3.4. Citoesqueleto
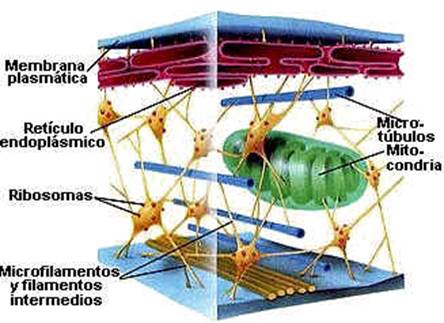
La célula debe su forma a un conjunto de proteínas globulares y filamentosas que forman el citoesqueleto. Este se distribuye en el núcleo, dando origen al carioesqueleto; en el citoplasma, originando el citoesqueleto propiamente dicho, o simplemente citoesqueleto, el cual se extiende desde el núcleo a todo el citoplasma y hasta la membrana plasmática y por ultimo forma el esqueleto de la membrana, que se dispone en la superficie interna de la misma.
Citoesqueleto
Se puede definir como un entramado tridimensional de filamentos y túbulos proteicos que ocupan el interior de la mayoría de las células eucariota y que adquiere una relevancia especial en las células animales, ya que estas carecen de pared celular. En este tipo de célula, el cito esqueleto mantiene la estructura y la forma de la misma actuando como un bastidor para la organización de esta y la fijación de orgánulos y enzimas.
El cito esqueleto también es responsable de diversos movimientos celulares. La motilidad celular es uno de los grandes logros de la evolución y el citoesqueleto es esencial como componente de soporte para este proceso. Las células eucariotas tienen la capacidad de organizar movimientos directos para migrar, alimentarse, dividirse y dirigir coordinadamente el transporte de materiales intracelulares. El mecanismo y dirección del movimiento se realiza de diferentes maneras y esta asociado con la utilización de energía. Otra función relevante es guiar el transporte de organelos intracelulares y de otros elementos. Por ultimo, componentes del citoesqueleto forman un papel primordial en la división celular como se vera en el momento oportuno.
El citoesqueleto no es una estructura permanente, sino que se desensambla sin cesar, y esta formado por tres componentes principales: microtúbulos, microfilamentos y filamentos intermedios.
Los tres componentes del citoesqueleto están interconectados y forman un retículo, que se extiende desde la superficie celular hasta el núcleo. Este sistema esta construido sobre la base de un modelo arquitectónico común que se encuentra en una sorprendente variedad de sistemas naturales y se conoce como modelo de integridad tencional. Con esta expresión se indica que el sistema se estabiliza mecánicamente así mismo, en razón del modo en que las fuerzas de compresión y tensión se distribuyen y equilibran dentro de la estructura.
Las estructuras q responden a este modelo de integridad tensional no alcanzan la estabilidad mecánica por la resistencia de los miembros individuales sino por la manera en q la estructura, en su conjunto distribuye y equilibra las tensiones mecánicas. Estas estructuras la tensión se transmite sin solución de continuidad a través de todos los elementos estructurales. En otras palabras, un incremento de tensión en un elemento cualquiera de la estructura se hace sentir en todos los demás elementos. Este aumento global de presión se equilibra por un aumento de la compresión de determinados elementos distribuidos por la estructura (Fig. 3.46).
Fig. 3.46. Esquema de la trama del citoesqueleto en el cual se observa su disposición tridimensional y su relación con los orgánulos celulares.
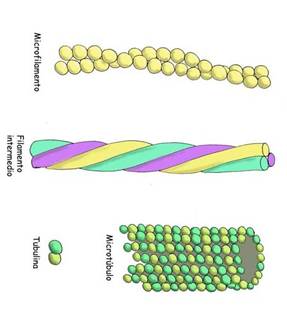
En la figura 3.47 aparecen los componentes del citoesqueleto que se localizan en el citoplasma. En la porción superior los microfilamentos, que son las estructuras de menor diámetro (7nm). En el medio aparecen los filamentos intermedios, los cuales presentan un diámetro intermedio entre los microfilamentos y los microtúbulos (10nm). En la porción inferior aparecen los microtúbulos con un diámetro de 25nm.
MICROTÚBULOS Y PROTEÍNAS ASOCIADAS
Los microtubulos son estructuras tubulares que se distribuyen de preferencia alrededor del núcleo y que al microscopio electrónico aparecen como si sus extremos se fijaran a la membrana plasmática o en formaciones cercanas a ella. Estos diminutos túbulos huecos actúan como entramado estructural de las células; al mismo tiempo, transportan sustancias de una parte a otra de la célula.
Cada uno de los microtubulos esta formado por 13 protofilamentos, que son polímeros formados por dos tipos de moléculas proteicas globulares, casi esféricas,
Llamadas á y â tubulina, que forman unidades heterodimericas. Los monómeros de tubulina presentan estructuras primarias muy semejantes y un peso molecular de 55000 daltons cada una. Los protofilamentos se disponen en forma de espiral, como muestra la figura 3.47
Los microtúbulos se pueden disponer Formando estructuras aisladas o agrupados formando organitos microtubulares como son: los flagelos y los centríolos.
Los microtúbulos aislados a su vez pueden encontrarse dispersos en el citoplasma celular o formando las fibras del uso acromático en la división celular.
