Capítulo 1
PREPARACIÓN PARA LA GESTACIÓN
FASE 1: ORIGEN Y MIGRACIÓN DE CÉLULAS GERMINALES
FASE 2: AUMENTO DEL NÚMERO DE CÉLULAS GERMINALES POR MITOSIS
FASE 3: REDUCCIÓN DEL NÚMERO DE CROMOSOMAS DURANTE LA MEIOSIS
ETAPAS DE LA MEIOSIS
MEIOSIS EN LAS CÉLULAS FEMENINAS
CORRELACIÓN CLÍNICA
MEIOSIS EN LAS CÉLULAS GERMINALES MASCULINAS
FASE 4: MADURACIÓN ESTRUCTURAL Y FUNCIONAL FINAL DE LOS ÓVULOS Y LOS ESPERMATOZOIDES
OVOGÉNESIS
ESPERMATOGÉNESIS
PREPARACIÓN DEL TRACTO REPRODUCTOR FEMENINO PARA EL EMBARAZO
OVARIOS Y TROMPAS UTERINAS O DE FALOPIO
CONTROL HORMONAL DEL CICLO REPRODUCTOR FEMENINO
CONTROL HIPOTALÁMICO
HIPÓFISIS (GLÁNDULA PITUITARIA)
OVARIOS Y PLACENTA
TEJIDOS DIANA DEL TRACTO REPRODUCTOR
INTERACCIONES HORMONALES CON LOS TEJIDOS DURANTE LOS CICLOS REPRODUCTORES FEMENINO
INTERACCIONES HORMONALES QUE INTERVIENEN EN LA REPRODUCCIÓN EN ELVARÓN
CORRELACIÓN CLÍNICA 1-2 Cómo determinar la fecha del embarazo
PREGUNTAS DE REPASO.
El embarazo humano comienza con la fusión de un huevo y un espermatozoide, pero hay una gran cantidad de preparativos que preceden a este acontecimiento. Primero, las células sexuales femeninas y masculinas deben pasar por una larga serie de cambios (gametogénesis) que las convierten desde los puntos de vista genético y fenotípico en gametos maduros, que son capaces de tomar parte en el proceso de la fecundación. A continuación, los gametos deben liberarse de las gónadas y encontrar su camino hacia la parte lateral de la trompa del Falopio, donde normalmente tiene lugar la fecundación. Por último, el huevo fecundado, cuyo nombre apropiado ahora es el de embrión, ha de entrar en el útero, donde se sumerge en la capa de revestimiento interno del útero (Implantación) para ser nutrido por la madre. Todos estos sucesos implican las interacciones entre los gametos o el embrión y el organismo adulto en el cual están alojados; la mayoría de esas interacciones están mediadas o bajo la influencia de las hormonas de los padres. Este capítulo se centra en la gametogénesis y en las modificaciones hormonales del organismo que permiten que se produzca la reproducción.
La gametogénesis suele dividirse en cuatro fases: 1) el origen extraembrionario de las células germinales y su migración hacia las gónadas, 2) un aumento en el número de células germinales por mitosis, 3) una reducción del número de cromosomas en virtud de la meiosis y 4) la maduración estructural y funcional de los óvulos y espermatozoides. La primera fase de la gametogénesis es idéntica en los machos y en las hembras, mientras que existen claras diferencias entre los patrones de los dos sexos para las últimas tres fases.
FASE 1: ORIGEN Y MIGRACIÓN DE CÉLULAS GERMINALES
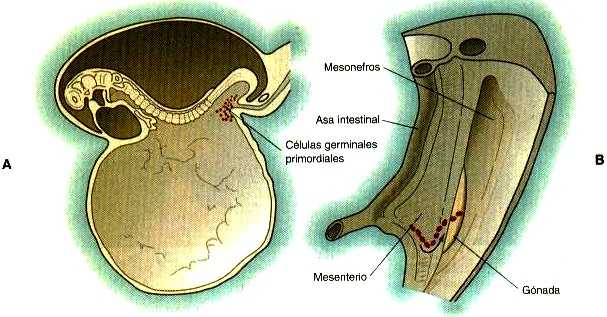
Las células germinales primordiales, los precursores de los gametos que son reconocibles más precozmente, tienen su origen fuera de las gónadas y migran hacia ellas durante el desarrollo embrionario temprano. Las células germinales primordiales humanas se reconocen fácilmente 24 días después de la fecundación en la hoja endodérmica del saco vitelino (Figura 1_1A) gracias a su gran tamaño y su elevado contenido de la enzima fosfatasa alcalina. En el ratón se han detectado de modo aún más precoz (Pág. 377). Las células germinales salen del saco vitelino y pasan al epitelio del intestino posterior; desde allí migran a través del mesenterio dorsal hasta alcanzar los primordios de las gónadas (Figura 1-1B). En el ratón se calcula que unas cien células salen del saco vitelino y, en virtud de la división mitótica (de seis a diez series de división celular), cerca de 4000 células primordiales entran en las gónadas primitivas.
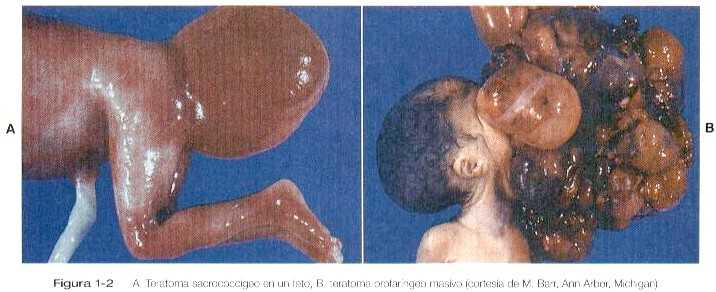
Las células germinales primordiales extraviadas que se alojan en localizaciones extragonadales por lo general mueren, pero si sobreviven pueden convertirse en teratomas. Los teratomas son tumores raros que contienen mezclas aleatorias de tejidos altamente diferenciados como la piel, cabello, cartílago e incluso dientes (Figura 1-2). Se encuentran en el mediastino, la región sacrococcígea y la región oral.
