3.2 SISTEMA DE MEMBRANAS
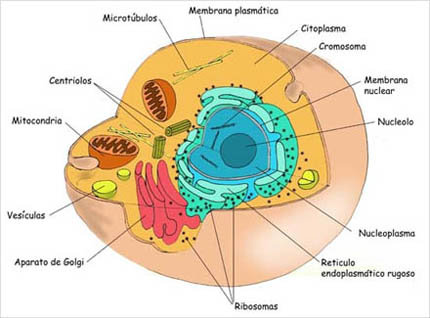
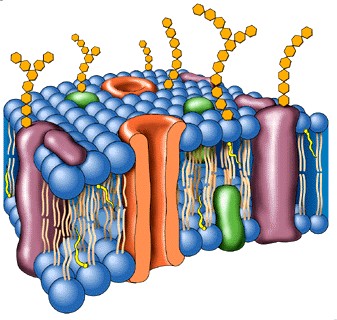
Las membranas celulares son esenciales para la vida de la célula. Las mismas pueden rodearla (membrana plasmática) o pueden localizarse en su interior (sistema de membranas), contribuyendo a la compartimentalización celular y por lo tanto rodeando orgánulos celulares u otros componentes membranosos (fig. 3.8).
La membrana plasmática envuelve a la célula, definiendo sus límites y manteniendo las diferencias esenciales entre su contenido y el entorno extracelular. El sistema de membranas intracelulares comprende las membranas que rodean a orgánulos tales como: el retículo endoplasmático, el complejo de Golgi, la mitocondria, los lisosomas y otros orgánulos membranosos. Estas membranas mantienen las diferentes características entre el contenido de cada orgánulo y el citosol.
A pesar de que realizan diferentes funciones, todas las membranas biológicas comparten una estructura molecular básica común: una bicapa lipídica, proteínas unidas por interacciones no covalente y oligosacáridos en mayor o menor proporción.
BICAPA LIPÍDICA
Lípidos de las membranas: Los lípidos de las membranas pertenecen a tres categorías: los fosfátidos de glicerina o glicerofosfátidos, los esfingolípidos y el colesterol.
Fosfátidos de glicerina o glicerofosfátidos
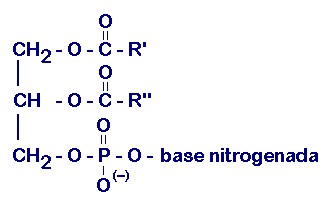
Los fosfátidos de glicerina o de glicerol son lípidos complejos saponificables y anfipáticos. Están formados por glicerol, 1 ó 2 residuos de ácidos grasos y un grupo fosfato; además, pueden contener, en dependencia del tipo, otros compuestos.
Los fosfátidos de glicerina pueden ser:
§ Fosfatidil serinas.
§ Fosfatidil entanolaminas.
§ Fosfatidil colina.
§ Fosfatidil inositoles.
§ Fosfatidil gliceroles y difosfatidilgliceroles ( cardiolipinas).
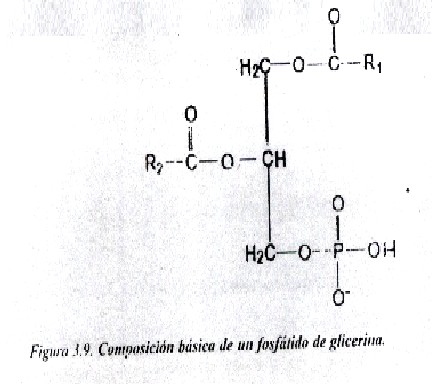
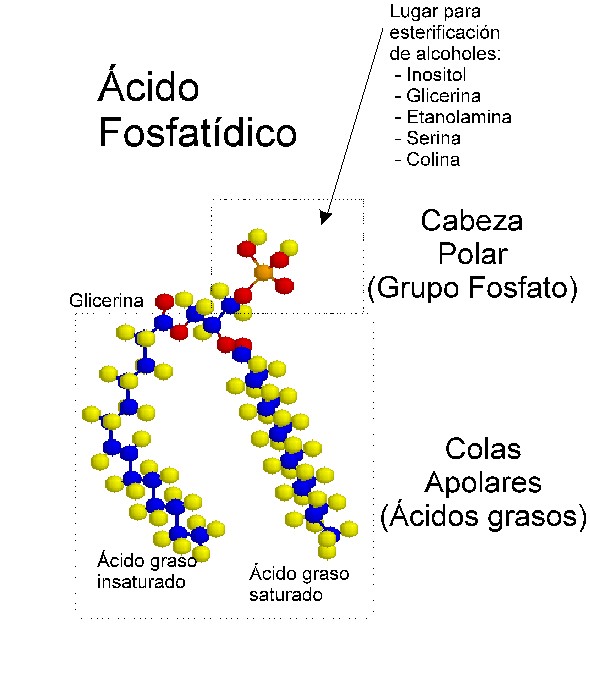
Ácidos fosfatídicos: la base estructural de la mayoría de los fosfátidos de glicerina es el ácido fosfatídico; este último apenas se encuentra en forma libre en los tejidos animales, aunque constituye la estructura básica del resto de los fosfátidos de glicerina y metabólicamente es precursor de su síntesis, e incluso un intermediario de la síntesis de los triacilgliceroles. Tiene eterificado al glicerol 2 o más ácidos grasos, uno de ellos casi siempre insaturado, y un grupo fosfato. Estos compuestos al igual que todos sus derivados presentan isomería óptica, ya que poseen asimetría molecular. La variedad más frecuente en los fosfátidos naturales es la L, que se presenta con el residuo del ácido graso del carbono 2 (ó á) hacia la izquierda y el grupo fosfato en el carbono 3 (ó á) (fig. 3.9).
Al unírsele al ácido fosfático la serina, la etanolamina, la colina y el inositol se originaran los fosfátidos de glicerina correspondientes.
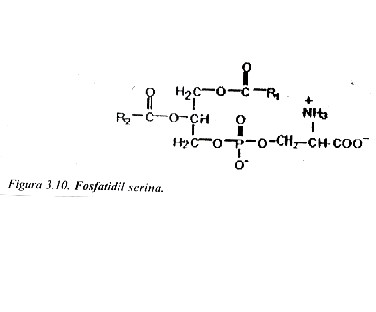
Fosfatidil serinas: En estos lípidos el ácido fosfático se une a la L-serina por esterificación de hidroxilo de la cadena lateral de este aminoácido con el grupo fosfato (fig. 3.10). Estos lípidos tienen carga negativa y se localizan de preferencia en la monocapa interna de la membrana.
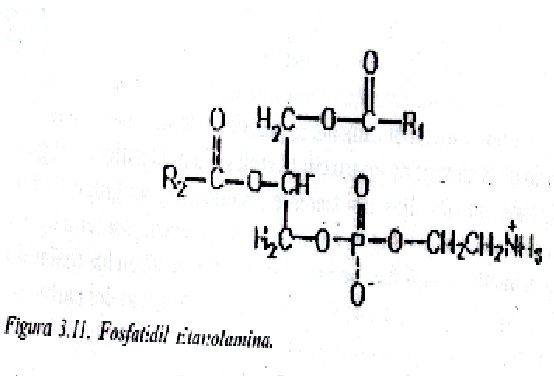
Fosfatidil etanolaminas: El sustituyente básico es el alcohol etanolamina (fig. 3.11). La etanolamina se forma por descarboxilación de la serina; junto a las fosfatidil serinas forman el grupo de la cefalinas. Estos lípidos se localizan en mayor proporción en la superficie interna de la membrana.
Fosfatidil colina: La base nitrogenada colina se une al ácido fosfórico por un enlace éster. Estos compuestos constituyen las lecitinas. Estos lípidos tienen carga es 0 y se localizan de preferencia en la parte externa de la membrana. En la fig. 3.12 puede apreciarse un modelo de la disposición espacial de este tipo de lípido.
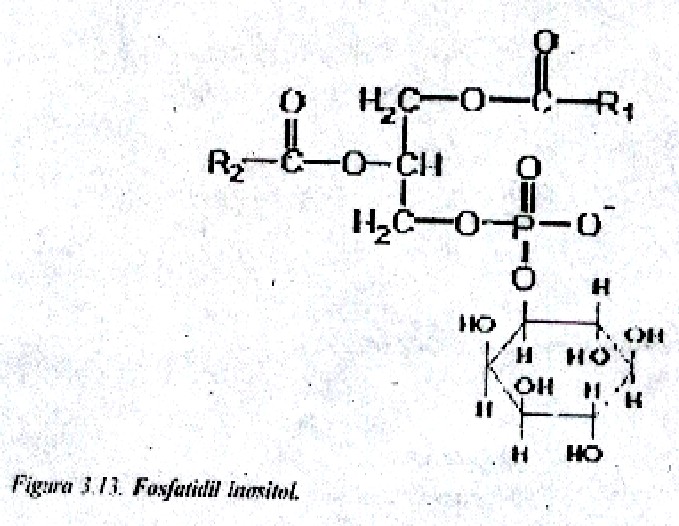
Fosfatidil inositoles o inositofosfátidos: Son fosfátidos de glicerina no nitrogenados, que contienen un alcohol cíclico de 6 átomos de carbono y cuya forma isomérica en los tejidos animales es el mioinositol o mesoinositol (fig.3.13). Se localizan en la superficie interna de la membrana.