Los microtúbulos pueden asociarse formando dobletes o tripletes; dobletes en caso de los cilios, flagelos y cuerpos básales de los cilios, y tripletes en los centríolos (Fig. 3.48)
PROTEINAS QUE ACTUAN COMO MOTORES CELULARES
Las células poseen proteínas que actúan como motores celulares que ligan dos moléculas y osando ATP como energía causan que una molécula cambien en relación a otra. Entre estos motores están: la miosina, la dineina y la quinecina.
Estas familias de proteínas tienen un extremo motor pero pueden tener varias clases de distintas estructuras moleculares en el extremo ligante. Estas proteínas motores son capaces de transportar organelos a lo largo de microtubulos convirtiendo la energía libre derivada de la hidrólisis del ATP en movimiento dirigido. (Fig. 3.49 y 3.50).
Entre las proteínas motoras que interactúan con los microtubulos se encuentran:
MICROFILAMENTOS.
Los microfilamentos son componentes del citoesqueleto que resultan de la polimerización de un tipo fundamental de proteína, la actina. En la actualidad se han descrito varios tipos de actina (al menos 6). Estas proteínas son codificadas por genes diferentes.
Los microfilamentos poseen un diámetro de unos 7 nm. Están formados por dos cadenas de actina que forman una hélice doble (Fig. 3.47). Su mayor concentración se encuentra por debajo de la membrana plasmática, porque entre sus funciones se encuentran: mantener la forma de la célula y formar protuberancias citoplasmáticas como los seudópodos y las microvellosidades. Otras de sus funciones son participar en la unión de dos células o entre las células y la matriz celular, en la trasducción de señales y por ultimo en la motilidad celular. En el caso de las células musculares, y en asociación con la proteína motora miosina, permiten la contracción muscular, aspecto que se vera en el tema de tejido muscular.
En el citoplasma, los microfilamentos se organizan de diversas formas:
· En las células musculares forman los filamentos finos que se asocian a los filamentos gruesos (proteína motora de la actina).
· En las microvellosidades forman hojas de 20 filamentos.
· En la cara interna de la membrana plasmática forman filamentos cortos, componentes del esqueleto de la membrana.
FILAMENTOS INTERMEDIOS.
Los filamentos intermedios están formados por proteínas fibrosas, estables y poco solubles (fig. 3.47). su diámetro es de aproximadamente 10 nm. Estas proteínas se combinan en dímeros helicoidales, que se asocian para formar tetrámeros alargados (protofibrillas). Cuatro protofibrillas conforman un filamento intermedio. Son apolares y tienen como funciones mantener la fuerza de tensión celular ( principal) y como soporte mecánico.
Los filamentos intermedios están formados por proteínas fibrosas no se desintegran fácilmente. Intervienen en la estructura de la membrana nuclear y desde allí pueden irradiar y asociarse con los microtubulos.
Clasificación de las proteínas de los filamentos intermedios:
· Tipo I: queratinas ácidas, por ejemplo los epitelios.
· Tipo II: queratinas básicas, por ejemplo en los epitelios.
· Tipo III: vimetina en mesenquima, desmina en el músculo, periferina en neuronas y proteína ácida fibrilar de la glia en las células gliales del sistema Nervioso.
· Tipo IV: neurofilamentos (L, M, H) en neuronas, internexina en el sistema nervioso en formación.
· Tipo V: laminas nucleares A, B y C en el núcleo de todas las células.
· Tipo VI: nestina presente en las células neuroepiteliales.
La mayoría de las células adultas posee solo un tipo de filamentos intermedios.
El patrón de distribución celular de los mismos puede ayudar al patólogo a diagnosticar un determinado tipo de cáncer. Las proteínas asociadas a los filamentos intermedios forman una red con estos organelos y la membrana plasmática.
3.5. ESPECIALIZACIONES DE LA SUPERFICIE CELULAR.
La superficie de algunas células presenta especializaciones relacionadas con su función. Entre estas especializaciones las mas conocidas son:
· Las zónulas ocluyentes.
· Las zónulas adherentes.
· Los desmosomas.
· Los hemidesmosomas.
· Las invaginaciones.
· Los contactos focales.
· Las uniones tipo nexo, uniones comunicantes (gap junctions).
· Los cilios y flagelos.
· Las microvellosidades.
Estas especializaciones están formadas en su mayoría por componentes del citoesqueleto cubiertos por la membrana plasmática, con excepción de las uniones tipo nexo, que solo están formadas por membrana plasmática.
ZÓNULAS OCLUYENTES.
Las zonulas ocluyentes son características de las superficies latero apicales de las uniones entre células epiteliales cilíndricas. Todas las zonulas son estructuras en forma de faja, que forman un cinturón alrededor de la célula. Las zonulas ocluyentes se caracterizan por la intima yuxtaposición periódica de las membranas celulares de las células próximas, con fusión de las hojas externas de las membranas plasmática respectivas. Las técnicas de criofractura demuestran que hay entrantes y salientes en las membranas adyacentes. La zonula ocluyente forma una barrera protectora que impide el paso de las moléculas entre células; tiene por tanto, un efecto de cierre, no permitiendo el paso de material al intersticio entre dos células.
