitocondria
En virtud de las actividades que realizan en la producción de ATP, las mitocondrias constituyen la "central energética" de la célula. Una célula puede tener unos pocos cientos o miles de mitocondrias. Las células con actividad fisiológica, como las de los músculos, hígado y riñones tienen un gran numero de mitocondrias porque usan ATP en grandes cantidades. Las mitocondrias por lo regular se localizan en la parte de la célula que tiene mayores requerimientos de energía, como entre las proteínas contráctiles del tejido muscular. Una mitocondria esta limitada por dos membranas, cada una de las cuales es similar en estructura a la membrana plasmática (fig. 0.3.24). La membrana mitocondrial externa es lisa., pero la membrana mitocondrial interna tiene una serie de pliegues o dobleces denominados crestas. La cavidad central Uena de liquido de una mitocondria delimitada por la membrana interna y las crestas es la matriz. Los pliegues complejos de las crestas proporcionan un área enorme para las reacciones químicas que son parte de la fase aeróbica de la respiración celular. Esas reacciones producen la mayor parte de ATP celular. Las enzimas que catalizan la respiración celular se localizan en la matriz y sobre las crestas.
Al igual que los peroxisomas, las mitocondrias se autorreplican, un proceso que ocurre cuando se incrementa la demanda de energía o antes de la división celular. Cada mitocondria tiene múltiples copias idénticas de una molécula circular de ADN que contiene 37 genes. Estos, junto con los genes del núcleo celular, controlan la producción de proteínas que forman los componentes mitocondricos. Dado que los ribosomas también se hallan presentes en la matriz de las mitocondrias, algunas síntesis de proteínas tienen lugar dentro de estas.
Aunque el núcleo de cada célula somática contiene genes tanto de la madre como del padre, los genes mitocondricos por lo general son heredados de la madre. La cabeza de un esperma (la parte que penetra y fertiliza al óvulo) normalmente carece de la mayoría de los organelos, como las mitocondrias, ribosomas, retículo endoplasmico y complejo de Golgi.
NÚCLEO
OBJETIVO
• Describir la estructura y función del núcleo.
El núcleo es una estructura de forma esférica u ovalada que comúnmente es la característica mas prominente de la célula (fig. 0.3.25). La mayoría de los cuerpos celulares tiene un solo núcleo, aunque algunos, como los eritrocitos maduro, no tienen ninguno. En contraste, las células del músculo esquelético y otras pocas células poseen varios núcleos. Una membrana de doble capa, denominada membrana nuclearr separa el núcleo del citoplasma. Ambas capas son lipidicas similares a la membrana plasmática. La membrana externa del núcleo es una continuación del RE rugoso y resulta similar en su estructura. La cubierta nuclear esta perforada por numerosos canales llamados poros nucleares (fig. 05.25c). Cada uno de ellos consta de una disposición de proteínas que circunda un canal central, el cual es unas 10 veces mas grande que el poro de un canal proteico en la membrana plasmática.
Los poros nucleares controlan el movimiento de las sustancias entre el núcleo y el citoplasma. Las moléculas pequeñas y los iones se difunden de manera pasiva a través de los canales acuosos abiertos en los poros.
No obstante, las moléculas mas grandes como el ARN (ácido ribonucleico) y las proteínas no pueden pasar a través de los poros nucleares por difusión. En vez de ello, lo hacen por medio de un proceso de transporte activo en el cual hay reconocimiento y traslado selectivo de las moléculas en una sola dirección. A medida que se lleva a cabo esta identificación y movimiento a través de los canales regulados, los poros nucleares se abren para acomodarlas. Esto permite que se lleve a cabo el transporte selectivo de proteínas al citosol dentro del núcleo, y del ARNs del núcleo dentro del citosol.
Dentro del núcleo hay uno o mas cuerpos esféricos llamados nucleolos. Estos son cúmulos de proteínas, ADN y ARN que no están envueltos por una membrana; constituyen sitios de ensamblaje de ribosomas, que desempeñan una función clave en la síntesis de las proteínas. Los nucleos son muy prominentes en las células que sintetizan enormes cantidades de proteínas, como las del hígado y las musculares. Durante la división celular los nucleolos se dispersan, desaparecen y reorganizan una vez que se han formado las células nuevas.
Dentro del núcleo se encuentran la mayoría de las unida-des hereditarias de las células, que reciben el nombre de genes (gen en singular), que controlan las estructuras celulares y dirigen la mayoría de las actividades celulares. Los genes están dispuestos en una sola fila a lo largo de los cromosomas. Las células somáticas humanas tienen 46 cromosomas, 23 heredados de la madre y 23 del padre. Cada cromosoma es una molécula larga de ADN que se halla estrechamente arrollada con varias proteínas (fig. 0.3.26). En una célula no dividida, los 46 cromosomas tienen la apariencia de una masa granular difusa, que recibe el nombre de cromatina. La micrografía electrónica muestra que la cromatina tiene una estructura como de cuentas ensartadas en un fillamento. Cada cuenta o esfera corresponde a un nucleosoma que consta de una doble hélice (o espiral) en torno a un centro de ocho proteínas llamado histona que ayuda a la organización helicoidal del ADN. El "filamento" entre las "cuentas" se denomina vinculo de ADN, cuya función consiste en mantener juntos los nucleosomas adyacentes. Otras histonas promueven el arrollamiento de los nucleosomas dentro del diámetro mas grande formado por las fibras de cromatina, que entonces se pliegan para formar grandes rizos. El ADN se organiza de esta manera en las células que no se hallan divididas. Sin embargo, justo antes de que se lleve a cabo la partición celular, el ADN se replica (duplica) y las espirales se condensan aun mas para formar pares de cromátidas hermanas, que se pueden apreciar fácilmente en el microscopio óptico como estructuras con forma de bastón.
SÍNTESIS DE PROTEÍNAS
OBJETIVO
• Describir la secuencia de los procesos que conlleva la síntesis de las proteínas.
Si bien las células sintetizan muchas sustancias químicas para mantener la homeostasis, muchos de los componentes celulares tienen como función sintetizar grandes cantidades de diversas proteínas, las cuales, a su vez, determinan las características físicas y químicas de las células y, por consiguiente, de los organismos. Algunas proteínas se utilizan para unir o ensamblar estructuras celulares como la membrana plasmática, el cito esqueleto y otros organelos. Otras proteínas sirven como hormonas, anticuerpos y elementos contráctiles del tejido muscular. Mas aun, otras son enzimas que regulan el número de reacciones químicas que se llevan a cabo en las células.
Las instrucciones para procesar las proteínas se encuentran principalmente en el ADN dentro del núcleo. Para sintetizar las proteínas, primero se transcribe (copia) la información codificada en una región del ADN, con el fin de producir una molécula especifica de ARN (fig. 0.3.27). Después la información contenida en este ácido se traduce en la secuencia correspondiente de aminoácidos que forman una molécula de proteína.
El ADN tiene la capacidad de codificar la producción de miles de diferentes proteínas. Como se vio en el capitulo 2, solo 20 aminoácidos están formados por bloques de todas las proteínas. Para que estas sean sintetizadas por las células, los ribosomas deben conjuntar los aminoácidos apropiados en una secuencia determinada por un segmento del ARN, el cual se produce de acuerdo con las instrucciones codificadas en la parte correspondiente del ácido desoxirribonucleico.
La información se almacena en el ADN (y también en el ARN) en conjuntos de tres bases nitrogenadas o nucleótidos. Una secuencia de tres nucleótidos en ese ácido recibe el nombre de triplete de bases. Cada uno de estos es trascrito (copiado) como una secuencia complementaria de tres nucleótidos de ARN, denominados codones. Un codon dado especifica un aminoácido. El código genético constituye un conjunto de reglas que se relacionan con la secuencia del triplete de bases del ADN con respecto a los cordones correspondientes del ARN y los aminoácidos que especifican.
El primer paso en la síntesis de las proteínas es la trascripción, que se analiza en el siguiente párrafo.
Trascripción
Durante este proceso, que ocurre dentro del núcleo, la información genética representada por la secuencia de tripletes de bases en el ADN sirve como molde o plantilla para copiar los datos en una secuencia complementaria de codones en un filamento de ARN. Las tres clases de ARN se originan a partir del molde de ADN: ARN mensajero (ARNm), el cual dirige las síntesis de una proteína; el ARN ribosómico (ARNr) que se una a las proteínas ribosómicas para producir ribosomas; y el ARN de transferencia (ARNt), que se enlaza a un aminoácido y lo mantiene en su lugar sobre un ribosoma hasta que es incorporado a una proteína durante el traslado. Existen mas de 20 distintas clases de ARNt, pero cada una se une solo a uno de los 20 diferentes tipos de aminoácidos. Por lo tanto, cada gen en un segmento de ADN es trascrito en un ARNm, ARNr, o ARNt especifico.
Una enzima ARN polimerasa cataliza la transcripción del ADN. No obstante, es necesario que se den instrucciones a la enzima sobre el sitio donde debe iniciar este proceso y el lugar donde ha de terminarlo. El segmento de ADN en el cual se inicia la transcripción es una secuencia nucleótida especial denominada promotora, que se localiza cerca del origen de un gen (fig. 3.28). Es aquí donde el ARN polimerasa se une al ADN. Durante la transcripción, las bases se complementan y forman pares (pareamiento). Las moldes o plantillas de bases de citosina (C), guanina (G) y timina (T) del ADN determinan como ha de ser la secuencia de estas bases en la cadena de ARN (fig. 3.28b). Sin embargo la plantilla de adenina en el ADN impone el uracilo (U) en vez de la timina en el ARN:
A U
T A
G C
C G
A U
T A
Molde de secuencia Secuencia de bases en el ADN
complementarias en el ARN
Solo uno de los dos filamentos de ADN sirve como plantilla para la síntesis de ARN, el cual es conocido como filamento con sentido. El otro (que no es trascrito) recibe el nombre de filamento en contrasentido.
La transcripción del filamento con sentido del ADN termina en otra secuencia nucleotida especial denominada de terminación, la cual especifica el fin del gen (fig. 03_08a). Cuando el ARN polimerasa alcanza este punto, la enzima se aparta de la molécula ARN transcrita y del filamento de ADN con sentido.
Dentro de un gen hay regiones llamadas intrones que no codifican ciertas partes de las proteínas, localizadas entre regiones denominadas extrones que codifican segmentos de una proteína. Inmediatamente después de la trascripción el ARNm incluye información de intrones y exones, por lo que recibe el nombre de ARN pre-m. Pequeñas ribonucleoproteínas (enzimas que recortan los intrones y los unen a los exones) del núcleo separan los intrones del ARNpre-m (fig. 0.3.28b). El resultado es una molécula ARN m funcional que pasa a través del poro de la membrana del núcleo hasta llegar al citoplasma, donde se realiza el transporte.
Traducción
Recibe el nombre de traducción el proceso mediante el cual la secuencia nucleotida en una molécula de ARNm determina el orden de los aminoácidos de una proteína. Los ribosomas del citoplasma llevan a cabo este proceso. La subunidad pequeña de un ribosoma tiene un sitio de enlace para el ARN; la subunidad grande posee dos de estos sitios (fig.0. 3.29). El primer lugar de unión de las moléculas de ARNt es el sitio P, donde la primera molécula ARNt, que lleva su aminoacido especifico, se une al ARNm. El segundo es el sitio A, que sostiene la siguiente molécula ARNt que lleva sus aminoácidos. El transporte se realiza en la siguiente secuencia (fig.0. 3.30):
1 Una molécula de ARNm se enlaza a una pequeña subunidad ribosómica en su sitio de unión. Un ARN especial, denominado iniciador, se une al codon de inicio (AUG) en el ARN donde empieza el transporte.
2 La unidad ribosómica grande se adhiere a la subunidad pequeña, con lo que da origen a un ribosoma funcional. El ARN iniciador se acopla al sitio P del ribosoma. Un extremo de un ARN tiene un aminoácido especifico y el extremo opuesto consta de un triplete de nucleótidos llamado anticodon. Por medio del apareamiento de bases, el anticodon ARN se une al codon ARN complementario. Por ejemplo, si el codon ARNm esta en el codon de inicio, entonces el ARNt con el anti codon UAC se unirá a el.
3 El anticodon de otro ARNt con su aminoacido se une al codon ARNm complementario en el sitio A del ribosoma.
4 El polipéptido en crecimiento se separa de ARNt en el sitio P y forma un enlace peptídico con el aminoácido transportado por el ARNt en el sitio A. Una enzima en la subunidad ribosómica grande cataliza la formación de un enlace peptídico.
5 Luego de la formación del enlace peptídico, el ARNt del sitio P se separa del ribosoma, y este cambia el filamento ARNm por un codon. El ARNt en el sitio A, que lleva la proteína recién formada, se desplaza al sitio P y permite que otro ARNt con sus aminoácidos se enlace a un codon recientemente expuesto en el sitio A. Se repiten de nuevo los pasos cuatro y cinco una y otra vez con-forme la proteína se alarga de manera progresiva.
6 La síntesis de las proteínas se detiene cuando el ribosoma alcanza un codon de alto (stop) en el sitio A, en el momento en que la proteína completa se separa del ARN final. Entonces el ARN deja vacante el sitio A y el ribosoma se divide en sus unidades constitutivas.
La síntesis de proteínas se lleva a cabo a una velocidad de 15 aminoácidos por segundo. Conforme el ribosoma se mueve a lo largo del ARN y antes de que termine la síntesis de la proteína completa, otro ribosoma puede unirse a la parte posterior de esta e iniciar la traducción del mismo filamento de ARN. De esta manera varios ribosomas forman un polirribosoma que puede estar unido a este mismo ácido mensajero. El movimiento simultaneo de varios ribosomas a lo largo de la misma molécula de ARNm permite que se realice su traducción en varias proteínas idénticas en un lapso breve.
APLICACIÓN CLÍNICA ADN recombinante
A partir de 1973, los científicos idearon técnicas para insertar genes de otros organismos dentro de diversas células huéspedes. Esta manipulación hace que el organismo huésped produzca proteínas que normalmente no sintetizaría. Los organismos alterados de esta manera son denominados recombinantes y su ADN una combinación de ADN de diferentes fuentes recibe el nombre de ADN recombinante. Cuando este funciona en forma adecuada, el huésped sintetiza la proteína determinada por el gen que se le ha insertado. La nueva tecnología que ha surgido de la manipulación de los materiales genéticos se denomina ingeniería genética.
Las aplicaciones practicas de la tecnología del ADN recombinante son enormes. En la actualidad se producen cepas de bacterias recombinantes que generan grandes cantidades de sustancias terapéuticas (para tratamiento). Entre ellas esta la hormona del crecimiento humano (hHG), la cual se requiere para el desarrollo durante la nifiez y es importante para el metabolismo de los adultos; la insulina, una hormona que ayuda a regular los niveles de glucosa en la sangre, se usa para controlar la diabetes; el interferon (IFN), una sustancia antiviral (y posiblemente anticancerigena), y la eritropoyetina, una hormona que estimula la producción de eritrocitos.
DIVISIÓN CELULAR NORMAL
OBJETIVO
Describir las etapas, los procesos y el significado de la división de las células somáticas.
La mayoría de las actividades celulares mantienen la vida de estas unidades día a día. Sin embargo, cuando las células somáticas sufren daño, enferman o se desechan son reemplazadas por medio de un proceso denominado división celular, mediante el cual se reproducen por si mismas. Hay dos tipos de división: de células somáticas y de células reproductivas.
En la división celular somática, una de estas unidades entra en un proceso llamado mitosis y de separación citoplasmática denominada citocinesis para producir dos células hijas idénticas. Cada una tiene el mismo numero y clase de cromosomas que la progenitora. La división de unidades somáticas permite reemplazar las células muertas o lesionadas y añade nuevas para el crecimiento tisular.
La división de las células reproductivas es el mecanismo que produce gametos espermas y óvulos que las células necesitan para formar la nueva generación de organismos sexualmente reproductivos. Este proceso consiste en una división especial, que consta de dos etapas, llamada meiosis, conforme a la cual el numero de cromosomas en el micleo se reduce a la mitad. La meiosis se describe en el capitulo 28; en esta parte nos enfocamos a la división de las células somáticas.
El ciclo de las células somáticas
El ciclo celular es una secuencia ordenada de sucesos por medio de los cuales una célula duplica su contenido y se divide en dos. Las células humanas, con excepción de los gametos, contienen 23 pares de cromosomas. Los dos cromosomas correspondientes a cada par —uno aportado por la madre y otro por el parte reciben el nombre de cromosomas homólogos. Ambos tienen genes similares, que por lo regular se hallan dispuestos en el mismo orden. Cuando una célula se reproduce debe replicar (duplicar) todos sus cromosomas para que sus genes puedan ser pasados a la siguiente generación de células. Este ciclo consta de dos periodos principales: interfase cuando una célula no esta en división y la mitotica cuando lleva a cabo este proceso (fig. 0.3.31).