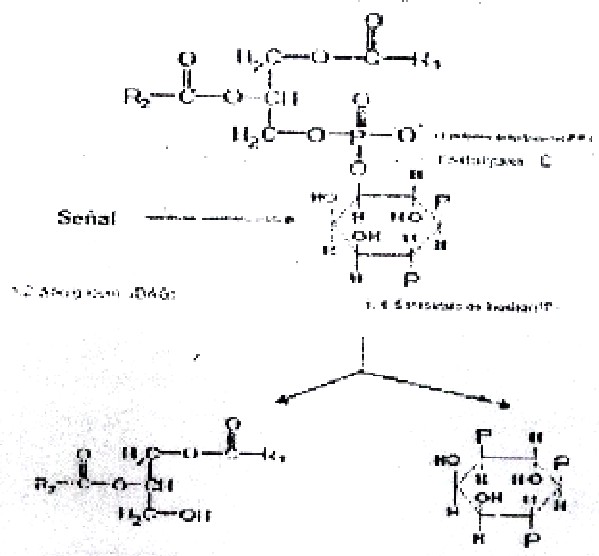
El derivado 4,5 bifosfato de fosfoinositol (PIP2) es un constituyente importante de los fosfolípidos de membrana, los que por la acción de determinados agonistas hormonales se escinde dando 1,2 diacilglicerol (DAG) y 1,4,5 trifosfato de inositol (IP3), los caules constituyen segundos mensajeros en la respuesta hormonal (fig. 3.14).
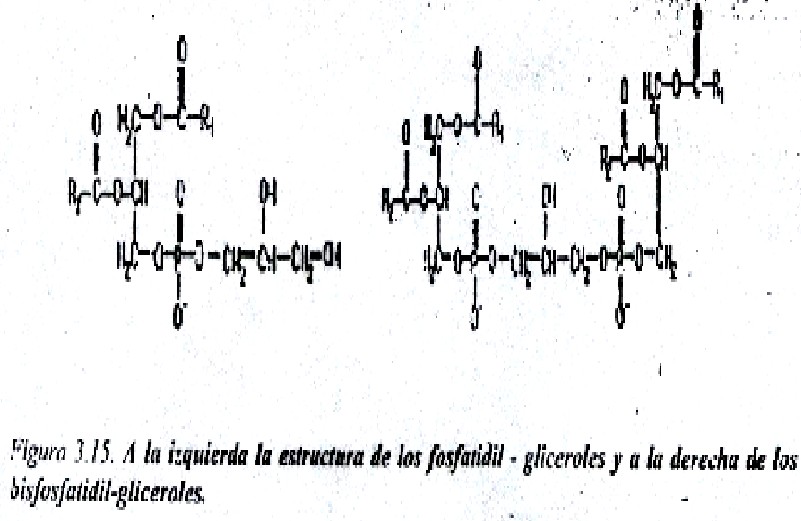
Bifosfatidil-gliceroles (cardiolipinas) y fosfatidil-gliceroles: Estos compuestos tampoco poseen nitrógeno; en su estructura contienen residuos de ácido fosfatídico que se eterifican a grupos OH del glicerol (fig. 3.15). La cardiolipina es abundante en la membrana interna de la mitocondria.
Funciones de los fosfátidos de glicerina.
1. Son componentes de las membranas celulares.
2. Los ácidos fosfatídicos son precursores de la síntesis de otros fosfátidos de glicerina y de los acilgliceroles.
3. Las fosfatidil colinas y fosfatidil inositoles son donadores de ácido araquidónico para la síntesis de prostaglandinas, tromboxanos, prostaciclinas, leucotrienos y otros compuestos relacionados.
4. Los compuestos formados a partir de un derivado del fosfatidil inositol (4,5 bifosfato de fosfatidil inositol): el diacilglicerol (DAG) y el trifosfato de inositol (IP3) actuán como segundos mensajeros de la acción hormonal.
Esfingolípidos
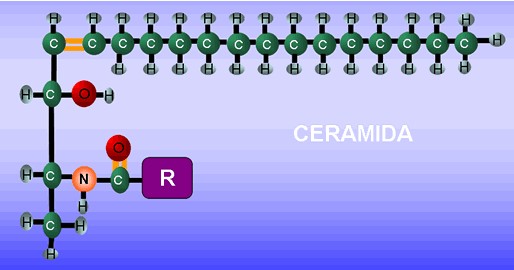
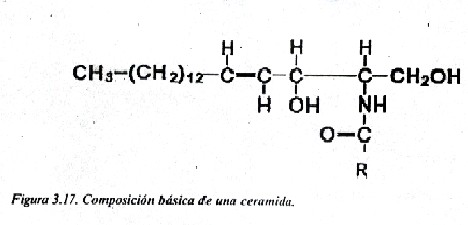
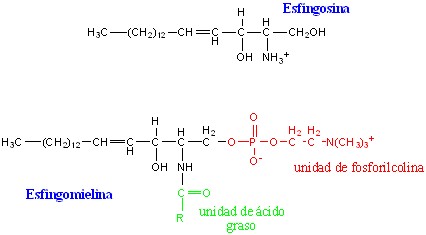
Los Esfingolípidos son lípidos complejos que contienen un alcohol nitrogenado e insaturado de 18 átomos de carbono, el esfingol o esfingosina (fig.3.16). Al esfingol se le une un ácido graso por el enlace amida, formando la ceramida (fig. 3.17), estructura básica de los esfingolípidos.
A la ceramida se le adicionan otros compuestos en dependencia del tipo de esfingolípido. Los esfingolípidos se clasifican en esfingomielinas y glicoesfingolípidos.
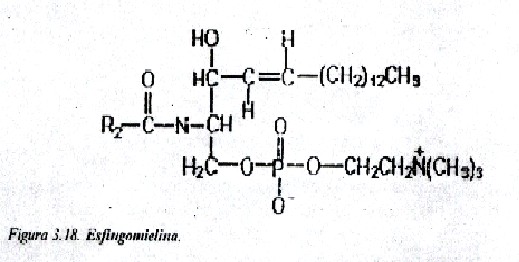
Esfingomielinas (fosfátidos de esfingosina): Estos lípidos contiene un grupo fosfato que se une por enlace éster al hidroxilo del carbono 1 de la ceramida, y también una molécula de colina esterificada al fosfato. La fuente de variación de estos compuestos radica en el ácido grado unido (fig. 3.18).
Con frecuencia se designan como fosfolípidos a los fosfátidos de glicerina y a las esfingomielinas, por su contenido de fósforo.
Glicoesfingolípidos: Estos compuestos, conocidos también como glicolípidos, carecen de grupo fosfato en el carbono 1 de la ceramida, y en su lugar se le une un glúcido, que puede ser un mono u oligosacárido. De acuerdo con el tipo de glúcido que contenganlos glicoesfingolipidos pueden ser: cerebrósidos, sulfolípidos o gangliósidos.
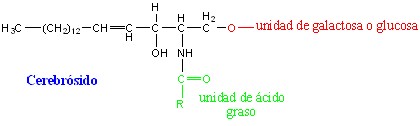
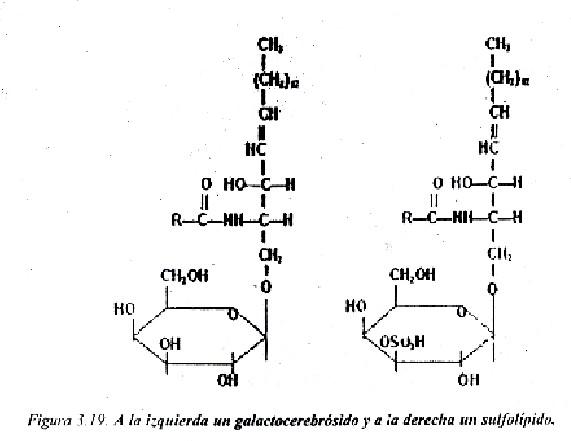
Cerebrósidos: el glúcido unido a la ceramida es un monosacárido, que puede ser la D-galactosa (galactocerebrósido) o de la D-glucosa (glucocerebrósido) (fig. 3.19).
Sulfátidos o sulfolípidos: constituyen derivados de los cerebrósidos a los que se les ha añadido un grupo sulfato al carbono 3 del monosacárido (fig. 3.19).