ZÓNULAS ADHERENTES
Esta unión rodea todo el perímetro de la célula y contribuye ala adhesión entre las células adyacentes. Esta formada por receptores de adhesión de la familia de las caderinas (caderina clásica E). En este tipo de unión hay una separación entre membranas celulares. En la superficie interna (citoplasmática) de la membrana de cada célula se encuentra un deposito de material electrón denso, o placa de adhesión ala cual se unen microfilamentos de actina del velo terminal (este ultimo se localiza por debajo de la superficie apical en las células epiteliales). Estas uniones se localizan por ejemplo: entre células que forman epitelios cilíndricos simples, entre células musculares cardiacas, entre otras.
DESMOSOMAS O MÁCULA ADHERENTE.
Este tipo de especialización esta presente en varios tipos celulares, como por ejemplo en los tejidos epiteliales que están sometidos a fuertes tracciones, presiones, como es el caso de la piel. En general se encuentran en membranas epiteliales simples y estratificadas, entre celulares epiteliales glandulares y entre células musculares estriadas cardiacas.
En este tipo de adhesión participan glicoproteinas trasmembranales conocidas como receptores de adhesión de la familia de las caderinas, que tienen como función adherir alas membranas plasmáticas de células vecinas. En ausencia del calcio las caderinas pierden su poder adhesivo.
El desmosoma es una estructura compleja en forma de disco constituido por las membranas de dos células continuas en la región del desmosoma, las membranas están separadas por un espacio de 20 nm; en dicho espacio se puede encontrar material electrón denso que corresponde con los dominios extracelulares de las glicoproteinas desmogleina y desmocalina de los receptores de adhesión. En los cortes, las membranas celulares aparecen rectas, lo que sugiere que el desmosoma presenta rigidez. en las caras citoplasmáticas de cada hemimembrana hay una placa circular formada por varios tipos de proteínas (placa de adhesión) ala cual se fijan filamentos intermedios de citoqueratina en los epitelios y de desmina en el caso de las células musculares (figs. 3.51 y 3.52).
En resumen, en los desmosomas:
· Las membranas de las células adyacente corren paralelas entre si, separadas por un espacio de unos 20nm el cual presenta una línea densa en su zona media.
· Adherida ala cara intracelular de la membrana plasmática se encuentra una gruesa banda llamada placa desmosomica.
· Insertados ala placa desmosomica aparecen numerosos filamentos intermedios.
Figura 3.51. a) se observan varias células unidas por desmosomas; en la porción basal se observan hemidesmosomas. b) se observa un esquema del desmosoma mostrando la placa de adhesión ala cual llegan los filamentos intermedios.
Fig. 3.52 fotografía electrónica mostrando un desmosoma.
HEMIDESMOSOMAS
Estructura similar a los desmosomas por sus características morfológicas pero solo presentan la mitad de estos últimos, ya que su mitad externa esta formada por fibrillas de colágeno; es decir, semejantes a los desmosomas por su forma pero difieren de estos tanto a nivel funcional como bioquímica. El hemidesmosma une el dominio basal de las células a la lámina basal.
Se localizan en la superficie basal de las células epiteliales estratificadas. Estas especializaciones están formadas por receptores de integrina en lugar de caderina, como sucedía en los desmosomas, y a estos receptores se van a unir filamentos intermedios de citoqueratina (Fig.3.51)
INVAGINACIONES.
En las células epiteliales del tubo contorneado del riñón, la membrana celular forma profundas invaginaciones entre las cuales se encuentran dispuestas mitocondrias. La presencia de mitocondrias entre las invaginaciones indica el gran intercambio de material a través de la membrana, por el gasto energético que esto provoca. (Fig. 3.51)
CONTACTOS FOCALES
Son uniones que se establecen entre célula y la matriz extracelular. Las gluco proteínas trans membranales de unión o receptores de integrinas conectan los haces de filamentos de actina (fibras de estrés) a la matriz extracelular.
Esta es una estructura transitoria que se forma y desaparece constantemente y mediante la cual las células pueden trasladarse o migrar en la matriz extracelular, fenómeno este particularmente importante durante la embriogenesis.
UNION CON HENDIDURA (GAP JUNCTION)
También llamadas nexo o unión comunicante. Se puede localizar en las superficies laterales de las células epiteliales, en las células musculares estriadas cardiacas, entre células nerviosas dando origen a sinapsis eléctricas, entre otras.
Las uniones comunicantes están formadas por hexameros proteicos que van a dar lugar a placas circulares de seis unidades proteicas. Estos hexameros forman estructuras llamadas hemiconexones, y al unirse dos de ellos forman la unión tipo nexo. Este tipo de unión posee un canal hidrofilico de 1.5 nm que permite el paso de iones, pequeñas moléculas y segundos mensajeros. Estas uniones tienen también un papel importante en la embriogenesis, coordinado el desarrollo de los tejidos del embrión (Fig. 3.54.)
Fig. 3.54 sinapsis eléctrica.
