5.3. GAMETOGÉNESIS Y
FECUNDACIÓN
La Gameto génesis el proceso por
el cual se forman los gametos en las gónadas masculinas y femeninas,
testículos y ovarios respectivamente. Durante este proceso ocurren cambios
morfológicos y cromosómicos que están regulados en el interior de las gónadas
en desarrollo. Las gónadas durante la órgano génesis se diferencian de acuerdo
al sexo cromosómico en condiciones normales, proceso conocido como
diferenciación sexual.
Ovogénesis
Las células sexuales femeninas se
originan de las Células Germinativas Primordiales (CGP), que son producidas
por el Epiblasto en la 2a semana del desarrollo. En la 3a semana se
mueven a la pared del saco vitelino, en la 4a semana comienzan la migración
hacia las gónadas y en la 5a semana llegan a las gónadas donde se forman las
células sexuales femeninas u ovocitos. La gónada femenina es el ovario y el
proceso de formación de las células sexuales femeninas u ovocitos recibe el
nombre de Ovogénesis.
En el ovario las CGP se
diferencian en Ovogonias, las cuales tienen sucesivas divisiones mitóticas
(Fase de proliferación). Hacia el final del tercer mes, las ovogonias
se organizan en grupos rodeados por una capa de células epiteliales planas
originadas del epitelio superficial que recubre al ovario. La mayoría de las
ovogonias continúan dividiéndose pero algunas de ellas se diferencian en
Ovocitos primarios, que tienen un mayor tamaño (Fase de crecimiento) que
inmediatamente comienzan la profase de la primera división meiótica, luego de
la duplicación del ADN (Fase de maduración).
Hacia el 5o mes del
desarrollo prenatal las CGP alcanzan el número máximo, alrededor de 7,
000,000. En este momento comienza la muerte celular de las CGP, de
ovogonias y ovocitos primarios. Sólo persisten las ovogonias próximas a la
superficie y ovocitos primarios que han quedado rodeados de una capa de
células epiteliales planas, denominándose Folículos primordiales.
La ovogénesis ocurre durante la
Folículo génesis. Los folículos pueden encontrarse en reposo, crecimiento,
degeneración o dispuestos para la ovulación. Existen diferentes tipos de
folículos de acuerdo a su grado de madurez: Primordial, Primario,
Secundario antral y Terciario o Folículo vesicular o de De
Graaf.
-De
utrisque sexus organis generatione inservientibus tractatus duo.
Genevae, J.A. Chouët et D. Ritter, 1699. Reignier De Graaf_grab.jpg
1641-1673-.
En el momento del nacimiento los
folículos que se encuentran el ovario son los primordiales y la
ovogénesis se detiene en la profase de la Meiosis I. Comienza una etapa de
reposo que se conoce como Periodo de Diploteno o Dictioteno, que se
caracteriza porque la célula presenta la cromatina en forma de red de encaje.
La meiosis se cree es inhibida por una sustancia de maduración del ovocito (OMI)
secretada por las células foliculares. Esta etapa de reposo dura hasta la
pubertad, en la cual sólo existen alrededor de 400 000 ovocitos y
sólo aproximadamente 500 llegarán a ser ovulados. Algunos ovocitos permanecen
en el estado de Diploteno por 40 años o más, lo que se asocia con defectos de
desarrollo.
En la pubertad cada mes varios
folículos primordiales comienzan a madurar, pero sólo uno alcanza la madurez
total, convirtiéndose en folículo primario, en el que las células
foliculares han formado un epitelio estratificado cúbico que recibe el
nombre de granulosa. En este folículo primario, entre la membrana plasmática
del ovocito y la granulosa, se encuentra una capa de glicoproteinas denominada
zona pelúcida. Este folículo primario da lugar al folículo secundario y
luego al terciario, que reanuda la Meiosis I, al final de la cual se
obtiene una célula con abundante citoplasma, el ovocito secundario, y una
célula pequeña, el primer corpúsculo polar. Antes de terminar la segunda
meiosis, en la metafase de la Meiosis II, el ovocito secundario es ovulado y
si no es fecundado degenera en aproximadamente 24 horas.
Si el ovocito ovulado es
fecundado termina la segunda meiosis; así, luego de concluida la Meiosis
II se obtiene una célula grande y con abundante citoplasma, viable, y tres
corpúsculos polares pequeños y con escaso citoplasma que normalmente
degeneran, (El primer corpúsculo polar puede dividirse también en dos
células). Simultáneamente con estos cambios morfológicos durante la meiosis
ocurren los cromosómicos. El ovocito primario tiene 46 cromosomas por lo que
es una célula diploide (2n). Cuando comienza el proceso de maduración, que
duplicaba su ADN, tiene 46 cromosomas dobles, cuando termina la primera
división tiene 23 dobles y sólo cuando termina la segunda división
cuenta con un juego haploide de cromosomas, sólo 23. De éstos últimos 22 son
autosomas y uno sexual X (22 + X) (Fig.5.1).
Durante este proceso los cambios
morfológicos y cromosómicos ocurren de forma simultánea o sea al mismo tiempo.
La ovogénesis se detiene en una
etapa avanzada de la vida de la mujer producto de la depresión hormonal, no es
un proceso continuo, el ciclo reproductor femenino solo se extiende hasta los
47 a 52 años de vida. Además va acompañado de otros cambios biológicos en la
mujer.