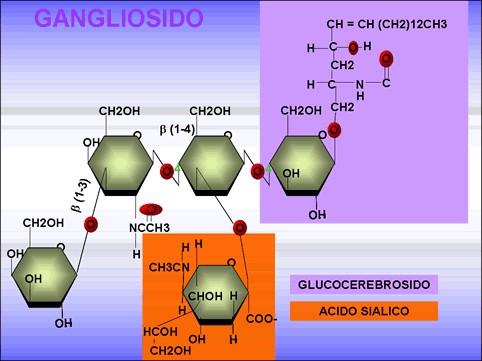
Gangliósidos: son esfingolípidos que contienen oligosacáridos como residuo glucídico. El oligosacárido está formado por diversos monosacáridos y uno o más residuos de ácido N-aciltilneuramínico o ácido siálico (fig. 3.20).
Los esfingolípidos son también anfipáticos; su porción polar corresponde a los sustituyentes del carbono 1 de la ceramida (grupo fosfato y colina en las esfingomielinas, y los glúcidos en los glicoesfingolipidos), en tanto que su porción apolar la conforman las cadenas hidrocarbonadas del ácido graso y del esfingol. Las glicoesfingolípidos son los lípidos más solubles en agua debido a su contenido glucídico.
Funciones de los esfingolípidos
1. Forman parte de las membranas biológicas. Se encuentran en grandes cantidades en la sustancia blanca del sistema nervioso central.
2. Las esfingomielinas son compuestos de las vainas de mielina de las fibras nerviosas.
3. Algunos glicoesfingolípidos por su carácter informacional le confieren acción antigénica a la superficie de algunas células, lo que contribuye al reconocimiento molecular de estas.
4. Los cerebrósidos y sulfátidos forman parte de órganos como el cerebro, los nervios, el bazo, los riñones entre otros.
5. Los gangliósidos aparecen en células ganglionares del sistema nervioso y en tejidos no nerviosos.
6. Se les atribuye participación en la transmisión del impulso nervioso.
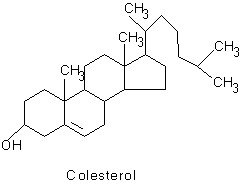
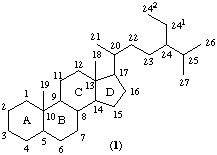
La característica más sobresaliente de los esteroides, y que es común para todos ellos, es la presencia del sistema policíclico: ciclo-pentano-perhidrofenantreno. De acuerdo con la cadena lateral unida al carbono 17 y a diferentes sustituyentes e insauraciones, se forman los diferentes esteroides. Los esteroides se pueden clasificar en: esteroles, ácidos biliares, corticoesteroides, progesterona, andrógenos y estrógenos.
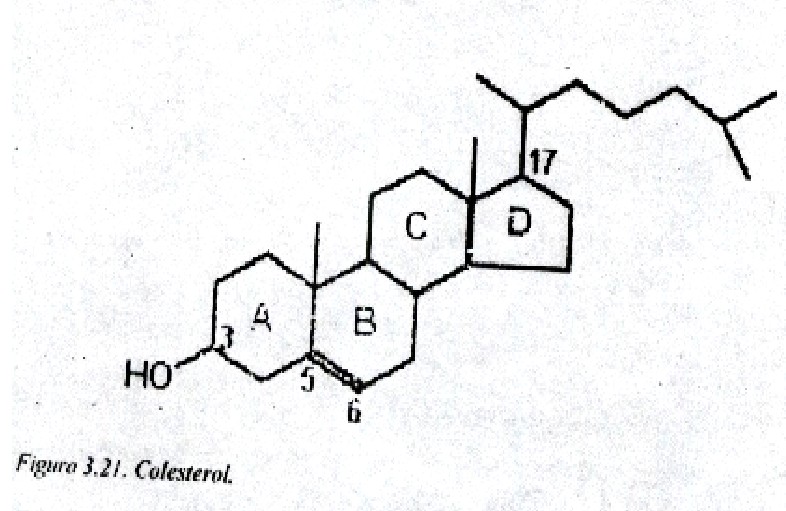
En este capitulo se hará referencia a los esteroles. En estos compuestos la cadena lateral unida al carbono 17 puede contener 7, 8 ó 9 átomos de carbono y por ello se originan los esteroides C26 C27 C28, respectivamente; poseen, además, un hidroxilo en la posición 3. Entre estos tipos de lípidos se encuentra el colesterol, el cual es un lípido de membrana y precursor de otros esteroides (fig. 3.21).
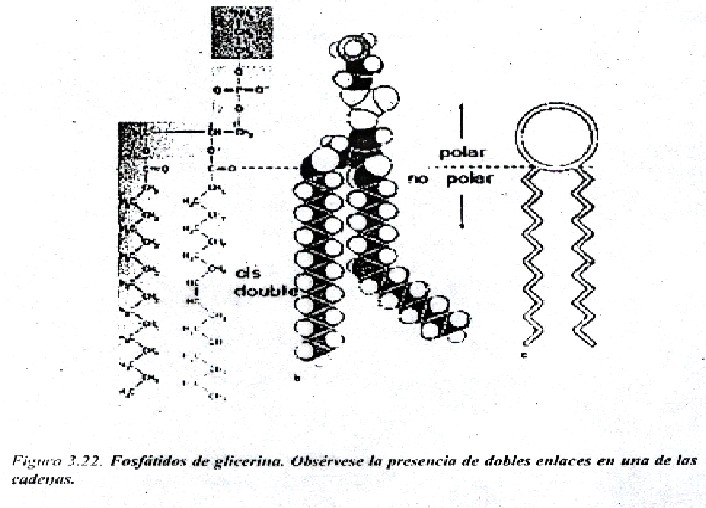
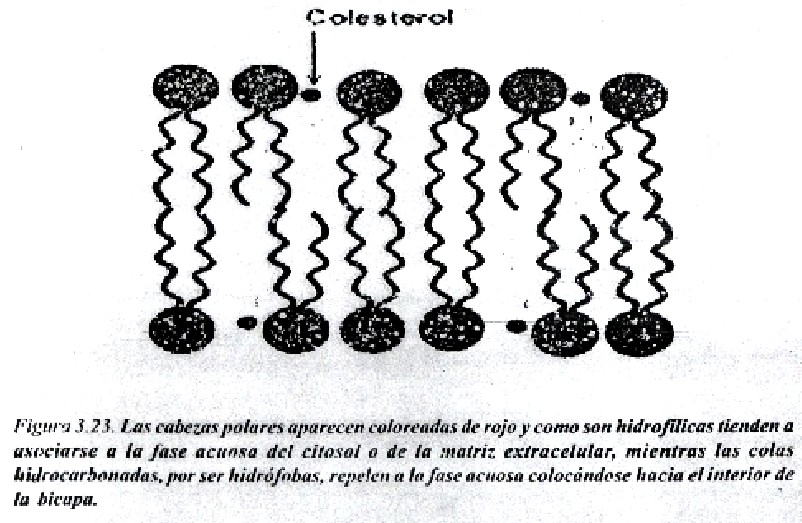
La bicapa lipídica ha sido establecida coma la base universal de la estructura de la membrana celular. Es fácil de observar al microscopio electrónico (M/E), aunque para revelar sus detalles se requiere de técnicas especializadas como la difracción de rayos X y la criofractura. Se estructura en bicapa se debe a las propiedades antipáticas de los lípidos que la forman. El carácter anfipático de los lípidos se debe a que poseen un extremo polar o hidrofílico (formado por las cabezas polares de los fosfátidos de glicerina y esfingolípidos y el grupo OH del colesterol), y otro apolar o hidrófobo, dado por las colas hidrocarbonadas del resto de las estructuras. Las colas hidrocarbonadas de los fosfátidos de glicerina, correspondientes a los ácidos grasos esterificados al glicerol, varían en longitud (14-24 átomos de carbono); una cadena es insaturada presentando doble enlace (cis) y la otra es saturada. Tales diferencias en longitud y saturación son importantes porque ello influye en la fluidez de la membrana. Los lípidos con estas características forman bicapas espontáneamente (Fig. 3.22 y 3.23).
La proporción de los lípidos varía entre las diferentes membranas, pudiendo llegar a constituir el 79% de las vainas de mielina y el 50% de la membrana plasmática de la mayoría de las células animales.
Otra característica de gran importancia de los lípidos de membrana es que ellos se distribuyen de forma asimétrica en ambas monocapas; de esta forma en la monocapa externa se localizan de preferencia las fosfatidil colinas y los esfingolípidos, en tanto que en la monocapa interna se localizan las fosfatidil serinas, las fosfatidil etanolaminas y los fosfatidil inositoles.