MICROVELLOSIDADES
Las microvellosidades le confieren al borde apical de las células un aspecto estirado, cuando se observan al M/O. en las células del tubo contorneado proximal, forman el llamado ribete en cepillo y, en las células de epitelio intestinal forman la chapa estriada con el M/E se observan como evaginaciones de la membrana celular, de 1.5 μm de alto por 0.1 μm de alto. El centro contiene un as de microfilamentos de actina que se relacionan con el velo celular o velo terminal, y con la membrana plasmática. En algunas células, la membrana plasmática asociada alas micro vellosidades presenta un glicocálix muy desarrollado. La función fundamental de estas especializaciones es aumentar la superficie de absorción. (Fig. 3.55).
Cilios y flagelos
Ambos son evaginaciones de la superficie celular. Los cilios son más pequeños y numerosos, y los flagelos más largos y únicos. Tanto los cilios como los flagelos tienen una estructura parecida y tienen como función el movimiento; por ejemplo, el flagelo del espermatozoide (única célula humana con flagelo) permite su desplazamiento a través de los conductos que debe atravesar. Los cilios permiten desplazar partículas, por ejemplo, en el sistema respiratorio, o células, como el ovocito a través de la tuba uterina. Los cilios tienen una longitud de 2-10 μm y un diámetro de 0.5 μm. Los flagelos alcanzan una longitud de 100 μm aproximadamente y 0.5 μm de diámetro.
En un corte transversal de un cilio se observa el M/E la estructura de los cuatro elementos que lo constituyen: cilio (propiamente dicho), placa basal, cuerpo basal y raicillas. El cilio o axonema es la prolongación cilíndrica que se proyecta a partir de la superficie de la célula, compuesta por nueve fibras y cada una constituida por dos microtúbulos; además en el centro de estos nueve pares de microtúbulos se observan un par de microtúbulos más (Fig. 3.56).
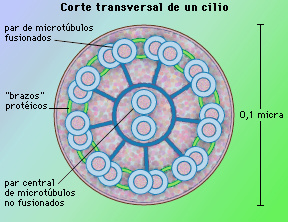
Figura 3.56. Corte transversal de un cilio donde se observa una parte central formada por microtúbulos, o axonema. El axonema esta constituido por dos microtúbulos centrales rodeados por nueve pares de microtúbulos.
3.6. Organelos citoplasmáticos. Inclusiones. Citosol
ORGANELOS MEMBRANOSOS
Retículo endoplasmatico
Este orgánulo se encuentra en la mayoría de las células. Es un orgánulo membranoso y consiste en un conjunto de túbulos, cisternas y sacos aplanados dispuestos en forma de red, conectados unos con otros, que se distribuyen por toda la célula irradiando desde el núcleo al aparato de Golgi. La cantidad de retículos en una célula depende de su actividad celular. Este orgánulo u organito se puede encontrar en una célula animal o vegetal pero no en una célula procariota.
Existen dos variedades: el retículo endoplásmico liso y el rugoso, que se diferencian por su estructura y funciones. El retículo endoplasmico liso posee contornos lisos y continuos, mientras que el rugoso presenta asociado a sus membranas ribosomas, los que le confieren este aspecto rugoso. El retículo endoplásmico liso predomina la organización de túbulos membranosos, mientras que en el rugoso predominan los sacos y cisternas (Fig. 3.57).
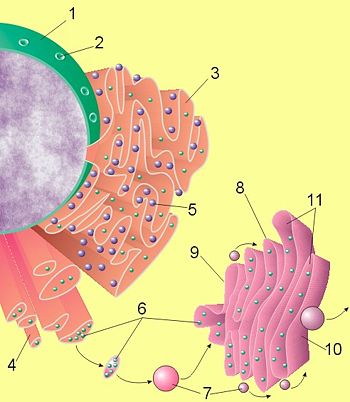
Figura 3.57. Los números 3 y 4 de la figura corresponden a los retículos endoplasmático rugoso y liso respectivamente.
Retículo endoplasmatico rugoso (RER): presenta ribosomas adosados a sus membranas, lo cual le confiere su carácter basófilo. Cuando las células presentan abundante RER se observa en la región donde este se localiza una intensa basofilia, se dice entonces que el RER se distingue al M/O por la basofilia localizada que le confiere al citoplasma. Este organito se localiza en la zona para basal en las células cilíndricas secretoras, mientras que en células no polarizadas ocupa una posición más bien excéntrica, como es el caso de las células plasmáticas.
Debido a que a esta estructura se adhieren los ribosomas durante la síntesis de proteínas y a que la variedad rugosa es mas abundante en los tejidos en los que tiene lugar una actividad importante de síntesis de proteínas –sumados a muchos otros datos experimentales- se le ha designado la síntesis de proteínas como su actividad primordial, participando tanto en la síntesis de proteínas exportable como de proteínas de membrana, como se verá más adelante (Fig. 3.58 y 3.59).
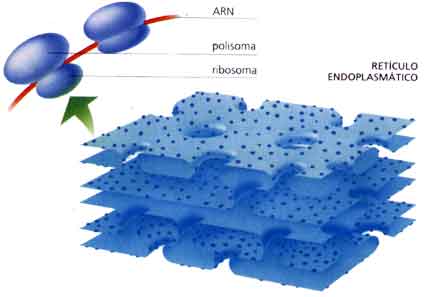
Figura 3.58. Imagen del retículo endoplasmático rugoso. En la parte superior se observan dos ribosomas unidos al ARN mensajero.
