Capítulo 8
SISTEMAS TEGUMENTARIO,
ESQUELÉTICO Y MUSCULAR
Células inmigrantes en la epidermis
Diferenciación epidérmica
Interacciones dérmico/ epidérmicas
Derivados epidérmicos
La construcción de los tejidos corporales implica procesos del desarrollo en dos niveles de organización. El primero es el de las células individuales, en el cual las células que componen un tejido se especializan en un proceso denominado citodiferenciación (véase la discusión sobre restricción, determinación y diferenciación en la Pág. 70). En el siguiente nivel de complejidad, varios tipos celulares se desarrollan de común acuerdo para formar tejidos específicos por medio de un proceso llamado histogénesis. En este capitulo se examina el desarrollo de tres importantes tejidos del cuerpo: la piel, los huesos y los músculos. La histogénesis de cada uno de ellos ejemplifica aspectos importantes del desarrollo.
La piel, que consta de la epidermis y la dermis, es una de las estructuras mas grandes del organismo. La epidermis representa la interfase entre el cuerpo y el ambiente externo, y su estructura esta bien adaptada a los requerimientos funcionales locales. La simple inspección de áreas como el cuero cabelludo y las palmas de las manos muestra que la estructura de la piel varia de una parte del cuerpo a otra. Estas variaciones locales son el resultado de las interacciones inductivas entre el ectodermo y el mesénquima subyacente. Las anomalías asociadas con el sistema tegumentario se presentan en la Correlación clínica 8_1, que se encuentra al final de esta sección.
Desarrollo estructural
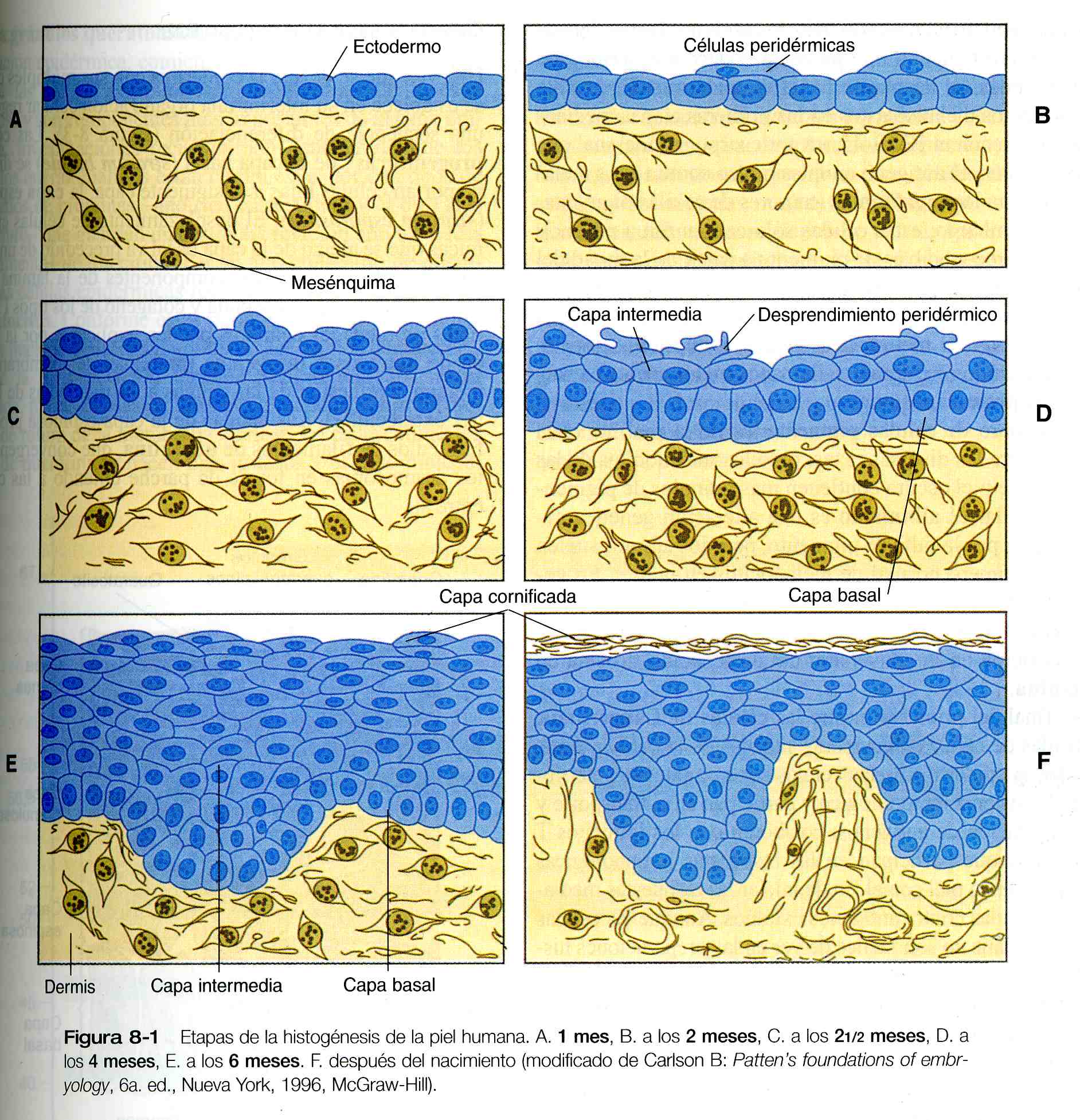
Al comienzo, la capa externa de la piel la forma una sola capa de células ectodérmicas (figura8_1 A). Conforme el desarrollo avanza, el ectodermo se transforma en varias capas y se hacen evidentes las diferencias regionales en su estructura.
La primera etapa en el desarrollo de las capas de la epidermis es la formación de una fina capa exterior de las células aplanadas conocida como peridermo, al final del primer mes de gestación (figura 8_1 B). Al parecer las células del peridermo, que esta presente en la epidermis de todos los embriones amniotas, participan en el intercambio de agua, sodio y posiblemente glucosa entre el liquido amniótico y la epidermis.
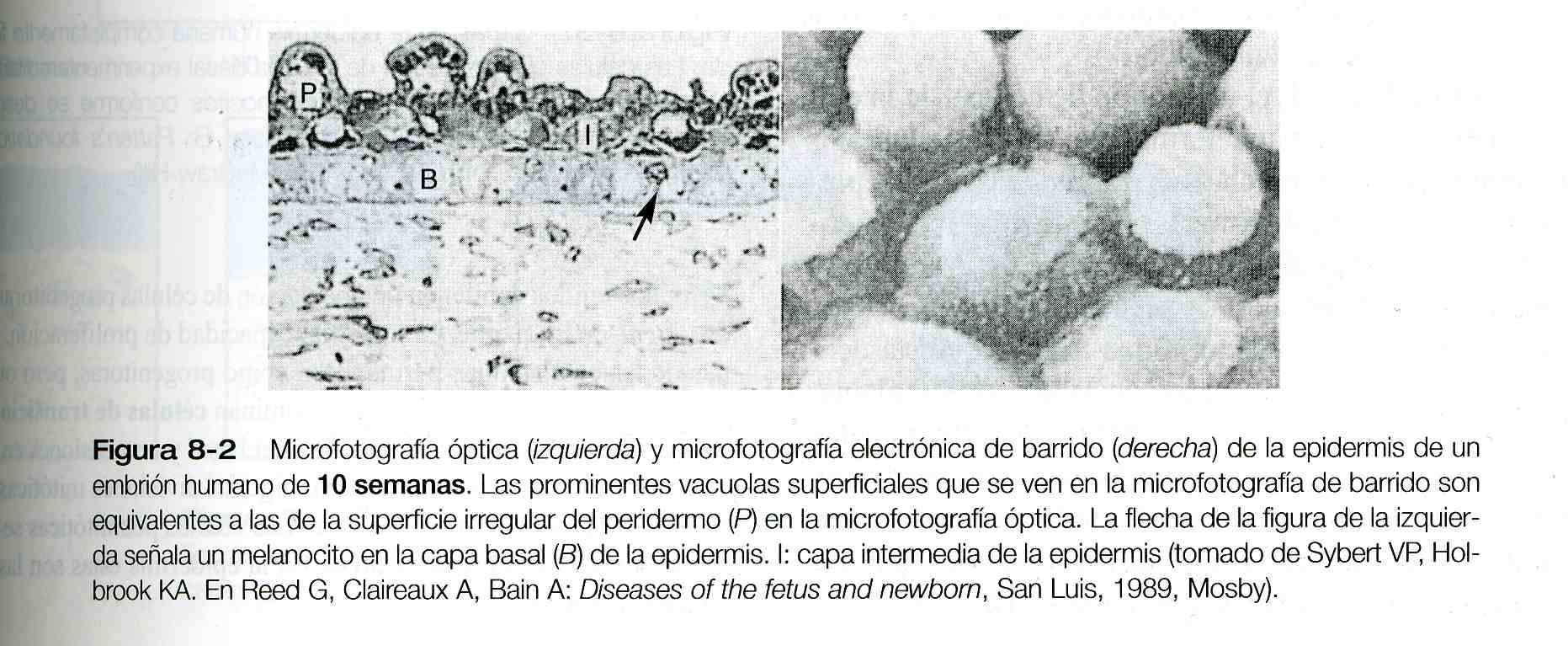
Hacia el tercer mes, la epidermis es una estructura de tres capas: una capa basal (o germinativa) que muestra mitosis activas, una capa intermedia de células (figura 8_1 D) que representa la progenie de las células progenitoras (stem cells) de la capa basal en división, y una capa superficial de células del peridermo que tienen unas vacuolas características en su superficie (figura 8_2). Las células peridérmicas contienen grandes cantidades de glucógeno, pero se desconoce la función de éste.
Durante el sexto mes la epidermis que esta por debajo del peridermo se diferencia en las capas definitivas características de la epidermis posnatal. Muchas de las células del peridermo sufren una muerte programada (apoptosis) y se desprenden en el liquido amniótico. La epidermis se convierte en una barrera entre el feto y el ambiente exterior, en lugar de una zona en el intercambio entre ambos. El cambio de función de la epidermis fetal puede ser de tipo adaptativo, ya que se produce cerca del momento en que los desechos urinarios comienzan a acumularse en el liquido amniótico.
Células inmigrantes en la epidermis
A pesar de su apariencia histológica homogénea, en realidad la epidermis es un mosaico celular, cuyas células se derivan no solo del ectodermo superficial sino también de otros precursores, como la cresta neural o el mesodermo, estas células desempeñan papeles específicos importantes en la función de la piel.
Al comienzo del segundo mes, los melanoblastos derivados de la cresta neural migran hacia la dermis embrionaria; poco después, se desplazan hacia la epidermis. Es posible reconocer los melanoblastos en las primeras etapas mediante la tinción con el HMB_45, un anticuerpo monoclonal que reacciona con un antígeno citoplasmático común a los melanoblastos y a los melanomas (tumores de las células pigmentarias); sin embargo, estas células solo comienzan a producir cantidades reconocibles de pigmento a partir de la mitad del embarazo. Esto ocurre mas pronto en las personas de piel muy pigmentada que en las blancas. La diferenciación de los melanoblastos en melanocitos maduros implica la formación de gránulos de pigmento llamados melanosomas a partir de los premelanosomas.
El numero de células pigmentarias de la piel no varia mucho entre las diversas razas, pero los melanocitos de las personas de piel oscura tienen mas gránulos de pigmento por célula. El albinismo es una alteración genética caracterizada por la falta de pigmento, pero los albinos suelen tener un numero normal de melanocitos en la piel. En general, los melanocitos de los albinos no pueden expresar la pigmentación debido a que les falta la enzima tirosinasa, que participa en la conversión del aminoácido tirosina en melanina.
Al final del primer trimestre las células de langerhans, derivadas de células precursoras situadas en la medula ósea, invaden la epidermis. Estas células son componentes periféricos del sistema inmunológico y participan en el transporte y la presentación de antígenos; colaboran con los linfocitos T (leucocitos que participan en las respuestas inmunológicas celulares) en la piel con el fin de iniciar las respuestas mediadas por células contra antígenos extraños. Aun que las células de Langerhans no son fáciles de ver en las preparaciones histológicas habituales, pueden distinguirse si se dirigen anticuerpos contra los antígenos específicos de la superficie celular o mediante la demostración histoquímica de su gran actividad de adenosintrifosfatasa ligada a su membrana. Las células de Langerhans están presentes en cantidades pequeñas (alrededor de 65 células /mm2 de epidermis) durante los dos primeros trimestres de embarazo, pero después su numero aumenta varias veces hasta constituir el 2 al 6 % del total de las células epidérmicas del adulto.
Las células de Merkel, el tercer tipo de célula de la epidermis, tiene un origen indeterminado. Algunos indicios sugieren que pueden diferenciarse a partir de las células precursoras de las células epidérmicas, sin embargo los experimentos en aves mediante el injerto de tejidos de la cresta neural marcados indican que los precursores de las células de Merkel migran hacia la extremidad desde la cresta neural. Estas células, que ya aparecen en la epidermis plantar entre las ocho y las doce semanas de gestación, se asocian con terminaciones nerviosas libres. Actúan como mecanorreceptores de adaptación lenta a la piel, pero las evidencias histoquímicas sugieren que en algún momento también puede funcionar como células neuroendocrinas.
Diferenciación epidérmica
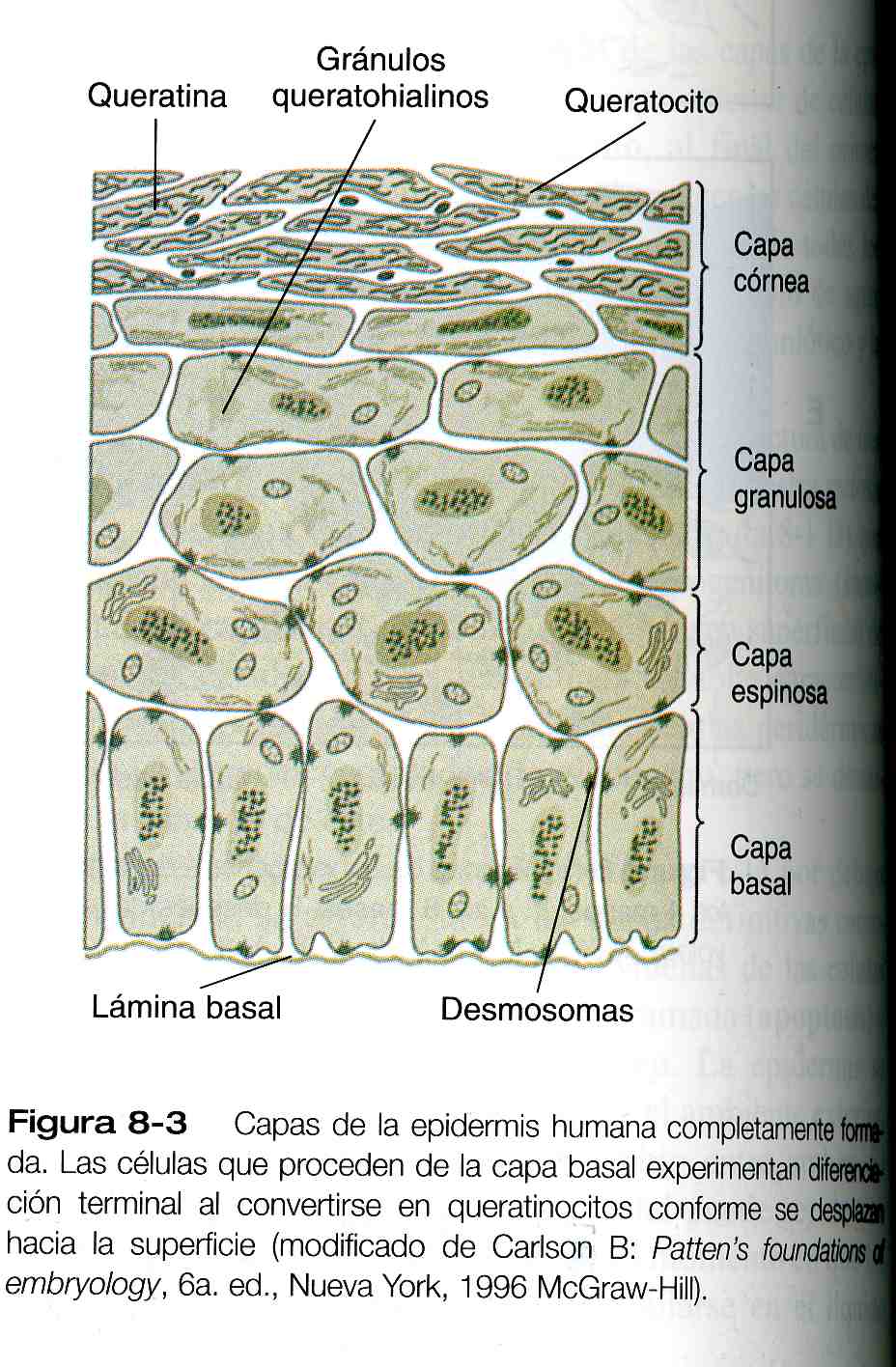
Una vez que se define la epidermis con sus múltiples capas, se establece en su interior una organización celular regular y una secuencia de diferenciación (figura 8_3). Las células progenitoras de la capa basal (stratum basale) se dividen y aportan células hijas a la siguiente capa, la capa espinosa (stratum espinosum). El desplazamiento de las células epidérmicas que se alejan de la capa basal va precedido de una pérdida de adhesividad a los componentes de la lámina basal (como fibronectina, laminina y colágeno de los tipos I y IV). Estas propiedades celulares pueden explicarse por la perdida de numerosas integrinas (proteínas de membrana que median la adhesión de las células a las moléculas de la matriz extracelular). Las células de la capa espinosa producen fascículos de filamentos de queratina, que convergen sobre los desmosomas en forma de parche uniendo a las células entre si.
Los gránulos queratohialinos, otro marcador de la diferenciación epidérmica, comienza a aparecer en el citoplasma de las células posmitóticas mas externas de la capa espinosa y son componentes mas importantes de la capa granulosa. Investigaciones recientes han demostrado por dos tipos de agregados de proteínas –uno rico en la histidina y el otro rico en sulfuro- que guardan una estrecha relación con los haces de filamentos de queratina. Por su alto contenido de queratina, las células epidérmicas reciben el nombre genérico de queratocitos. Conforme éstas pasan hacia la capa granulosa, sus núcleos comienzan a mostrar los signos característicos de la diferenciación terminal, como una apariencia aplanada, masas densas de cromatina nuclear, y signos precoces de rotura en la membrana nuclear. En estas células los haces de queratina se hacen mas gruesos y el peso molecular de las queratinas se sintetizan son más altos que los queratocitos menos maduros.
A medida que las células de desplazan hacia la capa exterior –la capa córnea (stratum corneum)-, pierden un núcleo y se parecen a sacos aplanados densamente almacenados con filamentos de queratina. Las células de esta capa están interconectadas mediante la filagrina, una proteína rica en histidina, que se deriva de unos de los componentes granulares de la queratohialina. Según la región de la superficie corporal, las células del estrato córneo se acumulan para formar alrededor de quince a 20 capas de células muertas terminan por desprenderse (por ejemplo, unas 1.300 células /cm2/h en el antebrazo humano) y por lo común se acumulan en el polvo casero.
La investigación bioquímica ha correlacionado la expresión de las proteínas de la queratina (miembros de una compleja familia de proteínas) con etapas especificas de la citodiferenciación epidérmica (figura 8_4). Las queratinas del grupo de los filamentos intermedios se expresan primero en células de la epidermis de dos capas durante el segundo mes del embarazo. Tres queratinas (40-, 45- y 52- kDa) son características de los epitelios simples y las otras dos (50- y 58- kDa), de los epitelios estratificados. Cuando la epidermis comienza a estratificarse (entre la novena y la duodécima semana de gestación), las células externas de la capa intermedia empiezan a expresar pequeñas cantidades de queratina 56,5 y 67- kDa, las cuales son características de la epidermis queratinizada. En las semanas siguientes las cantidades de estas dos queratinas aumenta considerablemente, mientras que las queratinas de los epitelios simples (40-, 45- y 52- kDa) disminuye al final del periodo fetal. La expresión de la filagrina, la proteína de unión intercelular, guarda una estrecha correlación con la posterior diferenciación de las capas cornificadas exteriores de la epidermis fetal.
La proliferación de la células epidérmicas basales ésta determinada por numerosos factores de crecimiento, algunos de los cuales estimulan la mitosis mientras otros la inhiben (cuadro 8_1). Los queratocitos suelen tardar cerca de cuatro semanas en pasar desde la capa basal de la epidermis hasta la ultima zona de descamación, pero en algunas enfermedades cutáneas como la psoriasis la proliferación de las células epidérmicas está mal controlada y los queratocitos pueden desprenderse a la semana de haberse generado.
Una de las principales características de la piel, en especial de la piel gruesa que cubre las palmas y las plantas, es la presencia de crestas y pliegues. Sobre las puntas de los dedos, las crestas forman pliegues y remolinos en los patrones de las huellas digitales que son únicos para cada individuo. Estos patrones son la base que permite el análisis genético y la investigación criminal a partir del estudio de tales diseños.
La formación de las crestas epidérmicas se asocia estrechamente con la aparición más precoz de las almohadillas de las superficies ventrales de los dedos de las manos y pies (figura 8_5). Las almohadillas se forman primero en las palmas, alrededor de las 6 ½ semanas y hacia 7 ½ semanas
se han formado en los dedos. Comienzan a regresar las 10 ½ semanas, pero mientras que están presentes establecen las bases para la información de las crestas epidérmicas, que se produce entre las semanas 11 y 17. Los sucesos equivalentes en los pies se ocurren casi una semana mas tarde que en la mano.
El patrón de las crestas epidérmicas se correlaciona con la morfología que tienen almohadillas ventrales cuando se forman las crestas. Si la almohadilla es alta y redonda, las crestas epidérmicas forman un remolino; si es baja y arqueada, el resultado es un arco. Una almohadilla de altura intermedia da lugar a una configuración en forma de pliegue de las crestas epidérmicas digitales. Al parecer, el momento de la formación también influye sobre la morfología; la formación precoz de las crestas se asocia con remolinos y la tardía con los arcos. Todavía no se ha comprendido la base de los patrones dermatoglíficos.
Cuando las crestas epidérmicas empiezan a formarse, las puntas de los dedos aún son lisas y la epidermis del feto está cubierta por células epidérmicas. Sin embargo, bajo la superficie lisa de las crestas dérmicas y epidérmicas comienzan a tomar forma (figura 8_6). Hacia el final del quinto mes del embarazo las crestas dérmicas se convierten en rasgos reconocibles de la superficie.
La dermis surge de células mesodérmicas derivadas del dermatoma de las somitas o de otras células mesenquimatosas localizadas justo por debajo del ectodermo. En la cara y en unas partes del cuello, las células dérmicas proceden del ectodermo de la cresta neural craneal (figura 11_10).
Al principio, la futura dermis está representada por células mesenquimatosas adheridas de forma laxa que están muy interconectadas mediante uniones focales apretadas entre sus procesos celulares. Estos precursores dérmicos primarios segregan una matriz intercelular acuosa rica en glicógeno y en ácido hialurónico.