PROTEÍNAS DE LAS MEMBRANAS
Aunque la estructura básica de las membranas biológicas está dada por la bicapa lipídica, la mayoría de las funciones de las membranas son desempañadas por proteínas. Son las proteínas las que confieren a las membranas sus propiedades funcionales, entre las que se encuentran las siguientes:
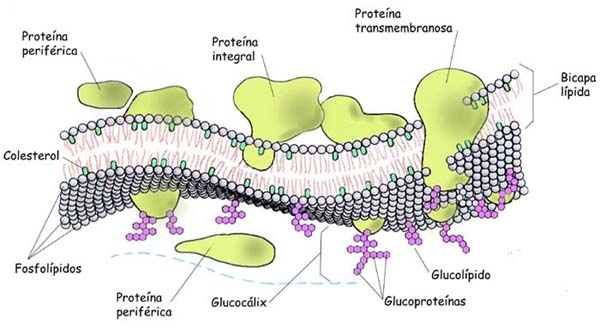
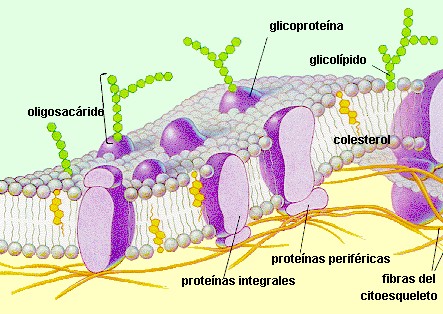
Según su localización en la membrana, las proteínas se clasifican en periféricas o extrínsecas e integrales o intrínsecas. Las proteínas de membranas, al igual que los lípidos, tienen distribución asimétrica.
Las proteínas integrales de membrana constituyen mas del 70% del total y se encuentran parcial o totalmente incluidas dentro de las membranas, por tanto, sólo pueden ser extraídas por medios drásticos como es el uso de detergentes o solventes orgánicos; estas proteínas son casi siempre insolubles en solventes polares y pueden disponerse de lado a lado de la bicapa lipídica, y en ese caso de denominan proteínas transmembranales. En la disposición de las proteínas intrínsecas se puede apreciar la orientación de los residuos de los aminoácidos apolares hacia la zona de la molécula incluida dentro de la membrana, con predominio de la estructura en hélice, y una localización de los radicales de aminoácidos polares hacia la superficie en la zona que queda por fuera de la bicapa. Las proteínas Integrales de las membranas al igual que los lípidos, tienen características antipáticas. Los receptores de membrana, las proteínas transportadoras, las proteínas que conforman canales y muchas enzimas son proteínas integrales.
Las proteínas periféricas se disponen en la superficie interna o externa de la bicapa y se unen, por interacciones débiles de tipo electrostático, a la cabeza polar del los lípidos de la membrana, por lo cual son fácilmente separadas con el uso de soluciones salinas. Estas proteínas son solubles en solventes polares.
GLÚCIDOS DE MEMBRANA
En todas las células eucariotas se encuentra una cantidad de glúcidos formando parte de las membranas, particularmente de la plasmática. Las glúcidos se hallan unidos de forma covalente a lípidos o proteínas para formar glicolípidos o glicoproteínas, respectivamente. La fracción glucídica constituye entre 2 y 10 %; desde el punto de vista estructural son oligosacáridos, que en su mayoría están unidos a proteínas y en menor cantidad a los lípidos. Los oligosacáridos de las membranas están dispuestos hacia la cara no citoplasmática y cumplen las funciones siguientes:
1. Contribuyen a la orientación de las proteínas de las membranas.
2. Participan en la interacción entre membranas de células distintas.
3. Tienen una función fundamental en las propiedades inmunológicas de las membranas.
Los azucares que se encuentran formando parte de estos glicolípidos y glicoproteínas son: glucosa, galactosa, manosa, mucosa, N-acetil-galactosalina y el ácido siálico. La asimetría se mantiene debido a que el paso espontáneo de moléculas de una capa a la otra es casi nulo.
MODELO DEL MOSAICO FLUIDO
El modelo de membrana conocido como “Modelo en Mosaico Fluido” fue propuesto por Singer y Nicolson en 1972. Este modelo es capaz de explicar numerosas propiedades físicas, químicas y biológicas de las membranas; se acepta como la organización estructural más probable de los componentes de las membranas biológicas. El modelo se propuso sobre la base del estudio de la composición química y propiedades físicas de las membranas biológicas, así como de resultados experimentales en los cuales se comparó el comportamiento de membranas sintéticas preparadas en el laboratorio, a partir de una mezcla de fosfolípidos y algunas proteínas, con el comportamiento de las membranas naturales.
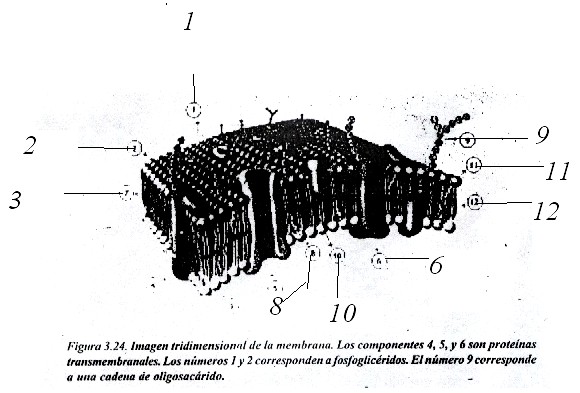
Según este modelo, se considera que las proteínas forman un mosaico dentro de la bicapa lipídica, que constituye la estructura básica, con glúcidos dispuestos hacia la cara no citoplasmática; el conjunto adopta una estructura tridimensional compacta y flexible (fig. 3.24).
La fluidez de la membrana esta dada por la facilidad de los lípidos y las proteínas de trasladarse dentro de la bicapa. El movimiento de las proteínas está mucho más restringido debido a su asociación con los elementos del citoesqueleto. El movimiento transmembranal o flip-flop es mucho más lento que el movimiento lateral. El colesterol juega un papel importante en la fuildez de la membrana. El grado de fluidez de una membrana influye en sus funciones: si aumenta su fluidez se incrementa su permeabilidad al agua y a otras moléculas e iones, en tanto que se obtiene un efecto contrario se disminuye la fluidez.
Resumiendo, el modelo del mosaico fluido considera:
1. Los lípidos y proteínas organizados en forma de mosaico, con los glúcidos unidos hacia la cara no citoplasmática.
2. Las membranas son estructuras fluidas donde los lípidos y proteínas pueden efectuar movimientos de traslación dentro de la bicapa.
3. Asimetría en la disposición de los lípidos, las proteínas y especialmente los glúcidos.
Al microscopio electrónico las membranas se observan como una estructura trilaminar, formada por dos capas obscuras periféricas y una capa central clara, no solo se observa en la membrana plasmática, sino también fue observada en las membranas de todos los organismos membranosos, por lo que surge el concepto de unidad de membrana.
El aspecto trilaminar de las membranas celulares se debe al depósito de osmio (durante la preparación de la muestra) sobre los grupos hidrofílicos, que se localizan en ambas superficies de la membrana (fig. 3.25).
La membrana plasmática se organiza como una doble capa continua, delgada, de 7,5nm de espesor. En algunas células, por fuera de la membrana, existe una cubierta celular que la cubre y la protege. A la zona externa de la membrana plasmática, abundante en glúcidos, se le conoce con el nombre de glicocalix. Con frecuencia se absorben glicoproteínas y proteoglicanos, secretados por la propia célula, a esta cara externa de la membrana plasmática.
3.3. FUNCIONES GENERALES DE LAS MEMBRANAS
Después de comprender bien la estructura supramolecular de las membranas celulares, es posible analizar en más detalle las distintas funciones que realizan estas estructuras. Algunas membranas, como las que rodean y delimitan cada organelo, tienen algunas particularidades que deben ser analizadas con cada uno de estos; por ejemplo, la membrana del retículo endoplasmàtico rugoso tiene la función de trasladar proteínas hacia el interior de ese organelo, las que posteriormente tendrán destinos específicos; la membrana plasmática tiene la función de servir de anclaje a algunos componentes del citoesqueleto para mantener la forma celular; la membrana interna de la mitocondria tiene varias biomolèculas que participan en el transporte de electrones en el proceso de respiración celular. Pudiera afirmarse que todas las propiedades de la materia viva (o del protoplasma), las más relacionadas con la membranas son el intercambio de sustancias, energía e información y la irritabilidad. A las membranas se le atribuyen las siguientes funciones:
1. Delimitan y aíslan las células y organelos
2. Intercambio de sustancias
3. Generación y/o transmisión de potenciales eléctricos
4. Comunicación e interacción con otras células y la matriz extracelular.
DIFUSION SIMPLE
La difusión simple es la forma más simple de paso de sustancias a través de una membrana de la célula e incluye:
1. La difusión a través de la matriz lipidica
2. Los poros y los canales
3. La ósmosis
La difusión simple es un mecanismo por el cual muchas sustancias se trasladan de un lado a otro de la membrana, a favor de su gradiente de concentración, por lo que no necesitan de ningún transportador que facilite este paso, siendo un proceso totalmente espontáneo que no requiere de energía (∆G<0).