Interfase
Durante la interfase la célula replica su ADN; también manufactura organelos adicionales y componentes citosolicos como paso previo a la división celular. La interfase constituye un periodo de gran actividad metabólica, durante el cual se efectúa la mayor parte del crecimiento celular.
La interfase a su vez consta de tres fases: G\, S, y G2 (fig. 0.3.31) . El filamento S participa en la síntesis del ADN. Dado que las fases G constituyen periodos en los que no hay actividad relacionada con la duplicación del ADN se consideraa que constituyen un intervalo de interrupción. La fase Gi e; el intervalo entre las fases mitotica y S. Durante la Gj la célula tiene actividad metabólica, duplica sus organelos y los componentes citosolicos con excepción del ADN. La replicación de los centrosomas comienza durante esta etapa pero no concluye en ella; virtualmente todas las actividades celulares descritas en este capitulo se llevan a cabo durante la Gi. En un cuerpo celular típico, cuyo ciclo tiene una duración de 24 horas, Gi puede durar aproximadamente de ocho a 10 horas. Sin embargo, este lapso es muy variable y puede comprender desde minutos, horas o anos segun los diferentes tipos de células. La fase S constituye el intervalo entre Gi y G2 y dura unas seis a ocho horas. Este lapso también resulta muy variable para los diferentes tipos de células. Durante la fase S se realiza la replicación del ADN, con lo cual se asegura que el material genético de las dos células hijas, formadas a partir de la división celular, será idéntico. La fase G2 es el intervalo entre la S y la mitotica; dura unas cuatro a seis horas. Durante ella prosigue el crecimiento de la célula, son sintetiza-das otras proteínas y enzimas para preparar la división, y se completa con la replicación de los centrosomas. Las células que permanecen en Gj, por mucho tiempo, quizá estén destinadas a no dividirse nunca mas, por ello se dice que se hallan en estado Go Por ejemplo, la mayoría de las células nerviosas se hallan en esta fase. No obstante, una vez que las células inician la fase S se ven forzadas a entrar en división.
Cuando se efectúa la replicación del ADN durante la fase S, su estructura helicoidal se desenrolla parcialmente y los dos filamentos se apartan en los puntos donde los enlaces de hidrogeno conectan con los pares de bases (fig. 0.3.32). Luego cada base expuesta es tomada por otra complementaria (con su azúcar y grupo fosfato asociado). Este desenrollamiento y la formación de pares de bases complementarios continua hasta que los dos filamentos originales del ADN se unen con el re-cien formado filamento de ácido desoxirribonucleico complementario. La molécula original de ADN tiene ahora dos moléculas idénticas de este mismo ácido.
Si se observa al microscopio la célula durante la interfase, es posible ver con claridad la envoltura nuclear, los núcleos y la cromatina (fig. 0.3.33). La ausencia de cromosomas visibles es otra de las características de la interfase. Una vez que la célula termina sus actividades durante las fases de interfase Gi, S y G2, da inicio la mitotica.
Fase mitotica
La fase mitotica (M) del ciclo celular consiste en la división nuclear o mitosis y la citoplásmica o citocinesis. Los sucesos que ocurren durante ella son plenamente visibles al microscopio porque la cromatina se condensa dentro de los cromosomas.
División nuclear: mitosis. Consiste en la distribución de los dos conjuntos de cromosomas, uno en cada uno de los núcleos separados. El resultado del proceso es la división exacta de la información genética. Por conveniencia los biólogos dividen el proceso en cuatro etapas: profase, metafase, anafase y telofase. Sin embargo la mitosis comprende un proceso continuo y las etapas se mezclan una con la siguiente de manera imperceptible.
• Profase. Durante la profase temprana, las fibras de cromatina se condensan y acortan (fig.0. 3.33b). La condensación puede evitar el proceso de arrollamiento de los filamentos largos de ADN, cuando estos se mueven durante la mitosis. Dado que la replicación del ADN ocurre durante la fase S de la interfase, cada cromosoma en profase contiene un par de filamentos cromátidas de doble hélice. Cada par se mantiene junto por una región constreñida denominada centrómero que se requiere para la adecuada segregación de los cromosomas. Unida al exterior de cada centro hay una proteína compleja conocida como cinetocoro, cuya función se resume en los siguientes párrafos.
En una etapa mas avanzada de la profase, el nucleolo desaparece (vease la fig.0. 3.33b) y la envoltura nuclear se rompe. Además, cada centrosoma se desplaza hacia uno de los polos opuestos (o terminales) de la célula y, al hacerlo, su área pericentriolar empieza a formar el huso mitótico, una estructura de microtubulos que se extienden en forma ovoidal. Al elongarse, los microtubulos empujan a los centrosomas hacia los polos celulares, de modo que el huso se extiende entre estos últimos y, al continuar su desarrollo, se forman tres tipos de microtubulos: 1. microtubulos ajenos a los cinetocoros que se extienden entre los dos centrosomas pero no fijan los cinetocoros; 2. microtubulos de los cinetocoros que se extienden por dentro y se unen a estos, y 3. microtubulos del aster que irradian hacia afuera desde el huso mitótico. Este ultimo es el que separa las cromátidas y conduce a cada una hacia un polo distinto.
• Metafase. En esta etapa, los microtubulos de los cinetocoros alinean los entromeros de los pares de cromatidesen el centra exacto del huso mitotico (fig.0. 3.33c). Esta región media se denomina piano ecuatorial o piano de la metafase.
• Anafase. Durante ella, los centrómeros se dividen y forman cromátidas hermanas. Esto permite que cada una, que forman un par, se dirija hacia el polo celular opuestoal que se desplaza su compañera (fig.03.33d). Cuando y están separadas, se les llama cromosomas hijos. A medida que estos se desplazan durante la anafase, impulsados porlos microtubulos de los cinetocoros, adquieren forma de V, mientras los centromeros gufan su trayecto y parece que arrastran las partes de los cromosomas hacia el polo.
• Telofase. Es la ultima etapa de la mitosis, que empieza alconcluir el desplazamiento de los cromosomas hijos (fig. 0.3.33e). Cada polo celular tiene un conjunto identico de cromosomas, los cuales se desenrollan y vuelven a formar cromatina filiforme. Entonces surge una envoltura en torno a cada masa de cromatina, aparece un nucleolo en las celulas hijas y fmalmente se rompe el huso mitotico.
División citoplasmica: citocinesis. Se da el nombre de citocinesis a la división del citoplasma y a la de los organelos de la celula madre. Este proceso principia en la anafase tardia o la telofase temprana, con la formación de un surco división, una ligera indentación en la membrana plasmática que por lo regular se forma en el piano medio entre los dos centrosomas y en torno al centro de la célula, junto a la cara interna de la membrana plasmática hay microfilamentos de actina, cuya acción da origen a dicho surco. Estos filamentos constituyen un anillo contractil que tira cada vez mas de la membrana, con lo que primero constriñe la célula por la mitad, a manera de cinturón, y finalmente la divide en dos. El piano del surco mitótico siempre es perpendicular al del huso, con lo cual queda asegurarce que ambos conjuntos de cromosomas siempre queden repartidos de manera equitativa entre las células hijas. Al terminar la citocinesis principia la interfase (fig. 0.3.33f)
La secuencia de fenómenos en el ciclo celular total es h siguiente:
Fase G1 —> fase S —> fase G2, —> mitosis —» citocinesis
En el cuadro 0.3.3 se presenta un resumen de los eventos del ciclo de las células somáticas.
Control del destino de las células
OBJETIVO
• Estudiar las señales que inducen la división celular y explicar la apoptosis.
Las células tienen tres destinos posibles: proseguir su y funcionar sin dividirse, crecer y dividirse o morir. Se llama homeostasis al equilibrio entre la proliferación y la muerte celulares. Durante los últimos 10 afios se han realizado intensas y fructíferas investigaciones acerca de las señales que indican a la célula si debe mantenerse en la fase Gj, en que momento tiene que dividirse y cuando ha de morir. Se sabe que la señal clave que induce la división celular es una sustancia denominada factor promoter de maduración (FPM). Uno de los componentes de dicho factor es un grupo de enzimas conocidas como proteínas cdc2, así llamadas porque participan en el ciclo de división celular (cdc). Otro componente del FPM es la clinica, proteína que recibe ese nombre en virtud de que su concentración aumenta y disminuye con el ciclo celular. Durante la interfase se incrementa el nivel de esta ultima, que activa las proteínas cdc y, por tanto, el FPM, con lo que se inicia la mitosis.
La muerte celular también esta regulada. Durante todos la vida de un organismo, ciertas células llegan de manera ordenada a una muerte programada genéticamente; este proceso se llama apoptosis. En este caso, un agente activador intracelular o extracelular, ocasiona que los genes de "suicidio celular" produzcan enzimas que lesionan la célula de diversas maneras, incluso con ruptura del cito esqueleto y el micleo. Como resultado, la célula se encoge y separa de sus vecinas; el ADN del núcleo se fragmenta y el citoplasma se contrae, pero la membrana plasmática permanece intacta, Entonces los fagocitos que se hallan en los alrededores la in-giren para destruirla. La apoptosis es un tipo de muerte celular normal y se diferencia de la necrosis, que es un tipo de muerte patológica debida a lesión de los tejidos. Con este tipo de muerte, muchas células adyacentes se hinchan, explotan y derraman su citoplasma hacia el liquido intersticial; por lo regular los detritos celulares estimulan una reacción inflamatoria del sistema inmunitario, lo que no ocurre con la apoptosis. Esta ultima resulta especialmente útil pues permite eliminar células innecesarias durante el desarrollo prenatal, pero prosigue después del nacimiento para regular el numero de células en los tejidos y eliminar las que resulten potencialmente peligrosas, como las cancerosas.
APLICAClÓN CLÍNICA
Genes supresores de tumores
Las anormalidades en los genes que regulan el ciclo celular o la apoptosis ocasionan muchas enfermedades. Por ejemplo, la causa de algunos canceres es una lesión de los llamados genes supresores de tumores, los cuales producen proteínas que normalmente inhiben la división celular. La perdida o alteración del gen supresor de tumores p53 en el cromo-soma 17 es el cambio genético causal de muy diversos tumores, corno los canceres de mama y colon. La proteína p53 normal secuestra células en la fase G1, con lo que impide su división; además es necesaria para iniciar la apoptosis.
CÉLULAS Y ENVEJECIMIENTO
OBJETIVO
• Explicar la relación entre los mecanismos celulares y los de envejecimiento.
El envejecimiento es un proceso normal que conlleva cambios progresivos en las reacciones homeostáticas corporales de adaptación. Ocasiona cambios observables en las estructuras y funciones, además de aumentar la vulnerabilidad a la tensión ambiental y las enfermedades. La geriatría es la rama de la medicina especializada en la atención de problemas y el cuidado de personas de edad avanzada.
Normalmente, cada minuto se producen millones de células en el cuerpo, pera varias clases de ellas (cardiacas, de músculo esquelético y neuronas) no se dividen porque siempre están secuestradas en la fase G0. En experimentos se ha demostrado que muchos otros tipos de células solo tienen capacidad limitada para dividirse; cuando se cultivan fuera del cuerpo, clnicamente se dividen cierto numero de veces y luego dejan de hacerlo. Estas observaciones son indicativas de que la interrupción de la mitosis es un fenómeno normal y programado genéticamente. De acuerdo con este punto de vista, los "genes de envejecimiento" constituyen una parte de la plantula genética congénita y se activan en momentos predeterminados para desacelerar o detener los procesos vitales .La glucosa es el azúcar mas abundante en el cuerpo y participa en el proceso de envejecimiento. Este carbohidratos se agrega de manera fortuita a proteínas intracelulares y extracelulares, con lo que forma enlaces cruzados irreversibles entre las moléculas proteinicas adyacentes. Al aumentar la edad se forman cada vez mas enlaces cruzados, lo que con-tribuye a la anquilosis y la perdida de elasticidad que sufren los tejidos con el envejecimiento.
Los radicales libres ocasionan lesiones oxidativas en lípidos, proteínas y ácidos nucleicos al "hurtar" un electrón para que acompañe a sus electrones no pareados. Algunos efectos son arrugamiento de la piel, rigidez de articulaciones y endurecimiento de las arterias. El metabolismo celular normal (p. ej., la respiración celular aeróbica en las mitocondrias) genera algunos radicales libres; otros se hallan en los contaminantes del aire, radiaciones y ciertos alimentos. Normalmente, las enzimas que se encuentran de manera natural en los peroxisomas y el citosol eliminan los radicales libres; en tanto que determinadas sustancias de la dieta, como las vitaminas E y C, el beta-caroteno y el selenio, son antioxidantes que inhiben la formación de radicales libres.
Algunas teorías del envejecimiento explican el proceso en el nivel celular, mientras que otras se concentran en los mecanismos reguladores que operan en todo el organismo. Por ejemplo, el sistema inmunitario puede iniciar ataques contra las propias células corporales. Es posible que esta respuesta auto inmunitaria se deba a cambios en los marcadores de identidad celular que se ubican en la superficie de las células, lo cual ocasiona que los anticuerpos las ataquen y marquen para destrucción. Al acumularse los cambios en las proteínas de la membrana plasmática celular, la respuesta auto inmunitaria se intensifica y da origen a los bien conocidos signos del envejecimiento.
DIVERSIDAD CELULAR
Hasta este punto, hemos investigado la complementaridad intracelular de la función y la estructura de las células. Casi todas ellas poseen un conjunto de organelos, cada uno de los cuales controla una parte especifica de los procesos fisiológicos que se realizan dentro de estas unidades vitales. Sin embargo, no todas las células corporales tienen el mismo aspecto ni realizan funciones idénticas en el organismo.
En promedio, un adulto humano tiene casi 100 billones de células, que se pueden clasificar en alrededor de 200 tipos distintos. Su tamaño varia considerablemente; así, para ver las mas pequeñas se requieren microscopios de alta potencia, mientras que el ovulo (la mas grande de todas) se puede observar a simple vista. El tamaño de las células se mide en micras, también llamadas micrómetros (um). (Una micra es igual a la millonésima parte de un metro, o sea 10" m.) El diámetro de un glóbulo rojo o eritrocito es de 8 mm, en tanto que el de un óvulo es de aproximadamente 140 micras.
Las células también tienen formas muy variables (fig. 0.3.34), pues las hay redondas, ovaladas, planas, cuboidales, columnares, alargadas, estrelladas, cilíndricas y discoides.
La forma de cada una se relaciona con las funciones que lleva a cabo en el organismo. Por ejemplo, el espermatozoide tiene una cola larga como látigo (flagelo) que utiliza para su locomoción. La forma discoide de los eritrocitos les confiere gran área superficial, lo que aumenta su capacidad para llevar oxigeno a otras células. Algunos otros tipos tienen microvellos que incrementan en gran medida su área superficial; tales micro vellosidades son comunes en las células epiteliales que recubren por dentro el intestino delgado, donde aumentan la capacidad de absorción de alimentos digeridos.
Las células nerviosas poseen largas prolongaciones que les permiten transmitir impulsos nerviosos a grandes distancias.
La diversidad celular constituye la piedra angular que hace posibles niveles mas complejos de organización biológica, como los correspondientes a tejidos y órganos.
CÁNCER
Se da este nombre a un grupo de enfermedades que se caracterizan por la proliferación. sin control de las células. Cuando estas se desarrollan sin orden en alguna parte del cuerpo se produce un exceso de tejido al que se conoce como tumor o neoplasia.
El estudio de los tumores se llama oncóloga. Las masas tumorales pueden ser can-cerosas, a veces letales o inocuas. Si la neoplasia es cancerosa se dice que se trata de un tumor maligno o enfermedad maligna. Una propiedad de estas neoformaciones es su capacidad para generar metástasis, es decir para diseminar las células enfermas a otros lugares del organismo. Los tumores benignos no son crecimientos cancerosos y, aunque no envían metástasis, en ocasiones resulta necesario extirparlos, cuando obstaculizan la función corporal normal o provocan alguna deformación a la persona.
Tipos de cáncer
Tejido donde se desarrolla. La mayoría de las neoplasias humanas son carcinomas, formaciones malignas que se desarrollan a partir de células epiteliales. Por ejemplo, los melanomas son crecimientos cancerosos de melanocitos, células epiteliales que producen el pigmento melanina. Sarcoma es un termino general que se utiliza para aludir a cualquier cáncer que se forma a partir de células musculares o en tejidos conectivos. Por ejemplo, el sarcoma osteogeno, que constituye el tipo mas frecuente de cáncer en los niños, destruye el tejido óseo normal. La leucemia es un cáncer de los órganos que generan la sangre, el cual se caracteriza por desarrollo rápido de leucocitos (glóbulos blancos) anormales, en tanto que los linfomas constituyen una enfermedad maligna de tejidos linfáticos, como los ganglios de este sistema.