Figura 3.59. En el esquema puede verse la asociación de polirribosomas al retículo endoplásmico durante la síntesis de proteína.
Las proteínas que se sintetizan en los ribosomas del RER son proteínas de membrana, de secreción y proteínas que permanecen dentro de la célula para realizará funciones metabólicas. En resumen las funciones más importantes del RER son:
· Participa en las síntesis y maduración de proteínas exportables o de membrana: las proteínas que se sintetizan en los ribosomas adosados a su membrana pasan a la luz o lumen de éste orgánulo y comienzan su procesos de maduración (formación de puentes disulfuro, glicosilaciones, entre otras), y cuando tienen la estructura necesaria son transportadas en sus cavidades hasta salir hacia el aparato de Golgi mediante vesículas de transferencia.
Retículo endoplasmático liso: se localiza a continuación del retículo rugoso (Fig. 3.57). Su superficie es lisa y se caracteriza por túbulos membranosos. Nos se observa al microscopio óptico.
En las células hepáticas se vuelve muy abundante cuando se consumen algunas sustancias tóxicas o medicamentos, aumentando su capacidad para activarlas o inactivarlas; por ello se le ha relacionado con la función de “destoxificación”, dentro de las cuales se encuentran ciertos medicamentos y/o drogas no medicamentosas.
La abundancia de este sistema en algunos tejidos, como por ejemplo en parte de la glándula suprarrenal y en las gónadas (testículo y ovario), que se encargan de producir hormonas de las llamadas esteroides, ha demostrado que este organito participa en su síntesis. Participa también en la síntesis de los ácidos grasos, principales componentes de la mayoría de los lípidos.
En el músculo, el retículo endoplásmico liso tiene una función especial, relacionada con el almacenamiento y transporte del calcio. En este tipo de célula recibe el nombre de retículo sarcoplásmico (Fig. 3.60). se dispone de forma regular en relación con las micro fibrillas; esto, aunado al hecho de que posee una gran capacidad para transportar calcio, pone en evidencia su participación en la regulación de la contracción muscular.
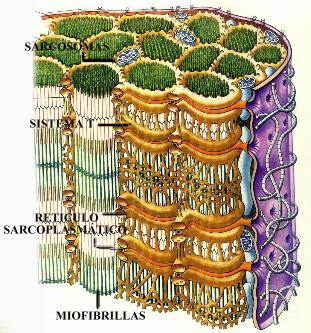
Figura 3.60 Retículo Sarcoplásmico del tejido muscular.
Aparato de Golgi
El aparato de Golgi (AG) es un orgánulo membranoso. Debe su nombre a Camillo Golgi, premio Novell de medicina en 1906. Al microscopio de campo brillante y utilizando como colorantes la hematoxilina y eocina, no se tiñe, originando una área no coloreada, o área de exclusión, que contrasta con el resto del citoplasma que si se colorea, por lo que a esta imagen se le llama imagen negativa del aparato de Golgi. Cuando se utilizan sales de planta presenta características argirófilas. Con ésta técnica se observa como una imagen acordonada carmelita, debido a que precipita las sales de planta. A esta se le llama imagen positiva del aparato de Golgi (Fig. 3.61).
Figura 3.61. Neuronas de ganglios cráneo-espinales teñidas con la técnica de impregnación argéntica. El núcleo se ve en blanco (imagen negativa) el aparato de Golgi se observa como filamentos pequeños alrededor del mismo.
El AG se localiza cerca del núcleo, asociado al centrosoma. En células secretoras se dispone en la región supranuclear (células secretoras cilíndricas), o alrededor del núcleo o peri nuclear, en neuronas y hepatocitos, siempre orientado hacia el polo secretor de la célula. El tamaño y desarrollo de estos orgánulos es variable, no sólo de una célula a otra, sino también con la actividad celular.
Cuando se observa al microscopio electrónico está formado por un conjunto de cisternas o sacos aplanados o dictiosomas (de 4 a 8 sacuelos aplanados rodeados de membrana y apilados unos encima de otros, formando una imagen en pila d moneda) cuyas partes laterales aparecen dilatadas. Está formado por tres tipos de estructuras membranosas: microvesiculas, provenientes del RER, un conjunto de cisternas aplanadas y por ultimo, vesículas grandes o vacuolas.
El AG es una estructura polarizada, es decir, tiene un lado diferente del otro los cuales describen dos caras o superficies: una convexa, cara cis o formadora, y otra cóncava, cara trans o de maduración. De este modo, el AG esta subdividido en tres regiones o fases: la región cis, orientada hacia el núcleo celular, la región intermedia y la región trans o fase de maduración. En la cara cis se encuentran las vesículas de transferencia, mientras q en la cara trans se localizan las vacuolas de secreción.