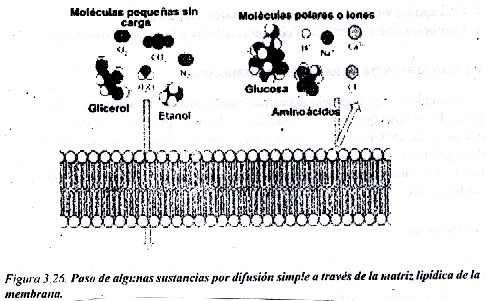
Por la difusión simple a través de la matriz lipídica pasan sustancias apolares y polares pequeñas sin carga. Las moléculas pequeñas que no tienen carga como los gases (CO2 y O2 ), el amoniaco, la urea, el glicerol, el etanol, que se mueven de manera libre y al azar por su energía cinética, pueden atravesar la membrana por difusión simple a través de la bicapa lipídica. Si las moléculas son polares y mas grandes como los monosacáridos, y polares con carga eléctrica como un aminoácido o un nucleótido, o un catión como el K- o el Ca2- , ya no pueden atravesar por este mecanismo (fig. 3.26). Otras sustancias que pasan fácilmente a través de las membranas por difusión simple son las sustancias apolares, como por ejemplo el benceno, las hormonas esteroides como la aldosterona, el cortisol, y las hormonas sexuales.
La velocidad de paso de las sustancias por difusión simple viene dada por la expresión siguiente:
V=P(Gradiente)= P (Aext-Aint)
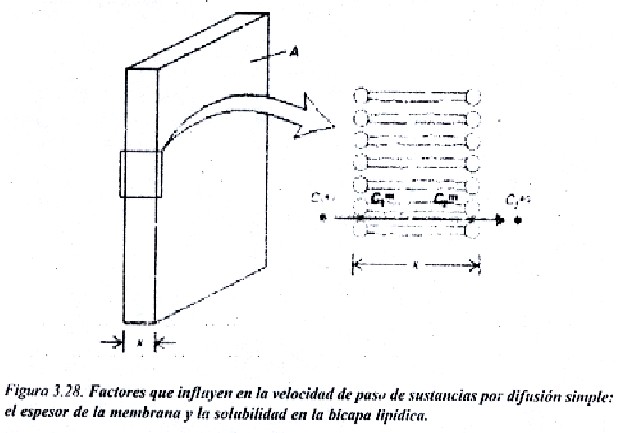
Donde P es coeficiente de permeabilidad, que depende a su vez del espesor de la membrana, del coeficiente de partición de la sustancia y de su coeficiente de difusión D. Esto se representa gráficamente en la figura 3.27, en la cual se observa que existe una relación lineal entre la velocidad de paso de la sustancia y el gradiente de concentración.
Como el coeficiente de difusión se puede considera similar para muchas sustancias que atraviesan la membrana por este mecanismo, y el espesor de la membrana tampoco es muy variable entre distinta membranas, el factor mas importante del cual depende P es del coeficiente de partición o relación entre la solubilidad de la sustancia en agua y en el componente lipídico Cm/Cagua (fig. 3.28).
Hay que tener en cuenta que cuando la sustancia que atraviesa se une de manera preferente a macromoléculas confinadas en uno de los lados de la membrana, o se modifica químicamente después que ha atravesado, mientras se continúe manteniendo el gradiente con la sustancia libre o no modificada esa sustancia continuara atravesando la membrana. Por ejemplo, si se mida la concentración de oxigeno en el interior del eritrocito es superior a la del plasma, pero resulta que la mayor parte del oxigeno esta unido a la proteína hemoglobina y existe otra fracción libre; si el gradiente de concentración con la forma libre es favorable para la entrada de este gas, el proceso de transporte continuara.
¿Y cuando la velocidad del paso neto de sustancia se detiene? Cuando ocurre lo que se muestra en la figura 3.29, es decir, cuando el paso de sustancias en un sentido y en otro es el mismo, el gradiente de concentración es igual a 0 y el sistema esta en equilibrio termodinámico.
Poros y Canales
Para muchas sustancias son precisos otros medios para poder pasar de manera rápida, al interior o exterior de la célula por las membranas. Uno de estos medios es la existencia en las membranas de poros y canales, formados por proteínas transmembranales. La diferencia fundamental entre ambos, aunque en algunos casos particulares puede ser sutil, es que los canales son mucho mas selectivos y estos pueden estar abiertos o cerrados, mientras que los poros siempre permanecen abiertos y son menos selectivos. Los canales por otro químico, a los que se abren por cambios de voltaje de la membrana. La cinética de este proceso es similar a bajas concentraciones de la sustancia a la de la difusión libre a través de la matriz lipídica.
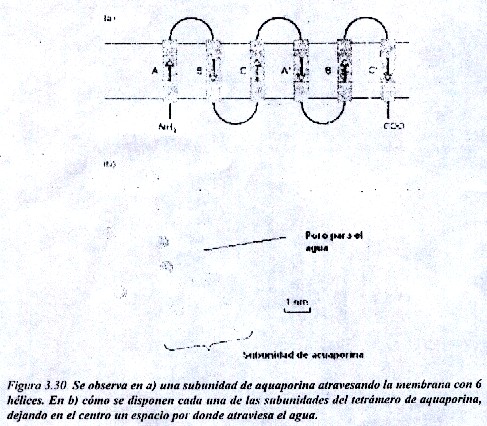
Un ejemplo de poro es el que forman las proteínas aquaporinas. El paso el H2O que es una molécula polar, se considero durante mucho tiempo que se realizaba únicamente por difusión a través de la matriz lipídica, pero en la actualidad se conoce que se realiza también y de manera muy favorecida a través de poros formados por estas proteínas.
Estas proteínas parecen jugar un rol relevante en las células con una alta permeabilidad al agua, como las células de los túbulos renales y los eritrocitos (fig. 3.30).
Otro ejemplo de difusión a través de poros es el intercambio de Cl- y –HCO3 (cloruro y bicarbonato), que es esencial para la función de los eritrocitos. Las membranas del eritrocito son muy poco permeables para estos dos iones, y por eso se produce un rápido intercambio de ellos en la proporción de 1:1 a través de poros formados por proteínas transmembranales, en particular en este caso se trata de proteína de banda 3, que atraviesa la membrana y forma un poro a través del cual pueden pasar los dos aniones. Existen algunos antibióticos como la Gramicidina A que actúan formando también poros con el interior hidrofílico en la membrana.
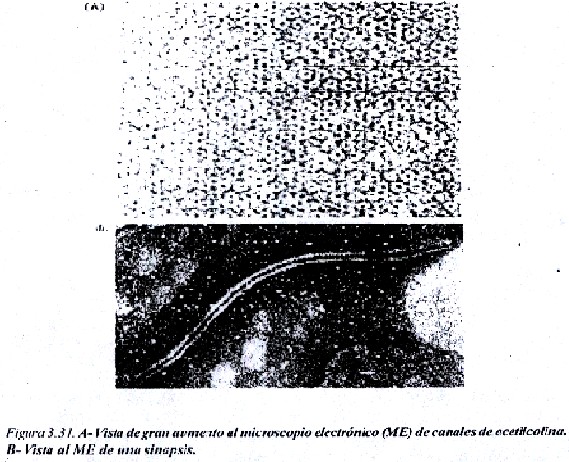
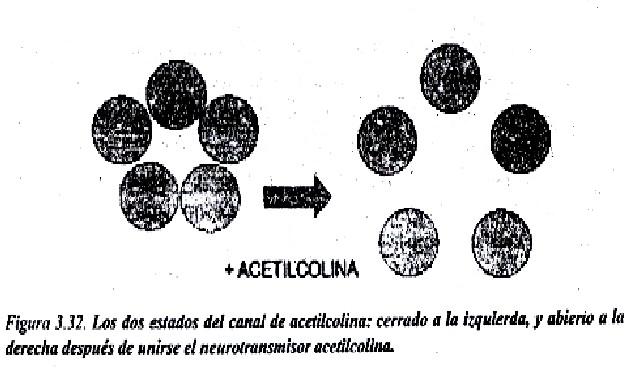
En las figuras 3.31 y 3.32, se trata de un canal, el canal de acetilcolina que existe en la membrana plasmática de las neuronas. La acetilcolina es un neurotransmisor que al unirse de manera muy especifica al receptor de acetilcolina, permite que se produzca un cambio conformacional en algunas subunidades proteicas de ese canal y entonces se abre, permitiendo el paso de iones de Na+ al interior de la neurona. En este caso se trata de un canal activado por un ligando química (la acetilcolina). Pasado cierto tiempo la acetilcolina se degrada por acción de una enzima, la colinesterasa, el receptor de acetilcolina queda libre y el canal se cierra.
A continuación, en la figura 3.33 se muestra otro canal, en este caso un canal dependiente de voltaje. Se trata de canales de Na+, que tiene entre 0.3 y 0.5 nm de diámetro y se encuentra también en las membranas plasmáticas de las neuronas. Estos canales se abren o cierran en dependencia de los cambios de voltaje, experimentando las proteínas que los forman cambios conformacionales que permiten el paso mas fácil del ion. La función de estos canales es imprescindible en la transmisión de los impulsos nerviosos a lo largo de estas células.