Desarrollo y diseminación del cáncer
Las células de los tumores malignos se duplican con rapidez y de manera continua y cuando invaden los tejidos circundantes inician la angiogenesis, que es el desarrollo de nuevas redes de vasos sanguíneos. Al grupo de proteínas que inicia este proceso en la neoplasia se le conoce como factor de angiogenesis totonoral (FAT). Al crecer, el cáncer empieza a competir con los tejidos sanos por espacio y nutrientes. Finalmente, el tejido normal reduce su tamaño y muere. Algunas células malignas pueden desprenderse de la neoplasia inicial (primaria) e invadir alguna cavidad corporal o entrar al torrente sanguíneo o la linfa, donde circularan hasta invadir otros tejidos y formar tumores secundarios. Las células malignas son resistentes a las defensas antitumorales del organismo. El dolor que provoca el cáncer surge cuando la neoformación presiona nervios o bloquea un conducto de salida en algún órgano, con lo que aumenta la presion de las secreciones.
Causas de cáncer
Muy diversos factores pueden provocar la perdida de los mecanismos de control en las células normales y convertirlas en cancerosas. Varios agentes ambientales constituyen una causa posible: partículas que se hallan en el aire, el agua y los alimentos que se consumen. Las sustancias químicas y las radiaciones que provocan cáncer reciben el nombre de carcinogenos. Estos agentes inducen mutaciones, que son cambios estructurales permanentes en la secuencia de bases del ADN genético .La Organización Mundial de la Salud estima que los carcinogenos causan de 60 a 90% de todos los canceres humanos. Algunos ejemplos de estas sustancias son ciertos hidrocarburos que contiene el alquitrán del tabaco, el gas radon que libera la Tierra y las radiaciones ultravioleta (UV) de la luz solar.
Algunos virus constituyen una segunda causa de cáncer. Estos agentes son pequeños paquetes de ácido nucleico, sea ADN o ARN, que pueden infectar las células y convertirlas en productoras de virus. El vinculo entre estos agentes y el cáncer esta bien establecido en diversas enfermedades malignas animales; sin embargo la relación con los canceres humanos es menos clara, pues el numero de personas infectadas por dichos virus es mucho mayor que el de quienes desarrollan enfermedad maligna. Las pruebas de que se dispone dan indicios de que las infecciones virales crónicas se relacionan hasta con 20% de todos los canceres.
En la actualidad se realizan intensos trabajos de investigación acerca de los genes causales de cáncer, conocidos como oncogenes. Cuando estos se activan de manera inadecuada, tienen la propiedad de convertir en cancerosas la células normales. Los oncogenes se desarrollan a partir de genes normales que regulan el crecimiento y el desarrollo, a los que se llama protocogenes. Estos últimos pueden sufrir alguna alteración que los hace producir sustancias anormales o perder su control, por lo que su expresión resulta inadecuada y ocasiona que generen productos en exceso o que lo hagan en momentos inapropiados. Los científicos creen que algunos oncogenes ocasionan que se formen los factores de crecimiento en exceso, los cuales son sustancias químicas que estimulan el desarrollo celular. Algunos otros provocan alteraciones en las superficies de los receptores, con lo que estos envían señales como si los hubiera activado un factor de crecimiento. En consecuencia, los patrones de desarrollo celular se vuelven anormales.
Los protoncogenes de todas las células realizan funciones celulares normales hasta que se produce el cambio maligno. Parece que algunos de ellos son activados como oncogenes por mutadones que alteran su ADN. La activación de otros depende de un reordenamiento de cromosomas en el que se intercambian algunos segmentos del ADN. Tales reacomodamientos activan a los protooncogenes al ocasionar que los genes que hacen aumentar su actividad queden junto a ellos. Se cree que algunos virus causan cáncer al insertar sus propios oncogenes o protoncogenes en el ADN de la célula huésped.
Carcinogenesis: un proceso de múltiples etapas La carcinogenesis es el proceso por el que se desarrolla el cáncer y se lleva a cabo en muchos pasos o etapas que comprenden hasta 10 mutaciones distintas, que deben acumularse en la célula antes que se vuelva cancerosa. Esta progresión de cambios genéticos que culminan con enfermedad maligna se ha estudiado mejor en el caso del cáncer de colon (colorrectal), un trastorno que tarda anos en desarrollarse, al igual que las neoplasias malignas de mama y pulmón. En el cáncer de colon el tumor principia como un área de proliferación celular incrementada a causa de una mutación. Dicho crecimiento prosigue y se vuelve anormal, aunque no canceroso, y forma tumores llamados adenomas. Luego de otras dos o tres mutaciones, se produce un cambio en p53 (un gen supresor de tumores) y se desarrolla el carcinoma. El hecho de que se requieran tantas mutaciones para que se forme un tumor maligno es indicativo de que el crecimiento celular normalmente depende de varios conjuntos de verificaciones y equilibrios.
Tratamiento del cáncer
Muchas neoplasias malignas se extirpan por cirugía. Sin embargo, se prefiere aplicar quimioterapia y radioterapia cuando el cáncer esta muy diseminado por el organismo o se halla en órganos como el cerebro donde la intervención quirúrgica puede afectar mucho su funcionamiento. La quimioterapia consiste en administrar fármacos tóxicos para las células cancerosas en tanto que la radioterapia las destruye, con lo que evita que se reproduzcan. Las células cancerosas se dividen con mucha rapidez, de modo que resultan mas vulnerables que las normales a los efectos destructibles de los fármacos y las radiaciones; no obstante estos tratamientos también pueden matar o anular las funciones de algunas células sanas.
La terapéutica contra el cáncer es complicada y difícil, ya que no se trata de una sola enfermedad y porque generalmente no todas las células de una población tumoral se comportan de la misma manera. Se cree que la mayoría de los canceres se derivan de una sola célula anormal, pero al momento en que el tumor alcanza Un tamaño clínicamente detectable, es posible que ya contenga una población de distintas células anormales. Por ejemplo, algunas células malignas envían metástasis en muy poco tiempo, pero otras no lo hacen; alguna; son sensibles a la quimioterapia, mientras otras son resistentes a lo: fármacos. Dada esta distinta capacidad de resistencia a los medicamentos, es posible que una sustancia destruya las células susceptibles pero permita que proliferen otras que la resistan.
Otro obstáculo que encuentran los medicamentos anticancerosos transportados por sangre es la barrera física que forman tumores sólidos como los que crecen en mamas, pulmones, colon y otros órganos. Las áreas profundas con alta presion en tales neoplasias ocasionan el colapso de los vasos sanguíneos tumorales, lo cual dificulta o hace imposible que el agente anticanceroso transportado en la sangre penetre en la neoformacion.
TERMINOLOGÍA MEDICA
Anaplasia. Perdida de la diferenciación y la función de los tejidos, que es característica de muchas enfermedades malignas.
Atrofia. Disminución del tamaño celular, con la subsecuente reducción en el volumen del tejido u órgano afectado; emaciación.
Displasia. Alteración del tamaño, forma y organización celular, debido a irritación o inflamación crónicas; puede progresar y convertirse en neoplasia (formación de tumor, generalmente maligno) o revertirse y recuperar la normalidad, cuando se elimina la irritación.
Hiperplasia. Aumento en el número de células de un tejido, a causa de un incremento en la frecuencia de división celular.
Hipertrofia. Aumento del tamaño de las células sin que haya división celular.
Metaplasia. Transformación de un tipo de célula en otro.
Progenia. Hijos o descendientes.
GUÍA DE ESTUDIO
INTRODUCCIÓN (p. 61)
1. La célula es la unidad básica con estructura y función vitales del organismo.
2. La citología es el estudio científico de la estructura celular. La fisiología celular estudia las funciones de la célula.
DESCRIPCIÓN GENERAL DE LA CÉLULA (p. 61)
1. En la figura 3.1 se observa un esquema general de la estructura celular, donde se combinan las características de muchas células del cuerpo.
2. Las partes principales de la célula son: membrana plasmática y citoplasma, que se compone de citosol, organelos y núcleo.
LA MEMBRANA PLASMATICA (p. 62)
1. La membrana plasmática envuelve y contiene al citoplasma dela célula.
2.La forman una mezcla de proteínas y lípidos en proporción de 50:50 por peso; estos componentes se mantienen unidos por fuerzas no covalentes.
3.Según el modelo de mosaico del liquido, la membrana plasmática constituye una especie de mosaico de proteínas que flotan en la bicapa de lípidos, como iceberg en el mar.
4.La bicapa lipidica se compone de dos membranas paralelas de fosfolípidos, colesterol y glucolípidos, con orientaciones opuestas. Esta configuración se debe a que los lípidos son anfipaticos, pues tienen una parte polar y otra no polar.
5.Algunas proteínas integras se extienden dentro de la bicapa de lípidos o a todo lo ancho de esta, en tanto que las proteínas superficiales se asocian con lípidos de la membrana o con proteínas
integras, ya sea en la superficie interna o en la externa de la bicapa.
6.Muchas de las proteínas integras son glucoproteínas que tienen grupos de azucares fijos en el extremo orientado hacia el líquido extracelular; estas moléculas, junto con las de los glucolípidos, forman el glucocaliz en la superficie externa de las células.
7. Las proteínas de membrana tienen muy diversas funciones. Así, constituyen canales y transportadores gracias a los cuales solutos
específicos pueden cruzar la membrana. También forman receptores, que sirven como sitios de identificación, y actúan como ligadores que fijan las proteínas en la membrana plasmática de células vecinas, enlazándolas una a otra, o a los filamentos que están dentro o fuera de la célula. Algunas proteínas de la membrana son enzimas y otras, marcadores de identidad celular.
8. La membrana tendrá mayor fluidez cuanto mas enlaces dobles tenga la cola de ácidos grasos de los lípidos que forman la bicapa. El colesterol brinda fortaleza a la bicapa lipidica, pero es
menos fluido a la temperatura normal del cuerpo. Debido a esta propiedad, la bicapa se sella cuando se le aguijonea o punciona.
9. La permeabilidad selectiva de la membrana permite que algunas sustancias la atraviesen con mayor facilidad que otras. La bicapa de lípidos es permeable al agua y a la mayoría de las moléculas no polares, sin carga, pero impermeable a iones y particulas cargadas o polares, excepto el agua. Los canales y transportadores aumentan la permeabilidad de la membrana a sustancias polares y sustancias con carga media o pequeña, incluso a iones que no pueden atravesar la bicapa. 10. La permeabilidad selectiva de la membrana soporta la formación de gradientes de concentración, que son diferencias entre las concentraciones de sustancias químicas en uno y otro lado de la membrana.
TRANSPORTE A TRAVÉS DE LA MEMBRANA PLASMÁTICA (p. 66)
1. En el transporte mediado, la fijación de sustancias a una proteína transportadora les ayuda a cruzar la membrana. Los monotransportadores desplazan una sola sustancia, en tanto que los transportadores acoplados pueden cruzar dos. Los cotransportadores llevan dos sustancias en la misma dirección, mientras que los contra transportadores las desplazan en sentidos opuestos.
2. El transporte pasivo se realiza con sustancias que se desplazan según su gradiente de concentración transmembrana, aprovechando su propia energía cinética para avanzar. En el transporte activo se requiere energía celular para impulsar el movimiento de sustancias "a contracorriente", es decir, contra
su gradiente de concentración.
3. En el transporte vesicular se forman pequeñas vesículas que primero se desprenden de la membrana plasmática y luego se funden con ella a fin de acarrear los materiales hacia el interior o hacia el exterior de la célula.
4. La difusión neta es el desplazamiento efectivo de moléculas iones de un área de mayor concentración a otra de menor nivel, hasta que se establece el equilibrio.
5. La velocidad de difusión a través de la membrana depende de la pendiente de la curva del gradiente de concentración, la temperatura, el tamaño o la masa de sustancia que se difunde, el área superficial disponible para la difusión y la distancia a la que debe diseminarse la sustancia.
6. Se llama osmosis al desplazamiento neto de agua a través de una membrana con permeabilidad selectiva, de un área donde la concentración acuosa es mas alta a otra en la que es más baja.
7. Cuando se encuentran en una solución isotónica, los eritrocitos conservan su forma normal; si están en una hipotónica sufren hemólisis y si es hipertónica ocurre su crenación.
8. Las moléculas hidrófobas no polares se difunden a través de la bicapa lipidica de la membrana plasmática; por ejemplo, las de oxigeno, dióxido de carbono, nitrógeno, esteroides, vitamina liposolubles (A, E, D y K), alcoholes de molécula pequeña y el amoniaco.
9. Los canales iónicos selectivos de K+, CT, Na+ y Ca"+ permiten la difusión a través de la membrana de estos iones pequeños que son demasiado lipofilos como para penetrar al interior no polar de la membrana.
10. La difusión facilitada consiste en que un soluto, como la glucosa, se fije a un transportador especifico en un lado de la membrana y sea liberado en el otro, debido a cambios estructurales que se producen en el transportador. Hay un numero restringido de esta base de transportadores, por lo que la difusión facilitada
tiene una capacidad máxima limitada de transporte y es saturable.
11. Algunas sustancias pueden cruzar la membrana "a contracorriente" —en contra de su gradiente de concentración— mediante transporte activo. Entre ellas se cuentan varios iones (como Na+, K+, H+, Ca +, I y Cl ), aminoácidos y mono-sacáridos.
12. Se emplean dos fuentes de energía para el transporte activo: la que se obtiene de la hidrólisis del ATP, que es la principal, y la energía almacenada en el gradiente de concentración Na o H , que es la fuente secundaria para este tipo de transporte.
13. La bomba de sodio, también conocida como Na /K ATPasa, es la de mayor prevalecía en el transporte activo.
14. Los mecanismos secundarios de transporte activo incluyen tanto cotransportadores como contratransportadores, que reciben energía del gradiente de concentración del Na+ o del H+.
15. El transporte vesicular incluye tanto a la endocitosis como a la exocitosis.
16. La endocitosis mediada por receptores consiste en la captación selectiva de moléculas o partículas grandes (ligandos) que se fijan a receptores específicos que se hallan en áreas de la membrana conocidas como poros recubiertos de clatrina.
17. Se llama fagocitosis a la ingestión de partículas sólidas. Este es un proceso importante que aprovechan algunos leucocitos para destruir bacterias que ingresan al cuerpo.
18. Se denomina pinocitosis a la ingestión de liquido extracelular. Este proceso consiste en la formación de una vesícula pinocitotica que envuelve al liquido que engullirá.
19. La exocitosis es la expulsión de productos de excreción o desecho al exterior de la célula mediante la fusión de las vesículas (que los contienen) con la membrana plasmática.
CITOPLASMA
1. Se llama citoplasma a todo el contenido de la célula que se halla entre el núcleo y la membrana plasmática.
2. Se compone de citosol y organelos.
Citosol
1. Es la porción liquida del citoplasma.
2. Contiene agua principalmente, además de iones, glucosa, aminoácidos, ácidos grasos, proteínas, lípidos, ATP y productos de desecho.
3. El citosol es el sitio donde se llevan a cabo muchas reacciones requeridas para la existencia de la célula. Organelos (p. 77)
1. Son estructuras especializadas, cada una con forma característica y funciones específicas.
2. Algunos, como el citoesqueleto, el centrosoma, los cilios, los flagelos y los ribosomas, no son membranosos (es decir, no contienen membranas).
3. Otros, como el retículo endoplasmico, complejo de Golgi, mitocondrias, lisosomas y los peroxisomas, si son membranosos (contienen una o mas membranas de fosfolipidos).
4. El retículo endoplasmico es una red de numerosos filamentos de proteínas que se extienden por todo el citoplasma.
5. El citoesqueleto esta formado por microfilamentos, filamentos intermedios y microtubulos.
6. El citoesqueleto constituye el marco estructural de la célula y es el encargado de realizar todos los movimientos celulares.
7. El centrosoma se compone de centríolos y área pericentriolar.
8. El área pericentriolar organiza los microtubulos en la célula que no se halla en proceso de división y el huso mitótico en la célula que se esta dividiendo.
9. Los centríolos participan en la formación y regeneración de cilios y flagelos.
10. Los cilios y los flagelos son estructuras móviles que se proyectan fuera de la superficie celular.
11. Los cilios desplazan líquidos a lo largo de la superficie celular.
12. Los flagelos impulsan a toda la célula.
13.Los ribosomas se componen de proteínas y ARN ribosómicos; constan de dos subunidades que se forman dentro del núcleo.
14. Los ribosomas libres no se adhieren a ninguna estructura citoplásmica; los que se hallan fijos a la membrana están anclados al retículo endoplasmico.
15. Los ribosomas son sitios donde se lleva a cabo la síntesis de proteínas.
16. El retículo endoplasmico (RE) consiste en una red de membranas que forman sacos aplanados y túbulos llamados cisternas; se extiende desde la membrana nuclear y abarca todas las zonas del citoplasma.