Regulación
de la ovogénesis:
Para la migración de las células germinativas primordiales (CGP), precursoras
de las células sexuales; es muy importante la fibronectina. Se ha demostrado
que en el ratón se secreta en las crestas gonadales de forma difusible TGF-B1,
que es capaz de atraer a las CGP. Estas células totí potenciales conservan
esta cualidad por el factor de trascripción nuclear Oct-4. Este factor,
después de la fecundación, se expresa en los primeros clivajes de les núcleos
de los blastómeros y es restringido por la masa celular interna. La
proliferación de las CGP ocurre por acción del Factor de las células madres.
Este factor es secretado por las células en el transcurso de la migración.
En la mujer la capacidad
reproductora es intermitente o cíclica y está regulada por los esteroides
ováricos que establecen una retroalimentación negativa sobre el Hipotálamo y
las hormonas gonadotrópicas de la Hipófisis, generando un patrón cíclico
característico. El ciclo ovárico y uterino corren paralelamente, comenzando en
la pubertad, se interrumpen durante el embarazo y la lactancia y cesan en la
menopausia. La ovogénesis comienza en el ovario fetal, cuando la hipófisis
funciona moderadamente, pero se reinicia en la pubertad cuando se ha alcanzado
la madurez endocrina necesaria. En la Folículo génesis, algunos de estos
procesos ocurren sin intervención hormonal, mientras que otros están regulados
por una compleja relación entre gonadotropinas, esteroides y factores
ováricos locales. Los detalles de estos procesos se estudiarán al abordar los
sistemas Reproductor y Endocrino.
El ovocito está rodeado por
diversas capas de células de soporte que protegen. Está rodeado de células
foliculares (granulosa), que lo alimentan y le suministran un ambiente
hormonal adecuado, de la membrana basal y de otras capas de células del
estroma que, cuando comienzan a madurar, la teca interna y la externa. La
primera de ellas tiene función secretora principalmente de estrógenos.
La participación de las células
de la granulosa es mediada por factores paracrinos como el GDF9, un miembro de
la familia TGF-B. El folículo también secreta factores de crecimiento y
diferenciación como el TGF-B2, VEGF, y Leptina y FGF2, que permiten que el
ovocito crezca y que los vasos sanguíneos a la región folicular. Para que el
ovocito madure necesita de señales de las gonadotropinas en determinadas
etapas.
La Hormona Estimulante de los
Folículos (FSH) determina que varios de ellos crezcan y proliferen. En los
mamíferos las hormonas controlan e inciden el ciclo uterino (menstrual y
cervical), y en el ciclo ovárico que permite que el organismo este listo para
recibir al embrión en un breve tiempo después que la Ovulación ocurra.
El ovocito primario se para en la
pro fase de la meiosis I, que es un estado equivalente a la fase G2 del ciclo
mitótico. Las células foliculares establecen entre sí uniones comunicantes por
las cuales intercambian pequeñas moléculas y precursores para síntesis de
moléculas mayores. Las células foliculares secretan macromoléculas que
contribuyen a formar la cubierta del ovocito y son incorporadas por este
durante el crecimiento mediante endocitosis o actúan como receptores de su
superficie controlando el patrón espacial y las asimetrías axiales de las
células.
La primera consideración sobre el
desarrollo de los embriones humanos comienza con la regulación por cambios
moleculares y la reorganización celular que ocurre antes de la fertilización.
Durante la fase terminal de la ovogénesis, es decir, en el periodo de la
meiosis que precede a la ovulación, ocurren cambios en el patrón de síntesis
de macromoléculas y modificaciones en la estructura y organización del
citoplasma y la membrana del ovocito. Estos cambios representan la expresión
de un programa sobre el desarrollo que prepara al ovocito para la
fertilización y el futuro posterior.
Existen en la actualidad grupos
de investigación que tratan de averiguar cómo esta programación a nivel
molecular y celular que posee el ovocito es regulada y coordinada en el
estadio pre ovulatorio y de la ovogénesis. Una vez que el ovocito es
fertilizado, el por qué las células embrionarias forman diferentes tejidos y
estos poseen una disposición ordenada en el espacio que de lugar a órganos, es
otro de los aspectos menos comprendidos de los organismos en desarrollo aunque
existen “respuestas provisionales”.
FASES
|
U
T
E
R
O
|
Menstrual |
Proliferativa o folicular |
Progestacional o secretora |
Gravidica |
|
0-4 |
Hasta el día 14 |
Termina día 28 |
Ocurre fecundación |
|
Menstruación |
Crece el espesor de la mucosa de 1-3mm, estimulando por
estrógenos ováricos.
Abundantes mitosis en las glándulas del estroma. Las
células de la mucosa de cúbicas se transforman en cilíndricas altas, también
se alargan las glándulas. El estroma se torna compacto por la abundancia
celular. |
El espesor del endometrio aumenta a 6-7 mm, debido a la
acción de la progesterona y estrógenos provenientes del cuerpo lúteo.