Las proteínas que vienen del RER entran al AG en su fase sis o su fase de entrada por endocitosis de las vesículas transportadoras q fusionan su membrana con la membrana de la fase cis. Posteriormente por gemación, las proteínas salen de la fase sis y entran ala intermedia nuevamente por gemación salen de la fase intermedia y van hacia la fase trans y finalmente también por gemación, salen en vesículas grandes electro densas y envueltas por una membrana que da origen a los gránulos de secreción que saldrán a su destino final. El destino final de esta vacuola puede ser la membrana plasmática o puede fusionarse con esta y puede verter su contenido al exterior por exostosis (Fig.3.62)
Funciones del AG:
Es pertinente aclarar que el proceso de glicosilación, que la mayoría de las veces se indica en el retículo endoplásmico, posee suma importancia, pues permite darle ala molécula procesada propiedades especiales. En el caso de las proteínas, por ejemplo, su glicosilación da lugar a los componentes básicos del glicocálix, que posee un papel fundamental en procesos de reconocimiento, comunicación celular y transducción de señales. En otros casos permite otorgarle ala molécula una resistencia mecánica adicional, como es común cuando se trata de hormonas o mensajeros a distancia.
Por ultimo, es conveniente señalar que una de las funciones mas importantes de las vesículas es transportar materiales hacia la membrana plasmática y desde ella hacia el interior de la célula; constituyen de este modo un medio de comunicación entre el interior celular y el medio externo hay un intercambio continuo de materiales entre el retículo endoplásmico, el aparato de golgi, los lisosomas y el exterior celular. Dicho intercambio esta mediado por pequeñas vesículas delimitadas por membrana que se forman por gemación a partir de una membrana y se fusionan con otra.
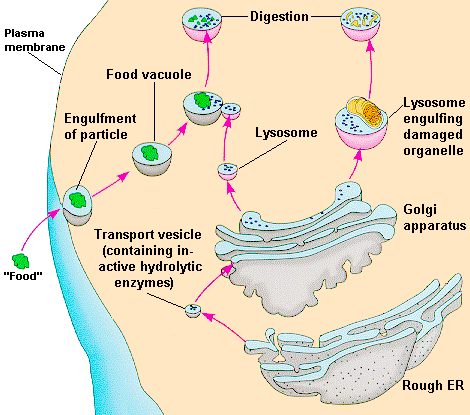
Lisosomas
Los lisosomas son orgánulos membranosos cerrados, constituidos por una solo membrana. Se pueden obtener en estado de pureza por métodos especiales de centrifugación que permiten separarlos de las mitocondrias, pues en los métodos generales de preparación se obtienen juntos. Estos organelos, si se les rompe colocándolos en agua o por medio de algún detergente, ponen en evidencia una serie de actividades enzimáticos muy diversas, capaces de romper por hidrólisis (introduciendo en algunos enlaces moléculas de agua) lípidos, carbohidratos, ácidos nucleicos, entre otros.
Funciones de los lisosomas: se considera que estos organelos representan los elementos necesarios para degradar compuestos intracelulares en caso necesario, o de material proveniente del exterior de la célula, por el alto contenido de enzimas hidrolasas que poseen. Por ultimo, constituyen una de las vías utilizadas en la producción de hormonas, como es el caso de hormonas tiroideas.
Los lisosomas son visibles al microscopio de campo brillante mediante el uso de técnicas histológicas como el azur y técnicas histoquímicas para la fosfatada ácida. Pueden ser clasificados de la siguiente forma:
Lisosoma primario. Lisosoma recién sintetizado y que acaba de salir por la fase trans del aparato de golgi. Contiene Blas enzimas hidroliticas o enzimas lisosomales y aun no se ha puesto en contacto con ningún otro cuerpo membranoso.
Lisosoma secundario. Este tipo de lisosoma resulta de la unión de de un lisosoma primario con otro cuerpo membranoso surgido a partir de elementos provenientes del exterior de la célula o del interior de esta.
Heterofagosoma. Es un lisosoma secundario que se forma por la unión de un lisosoma primario con una vesícula membranosa proveniente de la membrana plasmática y que contiene en su interior sustancias que han penetrado ala célula por endocitosis y en la cual tiene proceso de digestión celular.
Citolisosoma. Resulta de la unión de un lisosoma primario con un cuerpo membranoso surgido dentro de la célula y que contiene restos celulares en su interior como mitocondrias envejecidas, entre otras.
Cuerpo residual. Es el cuerpo membranoso que contiene los restos del proceso digestivo ya ocurrido en los casos anteriores, contenido es generalmente expulsado por exocitosis. En ocasiones el producto no es de desecho, sino una sustancia útil para el organismo, tal es el caso de las hormonas tiroideas que serán estudiadas en el Sistema Endocrino en la asignatura de Morfofisiología III. En algunos casos los cuerpos residuales permanecen en el interior de la célula sin ser expulsados, como los que contienen pigmentos de Lipofuscina (Fig. 3.63).
Peroxisomas.
Son organismos membranosos, con una membrana única, cuyo diámetro es de 0.5 a 1.2 μm. En estos orgánulos se degradan las purinas y otros compuestos. En los peroxisomas se produce agua oxigenada (H2O2), compuesto muy tóxico para la célula que es degradado rápidamente por una enzima. Entre las enzimas que posee el peroxisoma se encuentra la peroxidasa y la catalasa; esta última degrada al H2O2.