17. El RE rugoso esta cubierto de ribosomas y su función primaria es efectuar la síntesis de proteínas. También forma glucoproteinas y adhiere las proteínas a los fosfolípidos, a los que también sintetiza.
18. El RE liso carece de ribosomas. Sintetiza fosfolípidos, grasas y esteroides; libera la glucosa del hígado hacia el torrente sanguíneo; desactiva o destoxifica fármacos y otras sustancias potencialmente dañinas, y libera iones calcio que activan la contracciónen las células musculares.
19.El complejo de Golgi esta formado por sacos aplanados a los que se llama cisternas; estas tienen distintos tamaños, formas, contenido, actividad enzimática y numero de vesículas. Las cisternas de entrada están frente al RE rugoso y las de salida frente a la membrana plasmática; además hay cisternas intermedias entre las anteriores.
20. El complejo de Golgi recibe los productos sintetizados por el RE, luego los modifica, clasifica, empaca y transporta dentro de las vesículas hacia distintos lugares.
21. Algunas proteínas procesadas salen de la célula dentro de vesículas secretorias; otras se incorporan en la membrana plasmática y algunas mas entran a lisosomas.
22. Los lisosomas son vesículas envueltas en una membrana y contienen enzimas digestivas.
23. Algunos organelos tardíos, como endosomas, fagosomas y vesículas pinocitoticas descargan materiales en los lisosomas para su degradación.
24. Los lisosomas participan en la digestión de organelos desechados (autofagia), la de células huésped (autolisis) y la extracelular.
25. Los peroxisomas son similares a los lisosomas, pero mas pequeños.
26.Los peroxisomas oxidan varias sustancias orgánicas, como los aminoácidos, ácidos grasos, sustancias toxicas y, en el proceso, generan peroxido de hidrogeno.
27.El peroxido de hidrogeno que se produce en la oxidación se de-
grada por acción de una enzima peroxisomica llamada catalasa.
28. Las mitocondrias constan de una membrana lisa externa, otra interna con crestas y una cavidad Uena de liquido llamada matriz. Se les conoce como "generadores" celulares porque producen ATP.
NÚCLEO
1.Consta de una doble envoltura nuclear; poros nucleares que regulan el desplazamiento de sustancias entre el núcleo y el citoplasma; nucleolos, que producen ribosomas, y genes ordenados en cromosomas.