FASE 2: AUMENTO DEL NÚMERO DE CÉLULAS GERMINALES POR MITOSIS
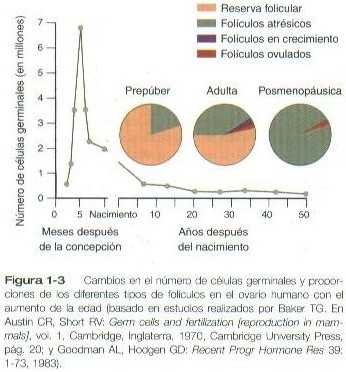
Una vez que llegan a las gónadas, las células germinales primordiales comienzan una fase de proliferación mitótica rápida. En la división mitótica, cada célula germinal produce dos progenies diploides iguales desde el punto de vista genético. A través de varias series de divisiones mitóticas el número de células germinales primordiales aumenta de forma exponencial desde cientos hasta millones. El patrón de la proliferación mitótica difiere apreciablemente entre las células germinales masculinas y las femeninas. Las oogonias, como se denominan a las células germinales femeninas con actividad mitótica, pasan por un período de intensa mitosis en el ovario embrionario desde el segundo hasta el quinto mes de la gestación en los seres humanos. Durante este período la población de células germinales aumenta desde sólo unos pocos miles hasta cerca de siete millones (Figura 1-3). Esta cifra representa el número máximo de células germinales que nunca habrá en los ovarios adultos. Poco tiempo después gran cantidad sufren una degeneración natural denominada atresia. La atresia de las células germinales es una característica continua del panorama histológico del ovario humano hasta la menopausia.
Las espermatogonias del varón, que son similares de las oogonias, siguen un patrón de proliferación mitótica que difiere en gran medida del de la mujer. La mitosis también comienza pronto en el testículo embrionario, pero en contraste con las células germinales femeninas, las masculinas conservan la capacidad de dividirse durante toda la vida. Los túmulos seminíferos de los testículos están tapizados por una población germinativa de espermatogonias. A partir de la pubertad, las subpoblaciones de espermatogonias presentan oleadas periódicas de mitosis. La progenie de estas divisiones entra en meiosis en grupos sincrónicos. Este patrón de mitosis de las espermatogonias continúa durante toda la vida.
FASE 3: REDUCCIÓN DEL NÚMERO DE CROMOSOMAS DURANTE LA MEIOSIS
ETAPAS DE LA MEIOSIS
La importancia biológica de la meiosis en el ser humano es igual que en otras especies. De relevancia primordial son:
1) la reducción del número de cromosomas del número diploide (2n) al número haploide (1n) de manera que puede mantenerse el número de cromosomas propio de la especie de generación en generación, 2) redistribución independiente de los cromosomas materno y paternos para una mejor mezcla de las características genéticas, y 3) distribución adicional de la información genética materna y paterna mediante el proceso de entrecruzamiento durante la primera división meiótica.
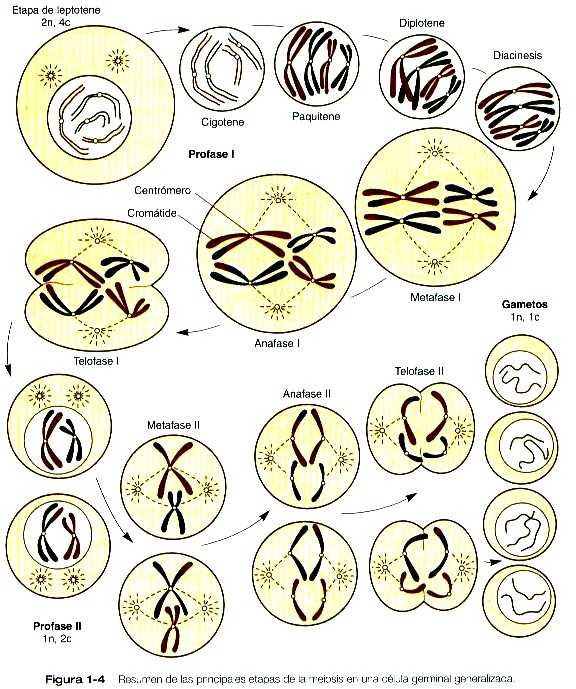
La meiosis comprende dos grupos de divisiones (Figura 1-4). Antes de la primera división meiótica, ya ha tenido lugar la replicación del ácido desoxirribonucleico (DNA), de manera que al comienzo de la meiosis la célula es 2n, 4c (en esta convención, n es el número de la especie y c es la cantidad de DNA en un juego sencillo (n) de cromosomas antes de que haya tenido lugar la replicación del DNA). La célula contiene el número normal de cromosomas (2n), pero como resultado de la replicación su contenido de DNA (4c) es el doble de la cantidad normal (2c).
En la primera división meiótica, que suele llamarse división de reducción, una profase prolongada (figura 1-4) redunda en el apareamiento de los cromosomas homólogos y los entrecruzamientos frecuentes, cuyo resultado es el intercambio de segmentos entre los miembros de los cromosomas pareados. Durante la metafase de la primera división meiótica, los pares de cromosomas (tétradas) se alinean en el plano de metafase (ecuatorial) de manera que en la anafase I un cromosoma de uno de los pares homólogos se desplaza hacia un polo del huso y el otro cromosoma se desplaza hacia el polo opuesto. Ésta es una de las principales diferencias entre la división meiótica y la mitótica. En la anafase de la mitosis, el centrómero entre las cromátides hermanas de cada cromosoma se divide después de que los cromosomas se han alineado en el plano ecuatorial de la metafase, y una cromátide de cada uno de los cromosomas migra hacia cada polo del huso mitótico. El resultado es que las células hijas son genéticamente iguales después de la división mitótica. Cada célula hija de la primera división meiótica contiene el número haploide (1n) de cromosomas, pero cada cromosoma todavía consta de dos cromátides (2c) conectadas por el centrómero. No se requiere una nueva duplicación del DNA cromosómico entre la primera y la segunda división meiótica por que cada célula hija haploide resultante de la primera división meiótica ya contiene cromosomas en estado duplicado.
La segunda división meiótica, denominada división ecuacional, es similar a una mitosis ordinaria, excepto que antes de la división la célula es haploide (1n, 1c). Cuando los cromosomas se alinean a lo largo del plano ecuatorial en la metafase II, en los centrómeros que están entre las cromátides hermanas se dividen, y esto permite que las cromátides hermanas de cada cromosoma migren hacia polos opuestos del huso durante la anafase II. Cada célula hija de la segunda división meiótica es haploide verdadera (1n, 1c).
MEIOSIS EN LAS CÉLULAS FEMENINAS
El período de la meiosis comprende otras actividades celulares además de la redistribución del material cromosómico. A medida que las oogonias entran en la primera división meiótica, durante el período fetal, se denominan oocitos primarios.
La meiosis en la mujer es un proceso muy lento. A medida que los oocitos primarios entran en la etapa de diplotene de la primera división meiótica en los primeros meses que siguen al nacimiento, se produce el primero de los dos bloques del proceso meiótico (figura 1-5). La fase suspendida de la diplotene de la meiosis es el período durante el cual el oocito primario se prepara para las necesidades del embrión.