Al comienzo del tercer mes la dermis en desarrollo hace su transición desde la forma embrionaria muy celular hasta un estado que se caracteriza por la diferenciación de las células mesenquimatosas en fibroblastos y la formación de cantidades crecientes de matriz intercelular de tipo fibroso. Los principales tipos de fibras son el colágeno y las fibras elásticas de tipo I y III. La dermis se vuelve muy vascularizada conforme una red capilar incipiente se transforma en grupos de vasos mayores. Poco después de la octava semana, los nervios ayudan a conformar arcos reflejos completos, lo que permite al feto responder a la presión y al contacto.
Interacciones dérmico/ epidérmicas
La transformación del ectodermo simple en una epidermis de múltiples capas depende de continuas interacciones inductivas con la dermis subyacente. Las interacciones dérmico/ epidérmicas también son la base de la formación de una amplia variedad de los derivados epidérmicos y la aparición de variaciones regionales en la estructura de la epidermis.
Por ejemplo, en una etapa precoz del desarrollo la epidermis que cubre las palmas y las plantas se vuelve mucho mas gruesa que cualquier otra parte del cuerpo. Estas regiones no producen vello, mientras que los vellos de cualquier clase, bien sean grueso o muy finos, se forman según patrones regulares en la epidermis de casi todo el resto del cuerpo.
Los experimentos de recombinación de tejidos realizados en diversas especies de vertebrados han demostrado que la dermis subyacente determina el curso del desarrollo de la epidermis y de sus derivados, y que el ectodermo también influye sobre el desarrollo de la dermis. Si los primeros componentes ectodérmicos y mesenquimatoso de la piel de disocian mediante la acción enzimática y se cultivan por separado, el componente ectodérmico sigue siendo ectodermo simple y no llega a diferenciarse en una epidermis de múltiples capas con derivados epidérmicos adecuados. Así mismo, el mesénquima subectodérmico aislado conserva su carácter embrionario sin diferenciarse en dermis.
Si el ectodermo de una parte del cuerpo se combina con epidermis de otra región, el ectodermo se diferencia aún patrón regional característico de la dermis subyacente en vez de el patrón epidérmico de su lugar de origen (figura 8_7). Los experimentos de recombinación entre especies han demostrado que aun en los animales que guardan entre si una relación filogenética distante, el ectodermo y el mesénquima de la piel pueden responder a las señales inductivas de cada uno de ellos.
Al igual que sucede con muchas otras partes del cuerpo, las interacciones inductivas y morfogénesis posterior de la piel y sus derivados están medidas por la producción y la secreción de moléculas de activación o de señal. La morfogénesis regional especifica de la piel y sus derivados parece modelarse gracias a las acciones de factores de transcripción, que probablemente funcionan bajo la influencia aún poco conocida del ácido retinoico, el cual ejerce poderosos efectos sobre la piel.
Derivados epidérmicos
Como resultado de las influencias inductivas ejercidas por la dermis, la epidermis produce una gran variedad de derivados como pelo, uñas, glándulas sudoríparas y sebáceas, glándulas mamarias e incluso el esmalte de los dientes (el desarrollo de los dientes se explica en el capitulo 13).
Los pelos son derivados epidérmicos especializados que surgen como resultado de estímulos inductivos de la dermis. Hay muchos tipos de pelos, que van desde los gruesos de las pestañas y las cejas hasta los casi invisibles del abdomen y espalda. La dermis subyacente determina las diferencias regionales en la morfología y la distribución de los pelos en la epidermis.
La morfogénesis del pelo se identifica por primera vez alrededor de las doce semanas de gestación como proyecciones epidérmicas descendientes situadas a intervalos regulares asociadas con pequeñas condensaciones de células dérmicas llamadas papilas dérmicas (figura 8_8). Bajo la continua influencia de una papila dérmica, la proyección epidérmica descendente continúa en las siguientes semanas y forma un bulbo piloso, En las semanas posteriores el bulbo epidérmico crece sobre la papila dérmica, y empieza a modelarse un folículo piloso precoz. En esta etapa el folículo piloso todavía no sobrepasa a la superficie externa de la epidermis, pero en la porción del folículo que penetrará profundamente en la dermis: 1)presagian la formación de las glándulas sebáceas, que segregan un lubricante cutáneo oleoso (sebo) y 2) son el punto de anclaje para el pequeño músculo piloerector. Este es un músculo liso derivado del mesodermo, que levanta el vello hasta una posición casi vertical cuando también está frío. En muchos animales esto aumenta las propiedades aislantes del pelo.
Se sabe más sobre los mecanismos que regulan la formación de las plumas que acerca de los del pelo. No obstante, el pelo es homólogo de la pluma, de manera que es muy probable que los mecanismos fundamentales que controlan la formación de las plumas también se apliquen al pelo. En primer paso en la formación de un derivado epidérmico, como el pelo o la pluma, es la actuación de factores de crecimiento producidos en el mesénquima –específicamente las proteínas morfogenéticas óseas (BMP) y los factores de crecimiento de los fibroblastos (FCF)- sobre el ectodermo suprayacente, lo que hace que esta capa se engruese para formar una plácoda para cada pelo que va a formarse. Las plácodas de ectodermo engrosado producen entonces las moléculas de señal –(FGF-2, sonic hedgehog y BMP-2) las cuales a su vez hacen que el mesénquima subyacente se condense (papila dérmica). En diferentes sistemas en desarrollo existen genes que contienen homeoboxes, Msx, que son producidos en los momentos y lugares en los cuales la proliferación celular es muy activa. Así ocurre en la formación de los derivados cutáneos; la producción de Msx por parte del ectodermo engrosado se asocia con una intensa proliferación tanto del ectodermo como de la dermis subyacente. Por último, la expresión de varios genes Hox parece tener importancia en la determinación del tipo de derivado epidérmico que se va a formar (por ejemplo, pelo de la ceja en contraste con el vello fino de la mejilla).
Los primeros pelos se ven sobresalir en las cejas poco después de la semana 16; al cabo de un par de semanas, ya cubren el cuero cabelludo. El brote capilar sigue un gradiente cefalocaudal en todo el cuerpo. Durante las últimas fases de la formación del pelo, el bulbo piloso es infiltrado por melanocitos, lo que determina su color. A partir del quinto mes, aproximadamente, las células epidérmicas del tallo del pelo comienzan a queratinizarse y se forman gránulos densos de tricohialina que le dan firmeza.
Los productos de las glándulas sebáceas se acumulan en la superficie de la piel como vérnix caseosa. Estas sustancia sirve de capa protectora para la epidermis, que está continuamente expuesta al liquido amniótico. Los primeros pelos del feto se conocen como lanugo, son de textura muy fina y crecen muy juntos; son muy abundantes durante el séptimo y el octavo mes. Es característico que se desprendan poco antes del parto y sean remplazados por vellos definitivos más gruesos que surgen de folículos recién formados.
En experimentos se ha demostrado que el patrón de los derivados epidérmicos como el pelo se relaciona con patrones generados en la dermis. Otros estudios han comparado los patrones del pelo del cuero cabelludo entre embriones normales y embriones con malformaciones craneales (figura 8_9), y han demostrado una correlación entre remolinos y la dirección del crecimiento del pelo y la tensión en la epidermis en el momento en que se forman los folículos pilosos.
Glándulas mamarias
Como sucede con muchas estructuras glandulares, las glándulas mamarias surgen como proyecciones del epitelio (en este caso ectodérmicas) hacia el mesénquima en respuesta a influencias inductivas procedentes de éste último. La primera evidencia morfológica del desarrollo de la glándula mamaria es la aparición de dos bandas de engrosamientos ectodérmicos llamadas líneas mamarias, que discurren a lo largo de las paredes ventrolaterales del cuerpo de los embriones en ambos sexos hacia la sexta semana (figura 8_10A). El nivel craneocaudal y la extensión hasta donde se desarrolla el tejido mamario a lo largo de líneas mamarias varían de una especie a otra. Si se compara la localización del tejido mamario en los bóvidos (caudal), en los humanos (en la región pectoral) y en los cánidos (a lo largo de la línea mamaria), queda demostrada la amplia variación en la localización y el número de glándulas mamarias. En los seres humanos, pueden encontrarse tejido mamario o pezones supernumerarios en cualquier punto a lo largo de las líneas primitivas mamarias (figura 8_10 B).
Las proyecciones epiteliales de los conductos mamarios (figura 8_11) se asocian con dos tipos de mesodermo: fibroblástico y adiposo. Los datos experimentales demuestran que las interacciones inductivas con los componentes adiposos del tejido conectivo son responsables de la forma característica del sistema de conductos mamarios. Como sucede con muchas estructuras glandulares en desarrollo, el mensaje inductivo parece estar mediado en alto grado por la matriz extracelular del tejido conectivo.
Aunque el mesodermo controla el patrón de ramificación del epitelio ductal, las propiedades funcionales de los conductos mamarios son intrínsecas del componente epitelial, según se demostró en un experimento en ratones, en el cual se combinó ectodermo mamario con mesénquima de la glándula salival. Los conductores mamarios desarrollan el patrón de ramificación característico del epitelio de las glándulas salivales; las células de los conductos mamarios produjeron alfa lactalbúmina, una de las proteínas de la leche.
En relación con su función y como estructura de característica sexual secundaria, las glándulas mamarias tienen una elevada capacidad de respuesta a estímulos hormonales del medio. Esto se ha demostrado en experimentos con ratones. En contraste con la aparición continua de proyecciones descendentes del epitelio ductal que se observan en las hembras, los conductos mamarios de los ratones machos responden a la presencia de testosterona e involucionan con rapidez. Los conductos mamarios femeninos responden de manera similar si se exponen a testosterona. Estudios posteriores han demostrado que el efecto de la testosterona está mediado por el mesénquima mamario y no por acción directa sobre el epitelio ductal. Por el contrario, si se permite que los conductos mamarios de los machos se desarrollen en ausencia de testosterona, adquieren una morfología femenina.
El papel del mesodermo y de los receptores de testosterona quedaba bien ilustrado en los experimentos en ratones con el síndrome de insensibilidad a los andrógenos, una mutación genética. Este síndrome es la contraparte de una afección en los seres humanos se llama síndrome de feminización testicular, en el cual los individuos con genotipo masculino carecen de receptores de testosterona y por tanto desarrollan fenotipo femenino, incluido el típico desarrollo mamario de mujer (figura 8_12 A) puesto que, sin receptores, el tejido no puede responder a la testosterona.
Los experimentos de recombinación in vitro en ratones con insensibilidad a los andrógenos han sido fundamentales para comprender el papel del mesodermo en la mediación de los efectos de la testosterona en el desarrollo de los conductos mamarios (figura 8_12 B). Si el ectodermo mamario de
los mutantes se combina con mesodermo normal en presencia de testosterona, los conductos mamarios regresan; sin embargo, el ectodermo normal combinado con mesodermo del mutante continúa formando conductores mamarios normales a pesar de exponerse a niveles elevados de testosterona. Esto demuestra que el defecto genético en la feminización testicular se expresa en el mesodermo.
El desarrollo posnatal del tejido de la glándula mamaria femenina también es regulado por la acción hormonal. El sencillo sistema de conductos mamarios que se creó en el embrión permanece en este estado hasta que se expone a la acción hormonal al comienzo de la pubertad (figura 8_13 A). Los niveles crecientes de estrógenos circulantes y otros cambios hormonales menos evidentes estimulan la proliferación de los conductos mamarios y el aumento de la almohadilla de tejido adiposo que está situada por debajo de ellos (figura 8_13 B). El otro gran cambio en el ciclo del desarrollo del tejido mamario tiene lugar durante el embarazo, aunque
Pueden detectarse cambios cíclicos menores en cada periodo menstrual. Durante el embarazo, los niveles elevados de progesterona, junto con la prolactina y el lactógeno placentario, estimulan el desarrollo de los alvéolos secretorios en los extremos de las ramificaciones de los conductos (figura 8_13 C). Con el continuo desarrollo de los alvéolos, en las células epiteliales se acumulan gran cantidad de organelos citoplásmicos, como el retículo endoplásmico rugoso y el aparato de Golgi, que están implicados en la síntesis y la secreción de las proteínas.
Durante la lactancia se producen interacciones recíprocas entre la glándula mamaria y el cerebro (éstas se resumen en la figura 8_13 D). Estimuladas por la secreción de la prolactina de la leche (caseína y alfalactalbúmina) y lípidos. En una rápida respuesta al estímulo de la succión, la liberación de oxitocina por la neurohipófisis desencadena la eyección de la leche. La oxitocina causa la contracción de las células mioepiteliales que rodean a los alvéolos. La succión también inhibe la liberación del factor liberador de hormona luteinizante por el hipotálamo, lo que ocasiona la inhibición de la ovulación y provoca una forma natural de anticoncepción.
Cuando se suspende la lactancia, la reducción de la secreción de prolactina y los efectos inhibitorios de la leche no eyectada de los alvéolos mamarios conducen al cese de la producción de leche. Los alvéolos mamarios involucionan y el sistema de conductos de la glándula mamaria recupera si estado previo al embarazo (figura 8_13 E).
El tejido esquelético está presente en casi todas las regiones del cuerpo, y los elementos esqueléticos individuales son bastante diferentes entre si en cuanto a su morfología y su arquitectura. No obstante, a pesar de esta diversidad, hay algunas características embriológicas comunes.
Todo el tejido esquelético se origina en células de morfología mesenquimatosa, pero el origen del mesénquima varía en las diferentes regiones del cuerpo. En el tronco el mesénquima a partir del cual se desarrolla el esqueleto axial segmentario (es decir, la columna vertebral, las costillas y el esternón), se origina de la porción esclerotómica de los somitas mesodérmicos, en tanto que el esqueleto apendicular (los huesos de las extremidades con sus respectivas cinturas) se deriva de la mesénquima del mesodermo de la placa lateral (somato pleura).
El origen del esqueleto de la cabeza es más complejo. Algunos huesos del cráneo (por ejemplo, los que constituyen la bóveda y gran parte de la base craneal) son de origen mesodérmico y gran parte de la base craneal) son de origen mesodérmico, pero los huesos de la cara y algunos que cubren el cerebro surgen del mesénquima derivado de la cresta neural ectodérmica.
Los elementos esqueléticos del cuerpo aparecen primero como moldes cartilaginosos de los huesos que después van a formarse (figura 8_14). En periodos específicos de la embriogénesis, el cartílago es remplazado por hueso mediante el proceso de osificación endocondral. En contraste, los huesos de la cara y el cráneo se forman mediante la osificación directa de las células del mesénquima sin que mediante una etapa cartilaginosa intermedia (formación intramembranosa de hueso). Los detalles microscópicos de la formación intramembranosa y de la endocondral se estudian en los textos de histología y no se repetirán aquí.
Un elemento común a las células mesenquimatosas precursoras de los elementos esqueléticos es su migración o su desplazamiento relativo desde su lugar de origen hasta el área donde se formará el hueso. El desplazamiento puede ser relativamente menor, como en la acumulación de células del esclerotoma del somita que se dispone alrededor de la notacorda para formar el cuerpo (centrum) de una vértebra, o puede implicar migraciones extensas de células de la cresta neural craneal hasta sus destinos para formar los huesos membranosos de la cara.
Para diferenciarse en elementos esqueléticos determinados, las células mesenquimatosas precursoras a menudo tiene que interactuar con elementos de su ambiente inmediato –por lo general, el epitelio con su lámina basal asociada- o con componentes de la matriz extracelular vecina. Los detalles de la interacción varían de una región del cuerpo a otra. Por ejemplo, en la especificación del esqueleto de la extremidad, está implicada una interacción continua entre la cresta apical ectodérmica (capitulo 9) y el mesodermo subyacente de la yema en la extremidad. Una interacción inductiva entre el esclerotoma y la notocorda o el tubo neural inicia la esquelotogénesis de la columna vertebral. En la cabeza, las células presqueléticas de la cresta neural pueden recibir información de niveles que van desde el tubo neural mismo, de lugares situados a lo largo de su trayecto migratorio y hasta de la región de su destino final. Las interacciones inductivas entre regiones del cerebro y el mesénquima suprayacente inducen la formación de los huesos membranosos de la bóveda craneal.
Cualquiera que sea la fuente y el tipo de información ambiental que usan las células precursoras mesenquimatosas, una vez que llegan a su destino casi siempre experimentan una condensación local antes de que se produzcan los primeros signos de su diferenciación. Es característico que la fase de condensación de la formación del hueso se acompañe de aumento de la regulación del N –CAM, una de las moléculas que median la adhesión de las células formadoras de hueso y favorece el establecimiento de la condensación previa al hueso.
Los tejidos esqueléticos se diferencian a partir de las células mesenquimatosas pluripotenciales que tienen la capacidad de diferenciarse en tejido conectivo fibroso, adipositos, células musculares, cartílago y hueso. Se sabe que hay factores de transcripción específicos que dirigen la diferenciación de las células mesenquimatosas tanto en adipocitos (células grasas) como en células musculares (pág. 178). Hace poco tiempo se demostró que el Cbfal (factor 1 de fijación del núcleo) – un factor de trascripción, que es una gen relacionado con el gen pair rule de la Drosophila- , controla la diferenciación de células mesenquimatosas en osteoblastos (células formadoras de hueso), pero la formación de cartílago es independiente de este gen. Los ratones portadores de mutantes homocigotos Cbfa 1 forman modelos cartilaginosos de los huesos endocondrales, pero estos huesos jamás se osifican; en cambio, experimentan cierto grado de calcificación pero permanecen como cartílago calcificado. Los huesos intramembranosos de la cabeza y la cara no se forman en estos ratones. Las proteínas morfogenéticas óseas (BMP), en especial las BMP-4 y BMP-7, están implicadas en la formación de huesos embrionarios. Las BMP actúan en varias etapas de la formación del hueso, pero algunos datos sugieren que su función inicial es estimular la expresión del Cbfal. La morfogénesis individual de los elementos esqueléticos suele estar bajo grupos complejos de control que van desde patrones de expresión de genes Hox en la etapa de somita, hasta interacciones inductivas entre los epitelios y las células mesenquimatosas precursoras y hasta influencias mecánicas que pueden actuar en cualquier momento desde las primeras etapas de la morfogénesis hasta las más avanzadas de la vida posnatal.
Columna vertebral y costillas
Las primeras etapas del desarrollo del esqueleto axial se describieron en el capitulo 5. sin embargo, la formación de esqueleto axial es mas compleja que la simple subdivisión del mesodermo paraxial en somitas y el desplazamiento en dirección medial de las células del esclerotoma para formar los primordios vertebrales. Cada vértebra tiene una morfología compleja que le es única y es especificada por controles que actúan a varios niveles y durante varios períodos del desarrollo.
De acuerdo con la división tradicional del desarrollo vertebral (figura 5-19), los esclerotomas se dividen en mitades craneales y caudales, y la mitad caudal más densa celularmente de un esclerotoma se une a la mitad craneal mas laxa celularmente de las siguiente para formar el cuerpo de la vértebra. Investigaciones morfológicas recientes sugieren que el desarrollo vertebral es más complejo que el propuesto por este modelo, según se indica en el esquema ilustrado en la figura 8_15.
La columna vertebral se divide en varias regiones (figura 8_14), a saber: 1) una región occipital, que se incorpora a la estructura ósea de la base del cráneo, 2) una región cervical, que incluye el atlas y el axis, vértebras muy especializadas que unen la columna vertebral con el cráneo; 3) la región torácica, en la cual se originan las costillas verdaderas, 4) la región lumbar, 5) la región sacra, en la cual las vértebras se fusionan en un solo hueso sacro,
y 6) la región caudal, que representa la cola de la mayoría de los mamíferos y el rudimentario cóccix en los seres humanos. Una vértebra típica surge de la fusión de varios primordios cartilaginosos, el cuerpo, que se deriva de las porciones ventromediales del esclerotoma de los pares de somitas (figura 5_18), rodea a la notocorda y sirve de suelo óseo para la médula espinal (figura 8_16). Los arcos neurales –que se originan en las células esclerotomales laterales- se fusionan a ambos lados con el cuerpo y, junto con otros arcos neurales, forman el techo protector de la médula espinal. El cierre incompleto del techo óseo da lugar a una anomalía llamada espina bífida oculta (figura 10_40). La apófisis costal forma las costillas verdaderas a la altura de las vértebras torácicas. En otros niveles de la columna vertebral los procesos costales se incorporan a las vértebras propiamente dichas.
El desarrollo de cada vértebra comienza con la inducción mediada por el sonic hedgehog (shh) realizada por la notocorda sobre el somita inicial para formar el esclerotoma. Bajo la influencia continua del shh, la porción medioventral del somita formará el cuerpo de la vértebra. La formación de si parte dorsal (el arco neural) la determina un grupo diferente de control del desarrollo. La inducción inicial de la placa del tubo neural redunda en la expresión del Pax- 9 y del gen Msx- 2 que contiene un homeobox, el cual guiará a las células del esclerotoma lateral para que formen el arco neural.
Las características regionales fundamentales de las vértebras se especifican de acuerdo con las acciones de combinaciones puntuales de genes que contienen homeobox (figura 8_17). La expresión de los genes Hox comienza con la aparición del mesodermo presomítico y para la mayoría de los genes persiste hasta que empieza la condrificación en los primordios vertebrales. La formación del patrón segmentado normal a lo largo del eje craneocaudal de la columna vertebral puede asegurarse por el hecho de que la mayoría de las vértebras son especificadas por una combinación única de genes Hox. Por ejemplo, en el ratón el atlas (C- 1) se caracteriza por la expresión de Hoxa- 3, Hoxb- 1 y Hoxd- 4. El axis (C-2) es especificado por estos cuatro genes más Hoxa- 4 y Hoxb- 4. el ácido retinoico (vitamina A) puede causar el desplazamiento del nivel craneal o caudal de la organización segmentaria global de las vértebras si se aplica en períodos determinados del desarrollo. Por ejemplo, si se administra en las primera etapas, el ácido retinoico produce un desplazamiento del nivel craneal (la última vértebra cervical se transforma en la primera torácica); si se administra más adelante causa un desplazamiento del nivel caudal (las vértebras torácicas se extienden hasta los niveles de las dos primeras vértebras lumbares). Estos desplazamientos de nivel se llaman transformaciones homeóticas y son características de la amplia familia de los mutantes homéoticos descritos en el capítulo 5. Cerca del 5% de los seres humanos presentan variaciones menores en el número o las proporciones de las vértebras. El síndrome de Klippel- Fiel, en ocasiones llamado brevicollis, se caracteriza por un cuello corto con una reducción del número de vértebras cervicales, una línea baja de implantación del pelo y otras anomalías. Es probable que estas variaciones de la organización vertebral guarden relación con desequilibrios en la expresión de los productos clave de los genes que contienen homeobox.
El axis y el atlas tienen una forma morfología poco usual y un origen característico (figura 8_18). El cuerpo del atlas es deficiente, pero el área del cuerpo es atravesado por la apófisis odontoides del axis. Ésta ultima consta de tres cuerpos fusionados que se presume son equivalentes: 1) la mitad de un segmento del cuerpo de un hueso transicional (el pro atlas) que no se encuentra en los seres humanos, 2) el cuerpo que debería haber pertenecido al atlas, y 3) el centro normal del axis. Esta distribución permite una mayor rotación de la cabeza sobre la columna cervical. Cuando se introdujo un transgen Hoxa- 7 de expresión ubicua en la línea germinal del ratón, la parte craneal de la columna vertebral se posteriorizó. La base del hueso occipital se transformó en una vértebra occipital (el proatlas) y el atlas se combino con su cuerpo, de lo cual resultó un axis desprovisto de apófisis odontoides.