Después de la ovulación aparecen vacuolas sub nucleares llenas de glucógeno
en las glándulas. El estroma continua proliferando y se edematiza lo cual
engruesa la mucosa. Al final de la etapa cambia la irrigación por la caída
de estrógenos y progesterona proveniente del cuerpo lúteo por ultimo ocurre
la menstruación. |
En caso de embarazo el blastocisto se introduce en la
mucosa uterina de 6-9 días después de la ovulación. Hacia el final de la
fase secretora el trofoblasto secreta hCG que estimula al cuerpo lúteo y
este continua secretando sus hormonas y no ocurre la menstruación. El
endometrio se transforma en hiperplásico edematoso y secretor. Las células
del estroma se tornan claras, grandes y ricas en glucógeno son las células
deciduales esta es llamada transformación decidua. |
|
OVARIO |
Folicular |
Formación de cuerpo amarillo |
Cuerpo amarillo gravídico |
|
Folículo génesis, crecimiento folicular junto con la ovogénesis acción de la
FSH de la hipófisis. |
Secreta progesterona y estrógenos continua secreción de LH de la Hipófisis. |
Continua secretando las hormonas anteriores por estimulo de la HCG |
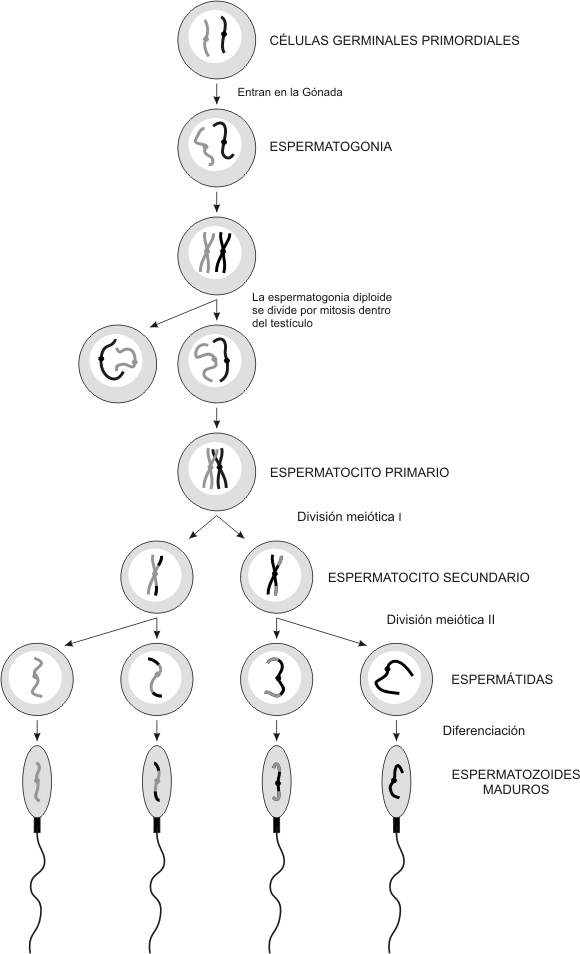
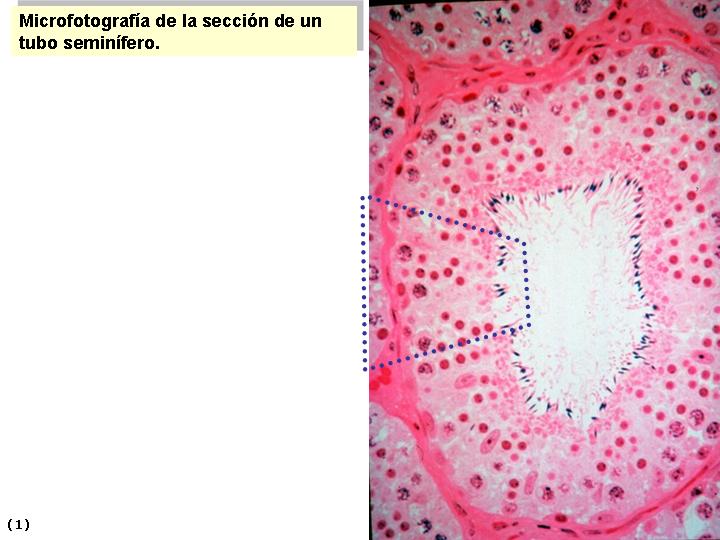
ESPERMATOGENESIS
Las células sexuales masculinas o espermatozoides se forman también de
las CGP en las gónadas masculinas o testículos, mediante el proceso de
Espermatogénesis. El proceso de la Espermatogénesis comienza en la pubertad,
por la necesidad de la madurez endocrina, pero es continuo hasta la muerte
del individuo. En el testículo del recién nacido se pueden observar las células
germinales, pálidas, redondeadas y rodeadas de células de sostén derivadas de
las células del epitelio superficial de las gónadas; ellas formarán las células
de
Sertoli.
Con la madurez del sistema reproductor masculino en la pubertad en vez de
cordones testiculares se pueden observar Túbulos seminíferos. Las CGP dan
origen a las Espermatogonias que son de dos tipos, las A y las B. Las primeras
continúan dividiéndose por mitosis para formar una reserva continua de células
madres (Stem cells). Algunas de ellas dejan de ser células madres, tienen
divisiones sucesivas y dan origen a generaciones de espermatogonias, aumentando
la diferenciación a medida que se dividen; la última división de estas células
forma las Espermatogonias de tipo B, que también se dividen por Mitosis, (Fase
de proliferación), y que formarán células de mayor tamaño, los Espermatocitos
Primarios (Espermatocitos
I),
(Fase de crecimiento). Estas últimas células duplican
su ADN (Fase de maduración) y comienzan la Meiosis I, con una Profase
prolongada de 22 días. A continuación termina rápidamente la Meiosis I y se
forman entonces los Espermatocitos Secundarios (Espermatocitos
II).