Un canal sobre el que se han realizado muchas investigaciones científicas en los últimos años es le canal CFTR localizado en la membrana plasmática de células del ser humano, por donde pasan los aniones de Cl- en dependencia de la unión con un ligando químico, el ATP. La proteína de este canal no se expresa adecuadamente en los enfermos con la enfermedad genética (autosómica recesiva) denominada Fibrosis Quìstica. Los enfermos, que se diagnostican en las primeras edades de vida, padecen de trastornos nutricionales e infecciones respiratorias frecuentes. Se han hecho intentos recientes de terapia génica para curar esta enfermedad.
El paso de sustancias a través de estos poros y canales es también un proceso con un (∆G<0), es decir, espontaneo en las condiciones de la célula, aunque como ya se dijo, en el caso de los canales estos necesitan abrirse ya sea por un ligando químico o por voltaje. Se ha calculado que la velocidad de paso de sustancias por los canales es de 108 moles/s.
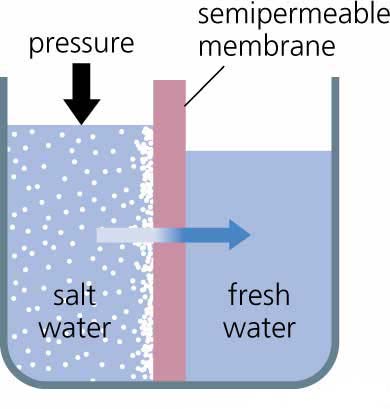
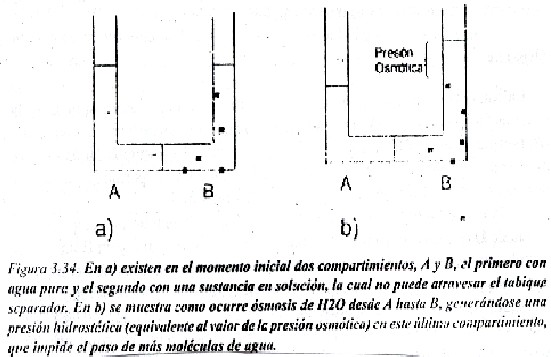
La osmosis es un caso particular de la difusión simple, muy importante en muchos fenómenos fisiológicos, donde lo que ocurre es difusión de agua. Ocurre porque se encuentra un soluto o sustancia disuelta a ambos lados de una membrana en diferente concentración y la membrana no es permeable al soluto pero si deja pasar libremente el agua por difusión simple.
En el esquema de la figura 3.34 en a) se ha representado un recipiente en forma de U que contiene en un compartimiento A, agua pura, y en B un soluto disuelto en agua a determinada concentración, separados esos dos compartimentos por un tabique que deja pasar libremente el agua, pero no las moléculas de soluto; en b) como proporción relativa de moléculas de agua en A es mucho mayor que en B, el agua difunde por el gradiente de concentración desde A hacia B, hasta que se genera un desnivel de liquido que genera una presión hidrostática (ñgh donde ñ=densidad del liquido, g=gravedad y h=altura) que se opone y llega a impedir el paso neto de mas agua desde A hasta B. a la presión necesaria para impedir la osmosis o difusión del agua se denomina presión osmótica, y depende de acuerdo a la ley de Van`t Hoff del numero de partículas disueltas y de la diferencia de concentraciones de las soluciones que se encuentran en A y B. Cuanto mayor sea esta diferencia mayor será la presión osmótica.
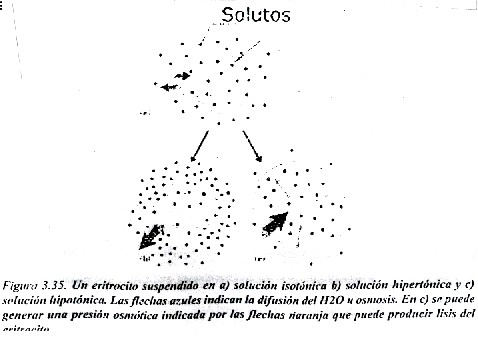
Los fenómenos osmóticos tienen gran importancia tanto en condiciones normales como en determinadas enfermedades del ser humano.
En la figura 3.35 se representa una célula, pudiera ser por ejemplo un glóbulo rojo de la sangre o eritrocito, una célula por cierto bastante especial. En a) se aprecia que la célula esta suspendida en una solución isotónica , o sea que tiene una concentración de soluto es igual a ambos lados de la membrana, no existe un gradiente de agua y no hay paso neto por lo tanto de esta sustancia a través de la membrana. En el caso de b) ocurre que la concentración del soluto en el exterior es mayor que en el interior celular, es decir, la célula esta suspendida en una solución hipertónica, como existe una proporción relativa mayor de moléculas de H2 O en el interior hay un flujo neto de agua, ocurre osmosis o difusión del agua hacia el exterior, y la célula puede deformarse y afectarse su función. Por ultimo, en el caso c) la concentración de soluto en la solución es menor que la del interior celular, es decir, es una solución hipotónica, por lo que el desplazamiento de las moléculas de H2 O va dirigido hacia el interior de la célula y puede llegar a ocurrir incluso que, por la gran presión osmótica que ejerce la solución en el interior celular, la membrana no resista y llegue a romperse. Esta es la razón por la que en la práctica médica es conveniente administrar soluciones isotónicas en vena, como por ejemplo NaCl 0.15 moles/l.
DIFUSIÓN FACILITADA O TRANSPORTE PASIVO
Se refiere a las moléculas que necesitan, por su naturaleza polar y su tamaño, de proteínas especificas de la membrana para poder pasar de un lado a otro de la misma, que actúan como transportadores. A esta forma de paso sustancia de un lado a otro de las membranas se la ha denominado difusión facilitada, porque la presencia de proteínas transportadoras en la membrana es lo que permite que las sustancias puedan atravesar la membrana.
El transporte pasivo se realiza siempre a favor de gradiente de concentración de la sustancia y no requiere de energía externa, es decir, es un proceso espontaneo como la difusión simple (∆G<0).
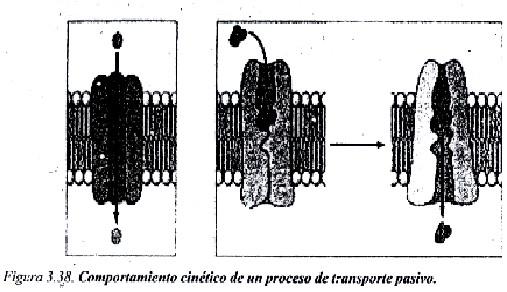
El transporte pasivo lo realizan proteínas localizadas en las membranas conocidas como Permeasas. Las sustancias que se transportan por este mecanismo se unen al transportador por un mecanismo de reconocimiento molecular, parecido a lo que ocurre cuando un sustrato se une al centro activo de una enzima, pero a diferencia de las enzimas, la sustancia que se transporta no es transformada, sino que se traslada al otro lado de la membrana sin ninguna transformación.
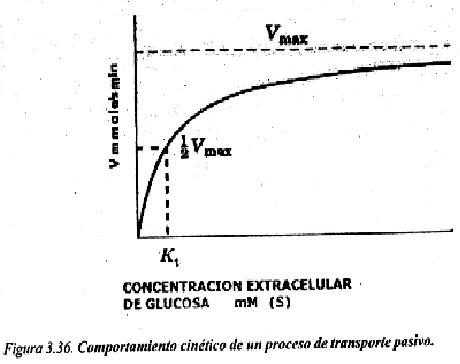
En el proceso de transporte de la sustancia, las Permeasas experimentan cambios conformacionales (al menos existen 2 estados conformacionales de estos transportadores). Como el número de transportadores es limitado en una membrana y se debe producir la unión por interacciones débiles de la proteína con cada molécula de soluto (o muy pocas moléculas), puede existir un fenómeno de saturación cuando la concentración de la sustancia a transportar es muy elevada. La velocidad de paso de sustancia por este mecanismo es más lenta que por difusión simple, y se ha estimado en 102 a 104 moles/s.
En la figura 3.36 se muestra el comportamiento cinético de este mecanismo es similar al de las enzimas; se puede observar que cuando la concentración de un soluto como la glucosa es baja, el incremento de velocidad es lineal, pero si la concentración llega a ser muy elevada, ya la velocidad tiende a hacerse constante.