Las costillas se originan en zonas de células mesenquimatosas condensadas lateralmente el centro (figura 8_16). Cuando comienza la osificación del las vértebras, las costillas se separan de ellas. Las costillas accesorias son frecuentes –en especial en los niveles lumbar superior y cervical inferior-, pero las cifras de su incidencia varían ampliamente de una serie a otra. Un cálculo realista sería una incidencia algo menor del 1%. Éstas y otras anomalías comunes (costillas bifurcadas o fusionadas) suelen ser asintomáticas y por lo general sólo se detectan en los exámenes de rayos X. Es probable que sean el resultado de la expresión equivocada de genes Hox específicos.
El esternón surge como un par de bandas cartilaginosas que convergen hacia la línea media ventral conforme la pared ventral del cuerpo se va formando (figura 8_19). Después de que las bandas esternales primordiales convergen, demuestran su verdadera naturaleza segmentaria al subdividirse de forma secundaria en elementos craneocaudales. Esta segmentación secundaria sigue en curso morfológico y molecular que inicialmente se asemeja bastante a la formación de las articulaciones sinoviales (pág. 202). Muchas de estas divisiones terminan fusionándose conforme se osifican para formar el cuerpo impar del esternón. Varias de las anomalías más frecuentes del esternón ( como la apófisis xifoides dividida) se entienden como facilidad a partir de su desarrollo embriológico. Las malformaciones de la apófisis xifoides se ven en ratones mutantes para genes Hoxc- 4 y Hoxa- 5, en tanto que los ratones mutantes para el Hoxb- 4 tienen el esternón dividido.
En ocasiones se ha considerado la clavícula como parte del esqueleto apendicular porque conecta el esternón con el húmero. La clavícula es uno de los primeros huesos del cuerpo en osificarse; hacia la octava semana el proceso está bastante avanzado. Estudios recientes en ratones heterocigotos para el gen Cbfa 1 han revelado no sólo la naturaleza de la clavícula, sino además un síndrome no muy bien comprendido en los seres humanos. Estos heterocigotos muestran hipoplasia de la clavícula, retraso de la osificación de los huesos membranosos (como el cráneo) y fontanelas craneales anterior y posterior abiertas. La displasia cleidocraneal en los seres humanos muestra todos estos componentes, así como dientes supernumerarios. Si no hay clavículas, los individuos afectados pueden juntar sus hombros en la línea media anterior. Los hallazgos en estos mutantes sugieren que la clavícula es un hueso de membrana y que podría ser una clase única de hueso, ya que no puede considerarse ni como hueso axial verdadero ni como un hueso apendicular en el sentido usual de la palabra.
Otro desarrollo tardío es la desaparición de la notocorda de los cuerpos vertebrales. Entre las vértebras, la notocorda se dispone en el primordio mesenquimatoso condensado de los discos intervertebrales. En el adulto la notocorda persiste como el núcleo pulposo que constituye el núcleo blando del disco. El disco intervertebral consta de capas de fibrocartílago que se diferencian de las células mesodérmicas derivadas del esclerotoma. El Pax- 1 s expresa de forma continua durante el desarrollo de los discos intervertebrales. En un ratón mutante, el ondulated, la expresión del Pax- 1 es deficiente, y como consecuencia los cuerpos vertebrales se fusionaron. Al parecer la expresión del Pax- 1 y la formación posterior de los discos intervertebrales son mecanismos importantes para mantener el carácter segmentario individual de la columna vertebral.
El extremo caudal del esqueleto axial está representado por un apéndice definido parecido a una cola durante la mayor parte del segundo mes (figura 8_20 A). Durante el tercero, la cola generalmente se corta, en gran parte por muerte celular y crecimiento diferencial, para persistir como el cóccix; sin embargo, en raras ocasiones puede persistir en los recién nacidos como una cola bien desarrollada.
El cráneo es una estructura compleja que consta de dos subdivisiones principales: el neurocráneo, que rodea el cerebro, y el viscerocráneo, que rodea a la cavidad oral, la faringe y las vías aéreas superiores. Cada una de estas subdivisiones a su vez costa de dos componentes: uno en el cual los huesos individuales están representados al comiendo por modelos cartilaginosos que más adelante son remplazados por hueso mediante la osificación endocondral, y otro en el cual el hueso surge por osificación del mesénquima.
La base filogenética y ontogenética del cráneo está representada por el condrocráneo, que forma la base cartilaginosa del neurocráneo (figura 8_21 A). El patrón fundamental del condrocráneo se ha preservado de forma notable durante la filogenia. Al comienzo está representado por varios juegos de cartílagos pares. Un grupo (los cartílagos paracordales, hipofisiarios, y las trabéculas del cráneo) guarda una estrecha relación con las estructuras de la línea media. En dirección caudal a los cartílagos paracordales se encuentran los cuatro esclerotomas occipitales. Junto con los cartílagos paracordales, los esclerotomas occipitales (que son homólogos de los precursores de las vértebras) se fusionan para dar lugar a la base del hueso occipital. En situación más lateral, el condrocráneo está representado por pares de cartílagos que están asociados con los primordios epiteliales de los órganos de los sentidos (órgano olfatico, ojos y órgano auditivo).
Los elementos primordiales individuales del condrocráneo experimentan varios patrones de crecimiento y fusión para formar los huesos estructuralmente complejos de la base del cráneo (los huesos occipital, esfenoides y temporal, así como gran parte del soporte óseo de la cavidad nasal) (figura 8_ 21 B). Además. Algunos de estos huesos (por ejemplo, el occipital y el temporal) incorporan elementos membranosos durante el desarrollo, de manera que en su forma final son estructuras compuestas (figura 8_ 21 D). Otros componentes del neurocráneo, como los huesos parietales y el frontal, son exclusivamente membranosos (cuadro 8_ 2).
Prácticamente todos los huesos del neurocráneo son el producto final de una influencia inductiva de una estructura epitelial sobre el mesénquima vecino. Estas interacciones suelen estar mediadas por factores de crecimiento y la matriz extracelular. Los estudios inmunocitoquímicos han demostrado la aparición transitoria de colágeno de tipo II (el principal tipo de colágeno del cartílago) en los lugares y en los momentos durante los cuales se presentan las interacciones que conducen a la formación del condrocráneo. Además del colágeno de tipo II, se acumula un proteoglicano específico de cartílago en las áreas de inducción de los elementos condrocraneales. Hay pruebas de inducción de que elementos epiteliales de la cabeza sólo inducen el esqueleto, sino que también controlan su morfogénesis. Esto contrasta con el control morfogenético del esqueleto apendicular, que esta determinado por el mesodermo y no por el ectodermo de la yema de la extremidad.
Los elementos del neurocráneo membranoso (los huesos apriétales y frontales y la parte interparietal del occipital)surgen como cúmulos planos, similares a placas, de espículas óseas (trabéculas) del mesénquima que ha sido inducido por partes específicas del cerebro en desarrollo. Estos huesos permanecen como estructuras
separadas durante el desarrollo fetal, e incluso en el nacimiento, están separados por suturas de tejido conectivo. Las intersecciones de las suturas donde se encuentran más de dos huesos están ocupadas por áreas más amplias de tejido conectivo llamadas fontanelas; las más prominentes son la fontanela anterior, localizada en la intersección de los dos frontales y los dos apriétales, y la posterior localizada en la intersección de los dos parietales y el occipital (figura 8_ 22).
A semejanza del neurocráneo, el viscerocráneo consta de dos divisiones: un viscerocráneo cartilaginoso y un viscerocráneo membranoso. En contraste con el neurocráneo, los huesos del viscerocráneo se originan en su mayor parte del mesénquima derivado de la cresta neural. Desde el punto de vista filogenético, el viscerocráneo está relacionado con el esqueleto de los arcos branquiales (arcos de las branquias). Cada arco branquial (que recibe también el nombre de arco faringeo en los seres humanos) está formado por un arco cartilaginoso que da lugar a varios de los elementos esqueléticos definitivos característicos de cada arco (cuadro 8_2). Los detalles de la organización y los derivados no craneales de los cartílagos de los arcos faríngeos se explican en la figura 13_22.
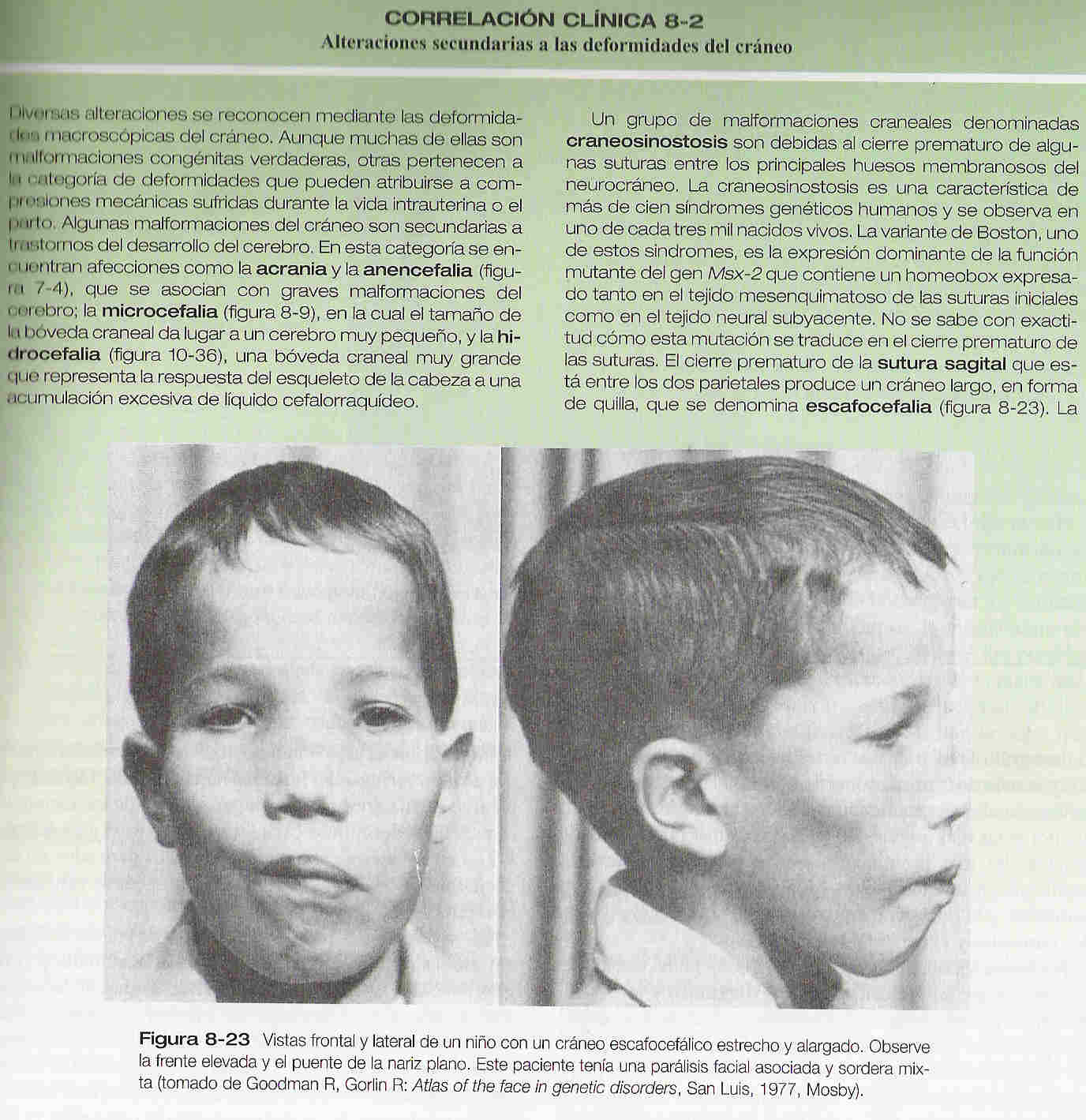
El viscerocráneo membranoso consta de una serie de huesos asociados con el maxilar superior, la mandíbula y la región del oído (figura 8_ 21 D). Estos huesos surgen del primer arco cartilaginoso (cartílago de Meckel) y asumen, entre otras, algunas de las funciones que originalmente desempeñaba el cartílago de Meckel, como la transmisión del sonido en el oído medio. La correlación clínica 8_ 2 explica las afectaciones resultantes de las deformidades del cráneo.
El esqueleto apendicular costa de los huesos y las cinturas de las extremidades. Existen diferencias fundamentales en la organización y control del desarrollo entre el esqueleto axial y el apendicular. El primero forma una cubierta protectora alrededor de los tejidos blandos internos (como, el cerebro, la médula espinal y la faringe) y el mesénquima que forma los huesos en inducido por los órganos a los cuales los huesos rodean. Por el contrario, los huesos del esqueleto apendicular forman un núcleo central para soporte de los miembros. Si bien la interacción con el epitelio –(la cresta apical ectodérmica de la yema de la extremidad - (capítulo 9) es necesaria para la formación de los elementos esqueléticos de la extremidad, el control morfogenético de ésta depende del mesodermo, y el epitelio sólo desempeña una función de estimulación.
Todos los elementos del esqueleto apendicular comienzan como moldes cartilaginosos que se convierten en hueso verdadero gracias a la osificación endocondrial durante la embriogénesis (los detalles de la formación del esqueleto apendicular se exponen en el capítulo 9).
Durante el desarrollo embrionario se forman tres tipos de musculatura: esquelética, cardiaca y lisa. Prácticamente toda la musculatura se deriva del mesodermo paraxial, específicamente de los somitómeros o de los somitas (figura 5_16). El mesodermo esplácnico da origen a la musculatura del corazón (músculo cardiaco) y a la musculatura lisa de los tractos intestinal y respiratorio (tabla 8_1). Otros músculos lisos, como los de los vasos sanguíneos y los piloerectores, se derivan del mesodermo local por diferenciación in situ.
El desarrollo muscular puede analizarse a diferentes niveles, que van desde la determinación y la diferenciación de las células musculares individuales, hasta la histogénesis del tejido muscular y por ultimo hasta la formación (morfogénesis) de los músculos completos. El músculo esquelético sirve como ejemplo para entender cómo se produce el desarrollo y cómo se controla en los diferentes niveles de organización.
Hay pruebas fehacientes de que ciertas células del epiblasto están determinadas para convertirse en células miogénicas incluso antes de que los somitas hayan terminado de formarse, pero conviene comenzar el análisis de aparición de las células precursoras del músculo en los somitas. Durante muchas décadas el origen de la musculatura esquelética era un tema controvertido; dos de los posibles orígenes eran los somitas y el mesodermo en la placa lateral. La controversia se resolvió cuando se hicieron estudios con marcadores celulares (cuadro 8_3), que hoy se sabe que prácticamente todo el músculo esquelético se origina en los somitas o los somitómeros. Los primeros pasos en la determinación de las células miogénicas en los somitas se resumen en la figura 5_18.
La fibra del músculo esquelético maduro es una célula compleja multinucleada que está especializada para contraerse. Los precursores de la mayor parte de los linajes musculares (células miogénicas) han sido seguidos hasta el miotoma del somita (figura 5_17). Aunque estas células se parecen a las células mesenquimatosas que pueden dar lugar a muchos otros tipos celulares en el embrión, han sufrido un suceso de restricción que las compromete con la línea formadora de músculo. Las células miogénicas pasan por varias divisiones mitóticas adicionales antes de llegar a la división mitótica terminal y convertirse en mioblastos posmitóticos.
Las células miogénicas en la proliferación mantienen el ciclo celular por medio de la acción de factores de crecimiento como el factor de crecimiento fibroblastos (FGF) y el factor beta de transformación del crecimiento. En el miotoma del somita, los factores reguladores miogénicos (véase la siguiente sección) se expresan en las regiones formadoras de músculo. El MyoD, uno de estos factores, saca las células mesenquimatosas del ciclo celular porque impide su entrada a la fase S y además estimula la diferenciación porque activa unos genes específicos del músculo. Otros factores de crecimiento, como el factor de crecimiento similar a la insulina, también participan en la promoción de la diferenciación muscular.
Los mioblastos posmitóticos comienzan a transcribir sus RNA mensajeros para las principales proteínas contráctiles, la actina y la miosina, pero el proceso fundamental en el ciclo vital del mioblasto posmitótico es fusión con otras células similares en un miotubo multinucleado (figura 8_26). La fusión de los mioblastos es un proceso preciso que implica su alineación y adhesión mediante mecanismos de reconocimiento celular mediados por Ca++ y la fusión final de sus membranas plasmáticas.
Los miotubos tienen una intensa participación en la síntesis del RNA y las proteínas. Además de formar actina y miosina, sintetizan varias otras proteínas, incluidas las proteínas reguladoras de la contracción muscular troponina y tropomiosina. Estas proteínas se ensamblan en miofibrillas, que son agregados de unidades contráctiles funcionales organizadas de forma precisa llamados sarcómeros. Conforme los miotubos forman miofibrillas, sus núcleos, que habían estado organizados en cadenas centrales regulares, migran hacia la periferia del miotubo. En esta etapa se considera que el miotubo se ha diferenciado en una fibra muscular, etapa final de la diferenciación de la célula muscular esquelética.
No obstante, el desarrollo de una célula muscular no está completo con la migración periférica de los núcleos del miotubo. Aunque los núcleos (mionúcleos) de la fibra muscular multinucleada ya no pueden proliferar, ésta tiene que seguir creciendo en proporción con el rápido crecimiento del feto y después del bebé.
El crecimiento de la fibra muscular se logra gracias a una población de células miogénicas llamadas células satélites, que adoptan posiciones entre la fibra muscular y la lámina basal en la cual está encerrada cada fibra muscular (figura 8_26). Por un mecanismo de control que aún no ha sido determinado, las células satélites se dividen lentamente durante el crecimiento del individuo. Algunas de estas células hijas se fusionan con la fibra muscular de manera que ésta última mantiene un número adecuado de núcleos para dirigir la continua síntesis de las proteínas contráctiles que requiere. Cuando las fibras musculares se dañan, las células satélites proliferan y se fusionan para regenerar las fibras musculares.
Un músculo típico no está compuesto por fibras musculares homogéneas. Por lo general se distinguen varios tipos de fibras musculares según sus propiedades contráctiles y su morfología, así como por su contenido de diferentes isoformas de las proteínas contráctiles. Para los propósitos de este texto, consideramos que las fibras musculares son de dos tipos: rápidas o lentas.
Factores de transcripción muscular. La miogénesis se inicia con un proceso de restricción que canaliza una población de células de mesénquima hacia un linaje de células miogénicas. La base molecular de esta restricción se debe a la acción de miembros de los grupos de factores reguladores miogénicos que, al actuar como reguladores genéticos maestros, activan genes específicos del músculo en las células mesenquimatosas premusculares.
La primera familia de factores reguladores miogénicos que se descubrió es un grupo de cuatro factores de transcripción básicos hélice- asa- hélice, en ocasiones también llamada familia MyoD (figura 8_ 27). El factor intensificador 2, otro factor regulador descubierto hace poco tiempo, parece trabajar corriente abajo del curso de activación de la familia MyoD, pero todos estos factores reguladores miogénicos pueden convertir células no musculares (como fibroblastos, adipocitos, condorcitos y células pigmentarias retinianas) en células que expresan el rango total de las proteínas musculares.
Como sucede con muchas proteínas hélix- loop- hélix, las proteínas reguladoras miogénicas de la familia MyoD forman dímeros y se fijan a una secuencia específica del ácido desoxirribonucleico (DNA) llamada E box, en la región estimuladora de los genes específicos del músculo. La especificidad miogénica de estas proteínas está codificada en la región básica (figura 8_ 27).
Las actividades reguladoras de la MyoD y otros miembros de esa familia están controladas a su vez por otras proteínas reguladoras que pueden modificar sus actividades (figura 8_ 28). Por ejemplo, muchas células contienen un activador de la transcripción llamado E12. Cuando una molécula de E12 forma un heterodímero una molécula de MyoD, el complejo se fija de forma más estrecha a la región de intensificación muscular del DNA que el dímero puro del MyoD; así se aumenta la eficiencia de la transcripción de los genes musculares. Por otra parte, un inhibidor de la transformación llamado Id (inhibidor de la unión del DNA) puede formar un heterodímero con una molécula de MyoD. El Id contiene una región hélice- asa- hélice pero no tienen región básica, que es la parte de la molécula que se une al DNA. La molécula Id tiene una mayor afinidad de unión para una molécula MyoD que otra molécula MyoD y puede entonces desplazar una de las unidades del dímero MyoD; el resultado de esta acción es la presencia de más heterodímeros Id- MyoD, los cuales se unen muy poco al DNA y a menudo no pueden activar los genes específicos de los músculos.
Durante el desarrollo muscular, los factores reguladores miogénicos de la familia MyoD se expresan en una secuencia regular (figura 8_ 29). En los ratones, los sucesos que llevan a la formación del músculo comienzan en el somita; allí, tanto el Pax- 3 como el myf- 5, trabajando por vías aparentemente distintas, activan la MyoD, lo que hace que ciertas células del dermomiotoma se dediquen a formar músculo. Con niveles elevados de MyoD, las células mononucleares se retiran del ciclo mitótico y comienzan a fusionarse en miotubos. En esta etapa se expresa la miogenina. Por último, en los miotubos en maduración se expresa el MRF- 4. Es interesante observar que los ratones KO (bloqueador), la ausencia de un solo factor regulador miogénico (por ejemplo, el myf- 5 o el MyoD) no impide que por sí misma la formación del músculo esquelético (aunque pueden observarse otros defectos menores), pero cuando el myf- 5 y el MyoD se eliminan al mismo tiempo, no se forma músculo. Otro ratón doble KO para el Pax- 3 y el myf- 5 produce ratones desprovistos por completo de músculo en el tronco y las extremidades, pero con la musculatura de la cabeza intacta. Estos demuestra que en las primeras etapas de la determinación las células formadoras de músculo de la cabeza y del tronco siguen diferentes vías reguladoras.
Puesto que cada regulador, activador e inhibidor es en sí mismo una proteína, su formación está sujeta a controles positivos y negativos similares. Los complejos ejemplos de la regulación de los primeros pasos de la miogénesis dan alguna idea sobre los múltiples niveles del control de la expresión genética y de las etapas de citodiferenciación en los mamíferos. Si bien los aspectos moleculares de la miogénesis se entienden mejor que las etapas que subyacen a la diferenciación de la mayoría de tipos celulares, resulta poco realista pensar en la diferenciación de casi todos los tipos de células no opere un grupo similar de mecanismos reguladores entrelazados.
El músculo como tejido no sólo consta de fibras musculares sino también de tejido conectivo, vasos sanguíneos y nervios. Ni siquiera las fibras musculares mismas son homogéneas, sino que pueden separarse en varios tipos diferentes desde los puntos de vista funcional y bioquímico.
Cuando los músculos empiezan a formarse, los mioblastos están entremezclados con el mesénquima del futuro tejido conectivo. El papel de este tejido conectivo en la morfogénesis del músculo se explica en la próxima sección. Los brotes de los capilares crecen en el músculo en formación para nutrirlo y las fibras nerviosas motrices entran poco después de que los primeros mioblastos comienzan a formar miotubos.
En una época se pensaba que todos los mioblastos eran idénticos en esencia y que sus características diferenciales (por ejemplo, ser rápidas o lentas) les eran impuestas por su inervación motriz. Sin embargo, investigaciones recientes han demostrado que en aves y en varias especies de mamíferos hay distintas poblaciones de células musculares rápidas y lentas ya desde la etapa del mioblasto, mucho antes de que las fibras nerviosas lleguen siquiera al músculo en desarrollo.