En los oocitos de los anfibios y de otros vertebrados inferiores, que han de desarrollarse fuera del organismo materno y a menudo en un ambiente hostil, supone una gran ventaja que las primeras etapas del desarrollo ocurran con mucha rapidez para que se alcance lo más pronto posible la etapa de locomoción y alimentación independientes. Estas condiciones requieren una estrategia de almacenamiento de los materiales necesarios para el desarrollo inicial bastante antes de la ovulación y la fecundación por que los procesos sintéticos normales no serían lo suficientemente rápidos como para producir materiales requeridos para el embrión en veloz división. En estas especies, se acumula la yema, se amplifican los genes para producir ácido ribonucleico ribosómico (RNAr) y se sintetizan y almacenan muchos tipos de moléculas de RNA en una forma inactivada para usarlos más adelante.
El sustrato morfológico para la síntesis del ácido ribonucleico (RNA) en el oocito del anfibio está representado por las cromosomas en cepillo que se caracterizan por muchas asas prominentes de DNA extendido en las cuales se sintetizan moléculas de RNA mensajero (RNAm). Los genes amplificados para la producción del RNAr se manifiestan por la presencia de 600 a mil nucléolos en el interior del núcleo. Los oocitos primarios también se preparan para la fecundación mediante la producción de varios miles de gránulos corticales, que son de gran importancia durante este proceso (capítulo 2).
El oocito de los mamíferos se prepara durante un período embrionario inicial, el cual es más prolongado que el de los anfibios y tiene lugar en el ambiente nutricio del tracto reproductor de la madre. Por tanto, no se enfrenta con la necesidad de almacenar una cantidad tan grande de materiales como en el caso de los óvulos de los vertebrados inferiores; en consecuencia, el aumento de la yema es desdeñable. No obstante, algunas evidencias señalan que hay un bajo nivel de amplificación del DNA ribosómico (DNAr) (2 a 3 veces) en los oocitos del ser humano en diplotene, lo cual sugiere que también se requiere algún grado de planificación molecular anticipada para sostener la división inicial en el ser humano.
La presencia de 20 a 40 pequeños micronúcleos (2μm) que contienen RNA (nucléolos en miniatura) en cada núcleo de oocito se correlaciona con la información molecular.
El análisis de la acumulación de RNA en los oocitos de los mamíferos se basa principalmente en los resultados obtenidos en el ratón, en el cual hay algo de acumulación de RNA que comienza durante la etapa de diplotene. Los cromosomas del ser humano en diploteno no parecen estar organizados en una verdadera configuración en cepillo, y parece improbable la síntesis de cantidades masivas de RNA. El oocito de un mamífero en desarrollo (ratón) produce 10 mil veces menos RNAr y mil veces menos RNAm y una acumulación proporcional de RNAr. Estas cantidades de RNA derivado de la madre parecen ser suficientes para llevar el óvulo fecundado a través de la primeras dos divisiones, después de lo cual el genoma embrionario asume el control de los procesos de síntesis macromoleculares.
Puesto que los gránulos corticales desempeñan un papel tan importante en evitar que entren demasiados espermatozoides durante la fecundación en los óvulos humanos (pág. 33), la formación de los gránulos corticales (principalmente el aparato de golgi) sigue siendo una de las funciones de la etapa diplotene que se preserva en los seres humanos. En el oocito de ratón se producen alrededor de 4500 gránulos corticales. En el oocito humano es probable un número poco mayor.a menos que degeneren, todos los oocitos primarios permanecen detenidos en la etapa de diplotene de la meiosis hasta la pubertad. Durante los años fértiles, un número pequeño (10 a 30) de oocitos primarios completan la primera división meiótica con cada ciclo menstrual y comienzan a desarrollarse aún más. Los otros oocitos primarios permanecen detenidos en la etapa de diplotene, algunos durante 50 años.
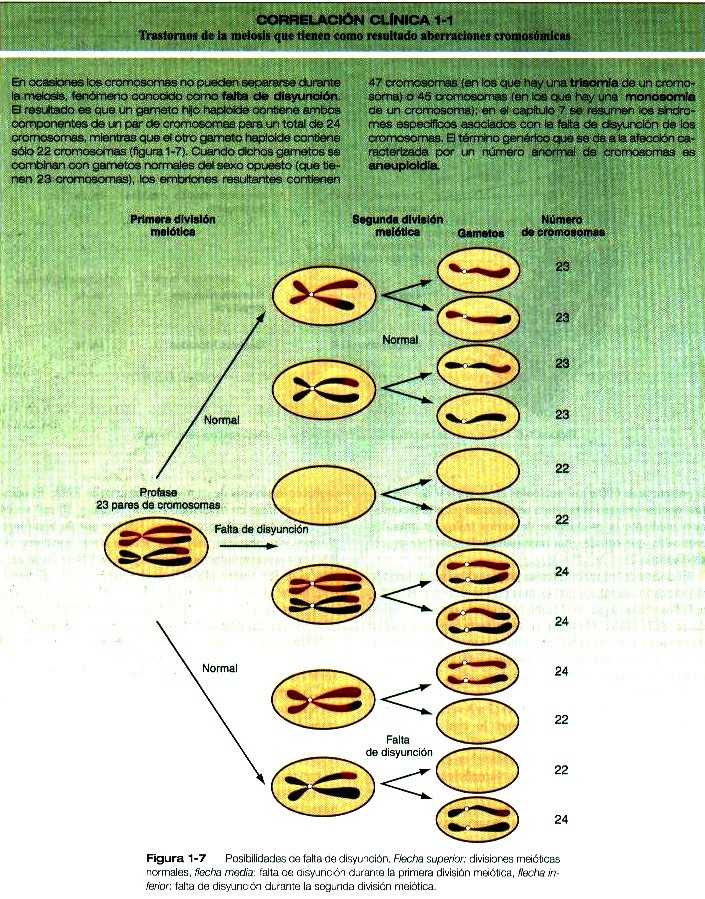
CORRELACIÓN CLÍNICA. Figura 1-7
|
CORRELACIÓN CLÍNICA 1-1 Trastornos de la meiosis que tiene como resultado aberraciones cromosómicas (Continuación) |
|
En otros casos, parte de un cromosoma puede ser translocada a otro cromosoma durante la meiosis, o parte de un cromosoma puede sufrir una deleción. Así mismo, a veces se presentan duplicaciones o inversiones de partes de cromosomas durante la meiosis. Estas afecciones pueden redundar en síndromes similares a los que se observan después de la falta de disyunción de cromosomas completos. En algunas circunstancias (por ejemplo, la fecundación simultánea por dos espermatozoides, falta de separación del segundo cuerpo polar respecto del oocito durante la segunda división meiótica), las células del embrión contienen más de múltiplos del número haploide de cromosomas (poliploidía). Las anomalías cromosómicas son la causa subyacente de un alto porcentaje de abortos espontáneos durante las primeras semanas de gestación. Más del 75% de los abortos espontáneos que se producen antes de la segunda semana y más del 60% de los que suceden durante la primera mitad de la gestación contienen anomalías cromosómicas que van desde trisomías de cromosomas individuales hasta poliploidía global. Aunque la incidencia de las anomalías cromosómicas disminuye con los partos de fetos muertos (mortinatos) que ocurren después del quinto mes de embarazo, se acerca al 6%, una incidencia diez veces mayor de la del 0.5% de los bebés vivos que nacen con anomalías cromosómicas. Cuando se asesora a los padres que han tenido un mortinato o un aborto espontáneo, puede ser útil mencionarles que de esta forma la naturaleza elimina a un embrión destinado a tener graves anomalías. |
Con la terminación de la primera división meiótica poco antes de la ovulación, resultan dos progeies celulares desiguales. Una es una célula grande llamada oocito secundario y la otra célula pequeña que carece de función y se llama primer cuerpo polar (figura1-5). Los oocitos secundarios comienzan la segunda división meiótica, pero de nuevo el proceso de la meiosis se detiene, esta vez en la metafase. El estímulo para la liberación respecto de este bloqueo de la meiosis es la fecundación por un espermatozoide. Los oocitos secundarios no fecundados no pueden terminar la segunda división meiótica. Ésta última es desigual: una de las células hijas de nuevo se ve relegada a convertirse en un segundo cuerpo polar pequeño y sin función. El primer cuerpo polar también puede dividirse durante la segunda división meiótica.