2.Los genes controlan la estructura y la mayoría de las funciones de la célula.
SÍNTESIS DE PROTEÍNAS
1.La mayor parte de los mecanismos celulares están dedicados a la síntesis de proteínas.
2.Las células producen proteínas mediante la transcripción y traducción de la información genética codificada en el ADN.
3.El código genético es el conjunto de reglas que relacionan la secuencia de los tripletes de bases del ADN con los codones correspondientes del ARN y los aminoácidos a los que codifican.
4. Para la transcripción, la información genética contenida en la secuencia de los tripletes del ADN sirve como molde o plantilla para que el ARN mensajero copie las instrucciones en una secuencia complementaria de sus codones.
5.La transcripción se inicia en una región del ADN conocida como promotora.
6.Las regiones del ADN que codifican la síntesis de las proteínas reciben el nombre de exones; las que no participan en esta función se denominan intrones.
7.El ARN prem recién sintetizado se modifica antes de salir del núcleo.
8.La tradución es el proceso por el que la secuencia de nucleotidos del ARN m especifica la secuencia de aminoácidos de una proteína.
9.En la traduccion, el ARNm se fija a un ribosoma, los aminoacidos específicos se fijan a ARNt, y los anticodones de este ultimo se unen a los codones del ARNm, de tal modo que ordenan los
aminoácidos específicos en la posición que les corresponde en
la proteína que se va a formar.
10. La traducción principia en el codon de inicio y concluye en el de terminación.
DIVISIÓN CELULAR NORMAL
1. La división celular es el proceso por el que las células se reproducen. Consiste en la división nuclear (mitosis o meiosis) y la del citoplasma (citocinesis).
2. La división celular somática es aquella por la que se incrementa el numero de células corporales e implica el tipo de partición nuclear llamado mitosis, así como la citocinesis.
3. La división celular que genera espermatozoides u óvulos es la reproductiva y se realiza con el tipo de división nuclear llamado meiosis y citocinesis. El ciclo de las células somáticas (p. 93)
1.El ciclo celular constituye una secuencia ordenada de fenomenos por los que la célula duplica sus contenidos y se divide en dos. Consta de una interfase y una fase mitotica.
2. Antes de la fase mitotica, las moléculas de ADN o cromosomas,