No sólo existen mioblastos rápidos y lentos, sino que también hay isoformas celulares incipientes y tardías en los mioblastos, con requerimientos diferentes de los factores séricos e interacciones nerviosas en su diferenciación. Cuando los mioblastos primarios se fusionan para formar miotubos, dan lugar a los miotubos primarios, que constituyen la base del músculo embrionario. La diferenciación de los miotubos primarios ocurre antes de que los axones nerviosos hayan entrado en el músculo recién formado. A continuación, a lo largo de los miotubos primarios se forman miotubos secundarios más pequeños que surgen de los mioblastos más tardíos (figura 8_ 30). Cuando esto ocurre, los primeros axones motores ya están presentes en los músculos, y hay pruebas de que los nervios son necesarios para la formación de los miotubos secundarios. Una fibra muscular primaria con sus fibras musculares secundarias asociadas están inicialmente rodeadas por una lámina basal común y acopladas desde el punto de vista eléctrico. Estas fibras musculares sintetizan de forma activa una amplia variedad de proteínas contráctiles.
Pronto en su historia vital, las fibras musculares embrionarias son inervadas por neuronas motrices. Aunque durante mucho tiempo se creyó que las motoneuronas rápidas y las lentas imponen sus propias características funcionales
a las fibras musculares en desarrollo, hoy parece que aquéllas pueden seleccionar fibras musculares de un tipo compatible gracias a información que llevan en sus superficies celulares. Al comienzo, el nervio motor puede terminar tanto en las fibras musculares rápidas como en lentas, pero al final, las conexiones inapropiadas se rompen de manera que las fibras nerviosas rápidas solo inervan fibras musculares rápidas y otro tanto sucede con las lentas. Si los datos de las investigaciones se confirman, se verá que ante todo las fibras nerviosas ayudan a las fibras musculares a mantener un estado de diferenciación y no a determinar las diferentes cualitativas entre fibras musculares rápidas y lentas.
Los fenotipos de las fibras musculares dependen de la naturaleza de las proteínas especificas que componen un aparato contráctil. Hay diferencias cualitativas entre las fibras musculares rápidas y las lentas en muchas proteínas contráctiles, y dentro de cada tipo de fibra muscular hay una sucesión de isoformas de las principales proteínas durante el desarrollo embrionario (las transiciones de las isoformas de la miosina en una fibra muscular en desarrollo se usan como ejemplo).
La molécula de la miosina es compleja, y consta de dos cadenas pesadas y de una serie de cuatro cadenas ligeras (figura 8_ 31), las fibras rápidas maduras tienen una cadena ligera LC1, dos LC2 y una LC3; la miosina de las fibras lentas contiene dos subunidades LC1 y dos LC2 de cadenas ligeras. Además, hay formas rápidas y lentas (MHCf y MHCs) de las subunidades de las cadenas pesadas de la miosina. Las moléculas de miosina poseen actividad de adenosintrifosfatosa y las diferencias en esta actividad explican en parte distintas velocidades de contracción entre las fibras musculares rápidas y las lentas.
Las moléculas de la miosina pasa por una sucesión de isoformas de transición durante el desarrollo. Desde el período fetal hasta la madurez tres isoformas de la cadena pesada de la miosina (embrionaria [MCHemb], neonatal [MHCneo] y adulta rápida [MHCf] se forman en la fibra muscular rápida en desarrollo (los cambios del desarrollo de las subunidades de las cadenas pesadas y ligeras se resumen en la figura 8_31). Otras proteínas contráctiles de las fibras musculares (por ejemplo, la actina y la troponina) pasan por isoformas transicionales similares. Cuando el músculo adulto sufre una lesión, las fibras musculares en regeneración pasan por grupos de isoformas transicionales celulares y moleculares que imitan de forma similar a las que se produjeron durante la ontogénesis normal.
El fenotipo de las fibras musculares no se fija de forma irreversible. Las fibras musculares posnatales poseen aun un alto grado de plasticidad; responden al ejercicio hipertrofiándose o haciéndose más resistentes a la fatiga; o se adaptan a la inactividad o a la denervación atrofiándose. Todos estos cambios se acompañan de varias alteraciones en la expresión de los genes. Muchos otros tipos de células también pueden modificar sus fenotipos en respuesta a los cambios en el ambiente, pero los cambios moleculares no siempre son tan evidentes como los que se ven en las fibras musculares.
En un nivel de organización más alto, el desarrollo muscular implica la formación de músculos identificables desde el punto de vista anatómico. La forma global de un músculo está determinada en principio por su marco de tejido conectivo más que por los mioblastos mismos. Los experimentos han demostrado que las células miogénicas del somita son intercambiables. Por ejemplo, las células miogénicas de los somitas que normalmente formarían músculos de tronco pueden participar en la formación de músculos normales de la pierna. En contraste, las células de componente de tejido conectivo de los músculos parecen formar parte del plano morfogenético.
Músculos del tronco y las extremidades. Los experimentos de injertos de codorniz / pollo han demostrado con claridad que los principales grupos de músculos esqueléticos del tronco y las extremidades se originan en los precursores miogénicos localizados en los somitas. En el tórax los músculos intrínsecos de la espalda se derivan de células de los miotomas, mientras que los músculos ventrolaterales (por ejemplo, los intercostales) surgen de yemas ventrales de los somitas en la fase epitelial (figura 8_32). En las regiones de las extremidades, las células miogénicas migran desde el epitelio ventrolateral del dermomiotoma en las primeras etapas del desarrollo.
Las células miogénicas que se originan en las regiones de los somitas craneales migran hacia la lengua en formación. En los niveles lumbares, los precursores de los músculos abdominales también salen del epitelio ventrolateral de las yemas somíticas. Es muy probable que el síndrome del vientre en ciruela, que se caracteriza por la ausencia de musculatura abdominal (figura 8_33), tenga como causa una deficiencia molecular en esta población de células miogénicas.
Algunos experimentos recientes han demostrado comportamientos celulares distintos entre áreas de los miotomas adyacentes a las regiones de las extremidades y las regiones diferentes a las de las extremidades. En los segmentos torácicos, las células del dermatoma rodean a los bordes laterales del miotoma; a continuación hay un aumento en el número de miotubos formados en el miotoma y los primordios musculares penetran en la pared del cuerpo. Por el contrario, el nivel de las yemas de las extremidades, las células del dermatoma mueren antes de rodear a los miotubos primarios que se forman en el miotoma, ni el numero de estos miotubos aumenta de forma importante ni sale de los miotomas para formar primordios musculares separados.
Varios experimentos sugieren que las influencias de la región corporal que rodea a los somitas intervienen en los primeros pasos de la liberación y en la morfogénesis de las células miogénicas precursoras en los somitas. Si se extirpa la yema de una extremidad de un embrión de pollo, las células del dermatoma no mueren; por el contrario rodena a los miotomas y permiten el aumento del número de miotubos que se forman en los miotomas. En cambio, cuando se injerta una yema de una extremidad en el nivel del tórax, las células del dermatoma mueren y los primordios del músculo subyacente nunca maduran. En otro análisis experimental, se demuestra que si se injertan somitas o trozos del mesodermo paraxial rotados de posición con su borde interno orientado lateralmente, las células miogénicas migran desde el nuevo borde lateral, lo que de nuevo indica que las actividades de las células del interior del somita responden a influencias ambientales locales.
Después de originarse en los somitas, los primordios de los músculos del tronco y el abdomen se organizan en grupos y capas bien definidos, como se ilustra en la figura 8_34 ( la morfogénesis de los músculos de las extremidades se examina en capítulo 9). Los resultados de varios experimentos han demostrado diferencias fundamentales en las propiedades celulares de los precursores celulares de los músculos de las extremidades y de los músculos axiales ( tabla 8_ 2)
Músculos de la cabeza y de la región cervical. Los músculos esqueléticos de la cabeza y cuello son de origen mesodérmico. Los experimentos de injertos de codorniz en pollo han demostrado que el mesodermo paraxial, específicamente los somitómeros, constituye la fuente principal de la musculatura craneal, aunque todavía no se sabe a ciencia cierta el origen de los músculos extrínsecos del ojo. Hay pruebas de que algunas de la células que los originan proceden lo la lámina procordal del embrión joven; también hay evidencias crecientes que indican que, en algunos aspectos, la miogénesis de la cabeza muestra importantes diferencias contra la del tronco (los diferentes controles en el nivel de la determinación miogénica entre la cabeza y el tronco ya han sido expuestos). Así mismo, varios músculos craneofaciales tienen propiedades fenotípicas diferentes de los músculos del tronco (por ejemplo, las isoformas de la miosina y posiblemente también elementos del control neuromuscular del fenotipo).
Los músculos de la cabeza y el cuello, al igual que los de la espalda , se originan por la migración de las células miogénicas que se alejan del mesodermo paraxial discurren por el mesénquima (ya sea derivado de la cresta neural o del mesodermo) en su trayecto hacia su destino final. Igual que en el tronco, la morfogénesis de los músculos de la región craneal parece estar determinada por la información inherente al tejido conectivo que constituye la vaina de los músculos. No hay especificidad para los primeros niveles en las células miogénicas paraxiales, según se ha comprobado injertando somitas o somitómeros de un nivel en dirección craneocaudal en otro nivel. En estos casos las células miogénicas que derivan de la estructura del injerto forman músculos normales para la región hacia la cual migran y no aquéllos correspondientes con el nivel de origen de los somitas injertados.
A pesar de que las pruebas apuntan hacia la intercambiabilidad de los somitas embrionarios en la formación del músculo, otras informaciones experimentales muestran la existencia de propiedades posicionales bien definidas de los músculos esqueléticos (por ejemplo, su capacidad de establecer conexiones con nervios de diferentes niveles axiales y ciertos aspectos de la expresión genética). Estos hallazgos indican una fuente impronta sobre las fibras musculares según un marcado gradiente rostrocaudal. Se desconoce en qué momento esta forma de especificidad axial de impone a las fibras musculares en desarrollo.
Algunos músculos de la cabeza, en particular de los de la lengua, surgen de los somitas occipitales de igual manera que los músculos del tronco y experimentan extensas migraciones hacia la cabeza en proceso de crecimiento. Su nivel de origen más caudal se evidencia por que son inervados por el nervio hipogloso (XII par craneal), el cual, de acuerdo con algunos anatomistas comparativos, es una serie de nervios espinales muy modificados. A pesar de su localización final en la cabeza, estos músculos están sujetos a la misma regulación molecular inicial de la miogénesis que los músculos del tronco.
Las anomalías y variaciones de los músculos esqueléticos son bastante comunes. Algunas, como la ausencia de partes del pectoral mayor, se asocian con malformaciones de otras estructuras.
La discusión in extenso de las anómalas de músculos específicos requiere un nivel de conocimientos de la anatomía que está mas allá del que se presume para este texto.
La distrofia muscular es un grupo de enfermedades genéticas caracterizadas por la degeneración y regeneración repetidas de varios grupos musculares durante la vida posnatal. En la distrofia muscular de Duchenne, que afecta a los niños mas pequeños, las fibras musculares carecen de una proteína asociada a la membrana llamada distrofina. Aunque aún no se conoce la función exacta de esta proteína, su ausencia parecer hacer que las fibras musculares sean más susceptibles al daño cuando se someten a esfuerzo físico.
Si bien es un músculo estriado, el músculo cardíaco difiere del esquelético en muchos aspectos su desarrollo embrionario. Derivadas del mesodermo esplácnico del embrión joven, las células musculares cardíacas se originan en células del miocardio. Las disimilitudes en la diferenciación de los músculos cardíaco y esquelético se evidencian pronto, ya que la MyoD y otros reguladores claves de la diferenciación del músculo esquelético no se expresan en el desarrollo inicial del músculo cardíaco. No obstante, las células musculares cardíacas y esqueléticas primitivas expresan isoformas de moléculas que son características de células maduras del otro tipo. Por ejemplo, tanto las células musculares cardíacas como las esqueléticas del embrión expresan altos niveles de alfa actina cardíaca; sin embargo, al nacimiento de la expresión de esta molécula disminuye en el músculo esquelético y permanece alta en el músculo cardíaco. Vale la pena anotar que en la hipertrofia cardíaca, las células maduras del músculo cardíaco comienzan a expresar grandes cantidades de RNAm de alfa actina esquelética.
Los primeros mioblastos cardíacos contienen cantidades relativamente elevadas de miofibrillas en su citoplasma y pueden contraerse de forma considerable. En el embrión los miocitos cardíacos mononucleados se enfrentan a un problema difícil: las células del corazón en desarrollo deben seguir contrayéndose al tiempo que la masa del corazón aumenta.
Este requerimiento funcional precisa que los miocitos cardíacos entre en mitosis aunque su citoplasma contenga muchos haces de filamentos contráctiles (figura 8_35). Las células del cuerpo suelen perder su capacidad para dividirse cuando su citoplasma contiene estructuras características del estado diferenciado. Para hacer frente a este problema los miocitos cardíacos desensamblan parcialmente sus filamentos contráctiles durante la mitosis. En contraste con los músculos esqueléticos, los miocitos cardíacos no se fusionan sino que más bien permanecen como células individuales, aunque pueden volverse binucleadas. Los miocitos cardíacos mantienen un contacto estructural y funcional estrecho a través de los discos intercalares, que unen las células adyacentes.
En el curso del desarrollo un grupo de células musculares cardíacas emprende una vía distinta de diferenciación caracterizada por aumento de tamaño, reducción de la concentración de miofibrillas y un gran aumento de la concentración de glucógeno en el citoplasma. Estas células forman el sistema de conducción, algunos de cuyas partes constituyen las células de Purkinje. Estas células también expresan un perfil diferente de isoformas de proteínas contráctiles de los miocitos ya sean auriculares o ventriculares.
Como sucede con el músculo cardíaco, gran parte del músculo liso del organismo se origina del mesodermo esplácnico. Las excepciones son el músculo ciliar y los músculos del esfínter de la pupila del ojo (que se deriva del ectodermo de la creta neural), y gran parte del músculo liso vascular (que suele originarse en el mesodermo local). Se sabe muy poco sobre la morfología y los mecanismos que subyacen a la diferenciación de las células del músculo liso.
|
CASO CLINICO
Un pediatra notó que un nuevo paciente, un niño de un año y medio de edad, tenia un cuello más corto de lo normal y una línea de implantación del cabello más baja de lo usual en el cuello. La información de los antecedentes familiares no dio evidencias de otros parientes con la misma afección. El examen radiológico reveló que el cuello del niño sólo tenia seis vértebras cervicales. El pediatra preguntó entonces si la madre recordaba haber tomado o estado expuesta a ciertos compuestos al comienzo del embarazo.
|
1. ¿Bajo cuál de estas condiciones se activan las células satélites del músculo?
A. Crecimiento normal de las fibras musculares
B. Regeneración de las fibras musculares
C. Hipertrofia de las fibras musculares
D. Todas las anteriores
E. Ninguna de las anteriores
2. ¿Cuál de los componentes celulares de la epidermis es una avanzadilla del sistema inmunológico y funciona presentando antígenos a otras células inmunológicas?
A. Células de Merkel
B. Queratocitos
C. Células basales
D. Melanocitos
E. Células de Langerhans
3. ¿Cuál de las siguientes estructuras es de origen mesodérmico?
A. El tallo del pelo
B. El conducto mamario
C. La glándula sebácea
D. El músculo piloerector
E. Ninguna de las anteriores
4. La craneosinostosis es causada por una anomalía en el desarrollo de:
A. El foramen magno
B. Las suturas craneales
C. La base del cráneo
D. Las mandíbulas
E. Ninguna de las anteriores
5. ¿En el desarrollo de la fibra muscular cuál de los factores reguladores de la miogénesis se expresa más tarde?
A. Miogenina
B. MyoD
C. MRF- 4
D. Myf- 5
E. Pax- 3
6. En la bajada de la leche durante la lactancia, las células mioepiteliales se contraen en respuesta a:
A. Progesterona
B. Oxitocina
C. Estrógenos
D. Lactalbúmina
E. Caseína
7. ¿Cuál componente de la piel en desarrollo determina la naturaleza del pelo que se forma o el grosor de la epidermis en el feto?
8. Un varón tiene dos manchas de color marrón simétricas bilaterales de cerca de 8 mm de diámetro localizadas en la piel, aproximadamente a 8 cm por debajo de cada pezón. ¿Cuál es la aplicación de ellas?
9. ¿Por qué no se encuentra hueso craneal sobre un área donde falta parte del cerebro?
10. ¿Cómo se determinó que la musculatura e las extremidades procede de los somitas?