2 H2O2 >>>>>>>à 2 H2O = + O2
catalasa
La actividad de la catalasa tiene gran importancia médica, porque bajo su acción muchas moléculas tóxicas, incluyen medicamentos, pueden ser transportadas por los peroxisomas del hígado y de los riñones. Aproximadamente el 50% del alcohol etílico ingerido es transformado por los peroxisomas del hígado y los riñones.
Otra de la funciones de los peroxisomas es la a oxidación de los ácidos grasos, actividad que realizan conjuntamente con las mitocondrias.
MITOCONDRIAS.
Son organismos que participan entre otras en la síntesis de enlaces ricos en energía, y dada su importancia para la célula seran analizadas cuando se trate el proceso de la respiración celular.
ORGANELOS NO MEMBRANOSOS.
RIBOSOMAS.
Los ribosomas son organitos u orgánulos no membranosos que participan en la síntesis de proteínas. Se pueden observar al microscopio óptico por características basófilas. Dicha basofilia está dada por los grupos fosfato que posee el ARN.
Los ribosomas pueden ser clasificados en libres o unidos a membranas. En el caso de los ribosomas libres la basofilia que confieren al citoplasma es difusa, por ellos se disponen de forma mas o menos azarosa. En el caso de los ribosomas unidos a membranas (formando el RER), confieren al citoplasma basofilia localizada, como ya se expreso en párrafos anteriores.
La función de los ribosomas libres es la síntesis de proteínas estructurales de la célula, mientras que los unidos a membranas sintetizan proteínas exportables y de membrana, como fue explicado con relación al RER.
A los ribosomas se les encuentra tanto en células eucariotas, donde poseen un coeficiente de sedimentación de 80s, como en células procariotas, donde su coeficiente de sedimentación es de 70s. Como se puede apreciar, los ribosomas de las células eucariotas son ligeramente mas grandes que los de las procariotas.
La estructura de los ribosomas consiste en dos subunidades: una subunidad mayor y otra menor (Fig. 3.64). Su composición macromolecular está dad por ARNr y cerca de 50 proteínas estructurales.
Centríolos.
Son organelos no membranosos, pares, pequeños que se encuentran cerca del núcleo de las células, y tienen capacidad de duplicarse antes de que se inicie la división celular, es decir en la etapa G2 del ciclo celular. En las células ciliadas o flageladas, la duplicación continuada de los centríolos representa el origen de los cuerpos basales, que dan lugar a los cilios y flagelos y a sus llamados centros cinéticos o de movilización; de alguna forma los centríolos están implicados en el movimiento de estos componentes de la célula.
Los centríolos son visibles al microscopio óptico con técnicas histológicas especiales. Son organelos que presentan una cavidad central, y una pared formada por nueve tripletes de microtúbulos, llamados A, B y C, inmersos en un material amorfo (Fig. 3.65). No tienen microtúbulos centrales como ocurre con los cilios. Al microscopio electrónico se observan como muestra la figura 3.66.
Inclusiones celulares.
A menudo pueden observarse en el citoplasma depósitos transitorios constituidos por una reserva de nutrientes u otras moléculas, a los cuales se les denomina inclusiones citoplasmáticas. De este modo, las inclusiones citoplasmáticas no son más que estructuras que existen en el citoplasma y que constituyen verdaderos almacenes de sustancias específicas disponibles para ser utilizadas por las células del organismo.
Se pueden clasificar las inclusiones celulares de la siguiente forma:
Nutrientes almacenados.
Entre estos se encuentran el glucógeno y la grasa.
Glóbulos de grasa. Estas inclusiones se encuentran en las células del tejido adiposo, son cúmulos de lípidos del tipo de los Triacilgliceroles (grasas neutras o ésteres de ácidos grasos y glicerol), que se explicaran más adelante. Ellos son reserva de energía muy eficiente, debido al elevado contenido calórico que posee esta clase de lípidos; en los adipositos pueden llegar a ocupar el 90% del volumen celular.
No obstante, estas inclusiones aparecen en otras células del organismo, como las musculares y los hepatocitos. En los hepatocitos pueden acumularse excesivamente y causar daño bajo determinadas circunstancias; en el denominado hígado graso que en algunos casos puede evolucionar hacia la Cirrosis Hepática, como sucede en los individuos alcohólicos (Fig. 3.67).
Los glóbulos de grasa no se observan cuando se utilizan técnicas histológicas corrientes pues la grasa se disuelve con los alcoholes y el Xilol utilizados en el método de inclusión en parafina, por lo que para poder observarla en necesario utilizar cortes por congelación y los métodos de coloración de Sudán; de esta forma se puede decir que las células poseen glóbulos de grasa son sudanófilas.
Características de los acilgliceroles. Conocidos antes como glicéridos, son ésteres de ácidos grasos con glicerol. En dependencia del número de ácidos grasos esterificados pueden ser monogliceroles, diacilgliceroles y triacilgliceroles (o grasas neutras, antes conocidas como triglicéridos) (Fig. 3.68).
Loa triacilglicéroles son los lípidos mas abundantes de la naturaleza, constituyen una fuente importante de energía en el organismo y es la forma de almacenamiento en el tejido adiposo. Loa acilgliceroles por sus características estructurales son moléculas apolares y sus propiedades físicas dependen del tipo de ácidos grasos esterificados. Los triacilgliceroles, cuyos ácidos grasos son de cadena larga y saturada, son sólidos a temperatura ambiente (mantecas); en tanto que si sus ácidos grasos son saturados de cadena corta (menos de 10 carbonos) o insaturados, son líquidos a la temperatura ambiente (aceites).