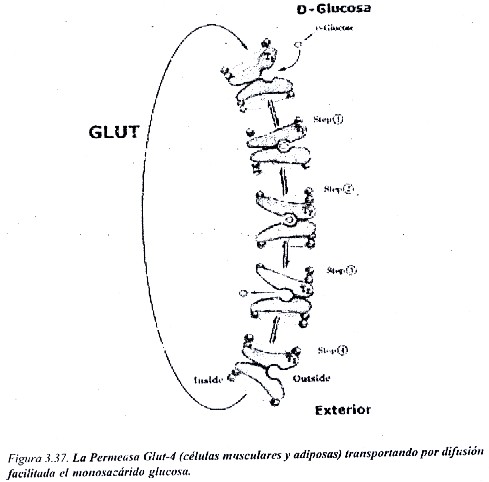
Un ejemplo de Permeasas son los transportadores del monosacárido glucosa (Glut) en las membranas plasmáticas, que tienen diferente distribución en los tejidos. El Glut-1 y el Glut-3 presentan una afinidad relativamente alta por la glucosa (Kt=1 mmoles/L), se encuentran en casi todas las células, y su función consiste en mantener una captación continua de glucosa es por las células. El Glut-2 se localiza en el hígado y su afinidad por la glucosa es menor (Kt= 15-20 mmoles/L); el Glut-5 se expresa en células musculares y del tejido adiposo (Kt=5 mmoles/L); el Glut-5 en el intestino. En la figura 3.37 se muestran los dos estados conformacionales del Glut-4 cuando realiza el transporte de la glucosa hacia el interior celular.
En la figura 3.38 se muestra que en el caso de las Permeasas se establece la unión con la molécula transportada, aunque sea por interacciones débiles, pero en el caso de los canales no ocurre así.
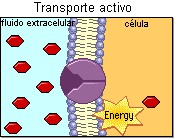
TRANSPORTE ACTIVO
La difusión facilitada es útil en muchos procesos celulares, pero en algunas situaciones es imprescindible que las células sean capaces de transportar sustancia en contra de gradientes de concentración, algunas veces muy desfavorables. Por ejemplo, en el retículo sarcoplasmático de las células musculares es necesario en algunas circunstancias trasladar átomos de Ca2+ contra un gradiente de 30 000/1, lo cual es una barrera impresionante. El transporte que se realiza cuesta arriba (fig. 3.39), en contra de un gradiente de concentración, se conoce como transporte activo, y en este caso también son necesarias proteínas integrales de la membrana. No es un proceso espontáneo pues se requiere energía. Generalmente la energía se obtiene de la hidrólisis del ATP por la misma proteína transportadora (transporte activo primario), o de la energía suministrada por otro proceso como se muestra en la figura 3.40 (transporte activo secundario); los gradientes iónicos por ejemplo, como se verá mas adelante, son muy importantes para la fosforilación oxidativa en la respiración celular y para la excitabilidad de los músculos y de las células nerviosas.
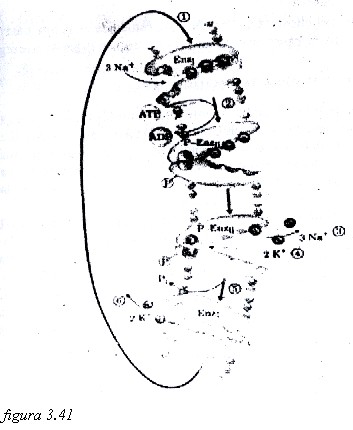
El ejemplo fisiológico mejor conocido de transporte activo lo constituye la bomba de Na-K o ATPasa Na+ / K+. Normalmente el fluido que rodea a las células tiene aproximadamente 140 mmoles/L (o mEq/L) de Na+ y 4 mmoles/L de K+. Sin embargo, el líquido intracelular tiene una concentración de Na+ de aproximadamente 10 mmoles/L y de 150 mmoles/L de K+. A pesar de que puede existir un paso extraordinariamente lento de estos iones por algún grado de difusión simple, estas desigualdades acabarían por desaparecer con el paso del tiempo si no existiera un mecanismo que mantuviera la entrada de K+ y la salida de Na+. Este movimiento se realiza mediante un sistema de transporte activo conocido por el nombre de la bomba de Na+/K+ (ATPasa de Na+/K+).
La bomba de Na+/K+ es una proteína integral de la membrana, tetramérica, formada por dos subunidades grandes (subunidades á), de 120 kDa cada una, y dos más pequeñas (β), de 55 kDa cada una. Las subunidades á intervienen directamente en el mecanismo de transporte y tienen actividad enzimática para hidrolizar el ATP. La energía libre para bombear los dos iones contra un gradiente de concentración la suministra precisamente este nucleótido, y en el proceso de transporte se extraen hacia el exterior celular 3 Na+ cada vez que ingresan a la célula 2 K+ (figura 3.41), favoreciéndose así una acumulación de cargas positivas en el exterior y la creación de un potencial eléctrico.
A los sistemas, como la bomba de Na+/K+, que transportan sustancia en sentidos opuestos, se les denomina antiportes. Si fuera como el caso del transporte activo secundario de la figura 15, en dos sustancias son transportadas en el mismo sentido, se les denomina simportes, y se les llama uniportes a los que transportan una sola sustancia.
La bomba de Na+/K+ también contribuye a regular el volumen celular, ya que coopera en el mantenimiento de la concentración de solutos dentro y fuera de la célula.
Un detalle interesante es que esta bomba es inhibida por la ouabaína, un fármaco que se utiliza para el tratamiento de la insuficiencia cardiaca, ya que al inhibirse la bomba se acumula calcio en el interior de la célula muscular cardiaca; este ion no puede intercambiarse de manera efectiva, por medio de un sistema antiporte con el Na+ extracelular por estar inhibida la bomba sodio-potasio; el calcio al acumularse incrementa la intensidad de la contracción muscular.
Otro ejemplo de transporte activo primario, es el transportador de Ca2+ de la membrana del retículo endoplasmático. En las células musculares esta bomba constituye el 80% de las proteínas de membrana de este organelo (denominado en la célula muscular retículo sarcoplasmático9 y es capaz de mantener una diferencia de concentración de Ca2+ de 15000 a 1 (0.1 micromol/L vs 1.5 mmol.L) entre su interior y el citosol. Otra bomba muy estudiada es la que transporta H+/k en el estómago, capaz de crear un pH muy bajo en la luz de éste órgano.
ENDOCITOSIS, EXOCITOSIS Y TRANSCITOSIS
Algunas sustancia, generalmente macromoléculas y agregados supramoleculares, penetran a las células utilizando un mecanismo muy diferente a los que fueron estudiados anteriormente, que implica una gran deformidad de la membrana plasmática, al producirse una invaginación que engloba a esa sustancia y la traslada al interior; este es el caso de la endocitosis. Por tanto, estas sustancias no atraviesan la membrana.
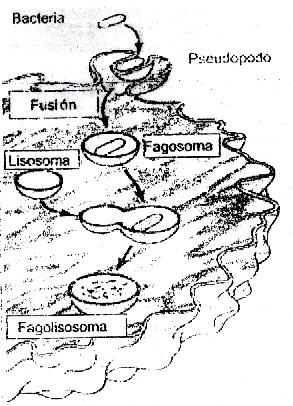
Por este mecanismo se produce el ingreso de grandes partículas como bacterias o restos de éstas, en un proceso mediado por la actina y conocido por fagocitosis (del griego phagein: comer) en los macrófagos. En esta forma de endocitosis las vesículas que ingresan a la célula conteniendo el material externo son de gran tamaño, de 250 nm de diámetro o mayores.
En la figura 3.42 se observa como después de penetrar a la célula la bacteria o restos de esta, se origina un fagosoma que después se une a un lisosoma primario para ser degradados los restos de la bacteria por las enzimas hidrolíticas de ese organelo. También se producen invaginaciones más pequeñas de la membrana para englobar partículas de menor tamaño, como agregados supramoleculares, y también macromoléculas y sustancias líquidas, proceso conocido como pinocitosis (del griego pinein: beber). En este caso las vesículas que se originan en la membrana tienen menor diámetro, de aproximadamente 100 nm. A la forma inespecífica de este mecanismo de endocitosis se le conoce por endocitosis o pinocitosis inespecífica y a la forma en que penetran sustancias por endocitosis pero uniéndose a receptores de la membrana plasmática muy específicos, se conoce como endocitosis mediada por receptores.
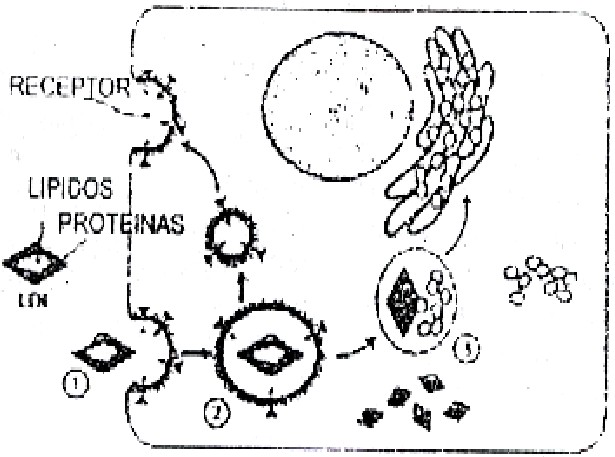
En la endocitosis mediada por receptores, donde son necesarios receptores específicos en la membrana plasmática, está implicada una proteína que rodea a esas vesículas membranosas denominada clatrina, y también otras proteínas; se requiere asimismo de la energía aportada por el ATP. El hierro procedente de la sangre ingresa a las células por este mecanismo. También ingresan por este mecanismo las lipoproteínas del plasma sanguíneo LDL, que transportan el colesterol plasmático hacia los tejidos periféricos: al llegar a las células de los tejidos periféricos, las LDL se unen a un receptor específico de la superficie celular, y se inicia el proceso de endocitosis. Las LDL son partículas esféricas, con un núcleo de lípidos en el centro donde se encuentra el colesterol y una superficie externa cubierta de varias proteínas, en particular la ApoB-100, que es precisamente el ligando que reconoce el receptor de membrana (figura 3.43).