se replican a fin de que puedan transmitirse cromosomas identicos a la siguiente generación de células.
3.Cuando la célula esta entre dos periodos de división y ejecutados los procesos vitales, excepto la reproducción, se dice que esta en la interfase, la cual se subdivide en tres fases: Gi, S
y G2.
4.Durante la fase Gi, la célula duplica sus organelos y componentes citosolicos; en la fase S se realiza la replicación del ADN y en la G2, se lleva a cabo la síntesis de enzimas y otras proteínas y se completa la replicación centrosomica.
5. La mitosis es la replicación y distribución de dos conjuntos de cromosomas entre dos núcleos separados e iguales entre si; consta de las siguientes etapas: profase, metafase, anafase y telofase.
6. Por lo regular, la citocinesis principia en la anafase tardía y termina en la telofase.
7. En el piano de la metafase celular se forma un surco divisorio, el cual se desarrolla hacia el interior para formar dos porciones separadas de citoplasma.
Control del destino de las células
1. Ciertas células pueden permanecer vivas y funcionar sin entrar en división, crecer y dividirse o morir.
2. El factor promotor de maduración (FPM) induce la división celular (tanto la mitótica como la meiotica) y se compone de cidina, que se desarrolla durante la interfase, y de proteínas cdc2,
a las que desactiva la del ink.
3. La apoptosis es la muerte celular programada y constituye un tipo natural de mortalidad. Se inicia durante el desarrollo embrionario y contimia por toda la vida del organismo.
4. Ciertos genes regulan la división celular y la apoptosis. Las anormalidades en estos genes es causa de muy diversas enfermedades y trastornos.
CÉLULAS Y ENVEJECIMIENTO
1 El envejecimiento es un proceso normal que se acompaña de alteraciones en las respuestas corporales homeostáticas de adaptación.
2. Se han propuesto muchas teorías sobre el envejecimiento, como el cese de la división celular genéticamente programado, la formación creciente de radicales libres y la intensificación de la respuesta auto inmunitaria.
DIVERSIDAD CELULAR
1. El organismo esta formado por casi 200 tipos de células, que varían considerablemente de tamaño y forma.
2. El tamaño de las células se mide en micras (micrometres). Una micra es igual a 10 mm.
3. La forma de las células se relaciona con sus funciones.
CUESTIONARIO DE AUTO EVALUACIÓN