{kind=link}
{kind=link}
{kind=link}
{kind=link}
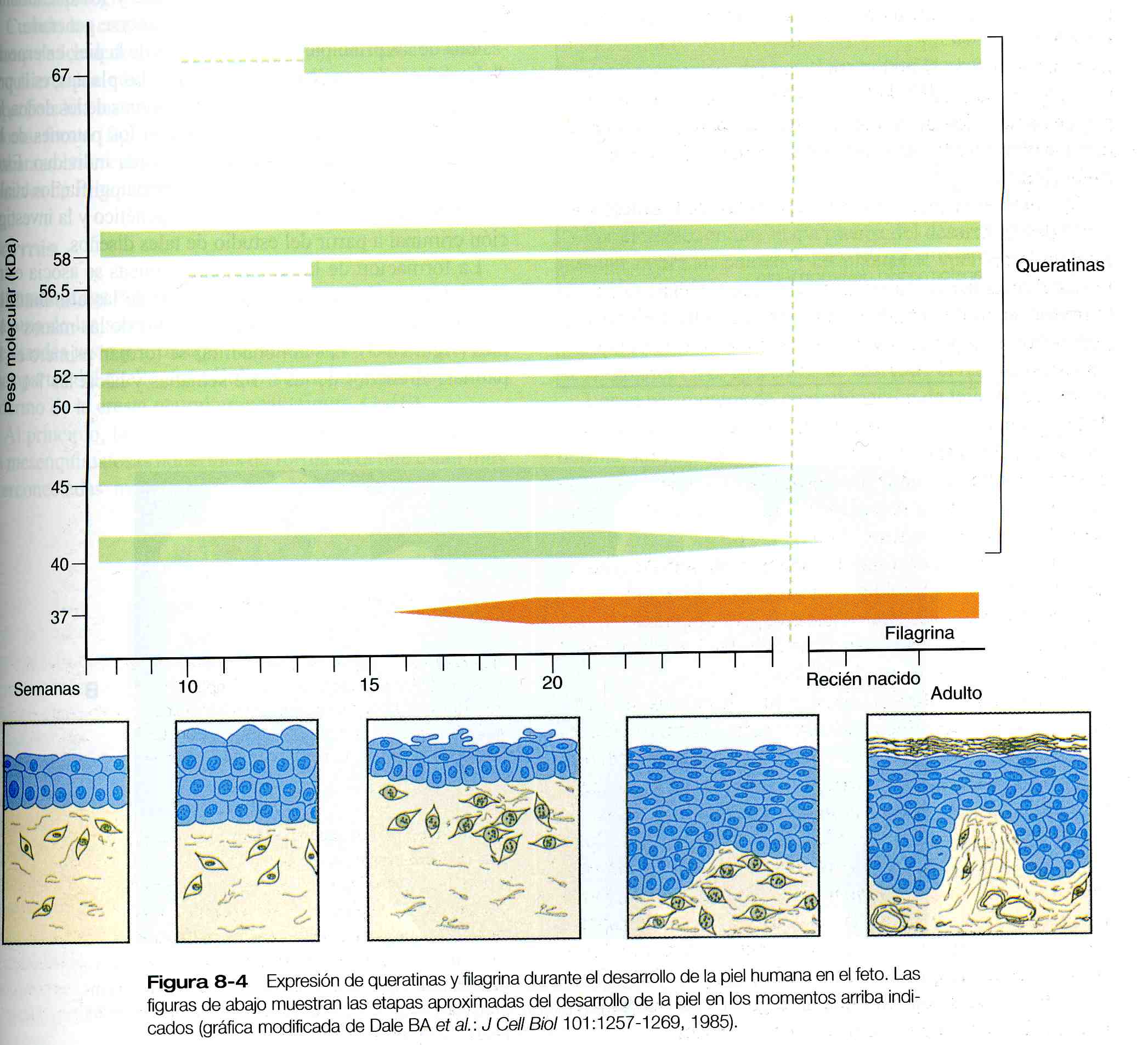{kind=link}
{kind=link}
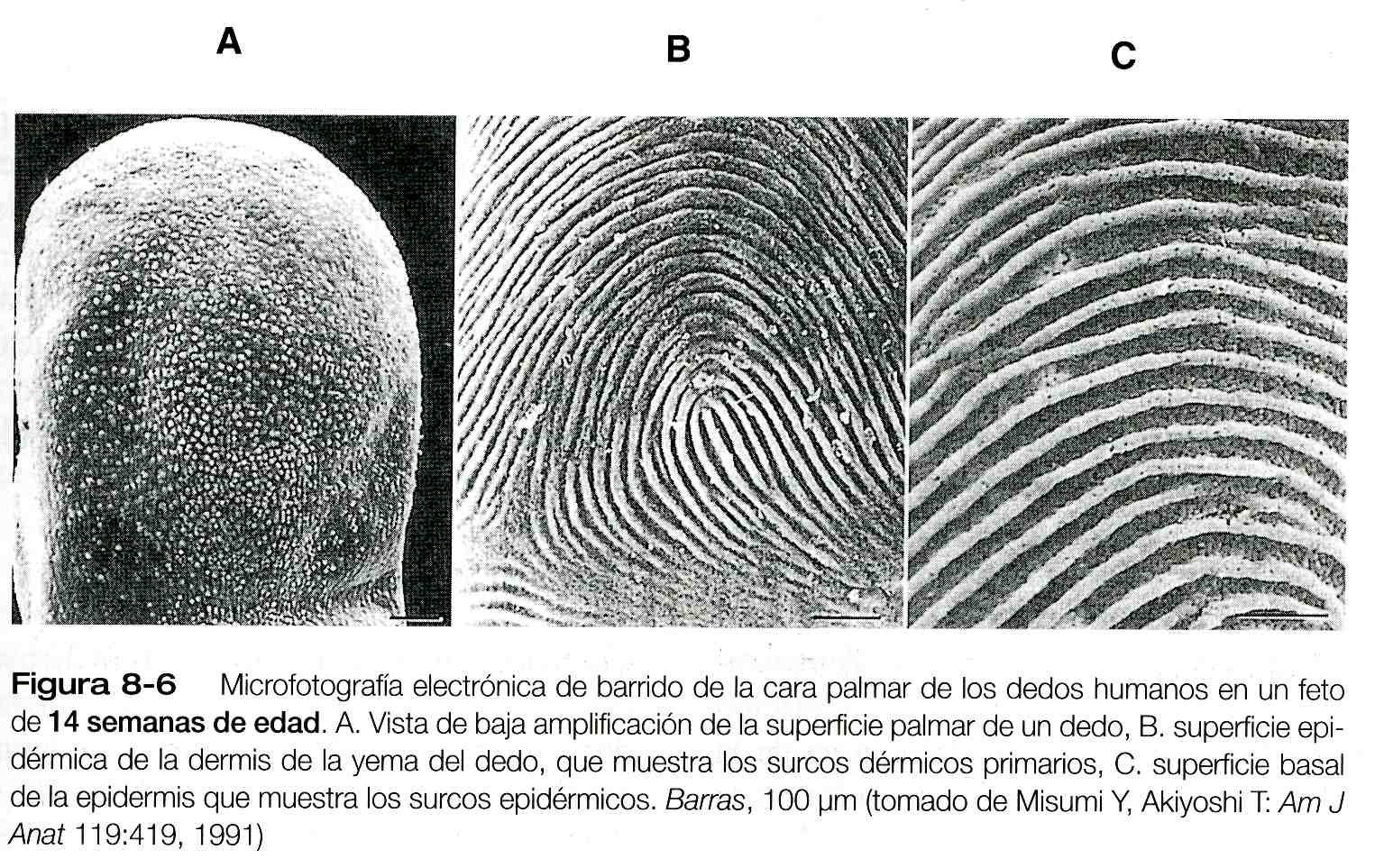{kind=link}
{kind=link}
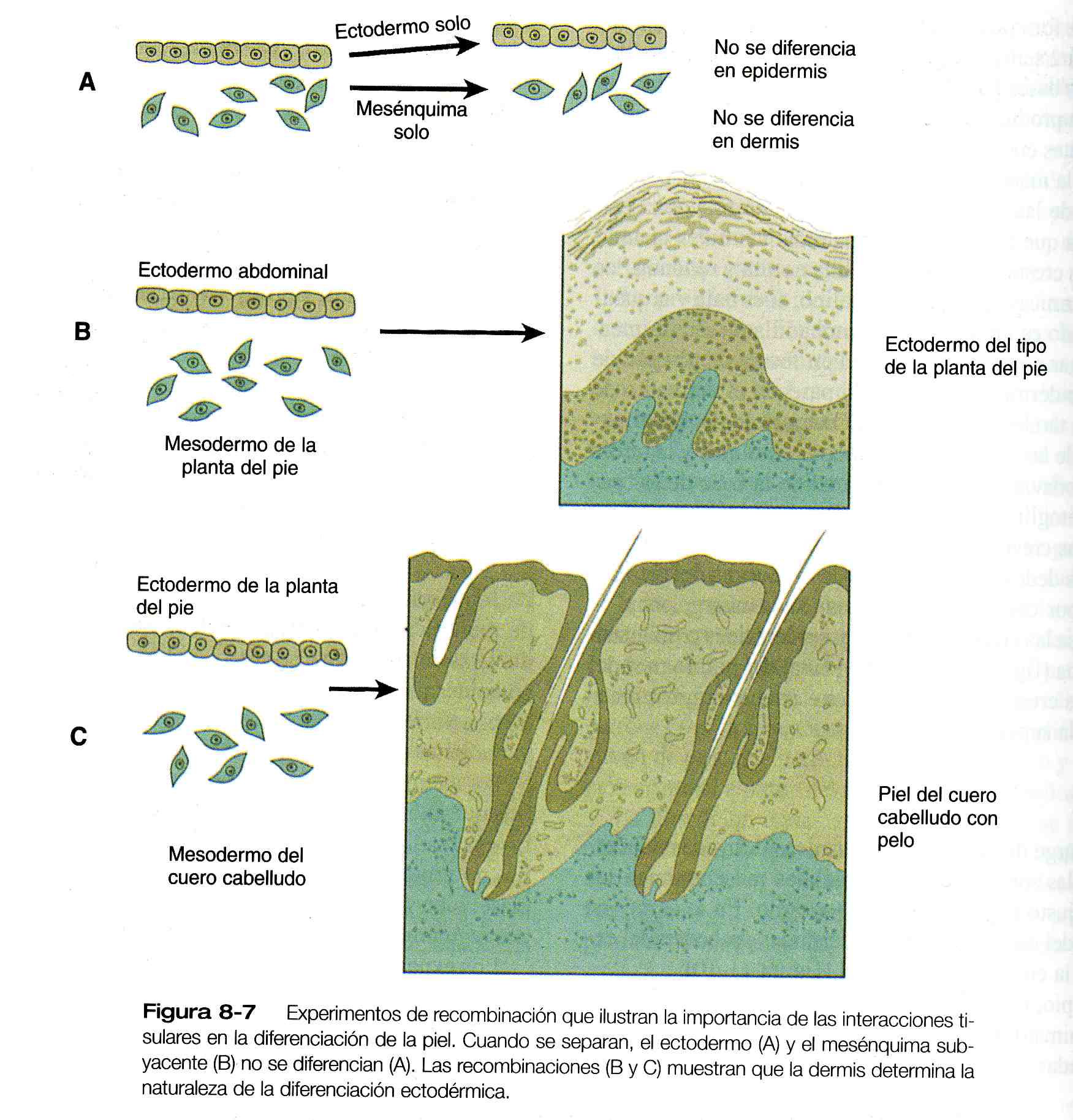{kind=link}
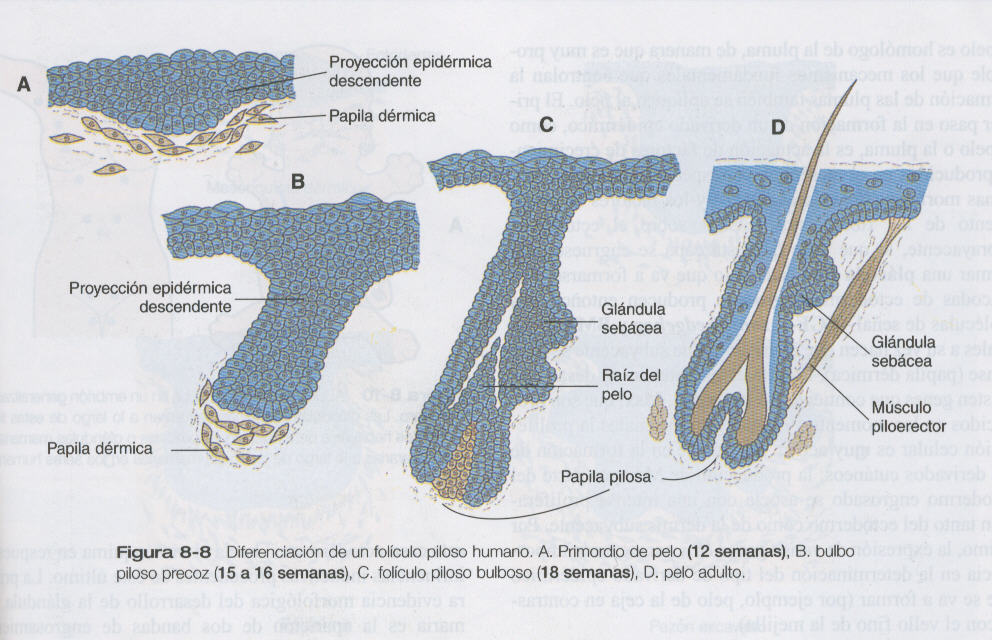{kind=link}
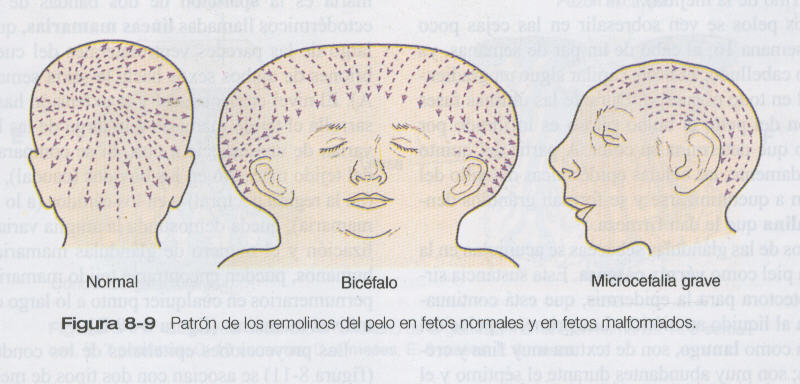{kind=link}
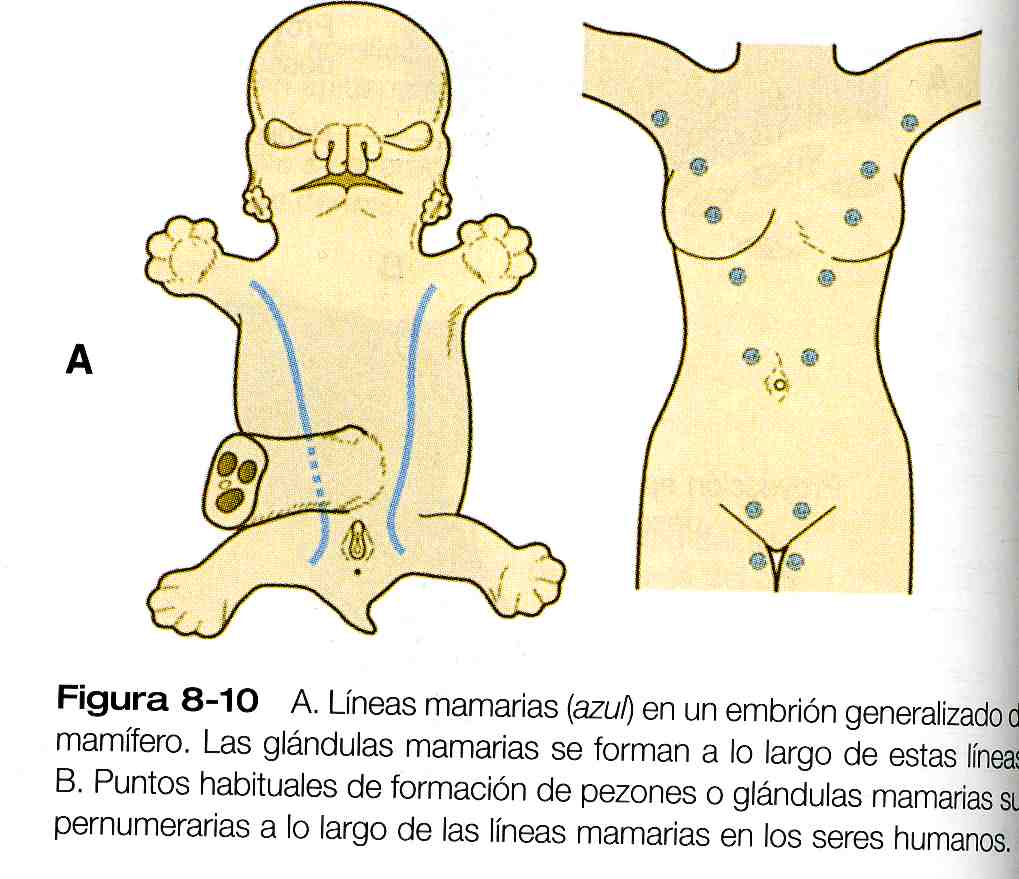{kind=link}
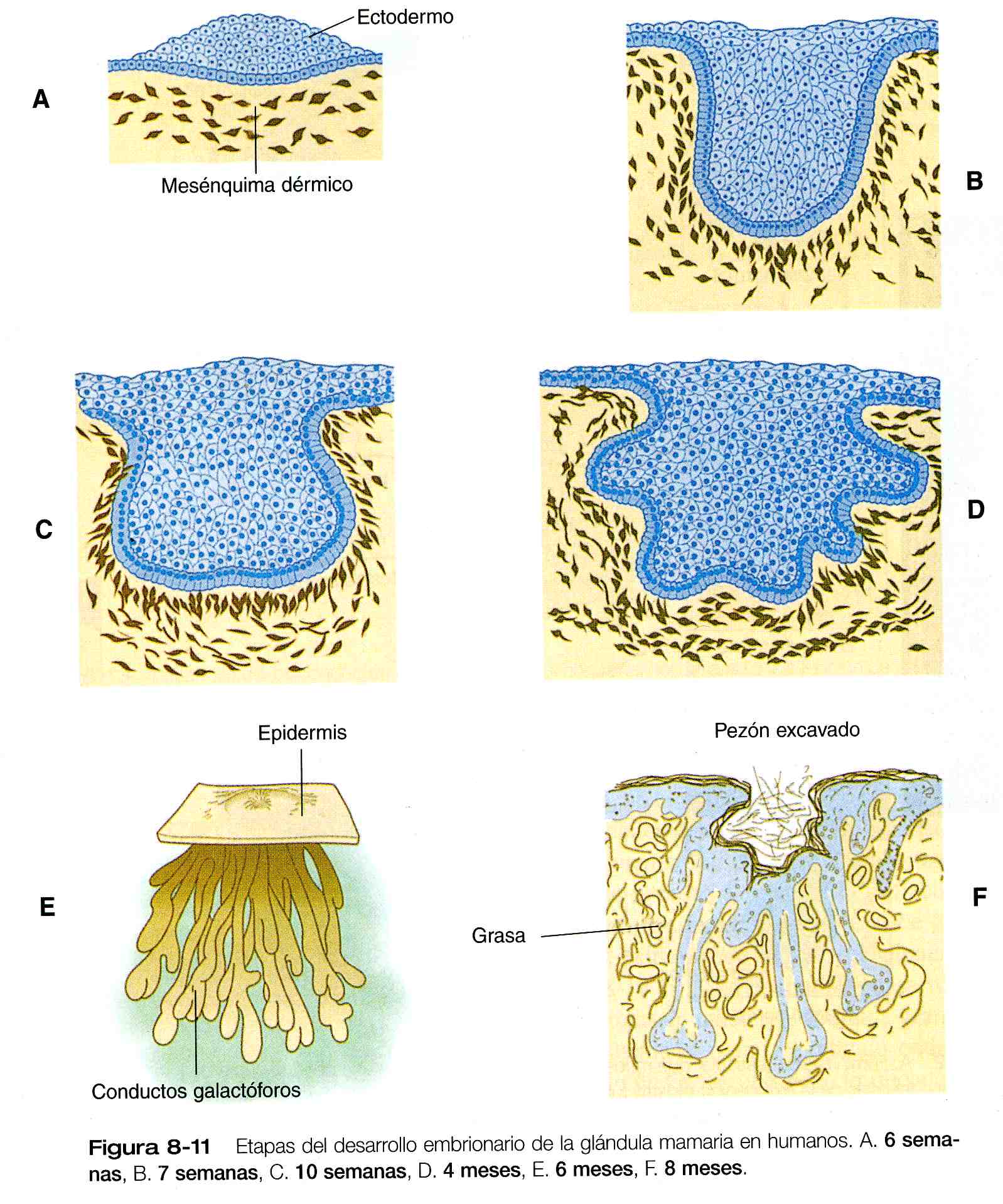{kind=link}