En estas células ocurre la Meiosis
II
y se forman entonces células haploides llamadas
Espermátidas.
Desde la formación de las Espermatogonias hasta la de las Espermátidas la
citocinesis es incompleta y las células forman un sincitio por la comunicación
entre ellas a través de puentes citoplasmáticos de un milímetro de diámetro.
Iones y moléculas pasan a través de esos puentes por lo que todas las células
.maduran sincrónicamente. Todas las células son morfológicamente iguales y
durante todo su desarrollo se encuentran "abrazadas" por las células de Sertoli.
Los núcleos de los Espermatocitos transcriben algunos genes cuyos productos
son usados más tarde para formar el axonema y el acrosoma.
Las células de Sertoli tienen varias funciones:
1. Estimulan las CGP para convertirse en espermatozoides.
2. Secretan la hormona antiMulleriana.
3. Estimulan la migración de células somáticas que se encuentran junto a la
gónada para formar tejido conjuntivo imprescindible para la producción normal
de espermatozoides.
4. Inducen a otras células somáticas a que se transformen en células de Leydig,
secretoras de Testosterona. La Testosterona masculiniza el cerebro en el
desarrollo temprano.
Las Espermátidas tienen a continuación un proceso progresivo de
transformaciones morfológicas que recibe el nombre de Espermiogénesis. Estos
cambios
son: se forma el Acrosoma, que ocupa la mitad de la superficie nuclear y
contiene las enzimas que ayudan a la penetración del espermatozoide en las
capas que rodean al ovocito, y el núcleo se condensa (estas dos estructuras
caracterizan la cabeza del espermatozoide); se forma el cuello, pieza intermedia
y cola; gran parte del citoplasma es eliminado. La transformación desde
Espermatogonias hasta Espermatozoides maduros es de 64 días. Después de
formados pasan a la luz de los túbulos seminíferos, desde donde se empujan
hacia el Epidídimo por los elementos contráctiles que se encuentran en la pared
de los túbulos seminíferos. Allí adquieren su movilidad y continúan por el sistema
de conductos masculinos en la misma medida en que se forma el semen
líquido que los contienen y los mantiene biológicamente viables.
Al igual que en la ovogénesis, los Espermatocitos I comienzan con un número
diploide de cromosomas cuando empieza la Meiosis I, y terminan siendo
células haploides, las espermátidas. El 50 % de de los espermatozoides forma
dos tendrán un cromosoma sexual
X
y el 50 % un cromosoma sexual Y (Fig.
5-2 y 5.3).
Gametos masculinos anormales: Las anomalías de los gametos masculinos
pueden ser morfológicas de diferente tipo: a veces en la cabeza, en la cola o en
ambas, pueden ser gigantes o enanos. También pueden presentar defectos
cromosómicos en el número o integridad de los cromosomas. En el hombre es
frecuente cuando hay algún problema en la reproducción realizar un
Espermograma o Espermiograma. Los valores
normales de las variables más
importantes del semen aparecen en la
tabla 5.2. - .
Tabla
5.2. Variables y valores normales del semen.
Volumen: 2 ml o más
Ph: 7.2-7.8
Cantidad espermatozoides: 20 x 10 a la 4 o
más
Motilidad: A + B = 50% o más o A = 25%
o más.
Morfología: 50 % o más con morfología normal
Viabilidad: 50 % o más vivos
Leucocitos: < I x 103/ml
Nomenclatura de algunas variables para el semen:
-
Normozoospermia: Eyaculado normal según la definición precedente.
-
Oligozoospermia: Concentración de espermatozoide » menor de 20 x 103
/ ml
-
Astenozoospermia: Menos del 50 % de los espermatozoides con progresión
antero grada (A+B).
-
Teratozoospermia:
Menos del 50 % de los espermatozoides con
morfología normal.
-
Oligoastenoteratozoospermia:
Perturbación de las tres variables.
-
Azoospermia: Ausencia de espermatozoides en el eyaculado.
-
Aspermia: Ausencia de eyaculado.
Regulación de la Espermatogénesis: En los Túbulos Seminíferos también
ocurre la esteroidogénesis ó síntesis de esteroides sexuales por las células de
Leydig que secretan la Testosterona, que es la principal hormona sexual masculina.
Ambos procesos están regulados por las hormonas .gonadotropinas, secretadas por
la Hipófisis anterior. La FSH tiene su célula blanca en las de
Sertoli y la LH en las células de Leydig.
La trascripción de genes por protaminas aparece tempranamente en las
células haploides o espermátidas, lo que las ayudara después a entrar a la luz
de
los tubos seminíferos. Las células germinativas espermatogénicas están unidas
a las células de Sertoli por la N-Cadherina.
diferenciación sexual
El sexo cromosómico se determina en el momento de la fecundación, lo que
decide una cascada de procesos que constituyen la diferenciación sexual del
individuo. Sin embargo, hay una etapa en la que las gónadas tienen la misma
apariencia morfológica, que es conocida como período o etapa indiferenciada.