Falso o verdadero:
6. De manera característica, la cara de la superficie interna de la membrana celular tiene carga mas positiva y la superficie externa, mas negativa.
7. La bomba de sodio constituye un ejemplo de transportes activo primario.
Complete los siguientes enunciados:
1. Las tres partes principales de la célula son:
2. La es la muerte celular programada, mientras que la alude a la muerte celular provocada por lesiones en los tejidos.
3. La porción liquida del citoplasma es el .
4. Los sitios donde se forman los ribosomas son los .
5. Relacione correctamente las dos columnas:
_ (a) mitosis (1) división citoplásmica.
_(b) meiosis (2) división de células somáticas que
_ (c) profase produce células hijas iguales.
_(d) metafase (3) división de celúlas reproductoras re-
_(e) que anafase duce a la mitad el mimero de cromo-
_(f) telofase somas.
_(g) citocinesis (4) etapa de crecimiento y metabolismo
_(h) interfase celulares
(5) etapa en que las fibras de cromatina
se condensan y acortan para formar
cromosomas.
(6) etapa en que los centromeres se divi-
den y las cromatides hermanas se diri-
gen hacia polos celulares opuestos.
(7) etapa en que los centromeres de los
pares de cromatides se alinean en el
centro del huso mitótico.
(8) etapa en que los cromosomas se de-
senrollan y se revierten a cromatina
Escoja la respuesta adectiada para las siguientes preguntas:
8. La unidad estructural básica de la membrana plasmática es: a) la bicapa de lípidos, b) la proteína Integra, c) la molécula de colesterol, d) la proteína periférica, e) el complejo glucoproteína-glucolipido.
9. Las proteínas íntegras pueden actuar en la membrana celular de todas las formas siguientes, excepto: a) como canal, b) como transportador, c) como receptor, d) como vesícula de exocitosis, e) como marcador de identidad celular.
10. ¿Cual de los siguientes factores influye en la velocidad de difusión de sustancias a través de la membrana plasmática? 1) el gradiente de concentración; 2) la distancia de difusión; 3) el área superficial; 4) el tamaño de la sustancia que se difunde; 5) la temperatura.
(a) 1, 2, 3 y 5; (b) 1, 2, 3, 4 y 5; (c) 2, 3, 4 y 5; (d) 1, 2 y 5; (e) 2, 3y5.
11. La solución en la que la célula conserva su forma y volumen normales se llama: a) hipertónica, b) hipotónica, c) isotónica, d) de solutos, e) de solventes.