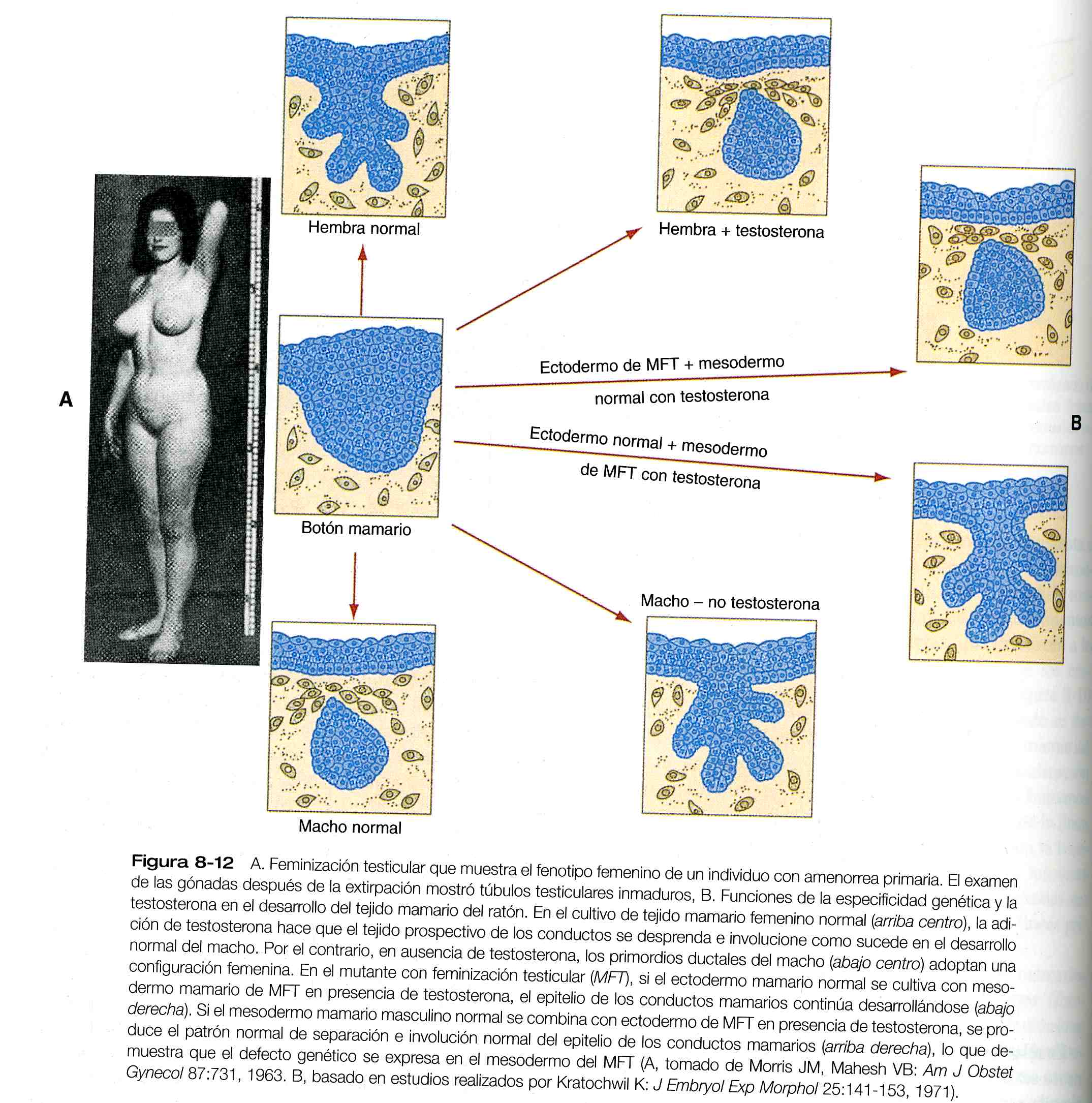{kind=link}
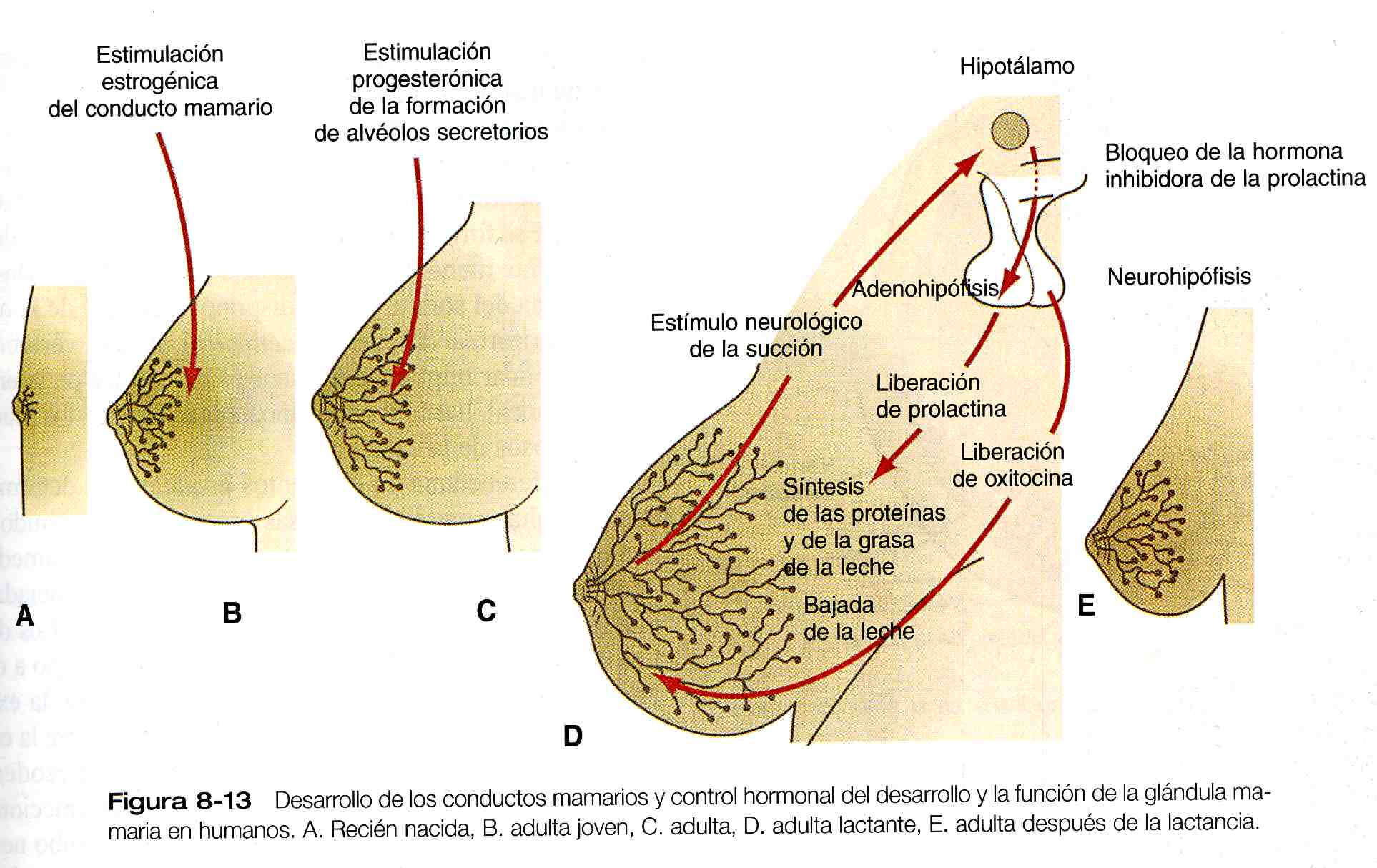{kind=link}
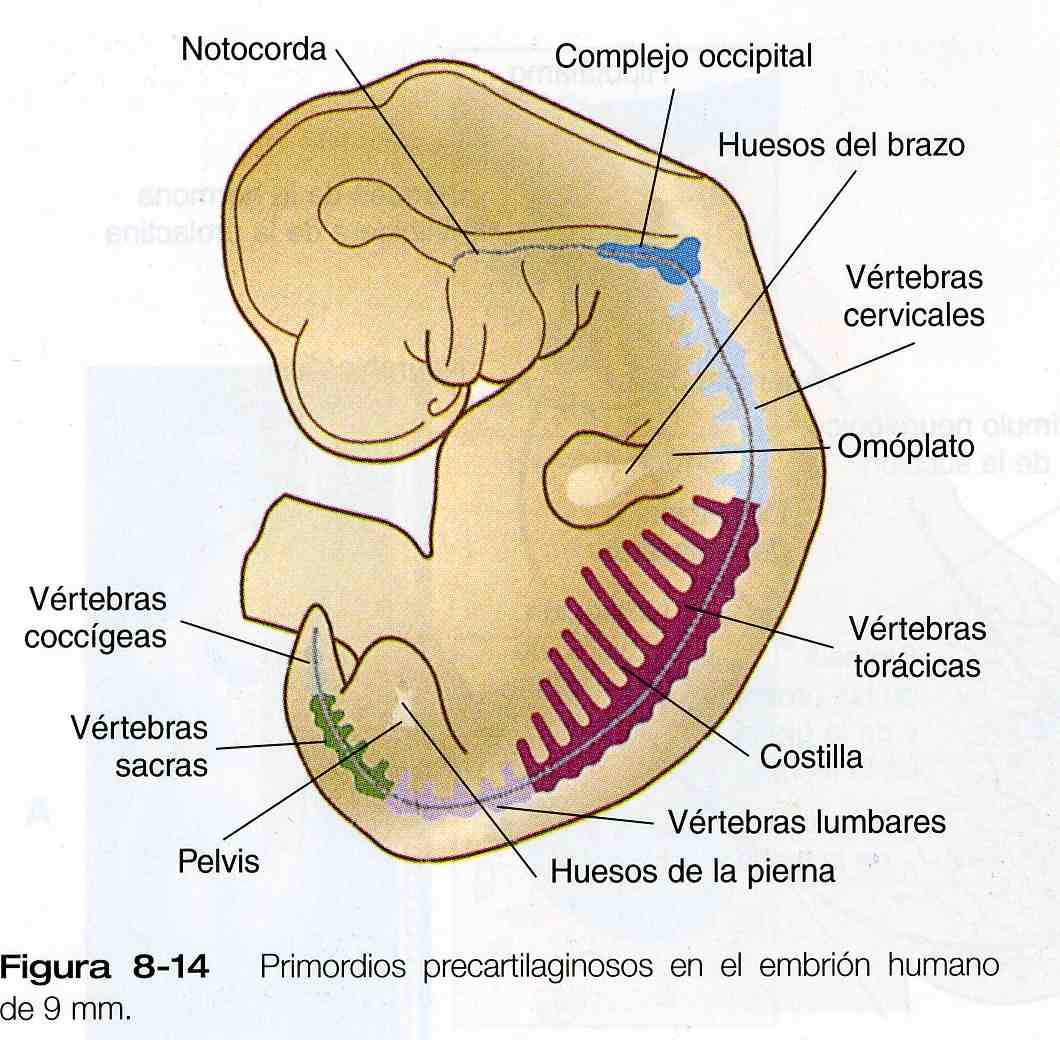{kind=link}
{kind=link}
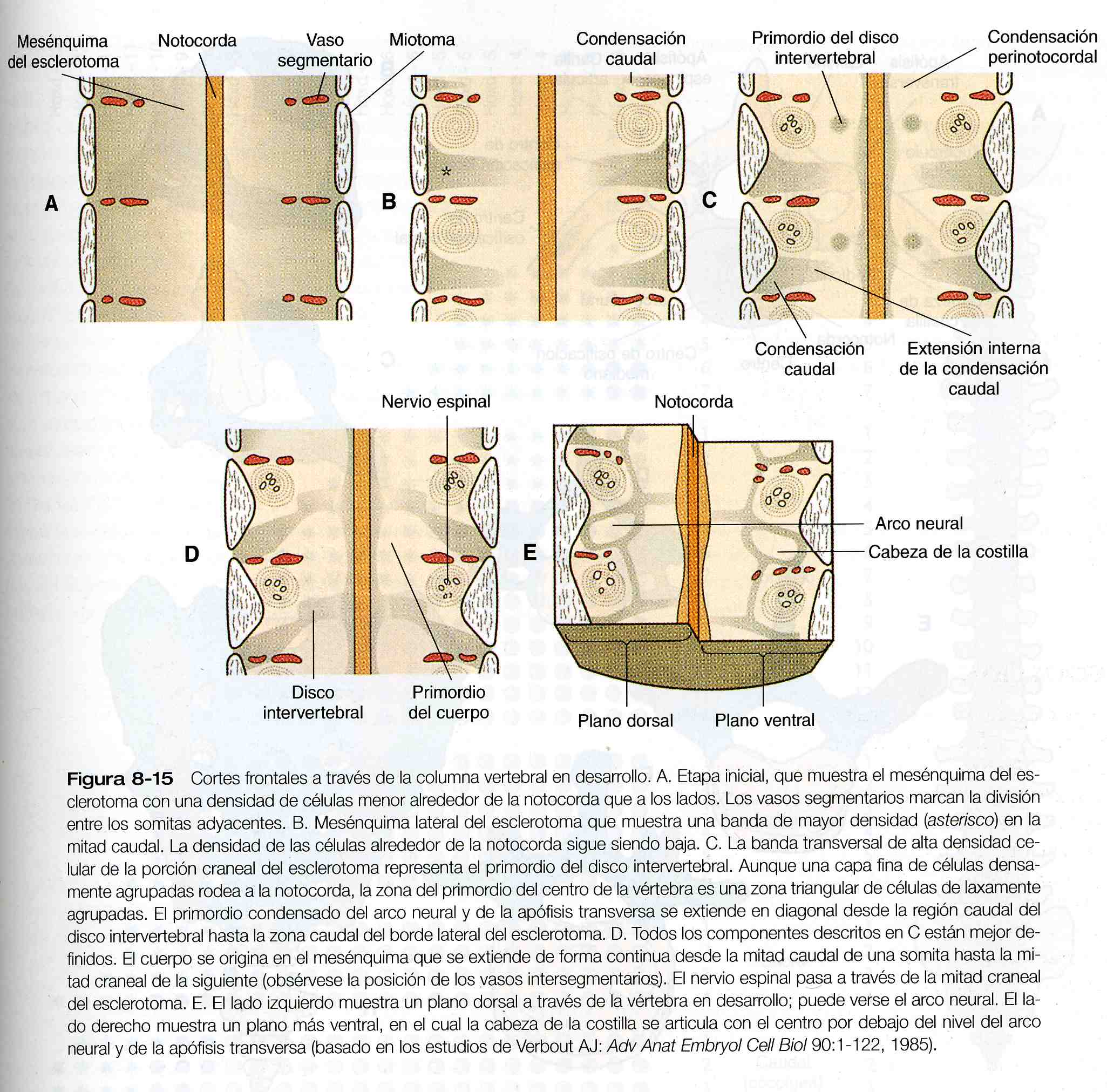{kind=link}
{kind=link}
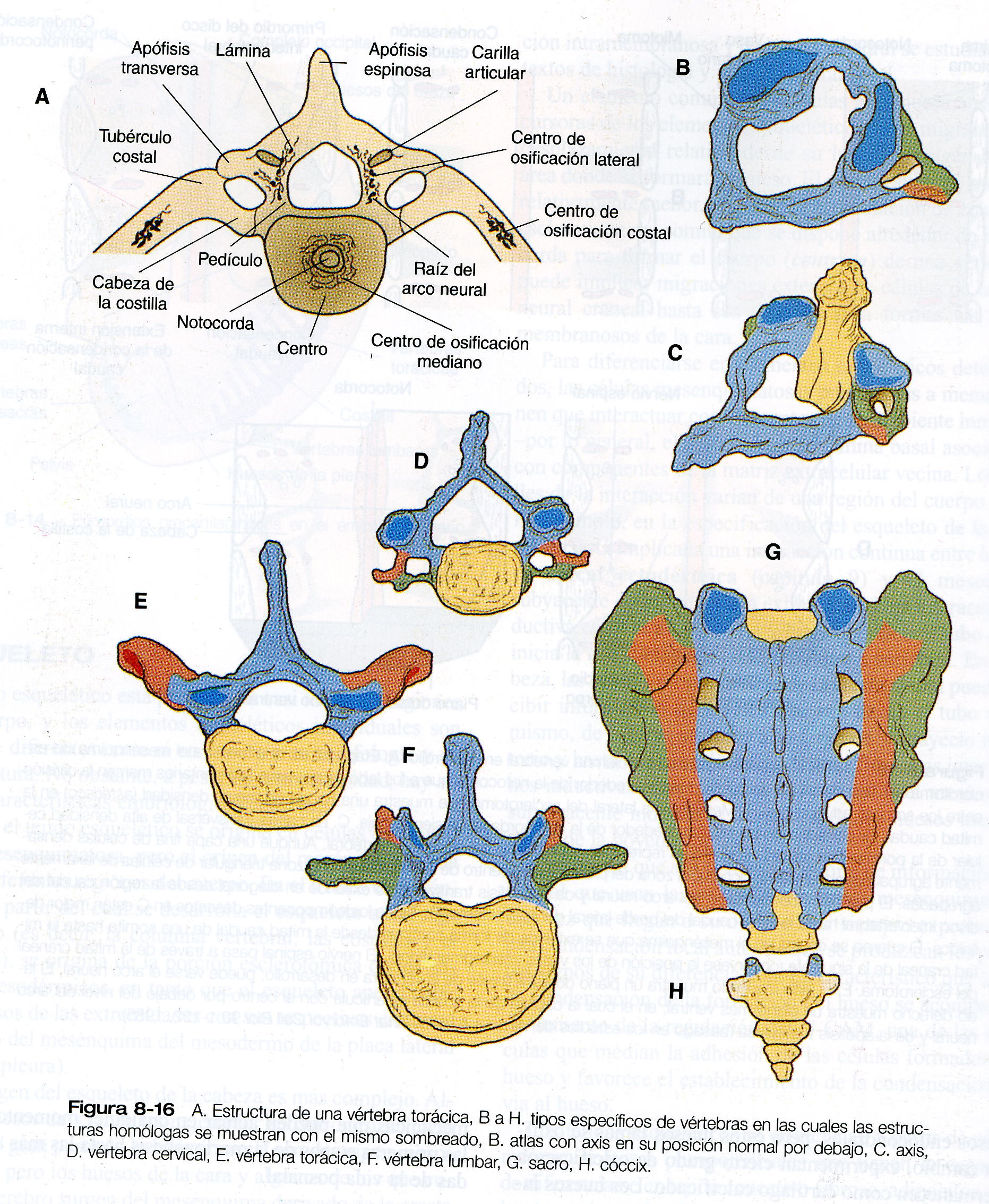{kind=link}
{kind=link}
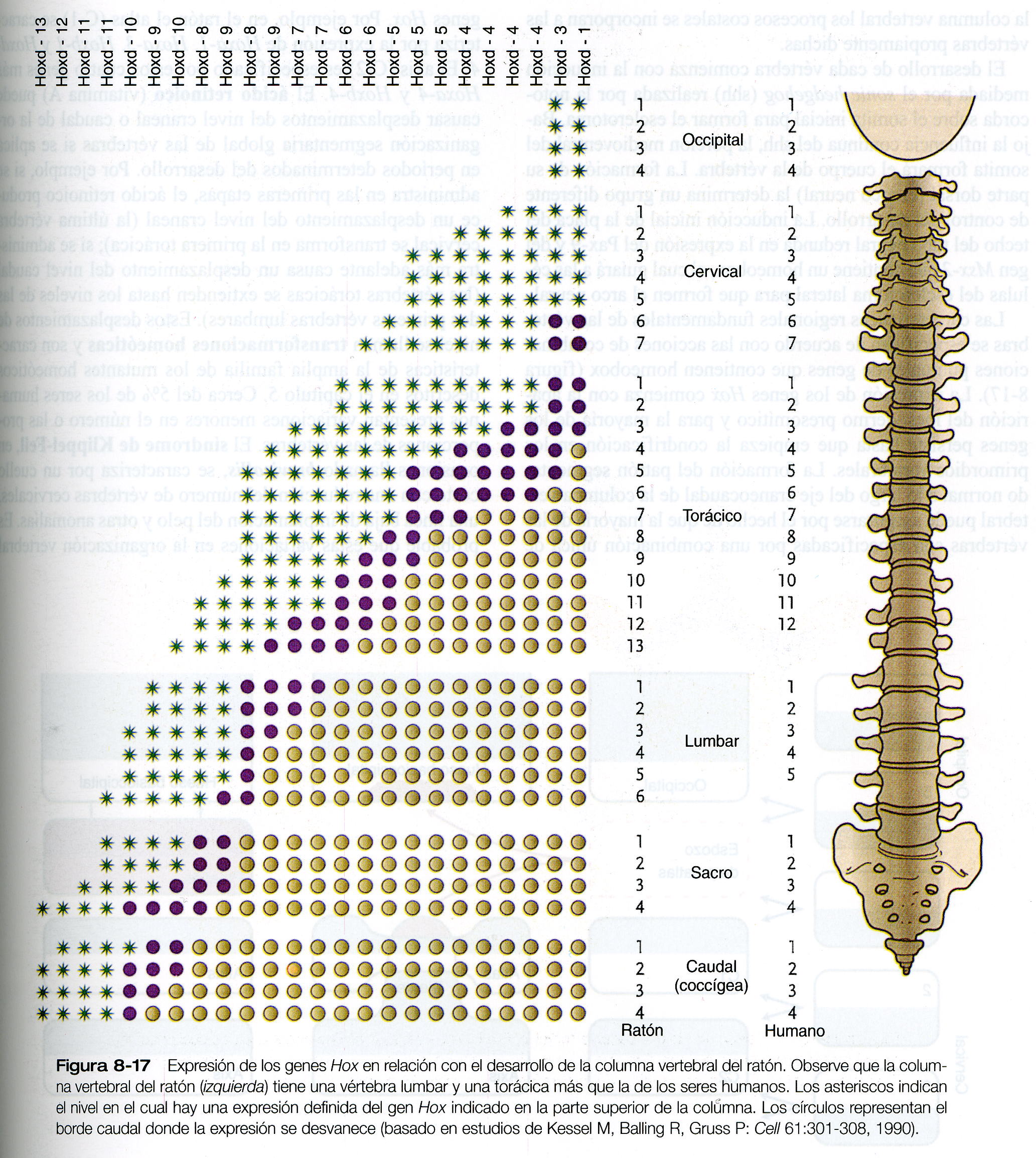{kind=link}
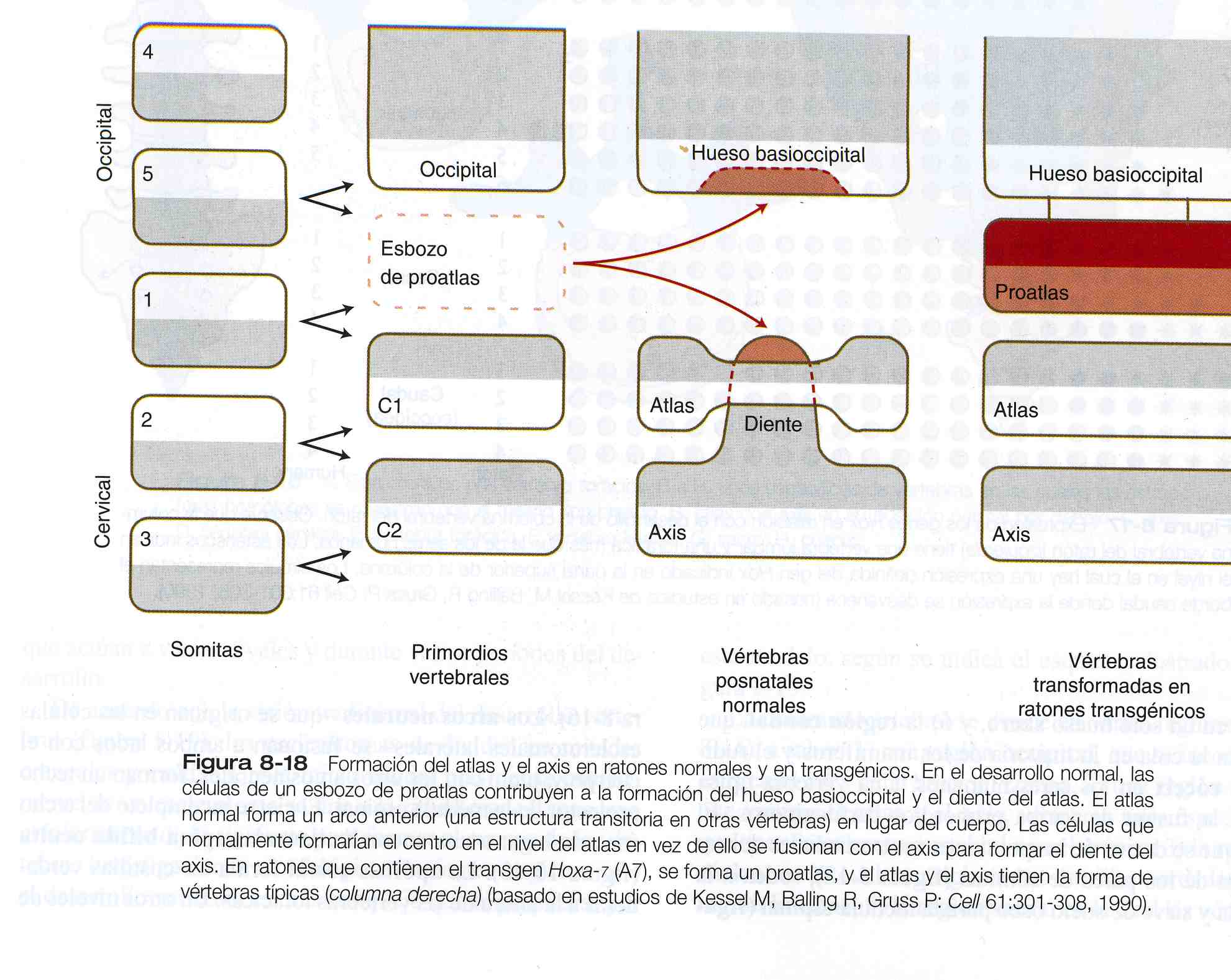{kind=link}
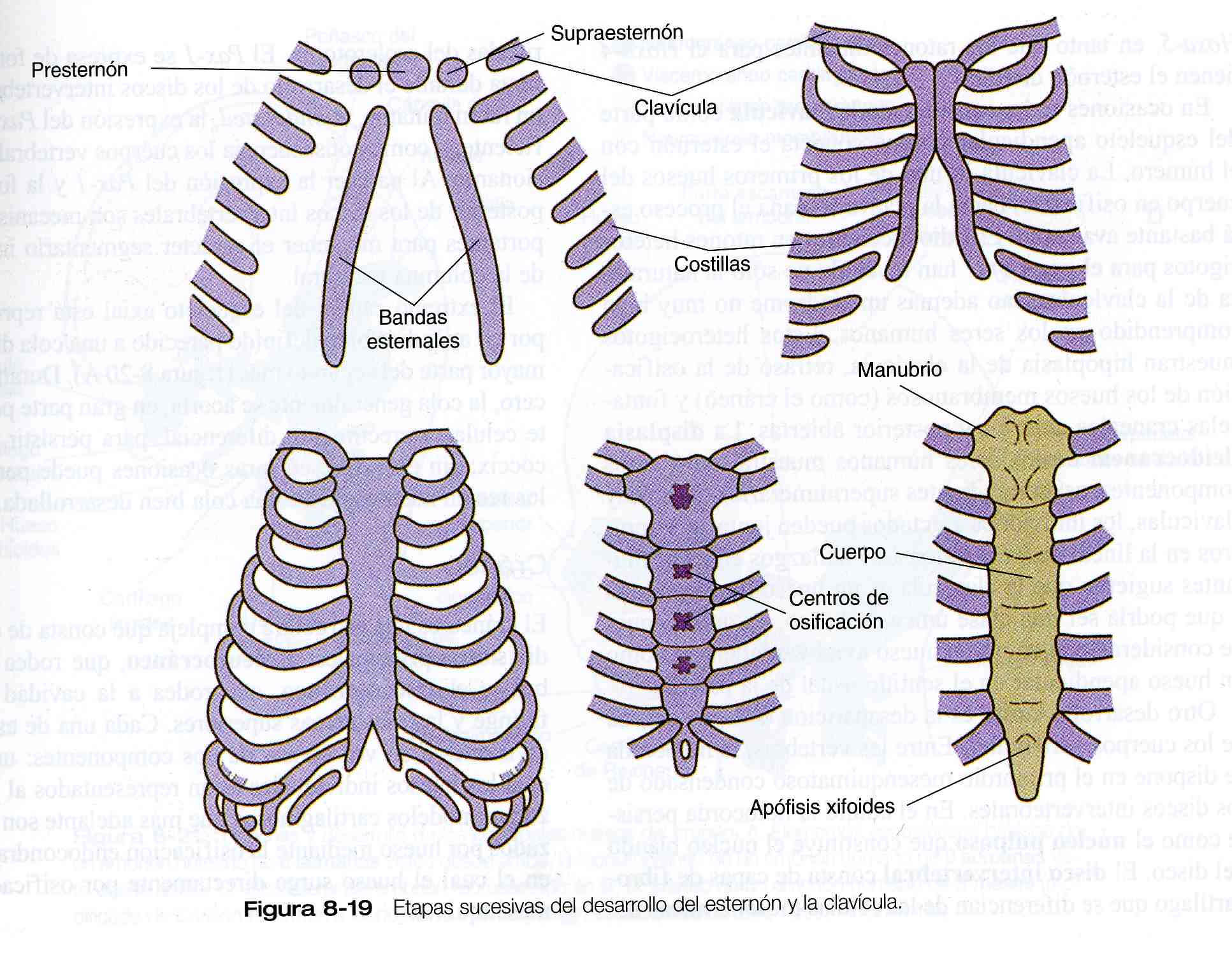{kind=link}