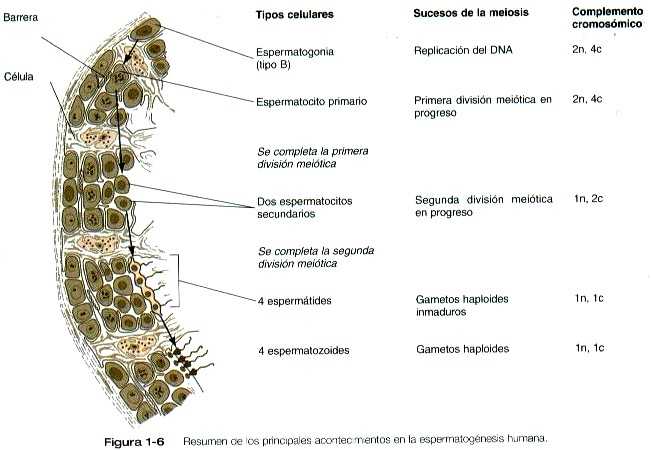
MEIOSIS EN LAS CÉLULAS GERMINALES MASCULINAS
La meiosis en el macho no comienza sino después de la pubertad. En contraste con el oocito primario de la hembra, no todas las espermatogonias entran en la meiosis al mismo tiempo. En realidad, gran cantidad de espermatogonias permanecen en el ciclo mitótico durante gran parte de la vida reproductora del macho. Una vez que la progenie de una espermatogonia ha entrado en el ciclo de la meiosis como espermatocitos primarios, tardan varias semanas en pasar por la primera división meiótica (figura 1-6). El resultado de esta primera división es la formación de espermatocitos secundarios, que entran de inmediato en la segunda división meiótica. Cerca de ocho horas más tarde se ha determinado esta fase y quedan cuatro espermátides haploides (1n, 1c) que forman la progenie de un solo espermatocito primario. La longitud total de la espermatogénesis humana es de 64 días.
Los trastornos que pueden presentarse durante la meiosis y que producen aberraciones cromosómicas se discuten en la Correlación clínica 1-1.
FASE 4: MADURACIÓN ESTRUCTURAL Y FUNCIONAL FINAL DE LOS ÓVULOS Y LOS ESPERMATOZOIDES
OVOGÉNESIS
ESPERMATOGÉNESIS
PREPARACIÓN DEL TRACTO REPRODUCTOR FEMENINO PARA EL EMBARAZO
La estructura y la función del tracto reproductor femenino están bien adaptadas para el transporte de los gametos y el mantenimiento del embrión. Muchas de las características más sutiles de esta adaptación están bajo control hormonal y son cíclicas. En esta sección se repasan en forma breve los aspectos de las estructuras reproductoras femeninas que son de mayor importancia para comprender el transporte de los gametos y el desarrollo embrionario.
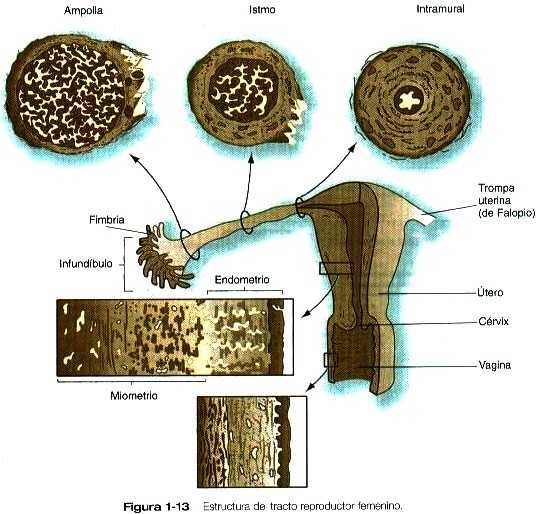
OVARIOS Y TROMPAS UTERINAS O DE FALOPIO
Los ovarios y las trompas uterinas (o de Falopio) forman un complejo funcional dedicado a la producción y el transporte de los óvulos. Los ovarios, órganos con forma de almendra localizados a ambos lados del útero, están ubicados muy cerca de los extremos abiertos con forma de embudo de las trompas uterinas. Numerosas proyecciones similares a dedos llamadas fimbrias (figura 1-13) se proyectan desde el infundíbulo abierto de la trompa hacia el ovario e intervienen en dirigir el óvulo ovulado hacia el interior de la trompa. La trompa de Falopio se caracteriza por un revestimiento interno muy complejo que tiene una gran densidad de pliegues longitudinales prominentes en la región ampular y una estructura más sencilla más cerca del útero. El epitelio interno de las trompas de Falopio contiene una mezcla de células ciliadas que ayudan en el transporte de los gametos y de las células secretoras de un líquido que contribuye al desarrollo inicial del embrión. Las capas musculares lisas a lo largo de la trompa sirven de base para las contracciones peristálticas. La cantidad y la función de muchos de estos componentes están bajo el control cíclico de las hormonas, y el efecto global de estos cambios es facilitar el transporte de los gametos y el óvulo fecundado.