12. ¿Cual de las siguientes afirmaciones acerca del núcleo es verdadera? 1) Los nucleolos que están dentro del núcleo son los sitios donde se sintetizan los ribosomas; 2) La mayoría de las unidades hereditarias de la célula, llamadas genes, se localizan dentro del núcleo; 3) La membrana nuclear es sólida e impermeable; 4) La síntesis de proteínas se realiza dentro del núcleo; 5) Cuando las células están en interfase, el ADN se halla dentro
del núcleo en forma de cromatina.(a) l,2y3;(b) I,2y4; (c) 1, 2y 5; (d) 2, 4y 5; (e) 2, 3 y 4.
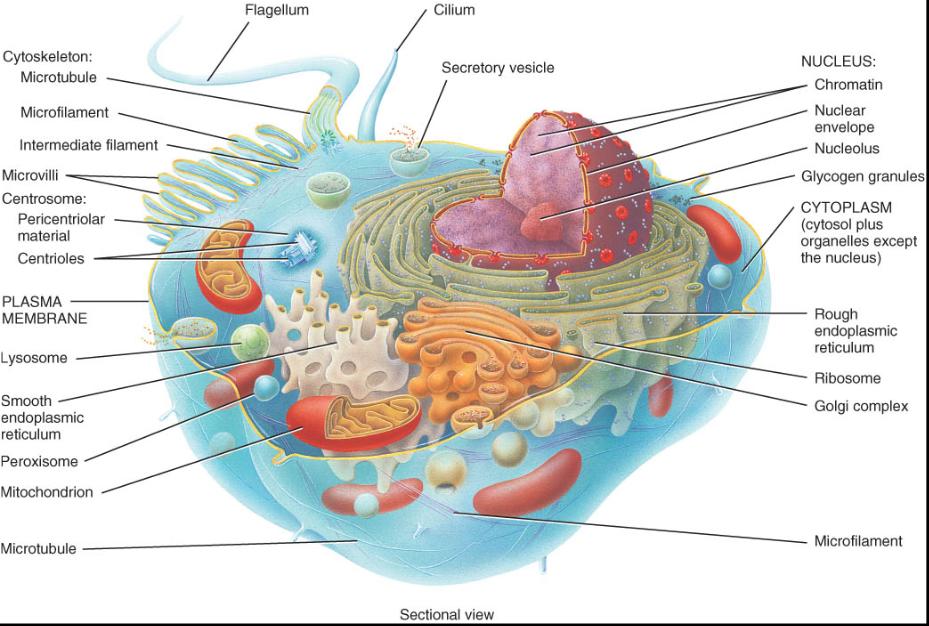{kind=link}
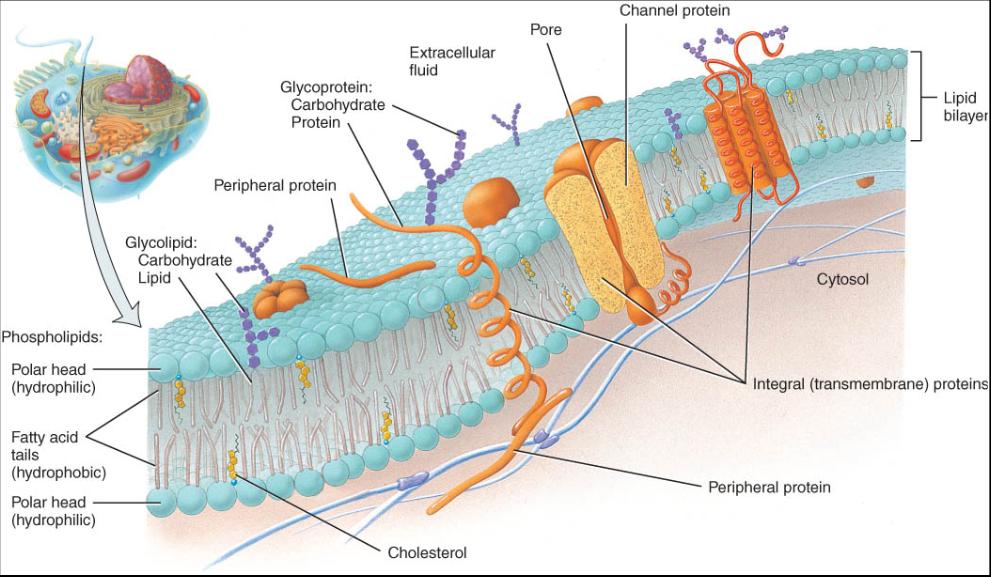{kind=link}
{kind=link}
{kind=link}
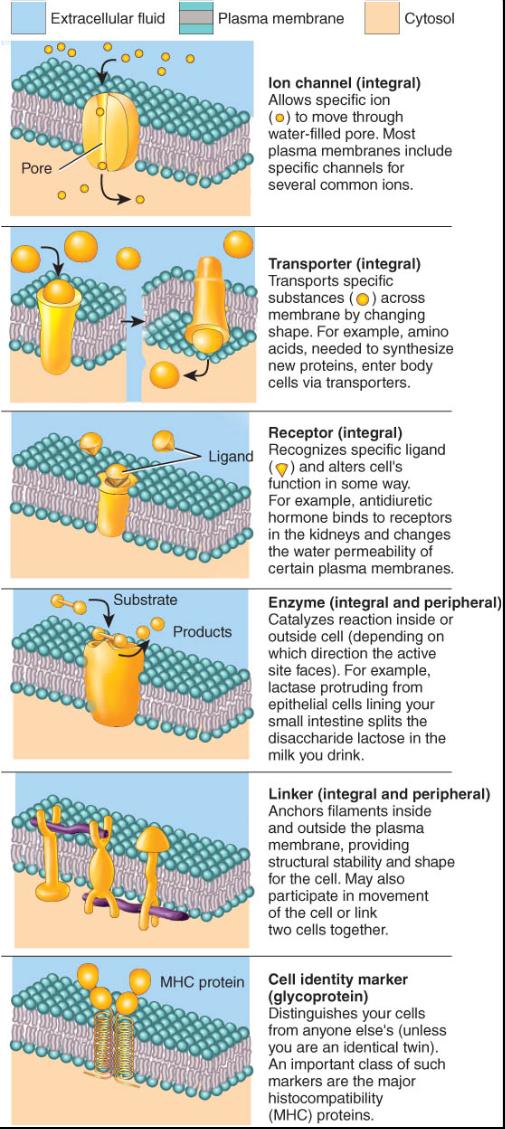{kind=link}
{kind=link}
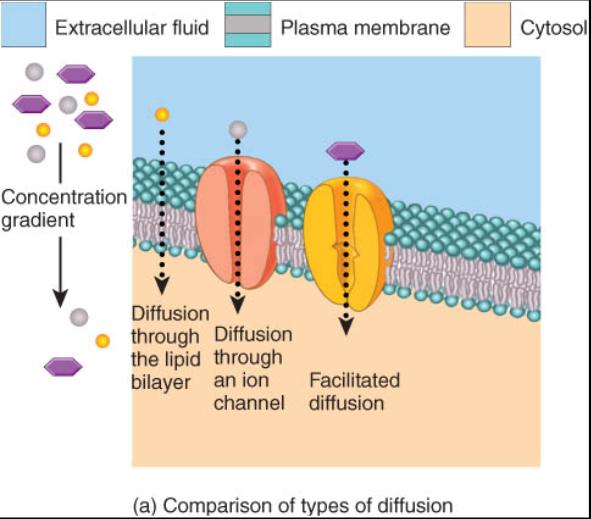{kind=link}
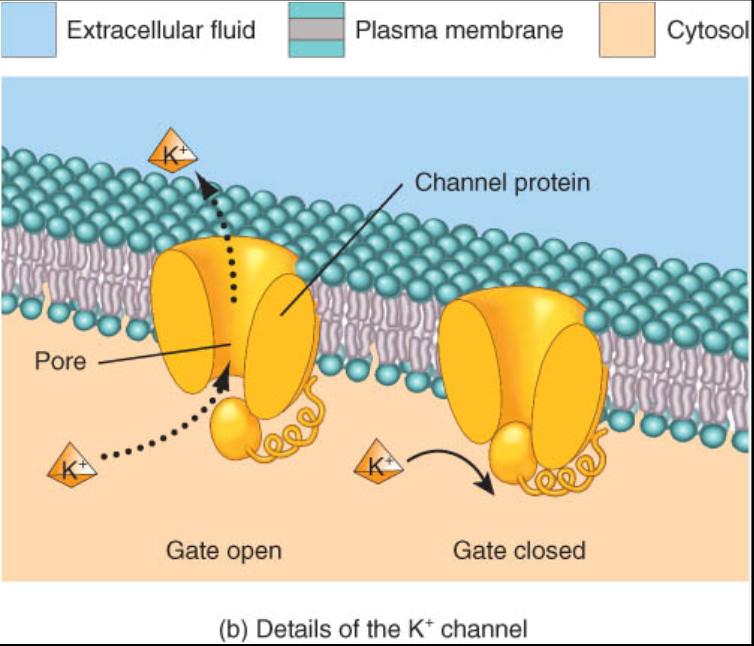{kind=link}
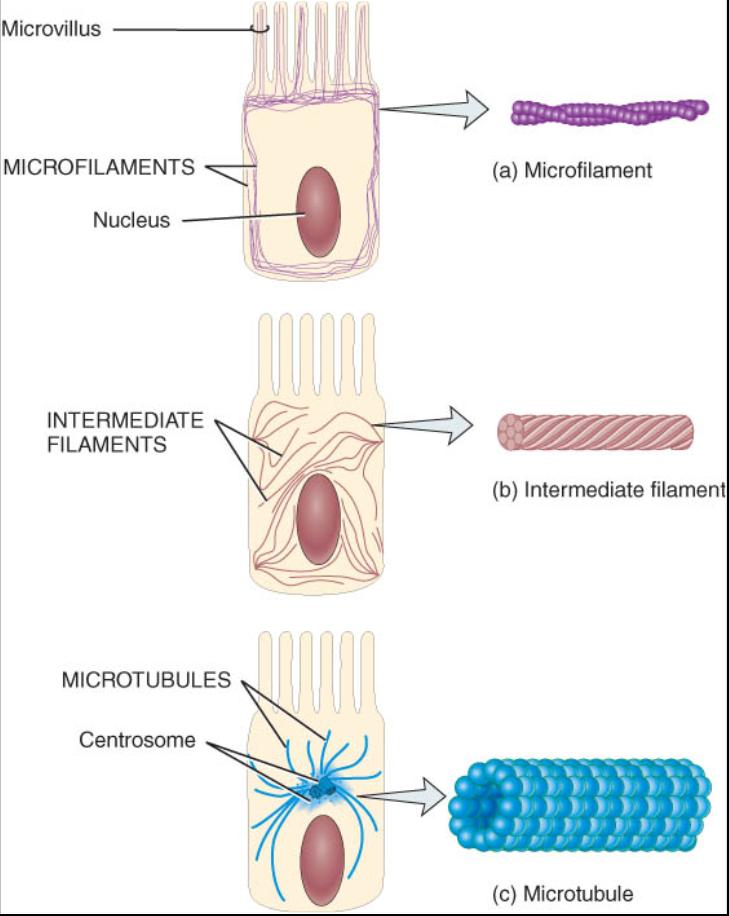{kind=link}
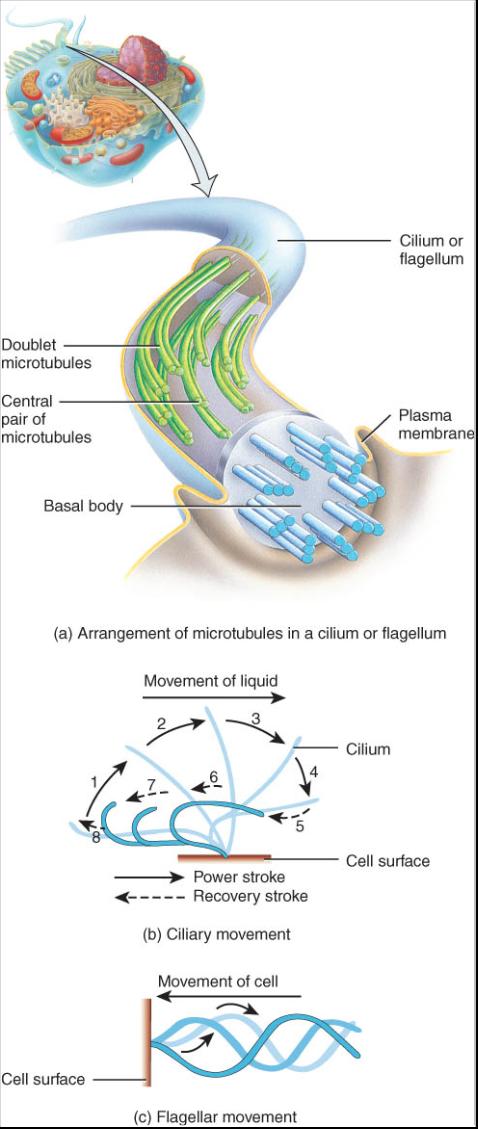{kind=link}
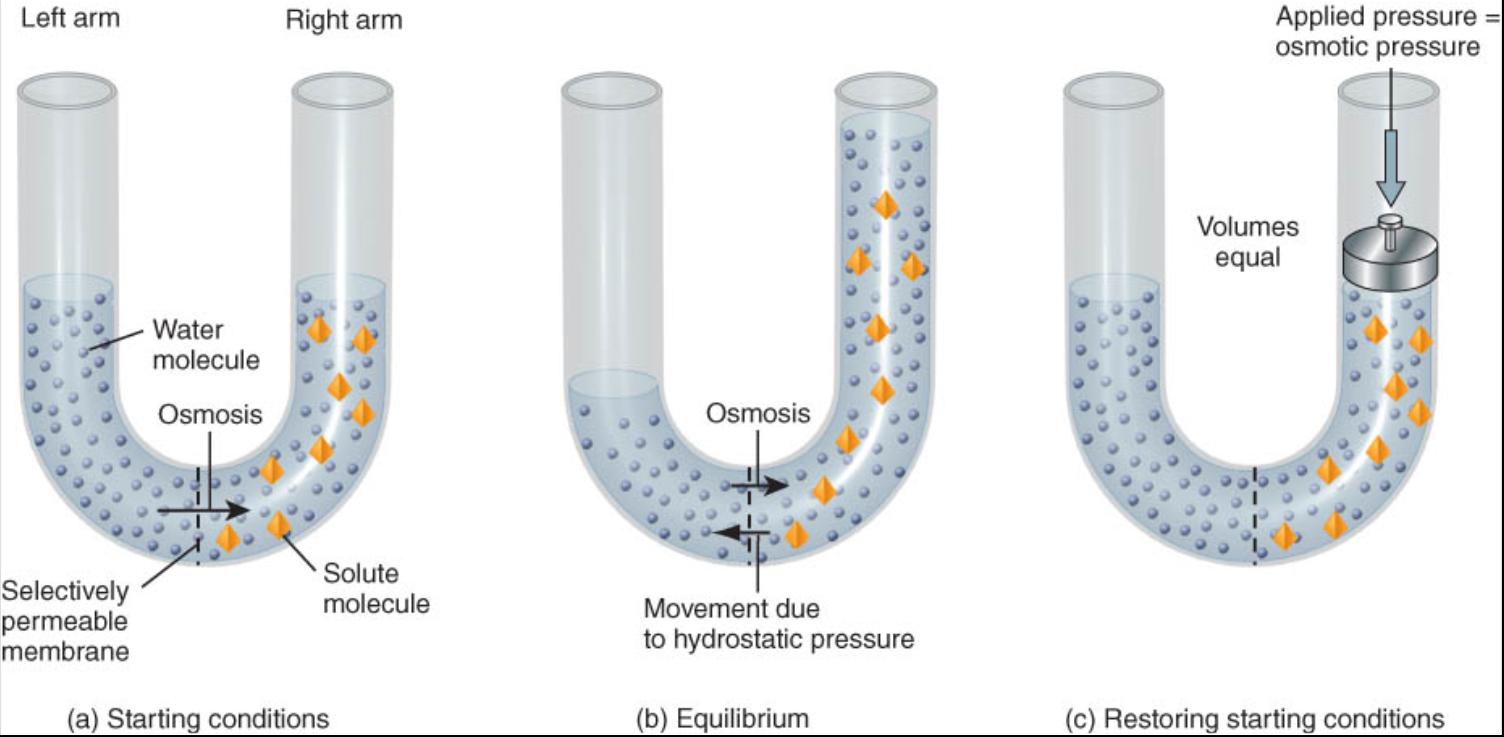{kind=link}
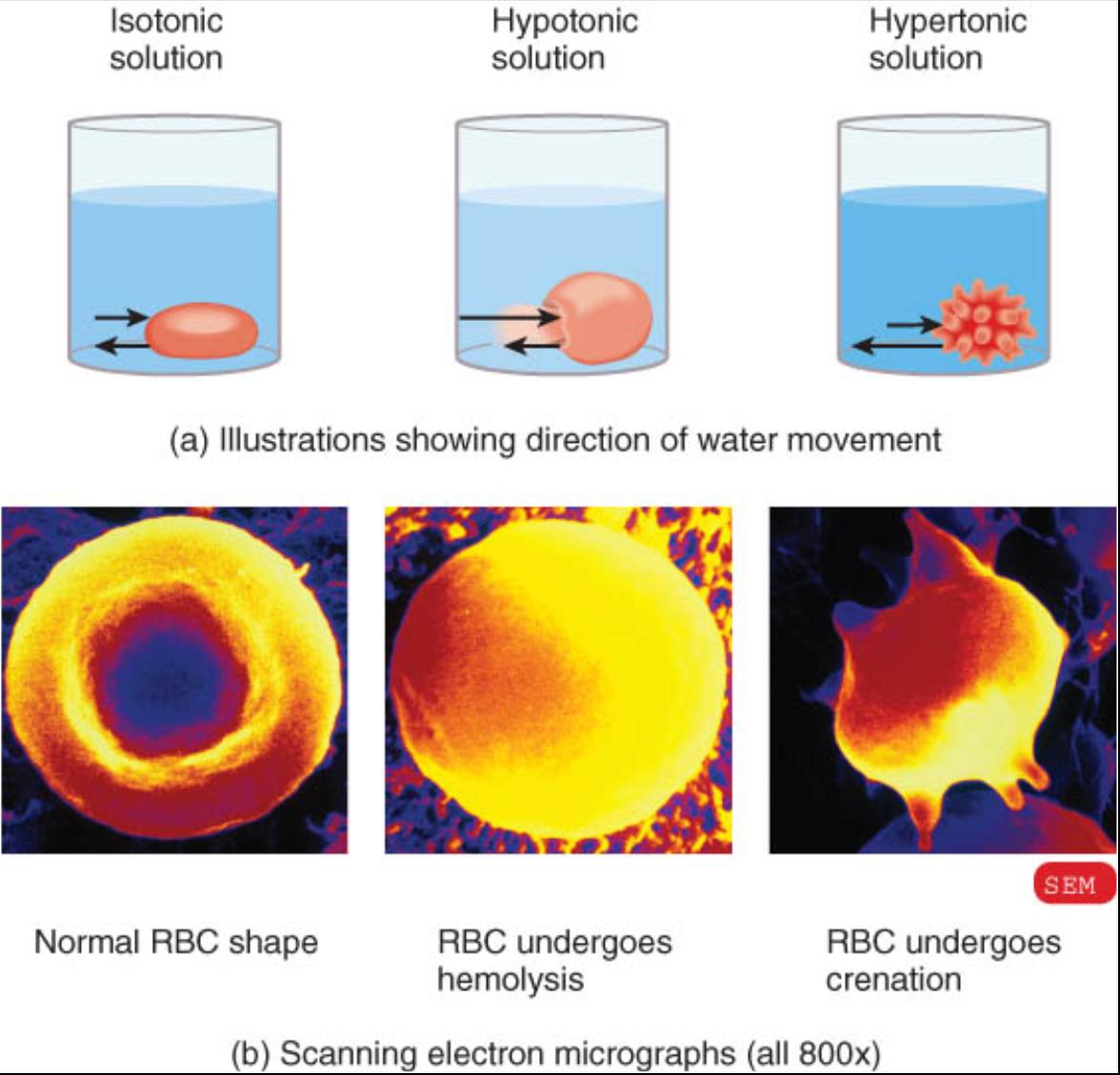{kind=link}
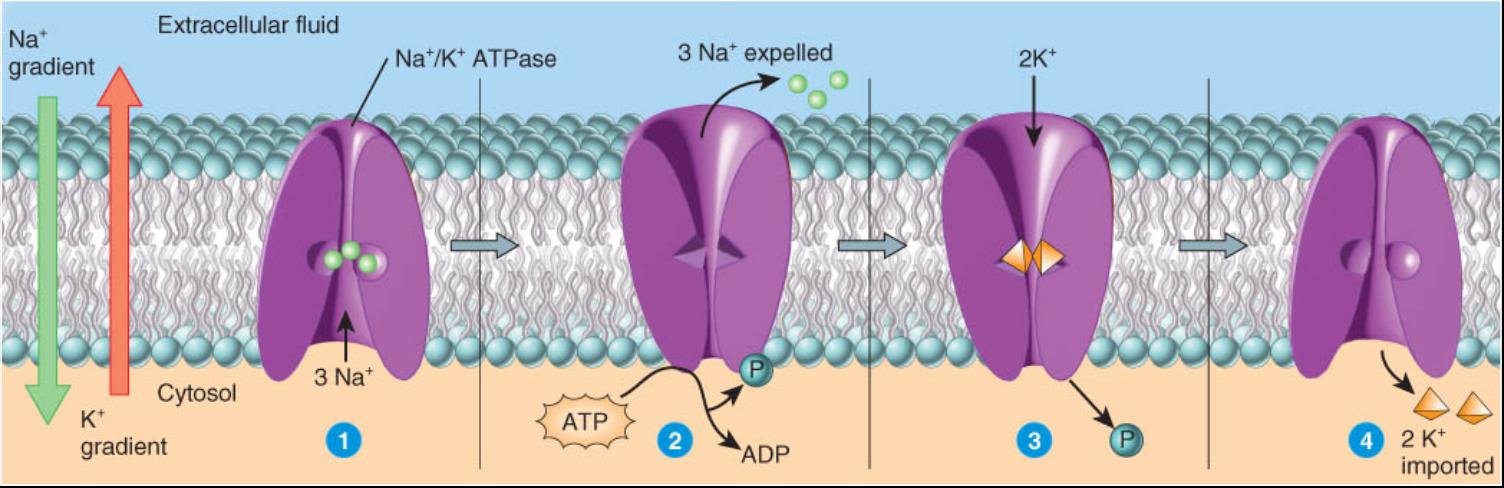{kind=link}
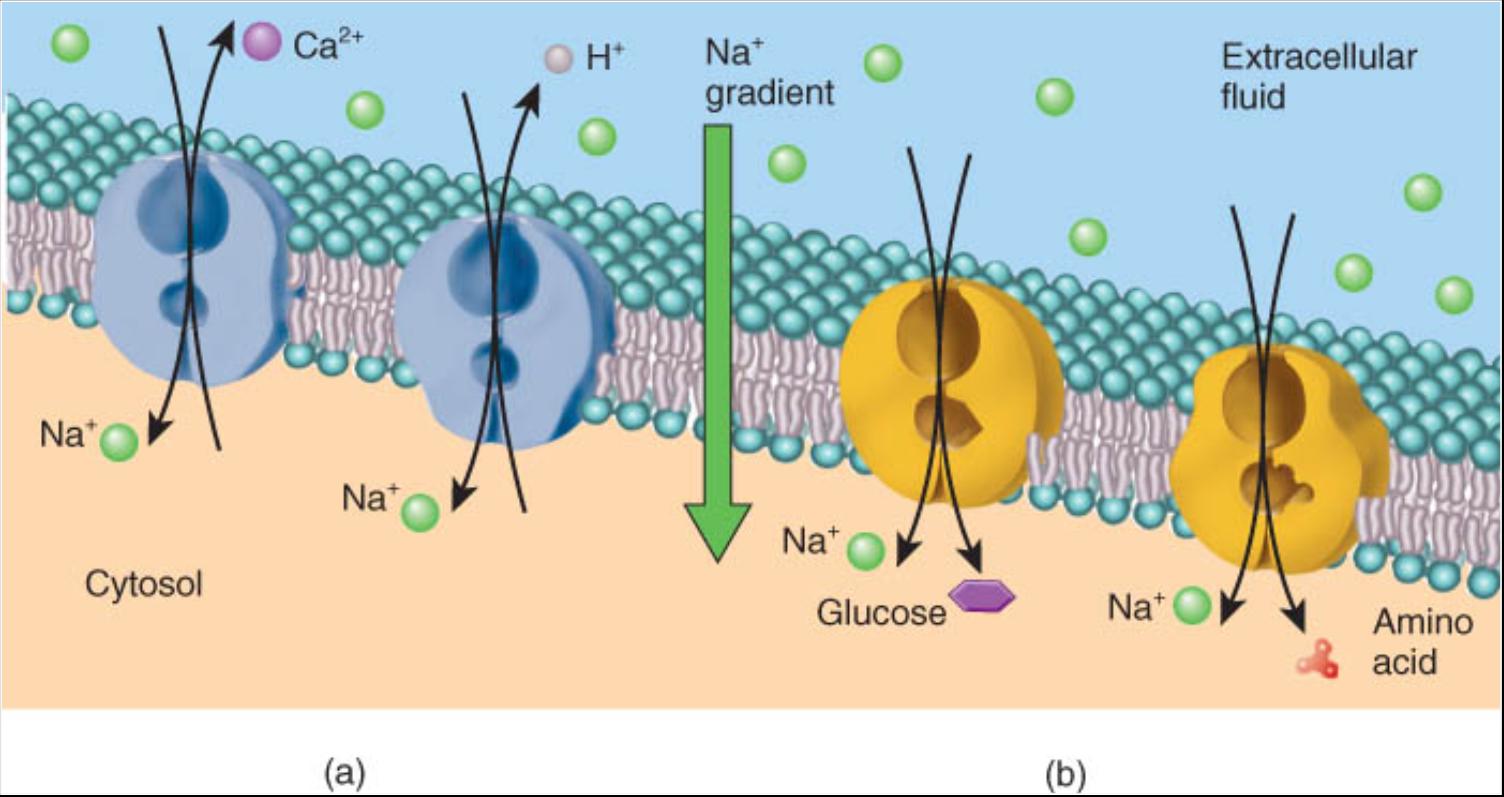{kind=link}
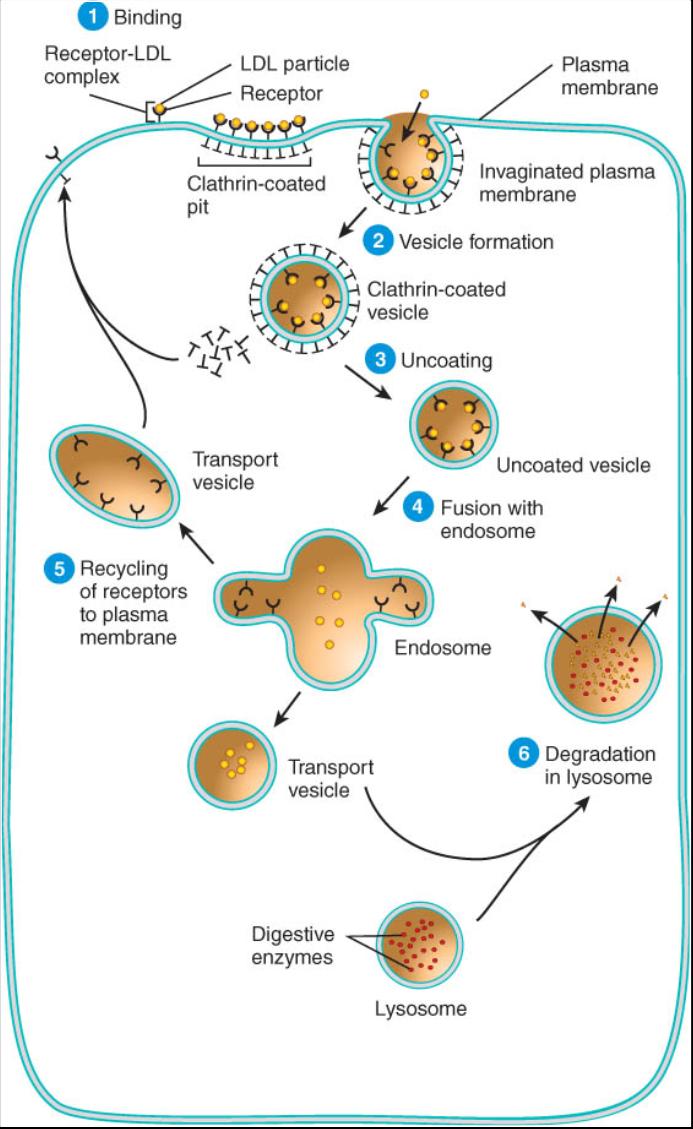{kind=link}
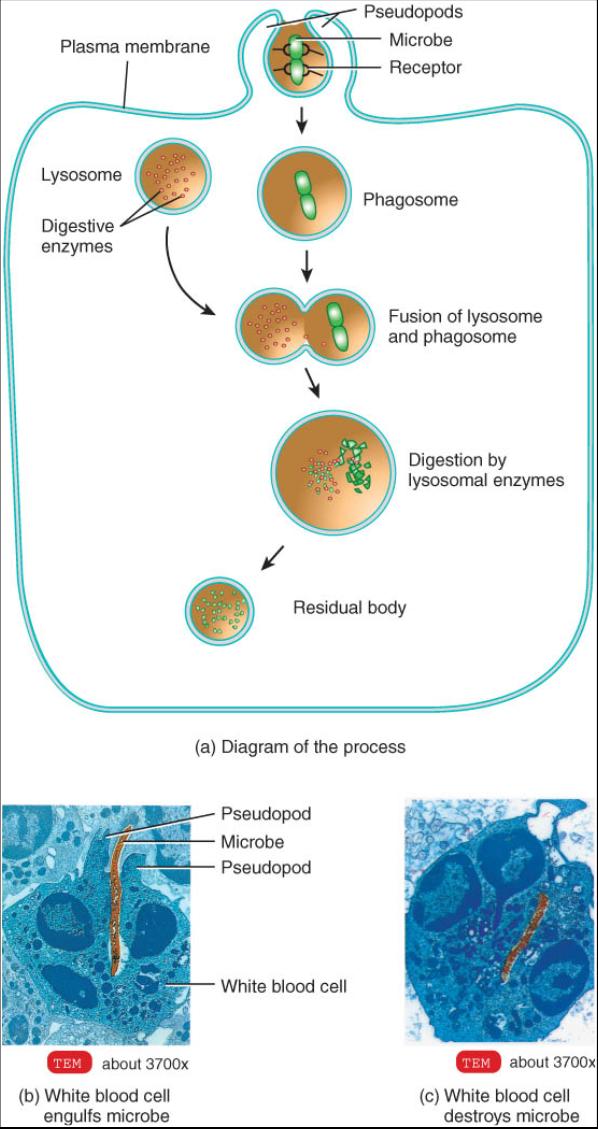{kind=link}
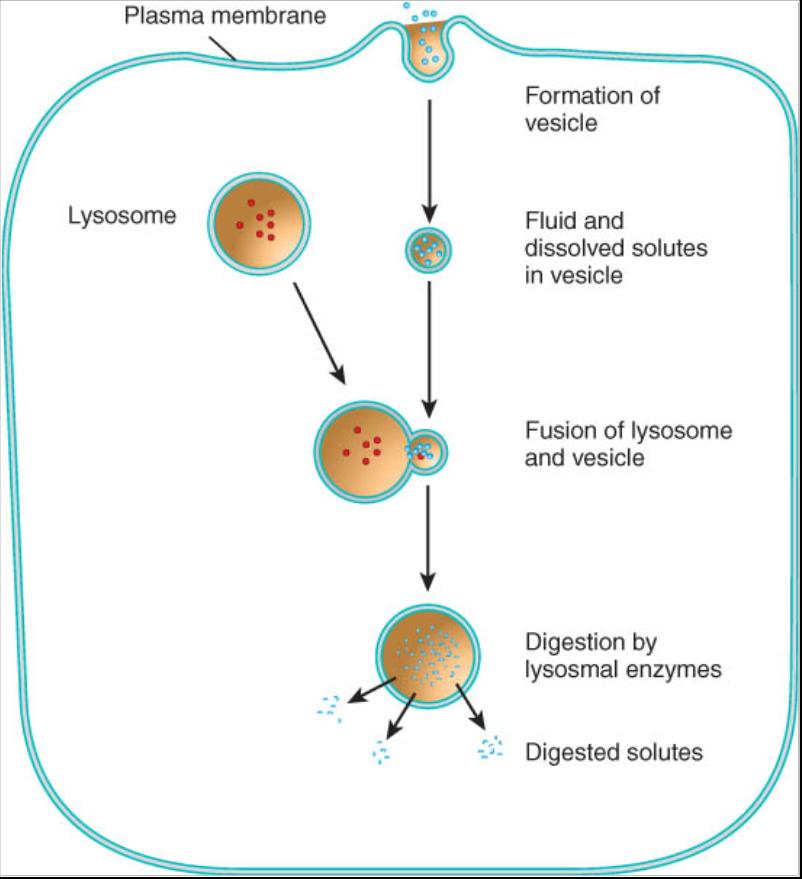{kind=link}
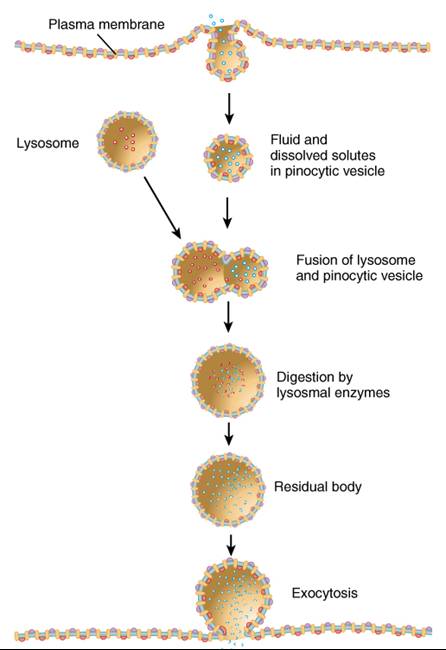{kind=link}
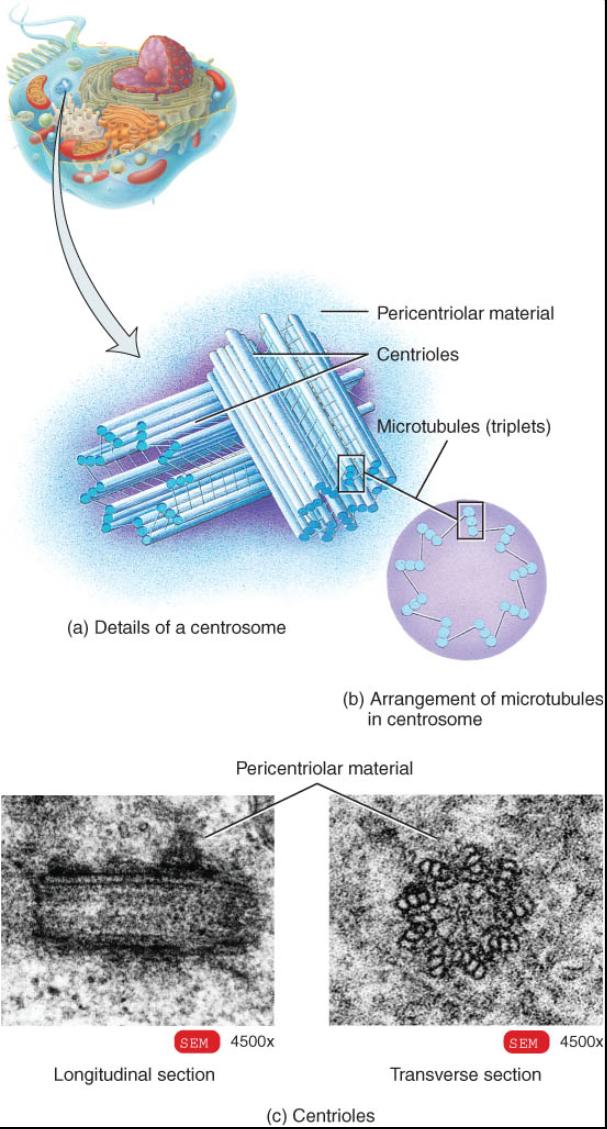{kind=link}
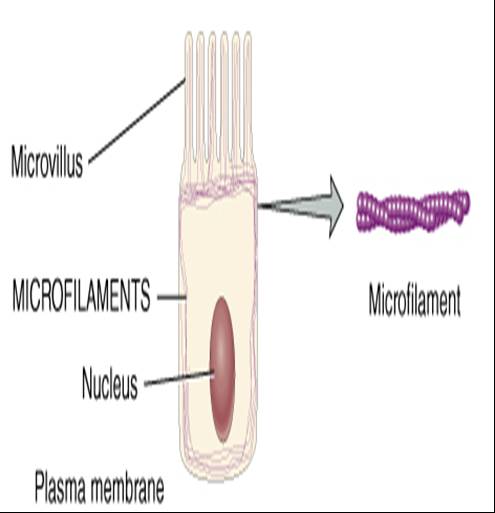{kind=link}
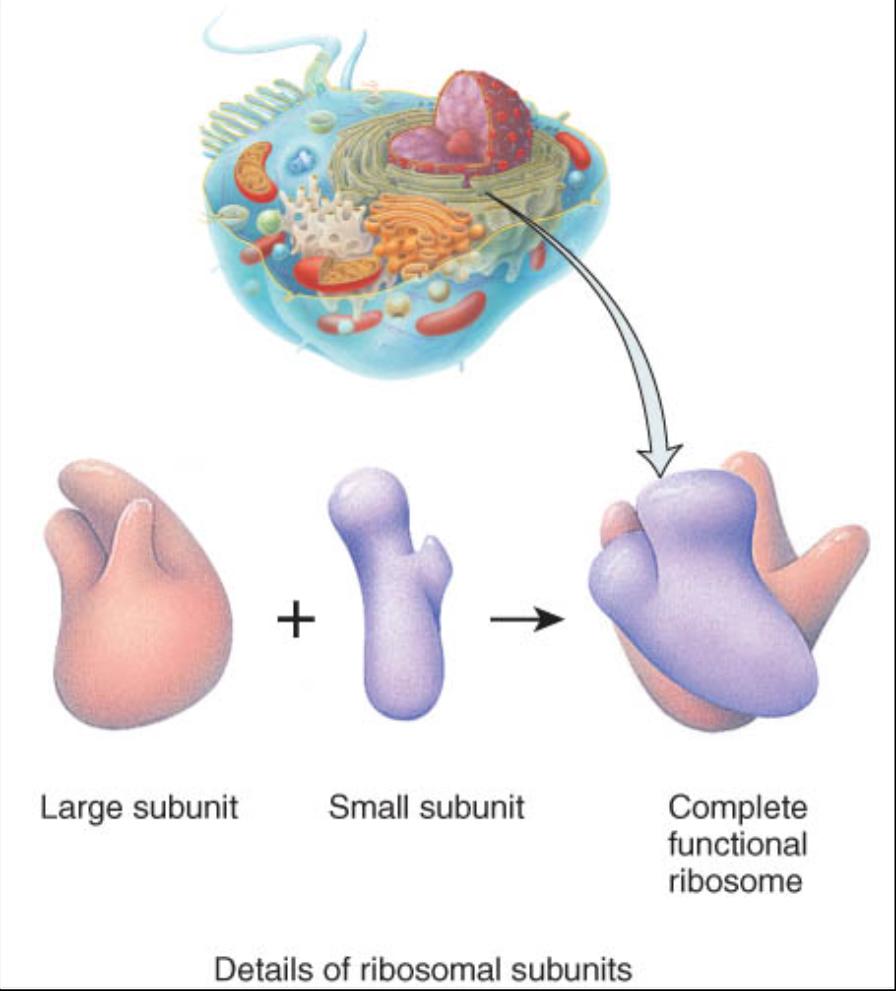{kind=link}
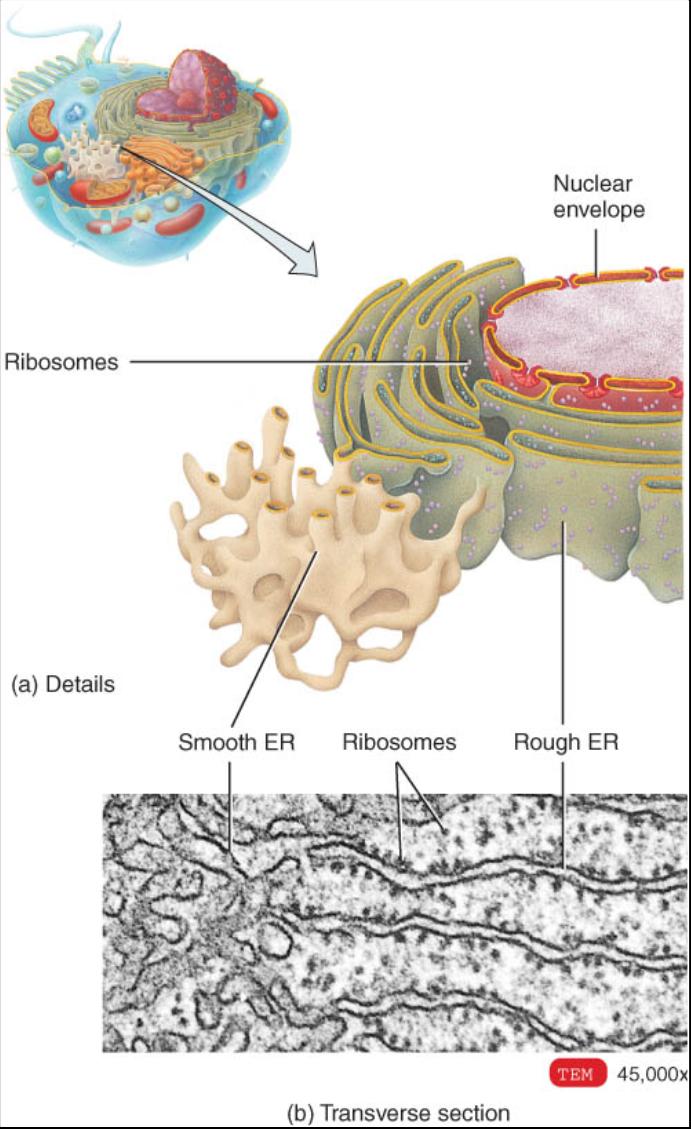{kind=link}
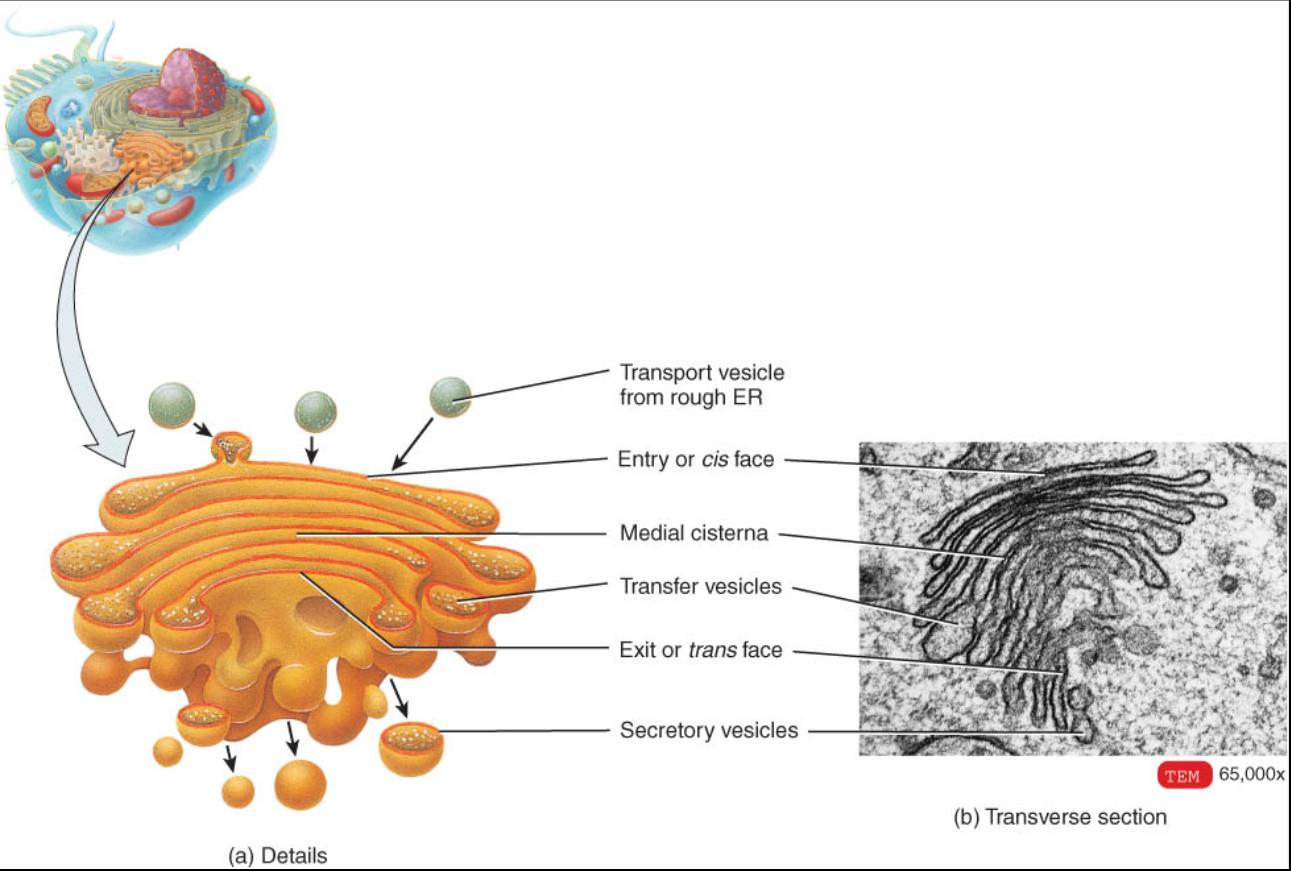{kind=link}
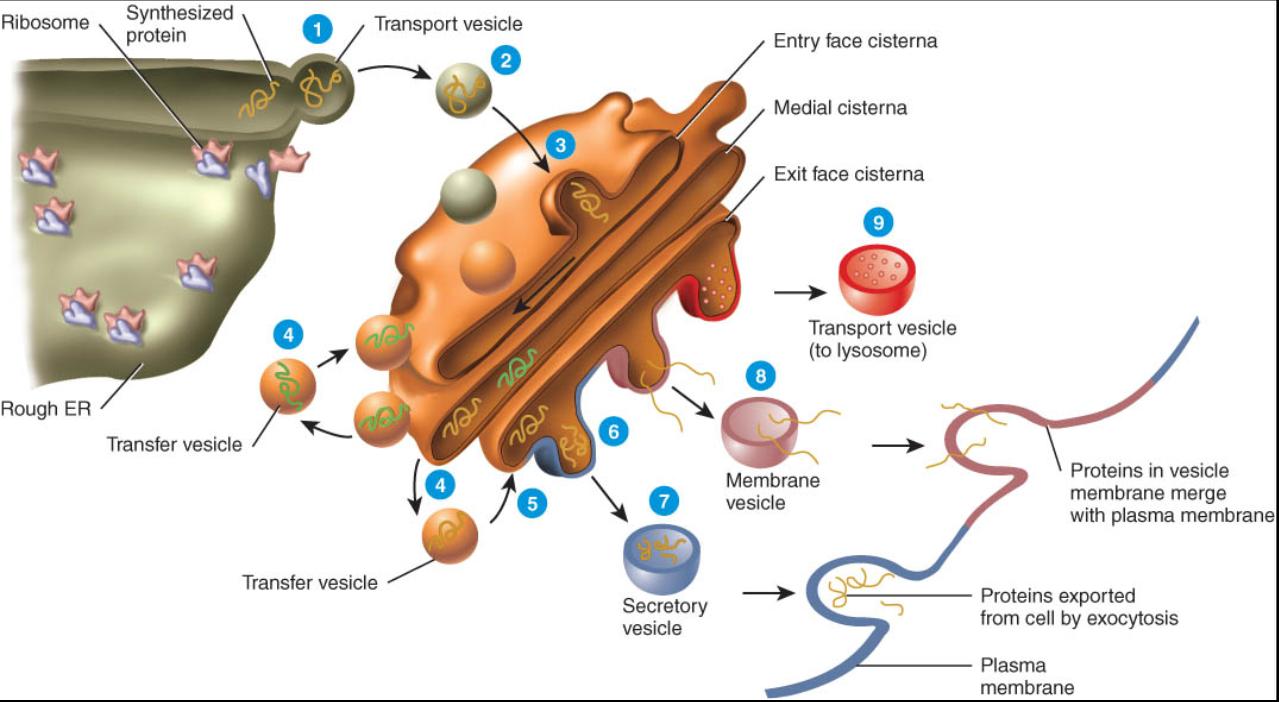{kind=link}
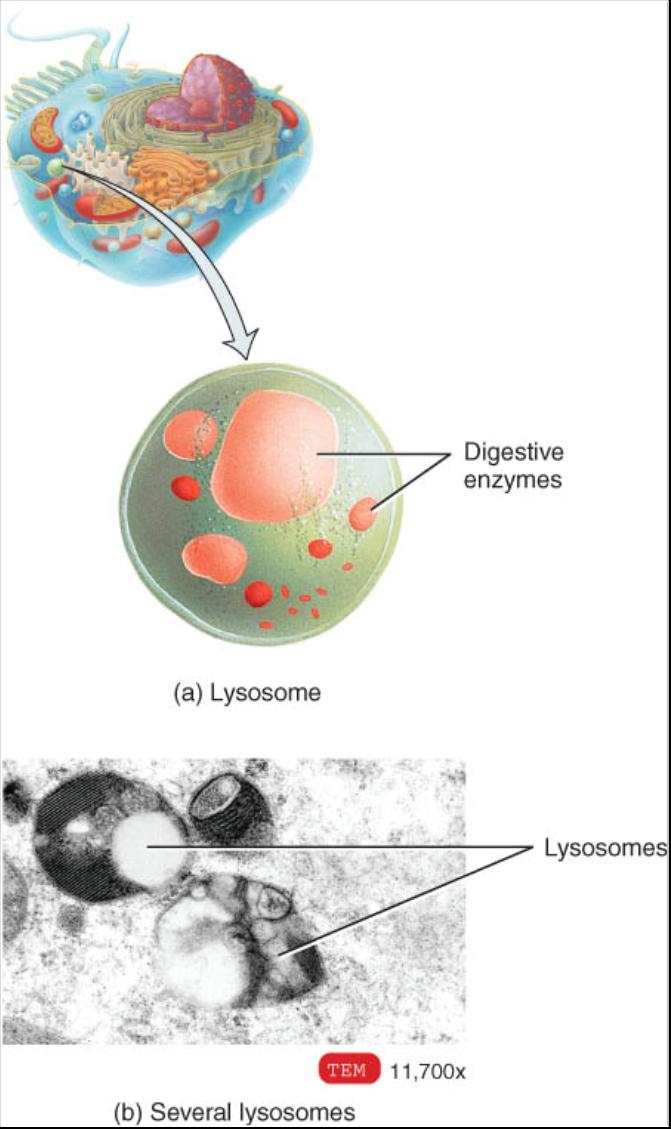{kind=link}
{kind=link}
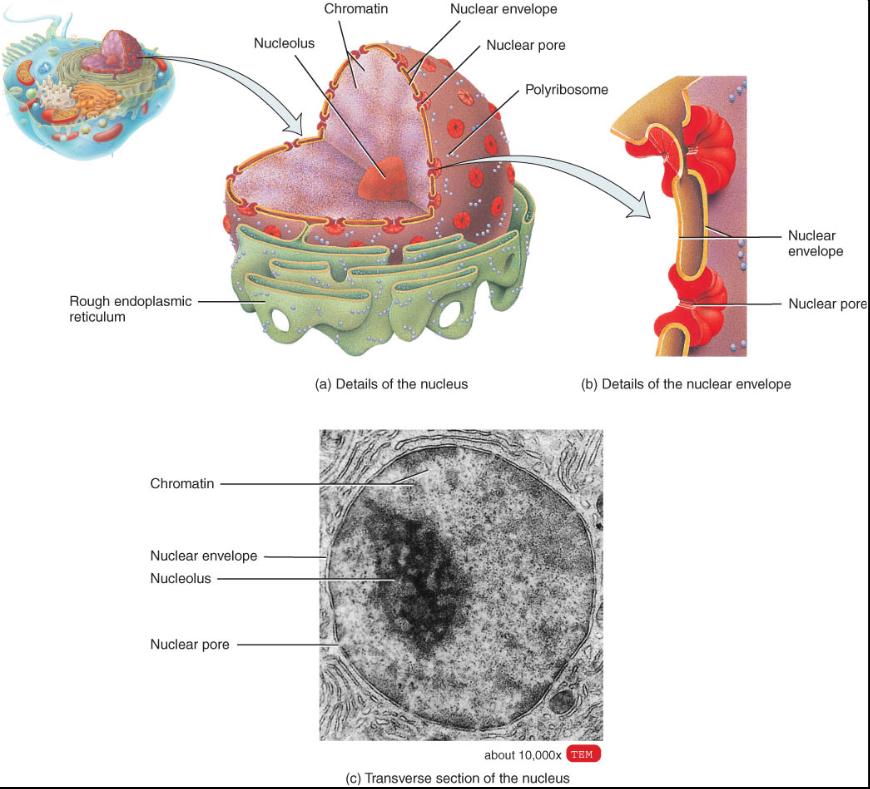{kind=link}
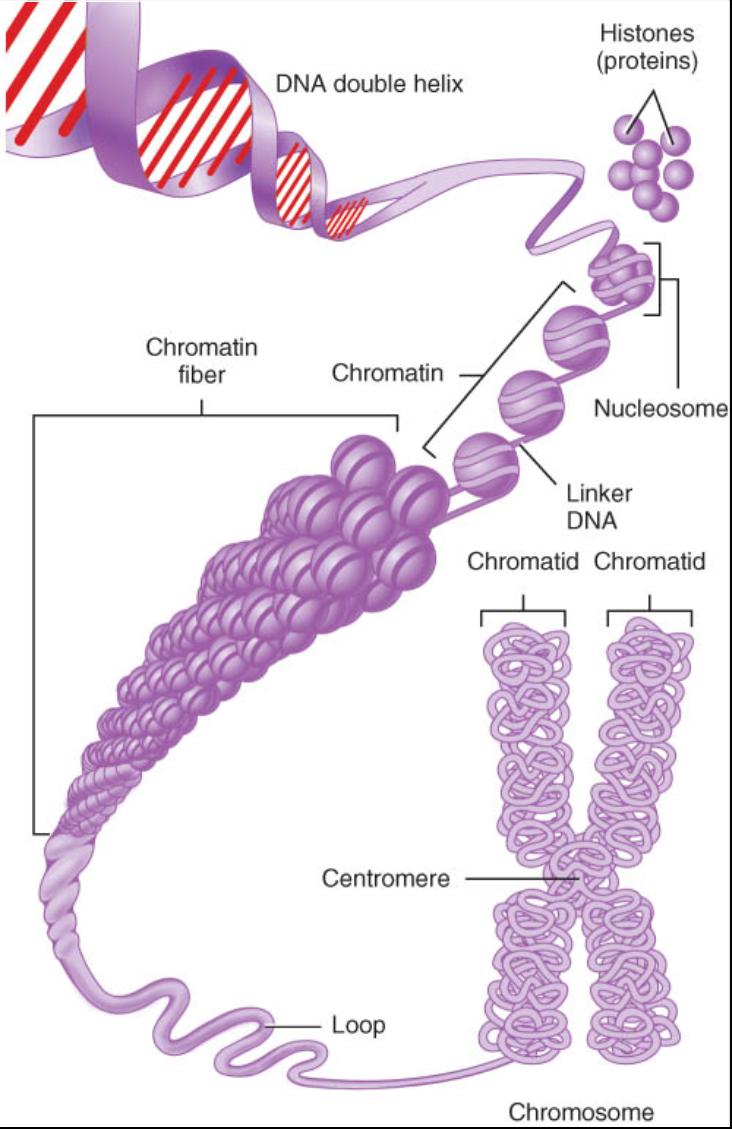{kind=link}
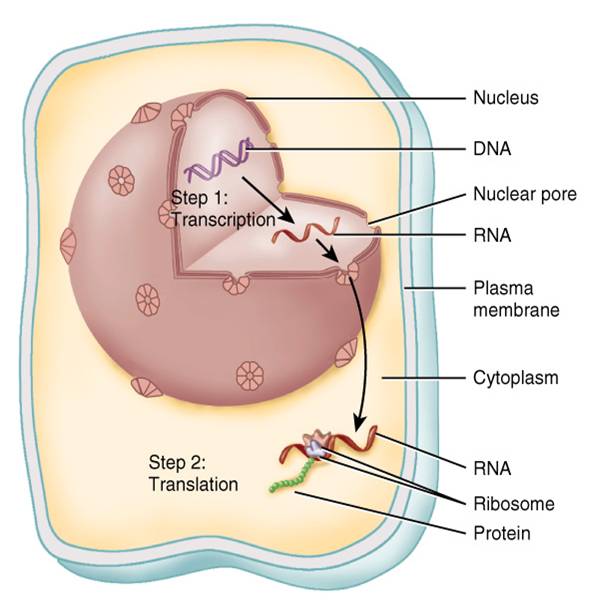{kind=link}
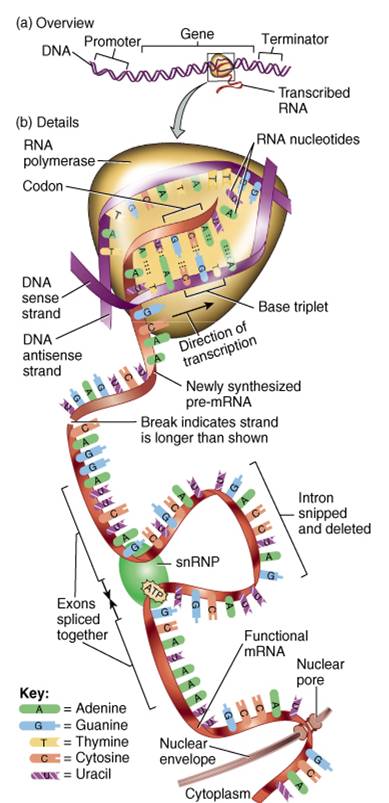{kind=link}
{kind=link}
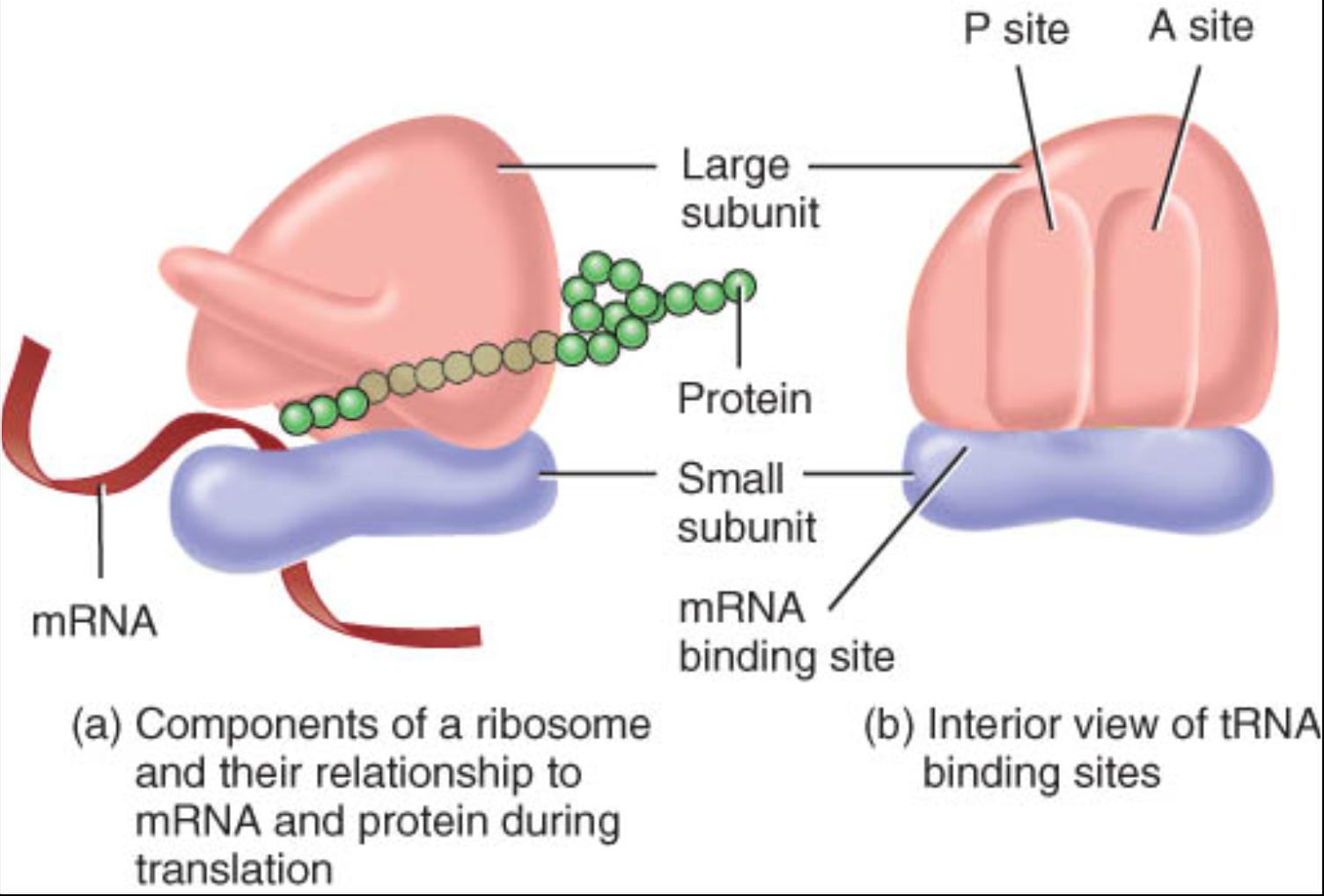{kind=link}
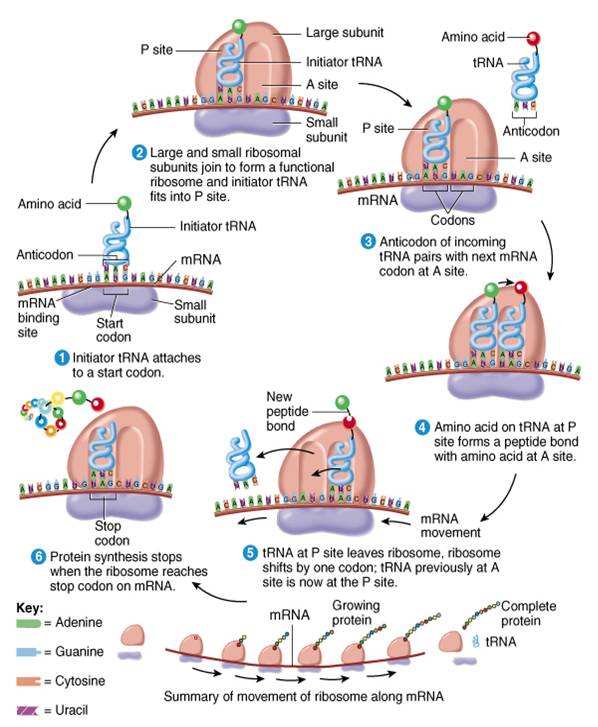{kind=link}
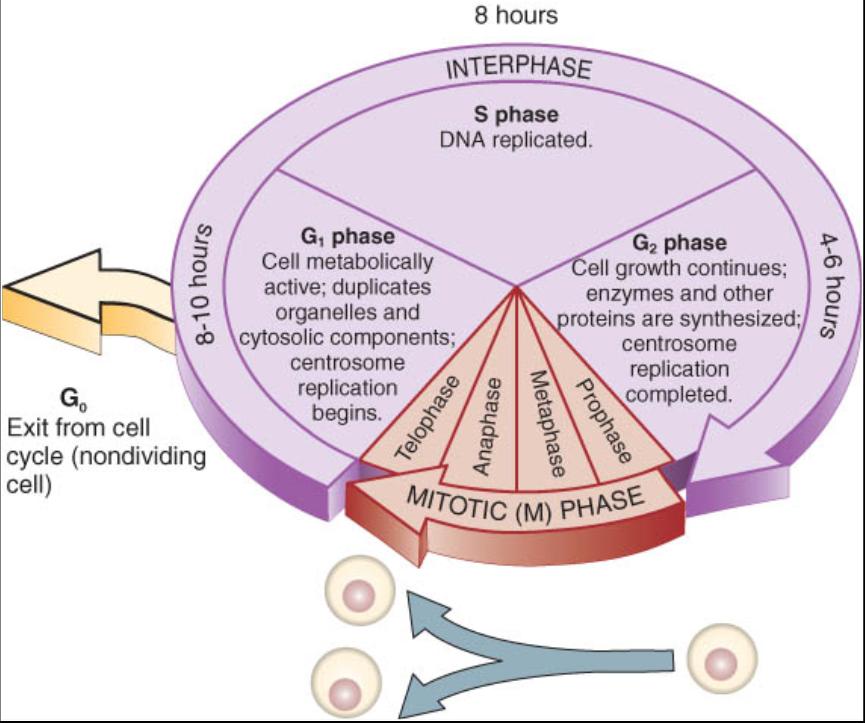{kind=link}
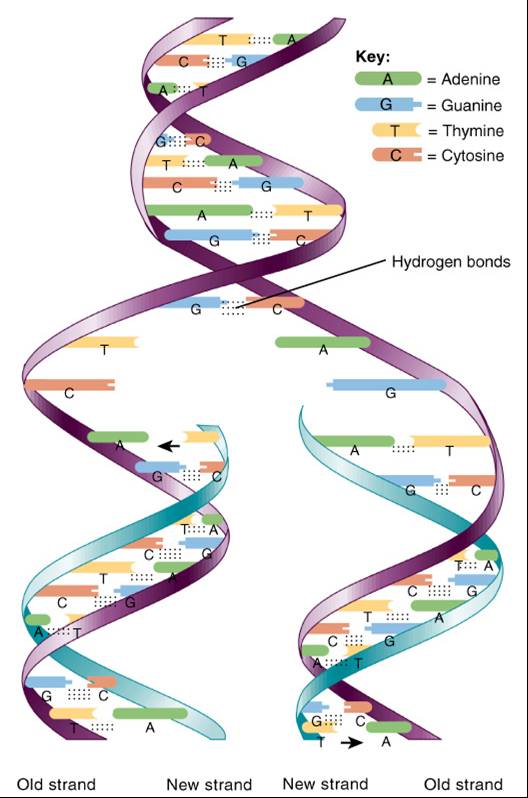{kind=link}
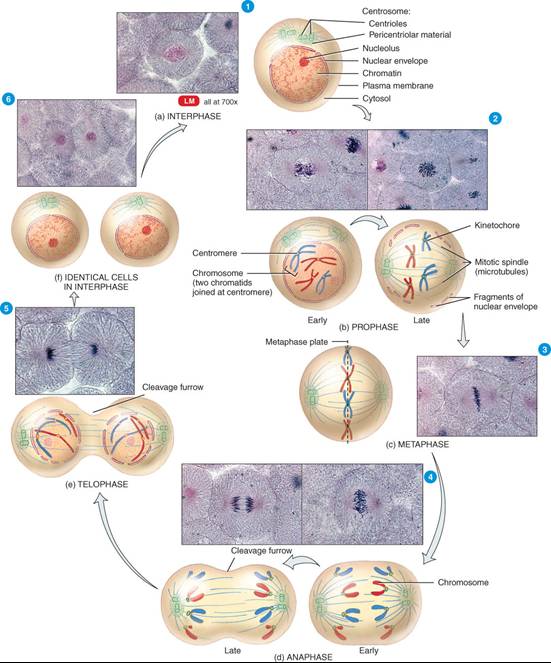{kind=link}
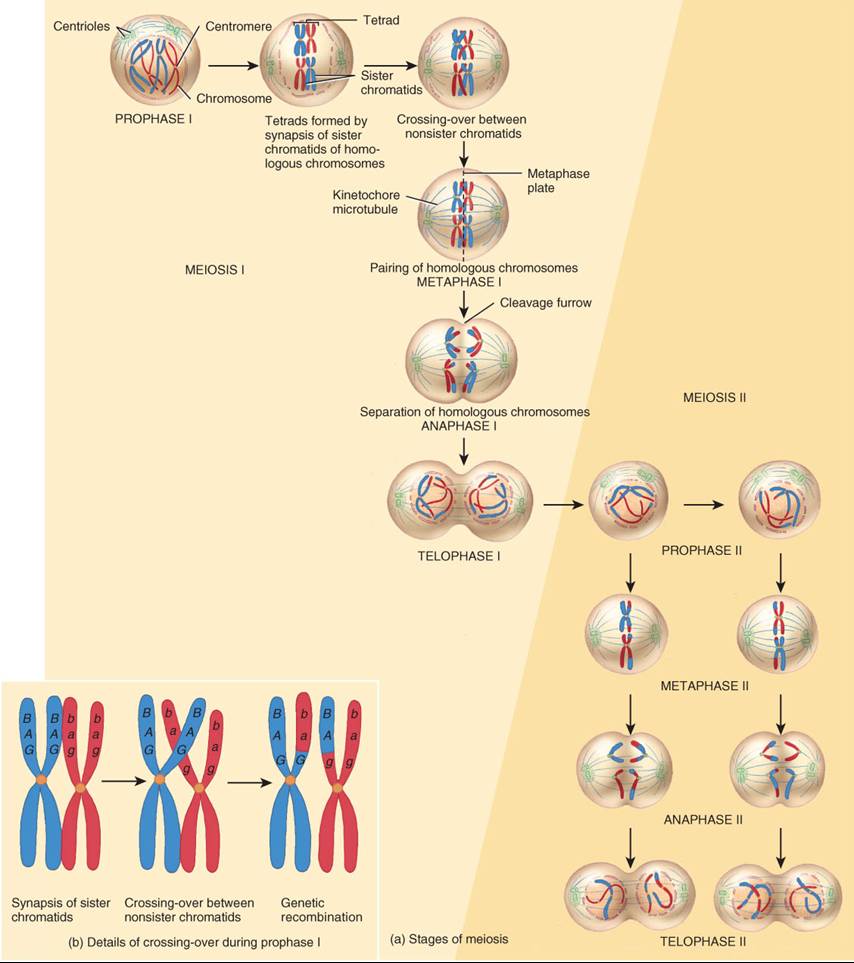{kind=link}
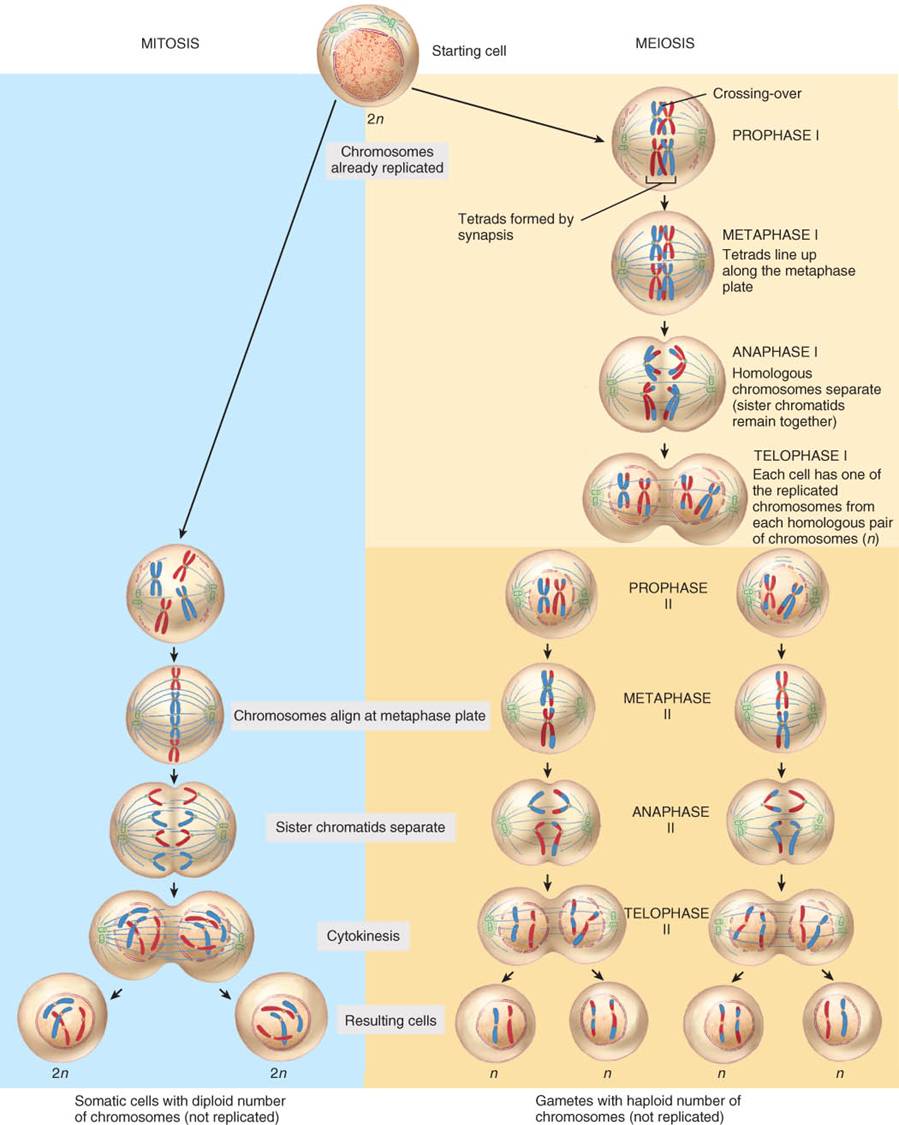{kind=link}
{kind=link}