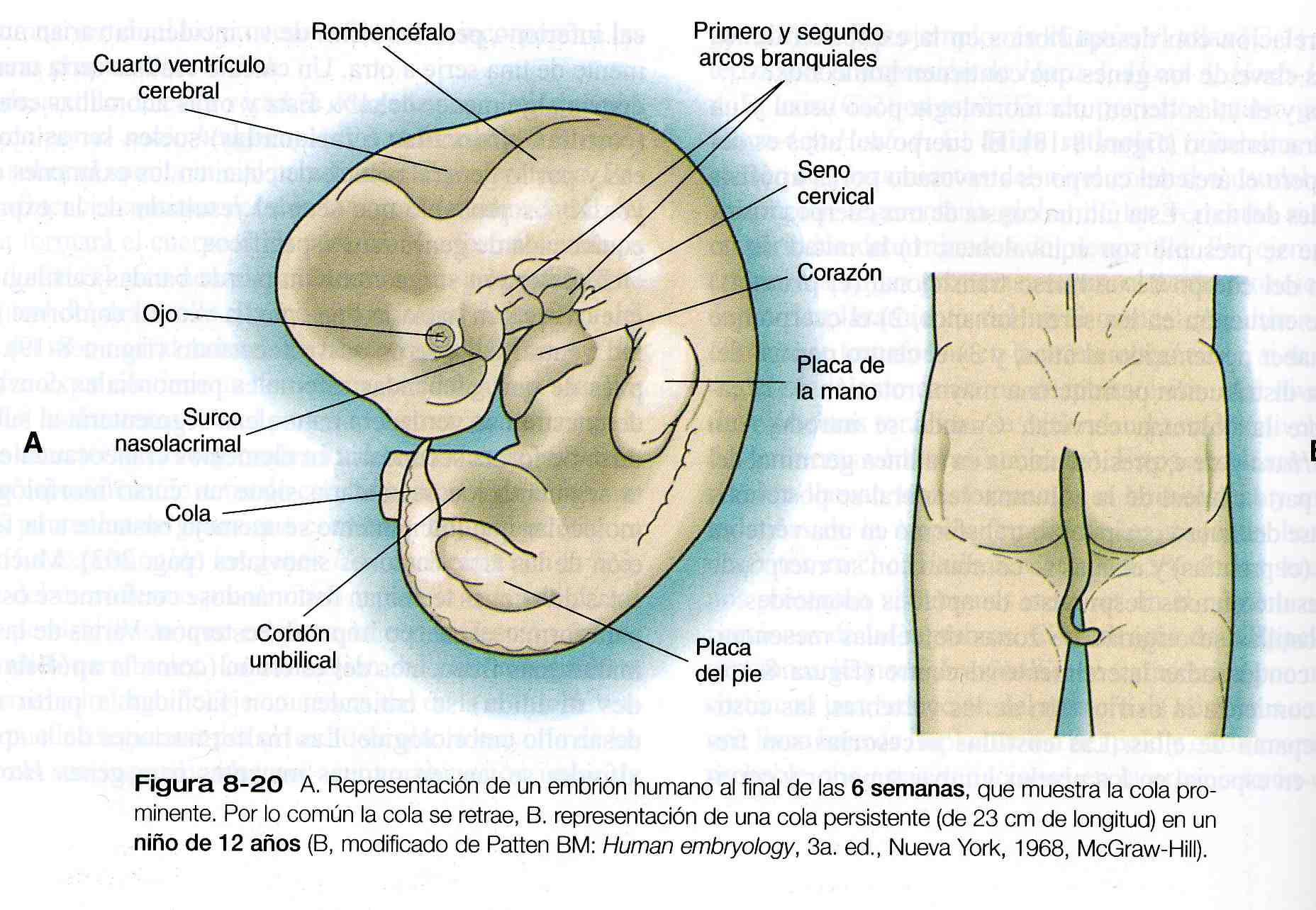{kind=link}
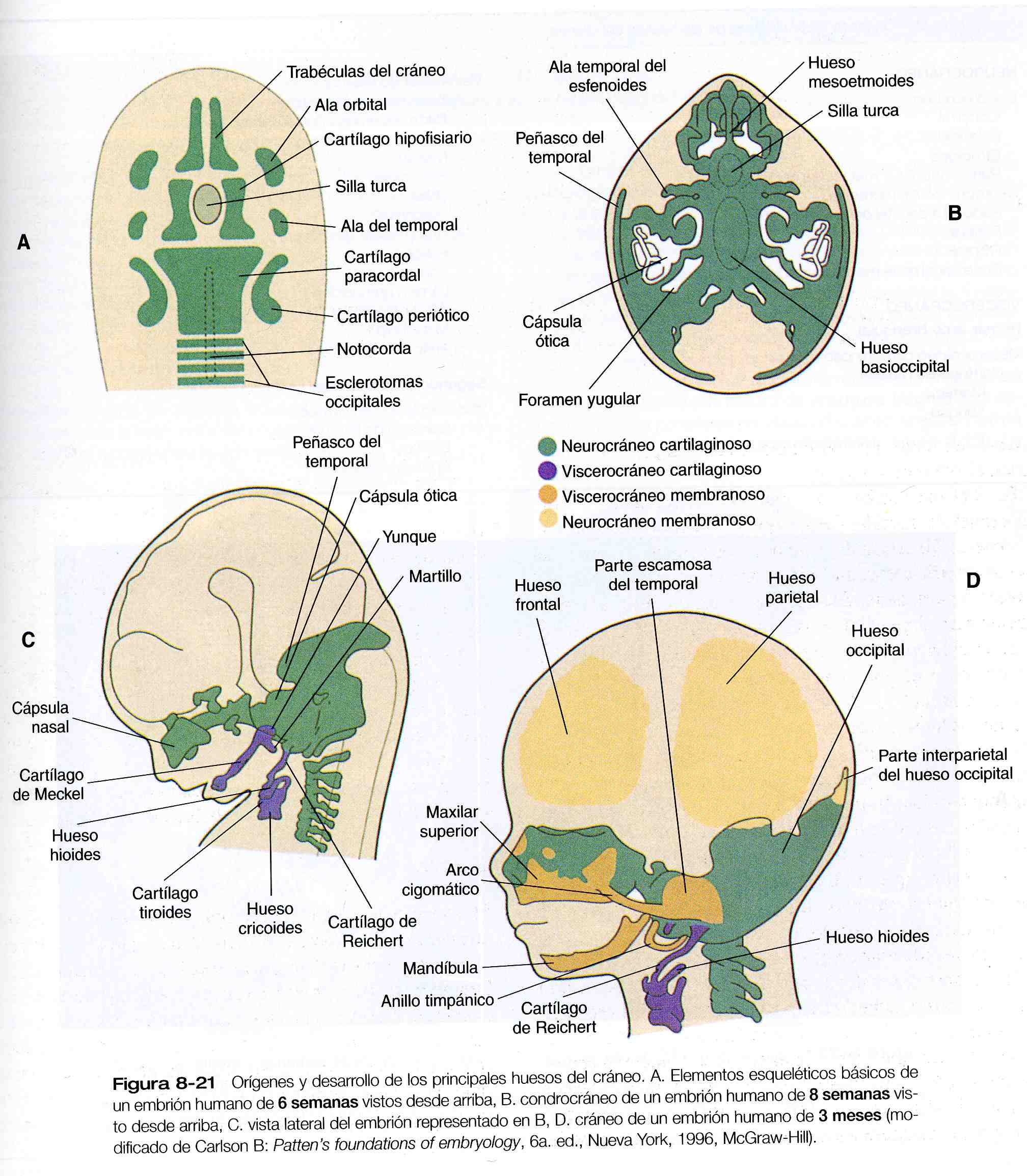{kind=link}
{kind=link}
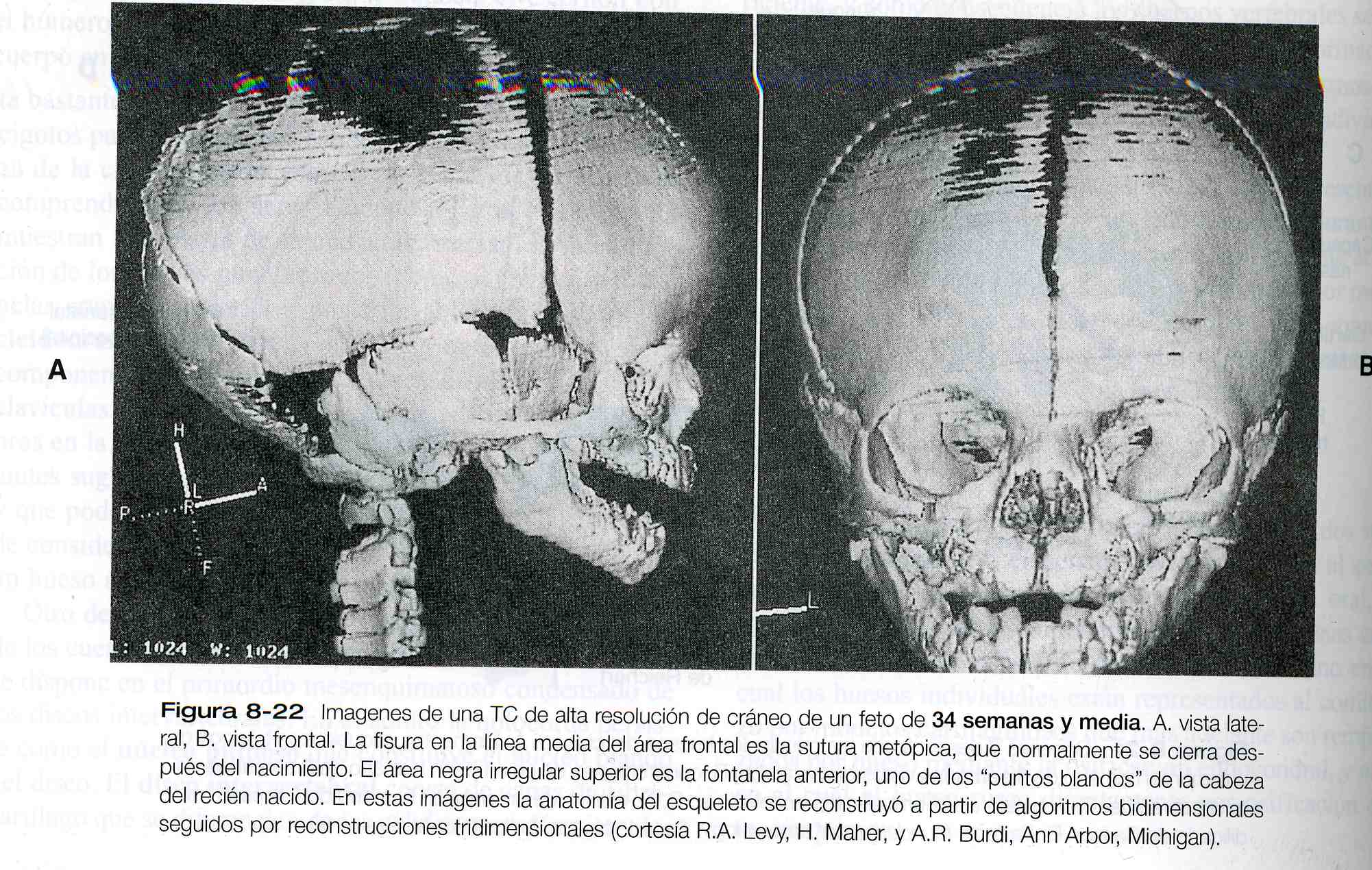{kind=link}
{kind=link}
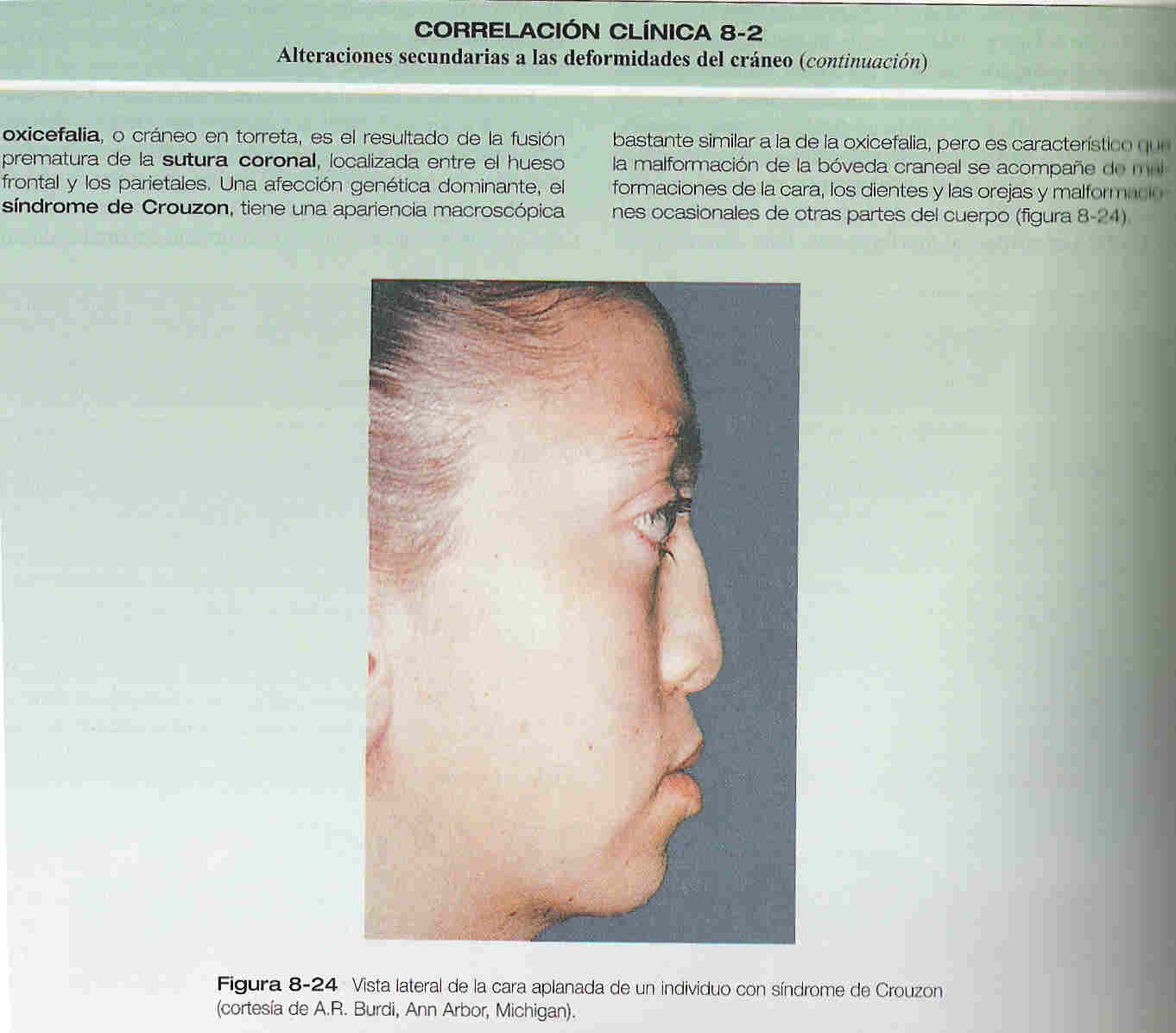{kind=link}
{kind=link}
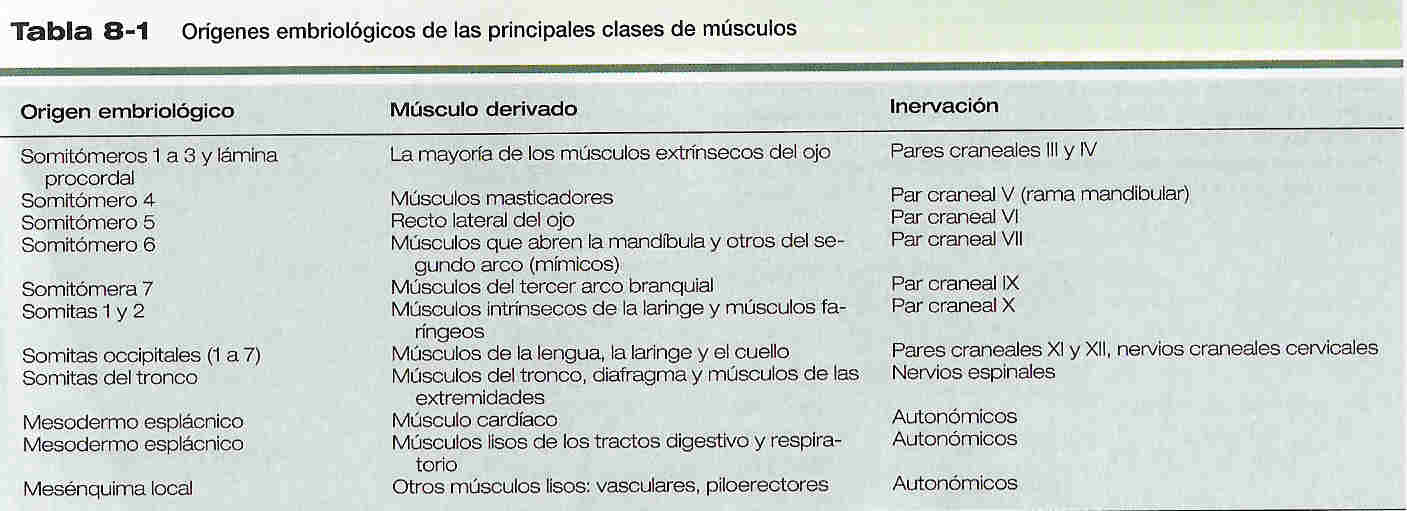{kind=link}
{kind=link}
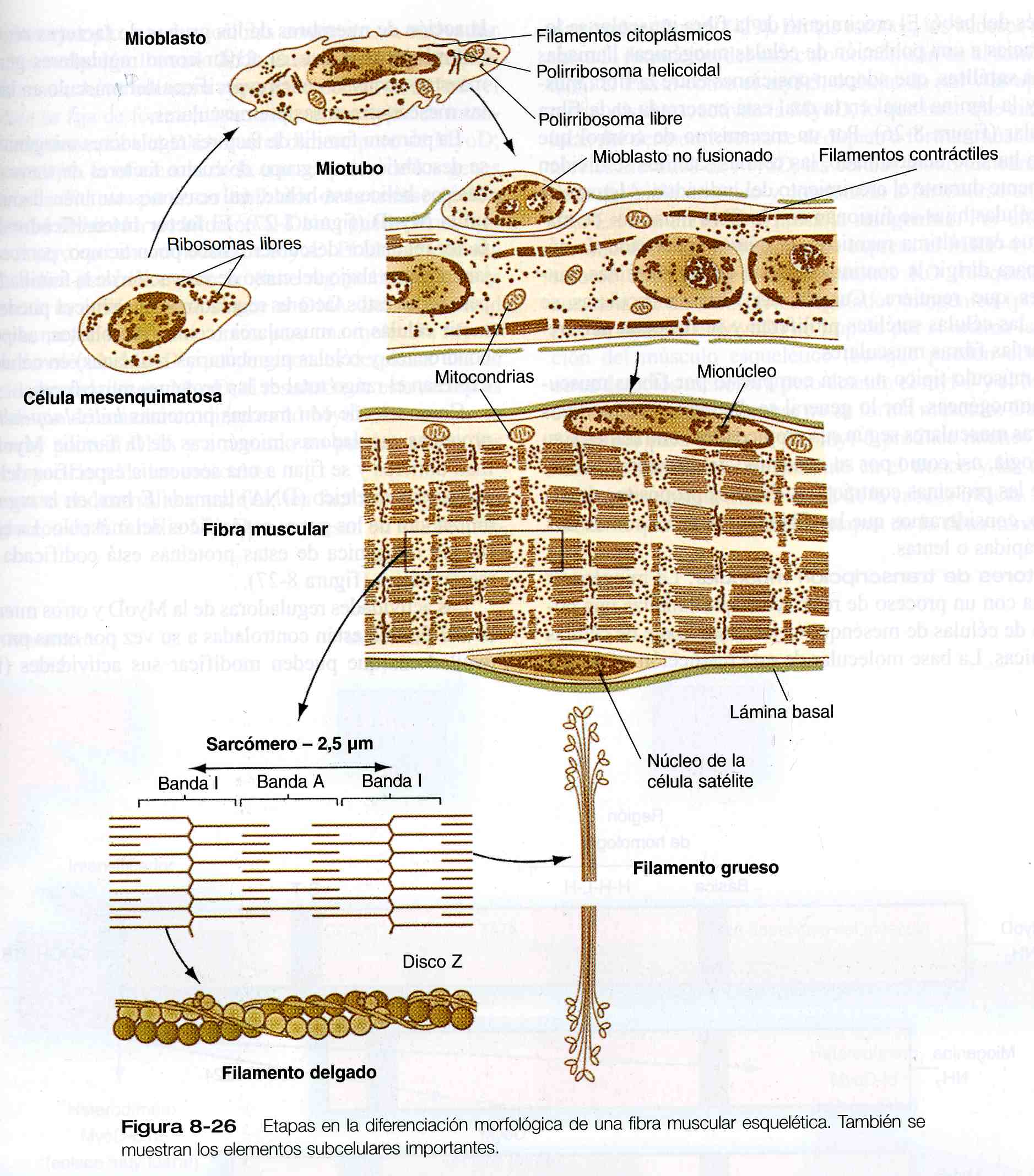{kind=link}
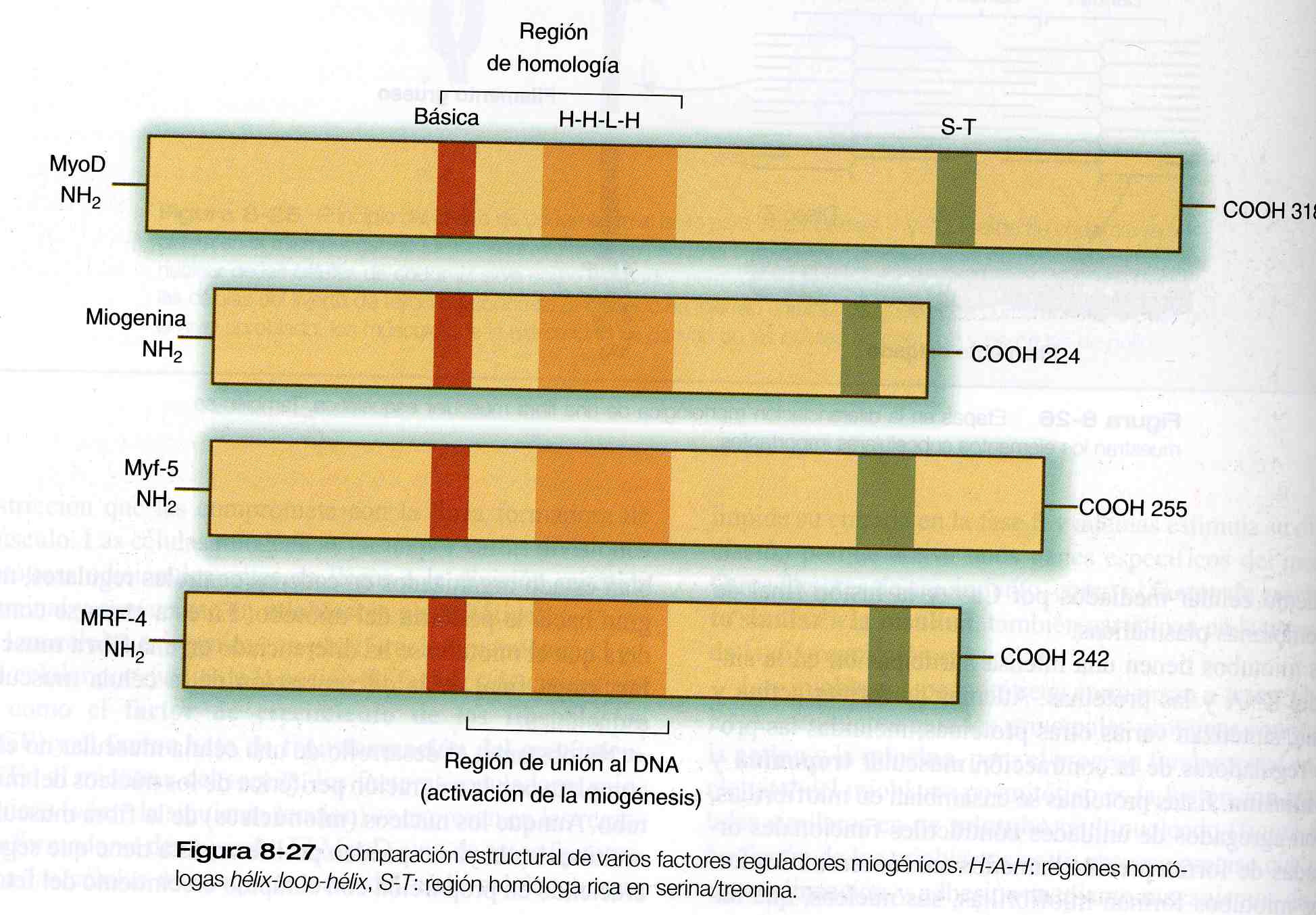{kind=link}