Pero en breve, la presencia de determinados genes condiciona diferencias evidentes,
propias de las características gonadales de cada sexo que serán estudiados
en el Sistema Reproductor. (Fig.5.4)
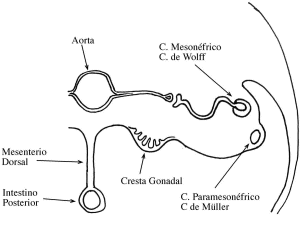
Numerosos son los genes encontrados que participan en la diferenciación
sexual. En los humanos tiene gran importancia el gen determinante testicular,
que está en relación con un gen que se encuentra en el brazo corto del cromosoma
Y.
Este gen es llamado SRY y codifica la proteína High Mobiliy Group Box. Se
conoce que el SRY determina que en la cresta gonadal se forme el epitelio de
células específicas del varón, las células de Sertoli. También induce a la
gónada
en formación y ella produce un factor quimiotáctico que atrae a las células
mesonéfricas a la gónada, las cuales tienen la importancia de inducir al
epitelio
en la formación de las células de Sertoli. Sin la presencia de estas proteínas
se
desarrollará un ovario en vez de un testículo.
SOX9 es un gen autosómico que también participa en la diferenciación sexual.
Codifica un factor de trascripción que es esencial para la formación testicular.
SF1/Sf1, es un factor de trascripción
que directa o indirectamente activado por
el SRY es necesario para permitir la bipotencialidad de la gónada. La
diferenciación tiene dos fases:
1. Formación de la gónada durante la órgano génesis
2. Desarrollo femenino o masculino en respuesta a las hormonas secretadpor
las gónadas durante la adolescencia y controladas por el eje Hipotálamo-Hipofisiario.
Estas dos etapas se estudiarán con más detalles en la asignatura
Morfo fisiología
III.
tránsito de los gametos masculinos
Antes de que ocurra la fecundación los gametos masculinos deben realizar
un tránsito por las vías reproductoras masculinas, con lo cual terminan su maduración
morfológica. Después de ser depositados en la vagina recorren un trayecto dentro
del sistema reproductor femenino hasta que llegan al tercio
externo de la tuba uterina donde ocurre la fecundación normalmente.
En el tracto genital femenino debe ocurrir la capacitación, que se calcula
que en humanos sea muy breve, lo cual es necesario para que los
espermatozoides puedan fecundar al ovocito.
Este proceso incluye la eliminación de la cubierta glicoproteica, que rodea al
plasmalema sobre el acrosoma y que el
espermatozoide recibió durante su tránsito por e! epidídimo, y del plasma
espermático. Además, se reorganizan las
moléculas de la membrana celular y el
patrón de movilidad se modifica (hiperactivación), con golpes mucho más
rápidos en la cola. Estas transformaciones permiten que el
espermatozoide pueda atravesar todas
las barreras que rodean al ovocito. Si no lo logra evidentemente
nos encontramos ante un problema de Infertilidad o Esterilidad de la
pareja. Después de estos cambios que ocurren
durante el tránsito, puede ocurrir la
Fecundación.
fecundación
El proceso de la Fecundación permite la auto perpetuación de la especie, lo
cual tiene una gran importancia para mantener el nivel de población. La fecundación
puede ocurrir ya en la pubertad, cuando existe la madurez de los sistemas
Endocrino, Nervioso y Reproductor. Es entonces que comienza el desarrollo
del individuo, desde el cigoto, pasando por la etapa Pre-embrionaria o primera
semana del desarrollo prenatal o intrauterino. Este proceso ocurre en la Ampolla
de la Trompa uterina.
La fecundación tiene varias fases las cuales son:
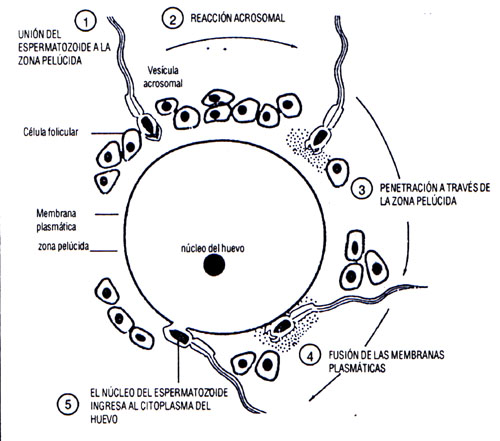
- Penetración de la corona radiada: Solo de 300-500 espermatozoides llegan al
sitio de la fecundación y solamente el que haya pasado al estado de capacitación
atraviesa esta capa de células foliculares pero los demás contribuyen
con él.
Con el contacto del espermatozoide con la corona radiada se desencadena
la reacción acrosómica, por lo que aparecen varios orificios en la parte superficial
de la membrana acrosómica y el plasmalema externo. Se forman poros
que permiten la salida de enzimas como la hialuronidasa, que permite atravesar
la corona radiante, y la acrosina, que contribuye a penetrar a la ZP y después
se
degrada.
-
Penetración de la zona pelúcida: La ZP es la barrera o capa donde existen
los primeros vínculos moleculares, está ubicada por fuera de la membrana
plasmática del ovocito, formada por glicoproteínas y tiene tres capas, ZP1,
ZP2 y ZP3. Esta última es muy importante en este proceso ya que determina un
reconocimiento mutuo entre los gametos.
Los ovocitos contienen vitelo o sustancias nutritivas en su interior de forma
moderada en los mamíferos: lípidos, proteínas y polisacáridos, formando las
llamadas plaquetas vitelinas. En su corteza presentan los gránulos corticales,
que cuando un espermatozoide activa el ovocito y ocurre la exocitosis; modifican
la cubierta de la célula e impiden la entrada de otro espermatozoide.