Las principales funciones del útero es recibir y mantener el embrión durante el embarazo y expulsar el feto al término de la gestación. La primera función la efectúa la mucosa uterina (endometrio), y la segunda, la pared muscular (miometrio). Bajo el efecto cíclico de las hormonas, el útero experimenta diversos cambios importantes en el curso de cada ciclo menstrual.
El útero es un órgano en forma de pera con paredes gruesas de músculo liso (miometrio) y un revestimiento mucoso complejo (figura 1-13). El revestimiento mucoso, llamado endometrio, tiene una estructura que cambia todos los días durante el ciclo menstrual. El endometrio puede subdividirse en dos capas: una funcional que se desprende con cada período menstrual o después del parto y una basal que permanece intacta. La estructura general del endometrio consta de: 1) un epitelio superficial columnar, 2) las glándulas uterinas, 3) un tejido conectivo especializado, el estroma, y 4) las arterias espirales que tienen un trayecto en forma de tirabuzón entre la capa basal y la superficie del endometrio. Todas estas estructuras participan en la implantación y la nutrición del embrión.
El orificio de salida del útero es el cérvix. La superficie mucosa del cérvix no es endometrio típico sino que está tachonada con diversas criptas irregulares. El epitelio cervical produce un moco rico en glucoproteínas, cuya composición varía considerablemente durante el ciclo menstrual. Las propiedades físicas variables del moco cervical facilitan o dificultan a los espermatozoides la penetración desde la vagina y el ascenso hacia el útero.
La vagina es un canal que sirve para la cópula y también como canal de parto. Está tapizada por un epitelio escamoso estratificado, pero las células epiteliales contienen depósitos de glucógeno cuya cantidad varía a lo largo del ciclo menstrual. Los productos de degradación del glucógeno contribuyen a la acidez de la secreción vaginal (pH: 4, 3). Al parecer, el pH bajo del tercio superior de la vagina una función bacteriostática e impide que los agentes infecciosos entren en el tracto genital superior por el cérvix y que en último término se diseminen hacia la cavidad peritoneal a través de los extremos abiertos de las trompas de Falopio.
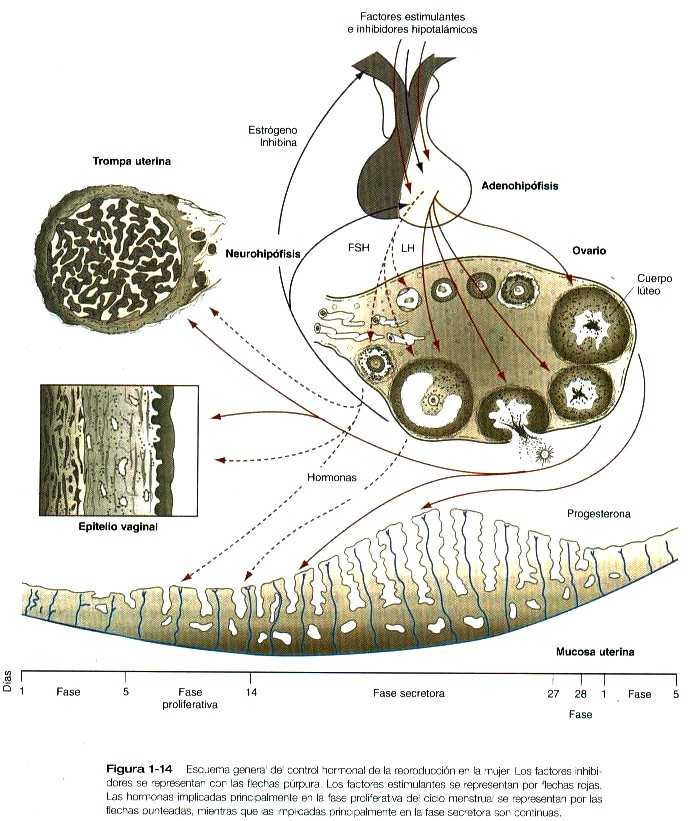
CONTROL HORMONAL DEL CICLO REPRODUCTOR FEMENINO
La reproducción en la mujer está regulada por una compleja serie de interacciones entre las hormonas y los tejidos sobre los cuales influyen. La jerarquía del control cíclico comienza en el hipotálamo situado en el cerebro (figura 1-14). El hipotálamo cerebral influye sobre la producción de hormonas actuando sobre el lóbulo anterior de la glándula hipofisiaria (adenohipofisis). Las hormonas hipofisiarias se dispersan por la sangre a todo el organismo y actúan sobre los ovarios, que a su vez se estimulan a producir sus propias hormonas sexuales esteroideas. Durante la gestación, la placenta ejerce un poderoso efecto sobre la madre mediante la producción de varias hormonas. El nivel final del control hormonal de la reproducción femenina es el ejercido por las hormonas placentarias u ováricas sobre otros órganos reproductores diana (por ejemplo, el útero, las trompas de Falopio, la vagina y las mamas).
CONTROL HIPOTALÁMICO
El primer nivel de control hormonal de la reproducción reside en el hipotálamo. Diversas señales estimulan a las células neurosecretoras del hipotálamo para que produzcan la hormona liberadora de gonadotropinas (GnRH) y factores liberadores de otras hormonas hipofisiarias. Los factores liberadores así como un factor inhibidor son transportados a la adenohipófisis por los vasos sanguíneos del sistema porta hipotalámico-hipofisiario, donde estimulan la secreción de las hormonas adenohipofisiarias (tabla 1-1).
HIPÓFISIS (GLÁNDULA PITUITARIA)
Mediante la producción de sus hormonas en respuesta a la estimulación del hipotálamo, la hipófisis constituye un segundo nivel de control hormonal de la reproducción. La hipófisis consta de dos componentes: la hipófisis anterior (adenohipófisis), una estructura glandular epitelial que produce diversas hormonas en respuesta a factores transportados por el sistema porta hipotalámico hipofisiario y la hipófisis posterior (neurohipófisis), una estructura neural que libera hormonas mediante mecanismos neurosecretorios.