Por hidrólisis en medio ácido los acilgliceroles dan origen a glicerol y ácidos grasos o glicerol y sales de sus ácidos (jabones) si el medio es alcalino.
Funciones de los triacilgliceroles:
Gránulos de glucógeno. Muchas veces se encuentran otras reservas energéticas en hidratos de carbono en forma de gránulos de glucógeno. Al microscopio óptico el glucógeno no se colorea con las técnicas corrientes de Hematoxilina y Eosina, apareciendo con un aspecto blanco en los cortes teñidos con estos colorantes. El glucógeno se tiñe con la técnica de PAS, brindando un color rojo magenta. Al microscopio electrónico el glucógeno se presenta como un agregado de pequeñas partículas electrón denso.
En el gránulo, además se encuentran las enzimas que participan en la síntesis y degradación del polisacárido, así como en la regulación del metabolismo del glucógeno. Tanto el peso molecular del glucógeno como la proporción en que se hayan la proteínas enzimáticas en el gránulo son variables, de modo que estas inclusiones poseen diámetros que van desde 1000 hasta 4000 nm.
El glucógeno es el homopolisacárido de reserva más importante en las células animales. El almacén del glucógeno es limitado siendo más importante en el tejido hepático, donde representa del 10 al 12% de su peso húmedo, y en el músculo esquelético, donde es de hasta el 2%.
Pigmentos.
Los pigmentos se clasifican en dos grupos: exógenos y endógenos.
Pigmentos exógenos: Son aquellos que formados fuera del organismo son incorporados a las células por una u otra vía. Entre ellos se encuentran: carotenos, polvos, minerales y marcas de tatuaje.
Los carotenos con un tipo de pigmento formado en varios tipos de vegetales. Son sustancias solubles en grasas, por lo que también se les denomina lipocromos. Algunas formas de caroteno son pro vitaminas que se convierten en Vitamina A. Un consumo excesivo de alimentos ricos en caroteno (zanahoria, tomates, etc.) pudiera proporcionar un color amarillento a la piel, aunque esto es algo poco frecuente.
Los polvos se inhalan por vía respiratoria y pueden producir una pigmentación característica en el tejido respiratorio (antracosis por carbón). La ingestión o absorción de minerales (plata o plomo) por la superficie corporal, puede producir en determinados sitios la acumulación de estas sustancias, dando una coloración a la piel y/o mucosas.
Pigmentos endógenos: Entre los pigmentos endógenos el más importante es la hemoglobina presenta en los eritrocitos, que es la responsable del color rojo de la sangre. En los eritrocitos “viejos”, los cuales son eliminados en el hígado, el bazo y la médula ósea, la hemoglobina es desintegrada en dos compuestos pigmentados, la hemosiderina que contiene hierro y otro que no contiene hierro, la bilirrubina. La hemosiderina se encuentra en los fagocitos de los órganos mencionados y la bilirrubina forma parte de la bilis, sustancia segregada por el hígado y que también se almacena en la vesícula biliar.
Otro de los pigmentos endógenos es la melanina, compuesto químico que le da color a la piel, a sus anexos y a los ojos. La melanina es de color pardo a negro y es producido por unas células denominadas melanocitos.
La lipofucsina es otro pigmento que tiene lípidos en su constitución y presenta en estado natural un color parduzco. Se ha observado en los cuerpos residuales de células nerviosas; es mas abundante en las personas de edad avanzada, por lo que se considera a la lipofucsina como un material de desgaste que no ha podido se eliminado de las células.
El citosol.
El citosol no es un organelo, ni puede considerarse como tal; sin embargo, se debe tener presente que no se trata de un simple ambiente inerte que sirva sólo de asiento a los organelos y otras estructuras celulares. El citosol es en primer lugar el componente mas externo de la célula y contiene una cantidad enorme de enzimas, muchas de la cuales funcionan de manera concertada para constituir vías metabólicas. Por otra parte, el citosol es el paso obligado en el camino de tantos miles de moléculas que van de uno a otro componente de la célula.
Entre los caminos metabólicos que tienen lugar en el citosol se encuentra la glucólisis, que es una serie larga de reacciones que convierten a la glucosa en ácido pirúvico y es ahí donde tienen lugar los cambios necesarios para llevar a muchas moléculas hacia el ciclo de los ácidos tricarboxílicos. Cuando las proteínas o parte de sus componentes se convierten en azúcares, como sucede durante períodos de ayuno prolongados, utilizan gran parte de la misma vía en un proceso que se llama gluconeogénesis, que también tiene lugar en el citosol.
La síntesis de los ácidos grasos sigue un camino que está organizado como un complejo multienzimático (supramolecular) y que se encuentra en el citosol.
Las fases preparatorias para utilizar los aminoácidos en la síntesis de las proteínas se realizan en el citosol. Estos con solo unos cuantos de los cientos de caminos metabólicos que se siguen para producir los varios miles de moléculas que constituyen a las células.