La ZP3 desencadena la reacción acrosómica induciendo la entrada de Ca2+
al citosol del espermatozoide. Estos iones Ca2+ también penetran al ovocito y lo
activan.
Quedan también al descubierto otras proteínas de la superficie del
espermatozoide que se unen a la ZP2, lo cual contribuye a mantenerlo unido a la membrana
del ovocito mientras penetra.
-
Fusión de las membranas celulares del ovocito y del espermatozoide: La
adhesión del espermatozoide a !a membrana del
ovocito es mediada en parte
por Integrinas; después se fusionan
las membranas y penetra al ovocito el
contenido del espermatozoide (Fig.
5.5 y 5.6). La membrana citoplasmática
queda en la superficie y después se
pierde. Cuando ocurre esta penetración
el ovocito responde con:
-
Reanudación de la Meiosis:
se forma así una célula con abundante citoplasma
y un segundo corpúsculo polar. El primer corpúsculo también puede dividirse, en
resumen hay 4 células pero sólo una con abundante citoplasma
que contiene los núcleos del ovocito propiamente y el del espermatozoide.
Estos núcleos se acercan y desaparece la membrana nuclear.
-
Reacciones corticales y de zona: se libera el contenido de los gránulos
corticales que contiene enzimas lisosómicas, lo que hace a la membrana del
ovocito impenetrable a la entrada de otro espermatozoide, por lo cual se
bloquea la polispermia.
-
Activación
metabólica del huevo: parece que el factor activador lo aporta el
espermatozoide y esta activación es importante para los procesos
celulares
moleculares que ocurren en las primeras etapas del desarrollo.
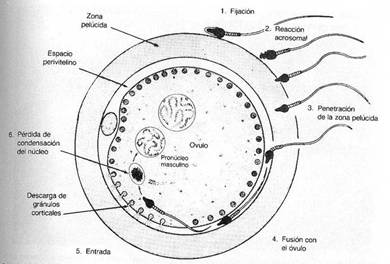
Los
resultados o consecuencias de la Fecundación son: Restablecimiento
del número diploide de cromosomas, determinación del sexo genético, iniciación
de la segmentación o clivaje.
vinculación con la práctica médicA
Todo el contenido explicado está en relación con diferentes problemas de la
reproducción.
Infertilidad y Esterilidad: es un problema frecuente que consiste en la no
concepción después de un año de relaciones sexuales sin ningún medio de
planificación. Aproximadamente el 20% de las parejas están aquejadas por
infertilidad la cual podría ser curable en mas del 90% de los casos si se
hiciera un diagnostico adecuado.
Sin embargo la falta de dicho diagnóstico impide lograr el embarazo.
Generalmente la infertilidad se origina en ambos miembros de la pareja y no en
uno sólo, por lo que es indispensable estudiar tanto al hombre como a la mujer.
La esterilidad
es la incapacidad total de concebir. Aproximadamente el 1% de las parejas son
estériles, lo
cual significa que la única opción que tienen es la adopción o, de acuerdo al
caso, alguna técnica de reproducción asistida cuando hay alguna posibilidad por
uno de los miembros de la pareja.
• Planificación Familiar y Anticoncepción. El embarazo debe ser deseado, para
lo cual; la pareja organiza y planifica el momento adecuado de acuerdo a sus
condiciones. Por lo tanto, también es necesaria la responsabilidad de la protección
para evitar un embarazo y contra las enfermedades de transmisión
sexual (ETS). Existen numerosos métodos anticonceptivos pero sólo se mencionarán
algunos: de barrera (preservativo masculino, el diafragma, el capuchón
cervical y las esponjas anticonceptivas); las píldoras, que por su contenido
.hormonal permiten la menstruación pero inhiben la ovulación; los implantes
subdérmicos, que inhiben la ovulación durante años; los dispositivos
intrauterinos (DIU), que se colocan en la cavidad uterina; drogas que provocan
aborto si se administran dentro de las 8 semanas siguientes a la última
menstruación, la Vasectomía y la ligadura de las Trompas.
• Técnicas de Reproducción asistida:
- Fecundación in vitro y Transferencia de embriones. Se recolectan los
ovocitos en Meiosis
II,
se fecundan en el laboratorio y cuando han terminado la
segmentación hasta la etapa de 8 células entonces son colocados en el útero
donde continúa ei desarrollo hasta el término.
- Transferencia intrauterina de gametos (GIFT). Se introducen ambos
gametos en la ampolla de la trompa y continúa el proceso normal.
- Transferencia intratubárica del cigoto (ZIFT). Se colocan los ovocitos
fecundados en la región ampollar de la trompa.
- Maternidad subrogada. Con los gametos de ambos cónyuges se realiza la
fecundación y se implanta el cigoto en la etapa de 8 células en un útero de
otra mujer fértil. Esta técnica lleva aparejados muchos problemas éticos y
legales.
5.4
primera semana del desarrollo
El cigoto humano tiene aproximadamente de 100 a 120 μm de diámetro y se
desarrolla en el interior del sistema reproductor femenino, específicamente
dentro
de las Tubas uterinas. El cigoto es del tipo Alecítico (tiene poco vitelo y está
ubicado centralmente alrededor del núcleo). Después de la ovulación, la fertilización
ocurre en la ampolla del oviducto, termina la meiosis y un día después
comienza el clivaje o segmentación.