Bajo la influencia de la GnRH y la retroalimentación directa de los niveles sanguíneos de las hormonas esteroideas, la adenohipófisis segrega dos hormonas gonadotrópicas polipeptídicas, la FSH y la LH, procedentes del mismo tipo celular (tabla 1-1). En ausencia de un factor inhibitorio (dopamina) procedente del hipotálamo, la hipófisis anterior también produce la prolactina, que actúa sobre la glándula mamaria.
La única hormona procedente de la neurohipófisis que contiene una participación directa en la reproducción es la oxitocina, un oligopéptido implicado en el trabajo del parto y en el estímulo para la bajada de leche de la glándula mamaria en las mujeres lactantes.
OVARIOS Y PLACENTA
Los ovarios y, durante el embarazo, la placenta constituyen el tercer nivel de control hormonal. En respuesta a los niveles sanguíneos de las hormonas de la adenohipófisis, las células de la granulosa de los folículos ováricos convierten los andrógenos (androtenediosa y testosterona) sintetizados por la teca interna en estrógenos (principalmente estrona y diez veces más potente 17β-estradiol), que luego pasan a la circulación sanguínea. Después de la progesterona es el principal producto secretorio del folículo luego de su conversión en el cuerpo lúteo (capítulo 2). Hacia el final del embarazo, la placenta completa la producción de la hormona esteroide ovárico mediante la síntesis de sus propios estrógenos y progestágenos. También produce dos hormonas polipeptídicas (tabla 1-1). La gonadotropina coriónica humana (HCG) actúa en el ovario manteniendo la actividad del cuerpo lúteo durante el embarazo. El lactógeno placentario humano (somatomatropina) actúa sobre el cuerpo amarillo; también promueve el desarrollo de la mama por que potencia los efectos de los estrógenos y la progesterona y estimula la síntesis de los constituyentes de la leche.
TEJIDOS DIANA DEL TRACTO REPRODUCTOR
El último nivel de jerarquía del control hormonal de la reproducción lo constituyen los tejidos diana, que se preparan tanto desde el punto de vista estructural como funcional para el transporte de los gametos o el embarazo en respuesta a la unión de las hormonas ováricas o placentarias a los receptores celulares específicos. Algunos de los ejemplos más prominentes del efecto de las hormonas sobre los tejidos diana son los cambios en el número de células ciliadas y en la actividad de la musculatura lisa uterina, así como las transformaciones profundas en el revestimiento endometrial del útero y los cambios cíclicos de los tejidos glandulares de la mama. Todos ellos se describen en mayor detalle más adelante.
Un principio general que se conoció hace ya algún tiempo es la efectividad de estimular primero los tejidos reproductores diana con estrógenos para que la progesterona pueda ejercer completamente sus efectos. Los estrógenos inducen a las células diana a producir grandes cantidades de receptores de progesterona, que deben estar en su lugar para que la progesterona pueda actuar sobre estas mismas células.
|
Hormona
|
Naturaleza Química |
Función |
|
HIPOTÁLAMO Hormona liberadora de las gonadotropinas (GnRH, LHRH) Factor inhibidor de la prolactina |
Decapéptido
Dopamina |
Estimula la liberación de la LH y la FSH por la hipófisis anterior
Inhibe la liberación de la prolactina por la hipófisis anterior |
|
HIPÓFISIS ANTERIOR Hormona folículo-estimulante (FSH)
Hormona luteinizante (LH)
Prolactina |
Glucoproteína (subunidades α y ß)
Glucoproteína (subunidades α y ß)
Polipéptido de cadena sencilla (198 aminoácidos) |
Varón: estimula las células de Sertoli para que produzcan proteína fijadora de andrógenos Mujer: estimula las células del folículo para que produzcan estrógenos
Varón: estimula las células de Leydig para que segreguen testosterona Mujer: estimula las células del folículo y al cuerpo amarillo para que produzcan progesterona
Promueve la lactancia |
|
HIPÓFISIS POSTERIOR Oxitocina |
Oligopéptido (PM, -1100) |
Estimula la eyección de la leche de la glándula mamaria. |
|
OVARIO Estrógenos
Progesterona
Testosterona
Inhibina
Activina |
Esteroide
Esteroide
Esteroide
Proteína (PM, -32.000)
Proteína (PM, -28.000) |
Tiene múltiples efectos en el tracto reproductor, la mama, la grasa corporal y el crecimiento óseo Tiene múltiples efectos en el tracto reproductor y el desarrollo de la mama Inhibe la secreción de FSH, tiene efectos locales en los ovarios
Estimula la proliferación de las células de la granulosa
|
|
TESTÍCULOS Testosterona
Inhibina |
Esteroide
Proteína (PM,-32.000) |
Tiene múltiples efectos en el tracto reproductor del varón, el crecimiento del vello y otras características sexuales secundaria Inhibe la secreción de la FSH, tiene efectos locales en los testículos |
|
PLACENTA Estrógenos
Progesterona
Gonadotropina coriónica humana (HCG)
Lactógeno placentario humano (somatomatropina) |
Esteroide
Esteroide
Glucoproteína (PM, -30.000)
Polipéptido (PM, -20.000) |
Tiene las mismas funciones de los estrógenos ováricos. Tiene las mismas funciones de la progesterona ovárica Mantiene la actividad del cuerpo amarillo durante el embarazo Promueve el desarrollo de la mama durante la gestación |
INTERACCIONES HORMONALES CON LOS TEJIDOS DURANTE LOS CICLOS REPRODUCTORES FEMENINO
Todos los tejidos del tracto reproductor femenino están bajo la influencia de las hormonas de la reproducción. En respuesta al ambiente hormonal del organismo, ellos experimentan modificaciones cíclicas que mejoran las posibilidades de éxito de la reproducción.
Para comprender las interacciones hormonales y las respuestas tisulares durante el ciclo reproductor femenino, es imprescindible conocer los cambios que experimentan los ovarios. En respuesta a la FSH y la LH segregadas por la hipófisis inmediatamente antes del período menstrual y durante el mismo, una cohorte de folículos ováricos secundarios comienza a madurar y segregar 17β-estradiol. Para el tiempo de la ovulación todos estos folículos, excepto uno, han sufrido atresia, y su contribución principal ha sido la de producir parte del suministro de estrógenos necesarios para prepara el organismo para la ovulación y transporte de los gametos.