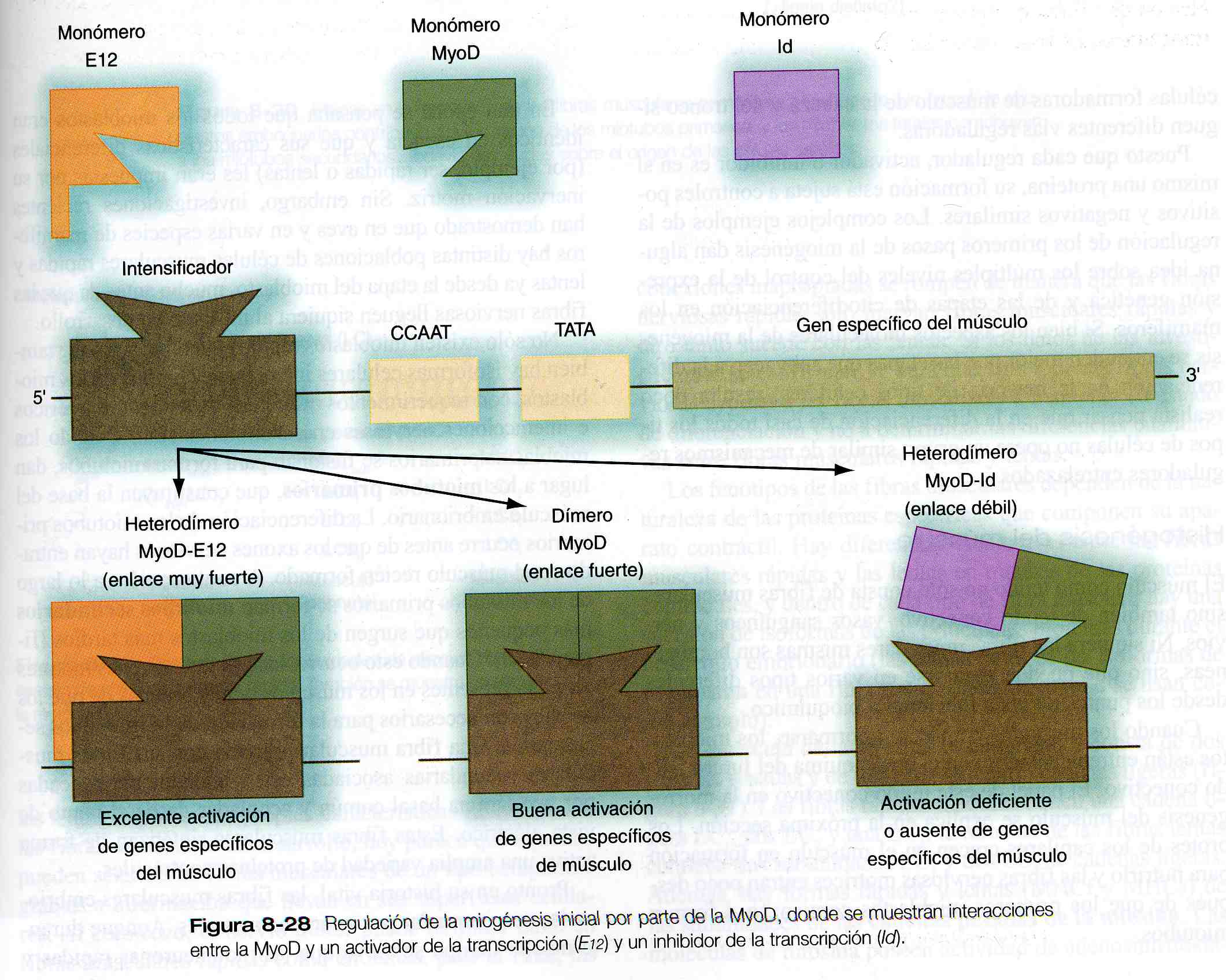{kind=link}
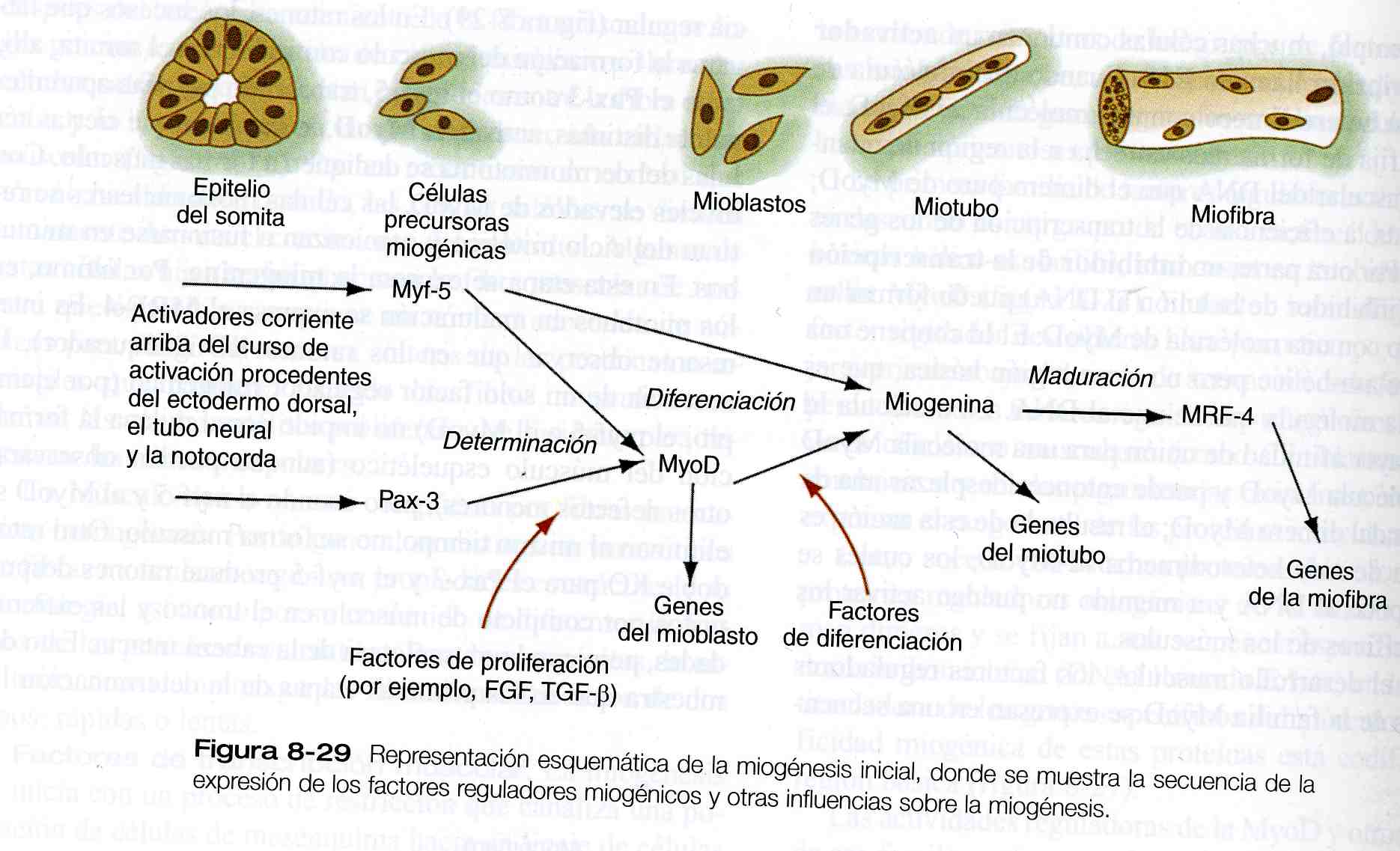{kind=link}
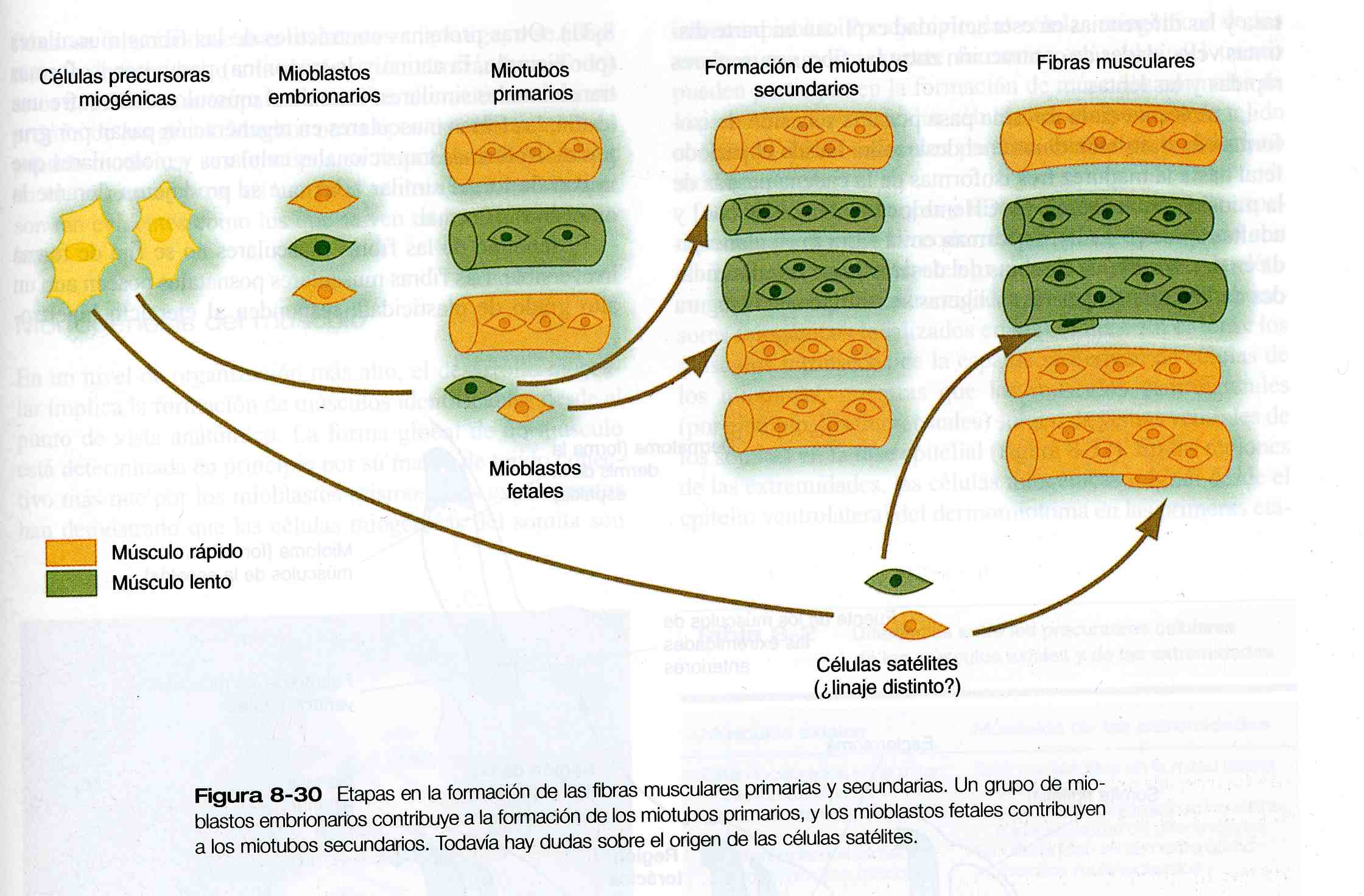{kind=link}
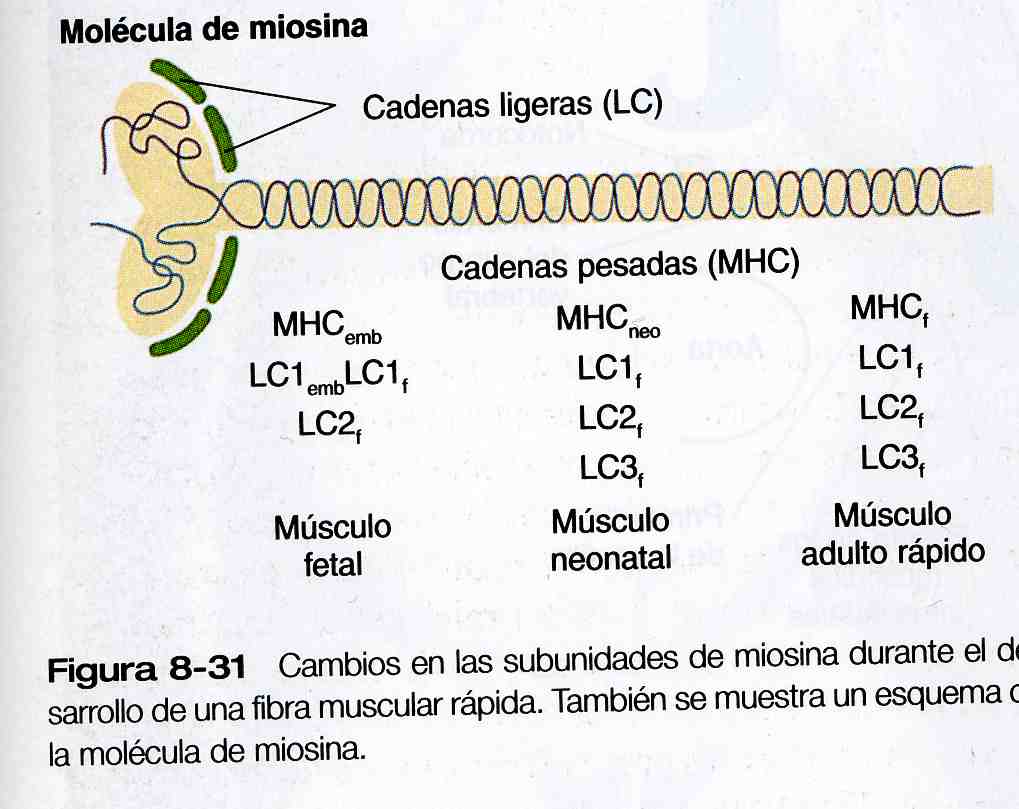{kind=link}
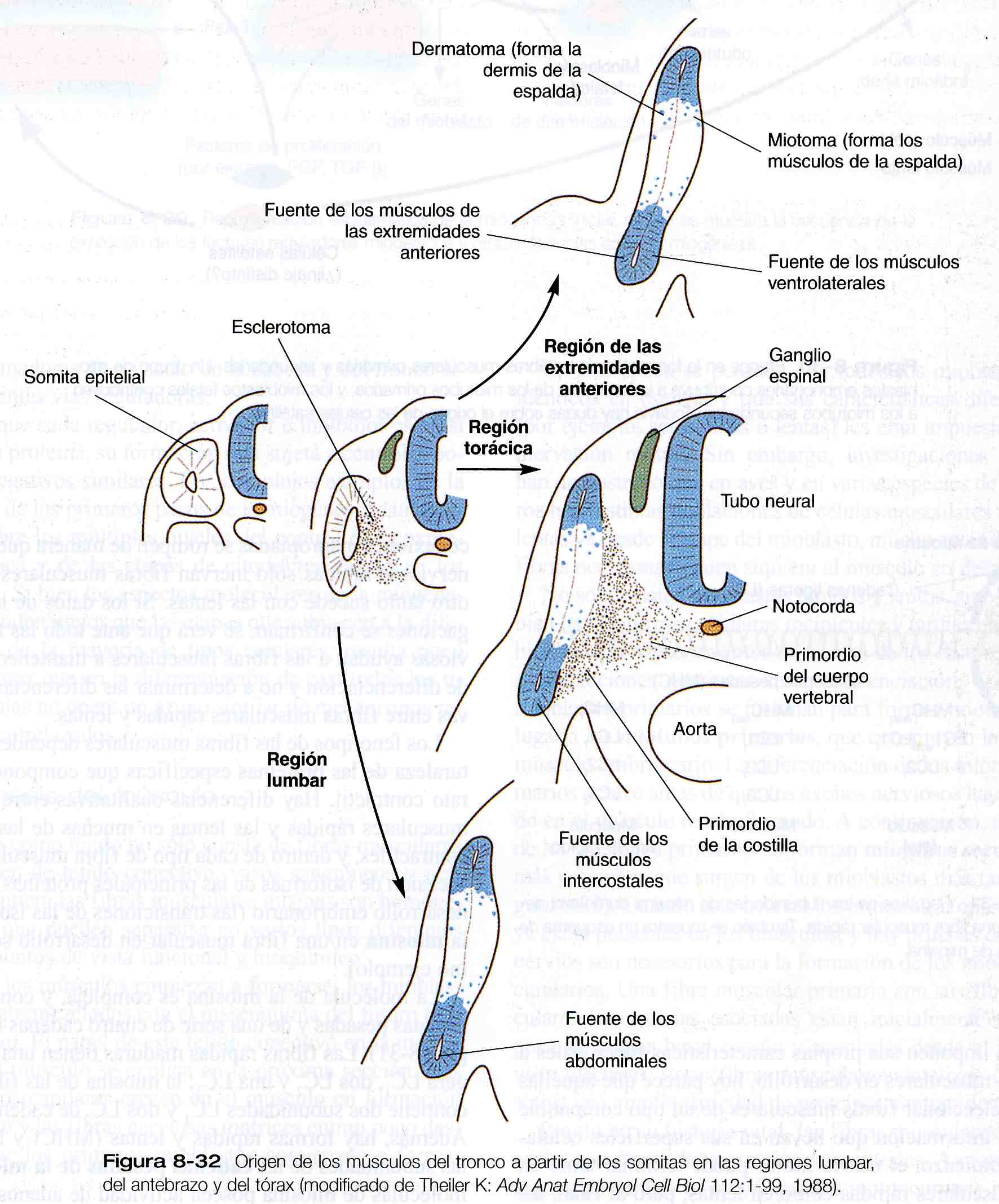{kind=link}
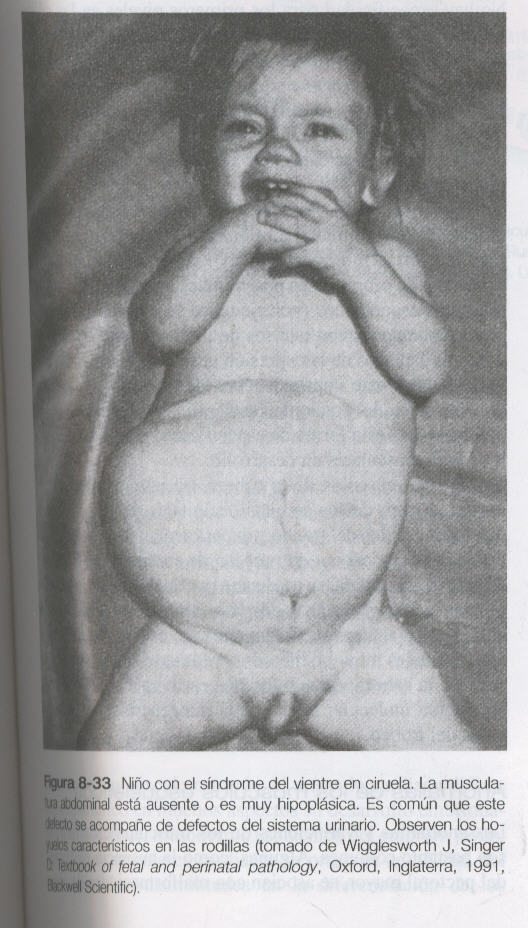{kind=link}
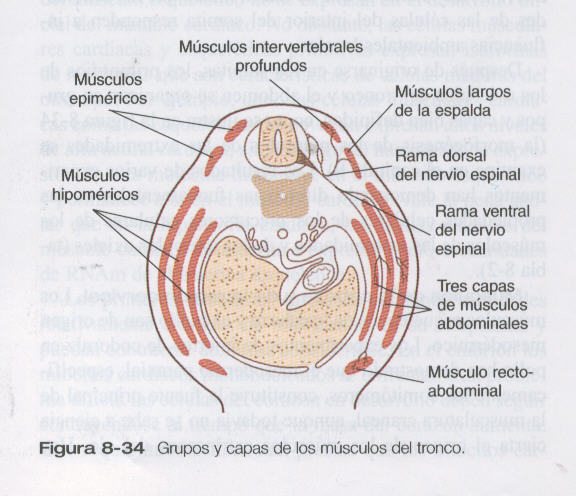{kind=link}
{kind=link}
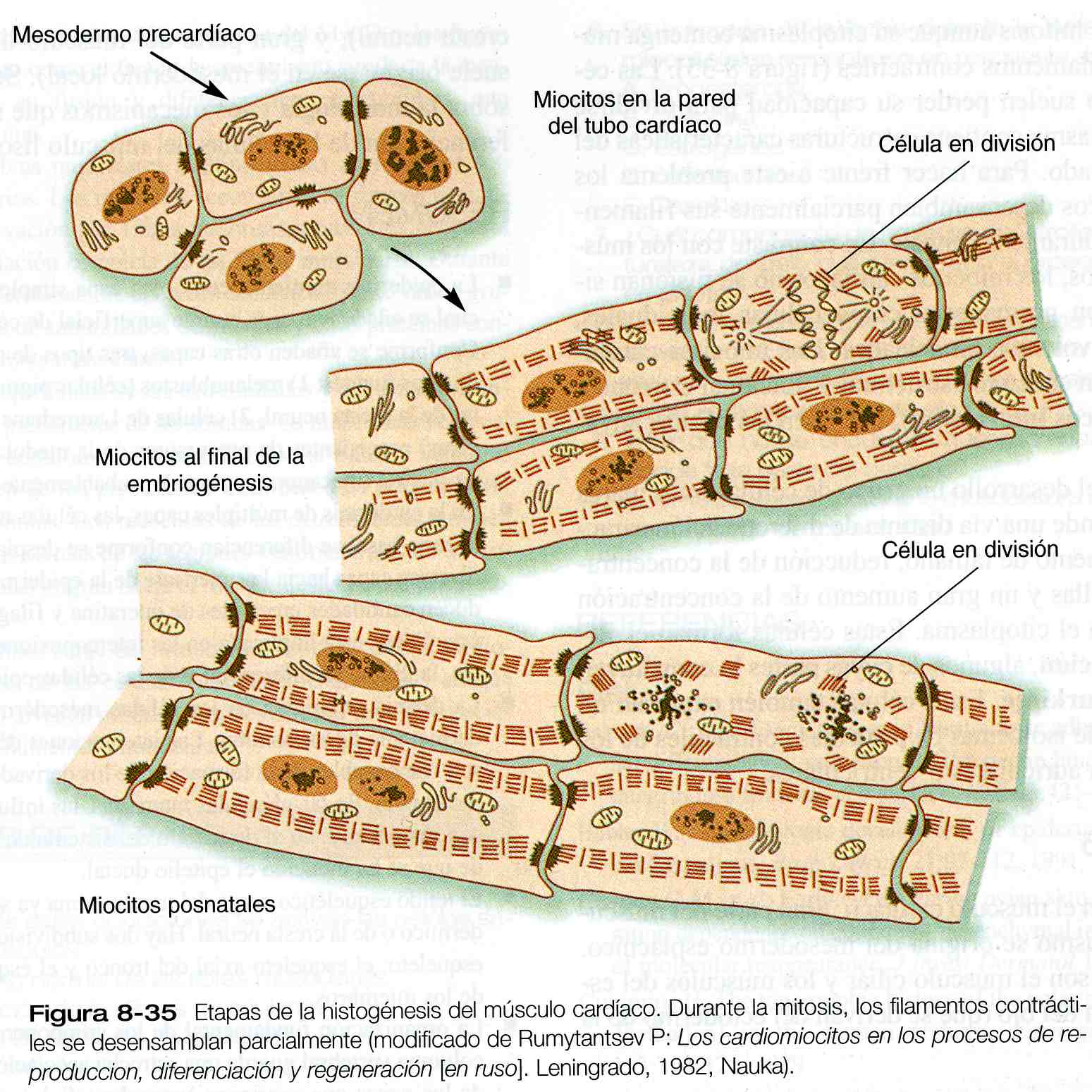{kind=link}