El primer clivaje es meridional, pero en la segunda división una blastómera
se divide meridionalmente y la otra ecuatorialmente. Este tipo de clivaje
es el
llamado clivaje rotacional. En estos primeros
momentos existen uniones comunicantes entre las células
que reciben el nombre de
Blastómeros(as). Este proceso
es además asincrónico y asimétrico. En el estadio de ocho células ocurre
la compactación, durante la cual las células se unen íntimamente unas con otras
y se conservan las uniones intercelulares que permiten que pasen iones y pequeñas
moléculas.
Cuando la segmentación ya ha llegado de 16 a 32 células estamos en presencia
de la Mórula. Las células que se encuentran en la periferia forman la
masa celular externa y las centrales la masa celular interna, las cuales por
diferentes procesos se van ubicando hacia un extremo. La Mórula aún permanece
rodeada por la Zona pelúcida, pero llega el momento que por un poro que
se forma en esta sale de esta envoltura convertida ya en un Blastocisto. La
capa externa de esta nueva estructura recibe el nombre de Trofoblasto y sus
células son planas; participa en la nutrición embrionaria primero y enla
formación
de la Placenta después. Las células que se encuentran hacia un extremo
del Blastocisto reciben el nombre de Embrioblasto, y formarán al embrión. En
e! centro hay una cavidad llamada Blastocele. Todos estos procesos ocurren en
el interior de la Tuba uterina; el cigoto en
transformación
es libre y allí ocurre un
intercambio con el medio materno donde se desarrolla y obtiene los nutrientes
necesarios. (Fig. 5.7).
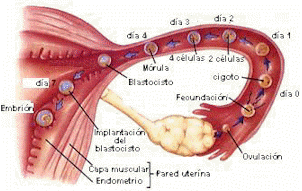
Figura 5. 7: Representación de los eventos que se producen durante la primera
semana del
desarrollo
Representación de procesos en orden de aparición:
1. Fecundación.
2. Cigoto.
3. Clivaje, dos Blastómeros.
5. Clivaje
6. Mórula
7. Blastocisto
8. Blastocisto adherido al Endometrio
La mucosa de la tuba posee un epitelio cilíndrico simple con células ciliadas
y no
ciliadas.
Las primeras contribuyen al movimiento del
cigoto en desarrollo
hacia la cavidad uterina, donde debe implantarse, y las segundas tienen función
de secreción. La secreción de estas células comienza a liberarse justo antes de
la ovulación. Ambos tipos de células tienen modificaciones debido a las influencias
hormonales.
Regulación del proceso: Desde la formación del cigoto o quizás antes, existen
distribuciones en el citoplasma del ovocito o del cigoto que marcan el destino
futuro, condicionado por numerosas variantes. Por ello la morfogénesis de
muchas regiones corporales esta determinada por campos morfogenéticos aún
desconocidos. Estas regiones del cuerpo están bajo el control de un plano global
del desarrollo genético.
Ni en las etapas más precoces los blastómeros de un embrión en segmentación
son homogéneos. En la etapa de cuatro células los niveles de síntesis de
ARN son bajos, mientras que en la de ocho células los niveles son muy altos en
algunas mientras que en otras se presenta el patrón de las de cuatro células.
Por lo tanto, existen blastómeros activos e inactivos desde el punto de vista de
la trascripción, lo cual implica diferencias entre ellos que deben tener una
implicación
en el desarrollo. Este hecho puede, estar en relación con su destino
prospectivo, o sea los tejidos que formarán en el futuro en una posición determinada.
A pesar de que aumenta el número de células por divisiones mitóticas hasta
la formación de la Mórula, la talla y las dimensiones espaciales no han cambiado,
por lo que se afirma que no hay crecimiento en esta etapa de divisiones. La
formación del Blastocisto coincide con la perdida de la zona pelúcida, con lo
que comienza la diferenciación del trofoblasto. Este proceso es producto de la
proteólisis de la ZP, la cual es estimulada por el Factor de Crecimiento Epidérmico
(EGF). Los estrógenos estimulan la síntesis de este factor, tanto en el
Blastocisto come en las células epiteliales uterinas, así como la de sus receptores
localizados en las células trofoblásticas. El efecto del EGF sobre la maduración
del Blastocisto es autocrina/ paracrina. La masa celular interna permite el
desarrollo trofoblástico produciendo proteínas tales como FGF4, que determina
las divisiones celulares en esta capa.
El endometrio está constituido por dos capas fundamentales: la basal y la
funcional. La primera de ellas posee un estroma más celular, muestra escasa
modificación cíclica, no se pierde con la menstruación y funciona como zona de
regeneración de la capa funcional. La capa funcional si es eliminada, quizás no
totalmente; con la menstruación y ocurren en ella modificaciones cíclicas.
Existe
una gran irrigación sanguínea en cada capa y cerca de la superficie se forma
una red capilar. La preparación del endometrio es paralela a la Folículo génesis
que está ocurriendo en el ovario.
Se ha demostrado con estudios experimentales que existen moléculas de
adhesión celular del tipo de las Integrinas en la superficie de las células
endometriales y células trofoblásticas que son responsables de la
fijación del Blastocisto y
posiblemente de que esto ocurra en un determinado lugar. La
implantación es intersticial porque queda
incluido el Blastocisto dentro de la
mucosa uterina.