Durante la fase preovulatoria o proliferativa (días 5 a 14) del ciclo menstrual, los estrógenos producidos por el ovario actúan sobre los tejidos reproductores femeninos (figura 1-14). La capa de revestimiento uterino vuelve a epitelizarse después del período menstrual que acaba de terminar. Luego, bajo la influencia de los estrógenos, el estroma endometrial experimenta un engrosamiento progresivo, las glándulas uterinas se alargan y las arterias espirales comienzan a crecer hacia la superficie del endometrio. Las glándulas mucosas del cérvix segregan un moco rico en glucoproteínas pero relativamente acuoso, que facilita el paso de los espermatozoides a través del canal cervical. A medida que avanza la fase proliferativa, un porcentaje mayor de las células epiteliales que tapizan las trompas uterinas se vuelven ciliadas y aumenta la actividad de la musculatura lisa de la trompa. En los días que preceden a la ovulación, las fimbrias de la trompa uterina se contraen y aproximan a la ampolla de la trompa a los ovarios.
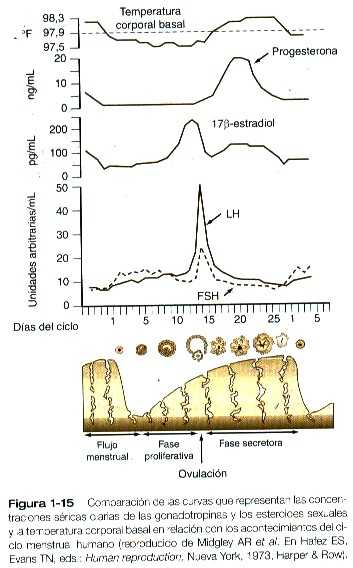
Hacia el final del período proliferativo, un marcado incremento en los niveles del estradiol segregado por el folículo ovárico en desarrollo actúa sobre el sistema hipotálamo-hipofisiario; allí causa un aumento de la capacidad de respuesta de la adenohipófisis a la GnRH y un pico de la secreción hipotalámica de la GnRH. Aproximadamente 24 horas después de que el nivel de 17β-estradiol alcanza su máximo en la sangre, la hipófisis envía un pico preovulatorio de LH y FSH a la circulación sanguínea (Figura 1-15). El pico de LH no es un aumento continuo de la secreción de la gonadotropina, sino que constituye una serie de pulsos agudos y momentáneos de secreción que parece responder a un mecanismo temporizador hipotalámico.
El pico de LH conduce a la ovulación, y el folículo de Graaf se transforma en cuerpo lúteo (cuerpo amarillo). La lámina basal que circunda a la granulosa del folículo se rompe y permite que los vasos sanguíneos de la granulosa crezcan dentro de la capa de las células de la granulosa. Mediante procesos de proliferación e hipertrofia, las células de la granulosa experimentan importantes cambios estructurales y bioquímicos y ahora producen progesterona como principal producto de secreción. El cuerpo lúteo conserva la capacidad de producir una pequeña cantidad de estrógenos. Después de la ovulación se dice que el ciclo menstrual, que ahora está bajo el dominio de la secreción de la progesterona, está en la fase secretora (días 14 a 28 del ciclo menstrual).
Después del pico de LH y con el aumento de la concentración de progesterona en la sangre, la temperatura corporal basal se eleva (figura 1-15). A causa de la relación entre la elevación de la temperatura basal y el momento de la ovulación, el registro preciso de la temperatura constituye la base del método del ritmo para el control de la natalidad.
Alrededor del momento de la ovulación, la presencia combinada de estrógenos progesterona en la sangre hace que la trompa uterina experimente una serie rítmica de contracciones musculares destinadas a favorecer el transporte del óvulo. La progesterona estimula a alas células epiteliales de la trompa para que segreguen líquidos que aportan los nutrientes al cigoto en división. Más adelante en la fase secretora, los niveles elevados de progesterona inducen la regresión de algunas células ciliadas del epitelio tubárico.
En el útero, la progesterona prepara al endometrio previamente estimulado por los estrógenos para la implantación del embrión. El endometrio, que se ha engrosado bajo la influencia de los estrógenos durante la fase proliferativa, sufre otros cambios. Las glándulas uterinas rectas comienzan a entorcharse y acumulan glucógeno y otros productos secretorios en el epitelio. Las arterias espirales crecen más hacia la superficie del endometrio, pero la mitosis de las células epiteliales endometriales disminuye. Mediante la acción de la progesterona, el moco cervical se vuelve muy viscoso y actúa como bloqueo de protección, que inhibe el paso de materiales hacia o desde el útero. Durante la fase secretora, el epitelio de la vagina se adelgaza.
En las glándulas mamarias, la progesterona impulsa el desarrollo previamente estimulado por los estrógenos, de los componentes secretorios, y ocasiona retención de agua en los tejidos. No obstante, el desarrollo más intenso del aparato de lactación espera su estimulación por hormonas placentarias.
A mitad de su camino en la fase secretora del ciclo menstrual, el epitelio tubárico ya ha experimentado considerable regresión desde su máximo a la mitad de su ciclo, mientras que el endometrio uterino está en el máximo de su preparación para recibir un embrión en división. Si no tiene lugar el embarazo, una serie de interacciones hormonales lleva el Ciclo menstrual a su fin.
|
Parámetro
|
Células de la granulosa (femeninas) |
Células de Sertoli (masculinas) |
Células de la teca (femeninas) |
Células de Leydig (masculinas) |
|
Origen Principales receptores Principales productos de secreción |
Rete ovari FSH Estrógenos, progesterona, inhibina |
Rete testis FSH Estrógenos, inhibina, proteína fijadora de andrógenos |
Mesénquima estromal LH Andrógenos
|
Mesénquima estromal LH Testosterona
|
Uno de los mecanismos iniciales de retroalimentación es la producción de la proteína inhibina por parte de las células de la granulosa. La inhibina se transporta por el torrente circulatorio hacia la hipófisis anterior, donde inhibe directamente la secreción de gonadotropinas, en especial la FSH. A través de mecanismos que todavía no son claros, también se reduce la secreción de la LH. Esta inhibición redunda en la regresión del cuerpo lúteo y en la marcad reducción de la secreción de progesterona por parte del ovario.
Algunas de las consecuencias principales de la regresión del cuerpo lúteo son la infiltración del estroma endometrial por leucocitos, la pérdida de líquido intersticial y la constricción espasmódica y la rotura de las arterias espirales que causa isquemia local. La isquemia produce hemorragia local y la pérdida de áreas del endometrio. Estos cambios inician la menstruación (por convención, constituye los días 1 a 5 del ciclo menstrual). En los días que siguen, toda la capa funcional del endometrio se desprende en pequeños trozos, junto con la pérdida acompañante de unos 30 mL de sangre. En el momento en que termina la menstruación, y como base para la curación y la reconstrucción del endometrio durante el siguiente período proliferativo sólo queda una pequeña capa endometrial denudada intercalada con el epitelio basal de las glándulas uterinas.
INTERACCIONES HORMONALES QUE INTERVIENEN EN LA REPRODUCCIÓN EN EL VARÓN
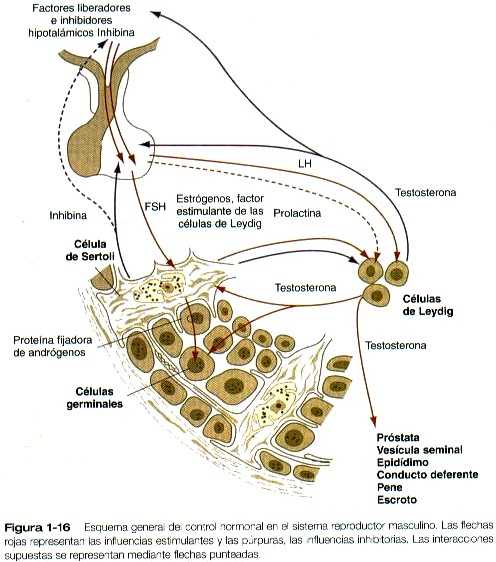
Además del paralelismo que existe entre ciertas estructuras del testículo y el ovario, también hay estrechos paralelismos entre las interacciones hormonales que intervienen en la reproducción de machos y hembras. Los más importantes son los que hay entre las células de la granulosa del folículo ovárico y las células de Sertoli del túbulo seminífero del testículo y entre las células de la teca del ovario y las células de Leydig del testículo (tabla 1-2).
La secreción hipotalámica de la GnRH estimula a la hipófisis anterior para que secrete FSH y LH. Ésta última se fija a los casi 20 mil receptores de LH que hay en la superficie de cada célula de Leydig (intersticial), y a través de una cascada de segundos mensajeros que llevan consigo al monofosfato cíclico de adenosina y a la fosforilación de las proteínas, la LH estimula la síntesis de testosterona a partir del colesterol. La testosterona se libera en la sangre y llega a las células de Sertoli y al resto del organismo, donde afecta a diversos tejidos sexuales secundarios, a menudo después de haberse convertido localmente en dihidrotestosterona.
|
CUADRO 1-2 Principales funciones de las células de Sertoli |
|
Mantenimiento de la barrera hematotesticular Secreción de líquido tubular (10 a 20 μL/g de testículo/h) Secreción de proteína fijadora de estrógenos Secreción de inhibina Secreción de una amplia variedad de otras proteínas (por ejemplo, factores de crecimiento, transferrina, proteína fijadora retiniana, proteínas fijadoras de metales) Mantenimiento y coordinación de la espermatogénesis Fagocitosis de los cuerpos residuales de las células espermáticas. |
|
CASO CLÏNICO |
|
Una mujer de 33 años de edad se ha sometido a la extirpación de ambos ovarios a causa de grandes quistes ováricos bilaterales. Al siguiente año se encuentra en una prolongada expedición en el norte de Canadá; su canoa se hunde y sus medicamentos de sustitución hormonal van al fondo del lago. Pasan más de seis semanas antes que ella pueda volver a conseguir un nuevo lote de medicamentos. ¿Cuál de los siguientes sería el aspecto menos afectado por la pérdida de los medicamentos de esta mujer?
|
Las células de Sertoli son estimuladas por la FSH de la hipófisis a través de los receptores superficiales de FSH y por la testosterona a partir de las células de Leydig a través de receptores citoplasmáticos. Después de ser estimuladas por la FSH, las células de Sertoli convierten parte de la testosterona en estrógenos (igual que lo hacen las células de la granulosa del ovario). Parte de estos estrógenos se difunde hacia las células de Leyding junto con un factor estimulante de las células de Leydig, que se procede en las células de Sertoli y alcanza a las de Leydig mediante un modo de secreción paracrino (no transportado por la sangre) (figura 1-16). La célula de Sertoli estimulada por la FSH produce la proteína fijadora de andrógenos, se fija la testosterona y se transporta hacia el compartimiento del líquido del túbulo seminífero donde ejerce una fuerte influencia sobre el curso de la espermatogénesis. A semejanza de las células de la granulosa que son sus homólogas en el ovario, las células de Sertoli estimuladas por las hormonas producen inhibina mediante un mecanismo de retroalimentación negativa inhibe la secreción de la FSH. Además de la inhibina y la proteína fijadora de andrógenos, las células de Sertoli tienen muchas otras funciones, las más importantes de las cuales se resumen en el cuadro 1-2.
CORRELACIÓN CLÍNICA 1-2 Cómo determinar la fecha del embarazo
Hay dos sistemas diferentes para fechar el embarazo. Uno, usado por los embriólogos, fecha de gestación desde el momento de la fecundación (edad de fecundación), de manera que un embrión de seis semanas (42 días) desde el día de la fecundación. El otro sistema, usado por los obstetras y muchos otros clínicos, fecha de embarazo a partir del último período menstrual de la mujer (fecha de la última regla; edad menstrual) por que es un punto conveniente de referencia desde el punto de vista de la historia obtenida del paciente. La edad menstrual de un embrión humano es de dos semanas más larga que la edad de fecundación por que suelen transcurrir dos semanas desde el comienzo de la última menstruación hasta la fecundación. Así pues, a un embrión con una edad de fecundación de seis semanas se le asigna una edad menstrual de ocho semanas y la duración típica del embarazo es de 38 semanas de edad de fecundación y 40 semanas de edad menstrual (figuras 1-17 y 1-16).
Por razones clínicas válidas, los obstetras subdividen la gestación en tres trimestres iguales, mientras que los embriólogos dividen la gestación en períodos desiguales correspondientes a acontecimientos importantes del desarrollo.
0-3 semanas Desarrollo Precoz (división, gastrulación)
4-8 semanas Período de organogénesis embrionaria
9-38 semanas Período Fetal
El reconocimiento de la existencia de diferentes sistemas para fechar el embarazo es esencial. En un pleito ante los tribunales que implique una demanda legal a propósito de un defecto de nacimiento, una confusión de dos semanas sobre la fecha del embarazo puede ser factor decisivo entre ganar o perder el pleito. En un caso de labio leporino o paladar hendido (pág. 308), la diferencia en el desarrollo de la cara entre las seis y las ocho semanas (Figura 13-6) haría imposibles algunas circunstancias. Por ejemplo, una noxa sufrida a las seis semanas podría ser la causa potencial de un labio leporino, mientras que a las ocho semanas los labios ya se han formado, de manera que en ese momento la aparición de esta malformación sería improbable.
PREGUNTAS DE REPASO
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}