ETAPAS FUNDAMENTALES DURANTE LA IMPLANTACIÓN.
1ra.
Anclaje del blastocisto. Parece que es por la mediación de ligandos de
las Integrinas de un lugar especifico del trofoblasto en el tejido
endometrial.
2da. Penetración del epitelio uterino. Se caracteriza por la invasión del
sincitotrofoblasto a la capa funcional endometrial que se encuentra en reacción
decidual. Esto con lleva la invasión de los tejidos subyacentes al epitelio y la
erosión de los vasos maternos. Comienza al final de la primera semana y culmina
en la segunda semana del desarrollo (Fig. 5_8)
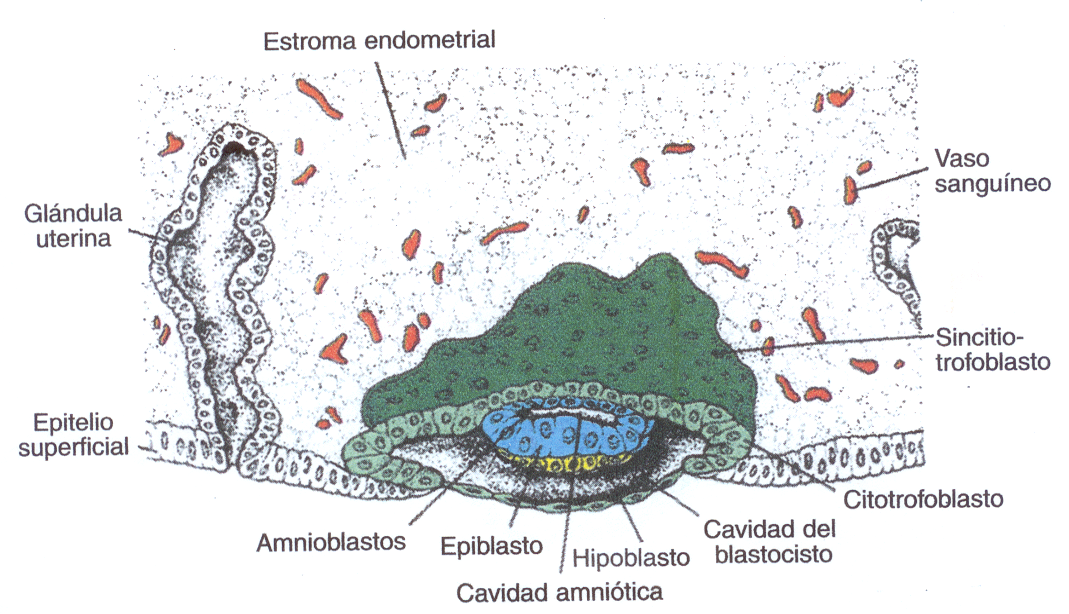
ASPECTOS
INMUNOLÓGICOS DURANTE LA IMPLANTACIÓN
El feto y la placenta son desde el punto de vista inmunológico diferentes
a la madre pero sin embargo no son reconocidos como tejidos ni tejidos extraños
ni rechazados por el sistema inmunológico de ella. El rechazo de tejidos
extraños ocurre normalmente por la activación de los linfocitos citotóxicos pero
también por posibles respuestas humorales inmunes. Se sugieren varias
explicaciones para la especial tolerancia de la madre a la presencia prolongada
del embrión inmunológicamente diferente durante el embarazo:
1.
Los tejidos fetales
en espacial el de la placenta, constituyen la interfase directa entre el feto y
la madre y no presentan antígenos extraños al sistema inmunológico de la madre
realmente ni el sincitotrofoblasto ni la cápsula citotrofoblástica expresan los
dos tipos principales de antígenos mayores de histocompatibilidad que
desencadenan la respuesta inmunológica del huésped en el rechazo de los injertos
de tejidos. Sin embargo, estos antígenos están presentes en las células del feto
y en los tejidos estromales de la placenta. La expresión de los antígenos
menores de histocompatibilidad, por ejemplo el antígeno HY, en fetos masculinos
sigue un patrón similar. No obstante los tejidos trofoblasticos expresan otros
antígenos menores. Por las brechas existentes en la membrana placentaria es
frecuente encontrar glóbulos fetales rojos y blancos circulando en la sangre
materna. Estas células deberían poder sensibilizar al sistema de defensa de la
madre.
En el sistema inmunológico de la madre ocurre una especie de bloqueo
durante el embarazo de manera que no reacciona a los antígenos totales a los
cuales se expone. Pese a ello la madre es capaz de poner en marcha una respuesta
inmunológica al injerto de un tejido extraño. Puede que exista una respuesta
selectiva del sistema inmunológico ante los antígenos fetales, aunque la
respuesta de incompatibilidad Rh muestra que esta no debe ser total.
La tercera posibilidad es que la barrera decidual local impida el
reconocimiento inmunológico del feto por parte de la madre o el paso de células
inmunocompetentes de la madre al feto. Hay pruebas de una barrera funcional
decidual funcional, pero un número importante de casos sabe que esta barrera se
rompe debido a traumatismos o enfermedades.
Aun falta mucho por dilucidar en las complejas interrelaciones
inmunológicas entre el feto y la madre.