Varias vesículas endocíticas tienden a agruparse y formar el endosoma, que tiene en su interior un pH relativamente más bajo que el del citosol, lo cual es un factor relacionado en muchos casos con la separación del ligando y el receptor o de la sustancia importante que ingresó a la célula como en el caso del hierro. También el mecanismo de degradación puede diferir. Ya se ha referido que las LDL, se degradan al unirse el endosoma al lisosoma, pero la proteína transportadora del hierro que se incorpora a las células por endocitosis no se degrada; sólo el hierro es separado de la transferían, al acidificarse el interior del endosoma, y es incorporado a la célula. Estas vesículas endocíticas también intervienen en el movimiento y reciclaje de receptores y ligandos dentro de la célula (figura 3.43).
La exocitosis es el proceso inverso, por el cual las vesículas membranosas conteniendo por ejemplo gránulos de secreción se dirigen a la superficie celular para fusionarse con la membrana plasmática y verter se contenido al exterior. La trancitosis es el paso a través de la célula de algunas sustancias al combinarse los mecanismos de endocitosis y exocitosis. En los endotelios por ejemplo, se observa con frecuencia este mecanismo de trancitosis, que pasa por alto los organelos celulares, y también el transito de inmunoglobulinas de la leche materna a través de las células intestinales del recién nacido es por trancitosis.
POTENCIAL DE MEMBRANA DE REPOSO
Muy asociada con la función de transporte de iones, otra función de las membranas de las células eucariotas es mantener un potencial de membrana en reposo, lo cual es la base de la transmisión del impulso nervioso, de la contracción y de otras funciones especializadas de células de nuestro organismo. Este potencial de membrana en reposo se origina, como se verá más adelante, es por el flujo de K a través de canales proteicos que existen en las membranas plasmáticas, principalmente de células nerviosas y musculares. Es equivalente a 200 KV por cm. y como dato curioso, las líneas eléctricas de una ciudad tienen un gradiente de 200 KV por Km., lo que indica la elevada capacitancia de las membranas biológicas.
Exterior+++++++++++++++++++++++++++++++++++
____________________________________
Interior -------------------------------------------------------------
A través de las membranas, como ya se ha mencionando, se crean desequilibrios iónicos por los sistemas específicos de transporte activo, y estas diferencias de concentración de los distinto iones unido a la permeabilidad selectiva de la membrana para cada uno, a través de canales proteicos, es lo que da lugar a un potencial eléctrico para un sistema en equilibrio con un ∆G=0, la conocida como ecuación de Nernst, que permite calcular el potencial eléctrico que se crea para un ion a través de una membrana. Según esta ecuación se mantiene de alguna manera una diferencia de concentración iónica a través de una membrana es en alguna medida permeable a ese ion. Si la membrana fuera totalmente impermeable, el potencial sería =0.
Δψ (mV)= 61 log10 CFUERA / CDENTRO
Suponiéndose que se trate del catión K+, y suponiendo una concentración extracelular de 4 mmol/L, e intracelular de 145 mmol/L, sustituyendo esos valores en la ecuación de Nernst se obtiene que:
Δψ (mV)= 61 log10 4/145=-95 mV
Lo que indica que la membrana estaría polarizada con un potencial de 95 mV y con cargas negativas en el interior. El K+ , por su gradiente de concentración tiende a difundir hacia el exterior a través de un canal proteico, conocido como canal de escape de potasio, dejando como es lógico cargas negativas en el interior, que se oponen por otra parte a la salida del catión. Llega un momento en que no ocurre difusión neta de K+ , hacia el exterior, pues alcanza un equilibrio entre dos tendencias opuestas, el gradiente de concentración del ion que tiende a sacarlo del interior y las cargas negativas que quedan por detrás que tienden a retenerlo.
Si se supone una concentración de Na+ fuera de la célula de 140 mmol/L y en el interior de 10 mmol/L, entonces el potencial calculado sería de +69 mV. También se puede hacer el cálculo para el Cl.
Para calcular la influencia de diferentes iones a la vez se utiliza la ecuación de Goldman, que se puede expresar como:
Δψ (mV)= 61 log10 ( ΣPC+FUERA + ΣPC- DENTRO / ΣPC+ DENTRO + ΣPC-FUERA )
Donde el símbolo son las sumatorias para los cationes y aniones con una concentración C y un coeficiente de permeabilidad relativa P a través de las membranas, a través de canales proteicos que existen para estos iones y que depende del número de canales iónicos que hay or unidad de superficie y del número de iones que dejan pasar por segundo. Para el caso K+ , Na+ y Cl- esta ecuación equivale a:
Δψ (mV)= 61 log10 CNa+ (fuera)PNa+ + CK-+ (fuera) PK+ + CCl- (dentro) PCl-
CNa+ (dentro)PNa+ + CK-+(dentro) PK+ + CCl- (fuera) PCl-
Se puede apreciar que si el coeficiente de permeabilidad P se hace 0 para algunos de esos términos, entonces esta ecuación se reduce a la de Nernst. Cuando se introducen los coeficientes de permeabilidad P para cada uno d estos iones (en las condiciones de la célula PK+ =1, PNa+ =0.1 y PCl- = 0.1) y sus concentraciones se obtiene un valor de ÄØ=-54 mV, lo cual se aproxima al potencial de medido en condiciones experimentales, que se ha reportado entre 60 y – 90 mV y también al calculado para el K+. Obsérvese que la permeabilidad para el Na+ y Cl- es muy baja.
Las permeabilidades de los canales proteicos de Na+ y K+ son precisamente las que pueden experimentar grandes cambios durante la conducción de un impulso nervioso, mientras que la permeabilidad de los canales de cloruro no sufre grandes cambios durante ese proceso y por eso a este anión se le considera menos importante.
Por todo lo anterior se considera que el catión K+ es el principal responsable del potencial de membrana en reposo. Desde luego, fue necesario previamente que la bomba sodio-potasio creará el elevado gradiente de concentración para este catión, y adicionalmente esta bomba tiene alguna influencia directa porque extrae de la célula 3 Na+ por cada 2 K+ que entra, es decir ve favoreciendo una mayor acumulación de cargas positivas en el exterior y negativas en el anterior pero esa contribución se considera modesta de unos 4 mV.
El potencial de membrana en reposo en las grandes fibras musculares esqueléticas es aproximadamente el mismo que en las grandes fibras nerviosas -90 mV. Sin embargo, tanto en las fibras musculares lisas así como en muchas de las neuronas del sistema nervioso central, el potencial de membrana es menor, de -40 a -60 MV.
En resumen los potenciales electrónicos aislados de sodio y potasio a través de la membrana, pero principalmente de este último catión, conjuntamente con la acción electrogénica de la bomba e sodio-potasio, aunque siendo menos importante este último factor, a corto plazo es lo que determina el potencial de membrana en reposo.
POTENCIAL DE ACCIÓN
Una propiedad característica de las células vivas es, como se ha analizado una distribución desigual de los cationes u aniones en las caras externa e interna de la membrana plasmática. Esto produce el potencial de membrana en reposo que puede tener valores entre -60 y +90 mV.
Las señales nerviosas se transmiten mediante potenciales de acción, que son cambios bruscos de ese potencial negativo de membrana en reposo, a un potencial positivo, y se termina con un regreso rápido al potencial negativo todo esto ocurre en 2 o 3 ms. Estos potenciales de acción se van desplazando a lo largo de toda la fibra nerviosa hasta alcanzar el extremo de la misma, donde por transmisión química o eléctrica en la sinapsis o punto de contacto de una célula nerviosa con otra, se saltan de la célula presináptica.
Debe tenerse en cuenta que las membranas de las células nerviosas tiene canales iónicos para el Na+, K+, Cl- y Ca2+, los cuales tienen diferente grado de permeabilidad para cada ión y en general se mantienen cerrados y se abren durante un lapso muy corto para dejar pasar esos iones, pero estos canales pueden abrirse o cerrarse por cambios de voltaje o de estímulos químicos en algunos casos. La concentración de algunos de estos iones pueden influir en la permeabilidad del otro, así por ejemplo se conoce que un aumento del Ca2+ extracelular reduce la permeabilidad de los canales de Na+ y desde luego al analizar la ecuación de Goldman anteriormente descrita, se comprende que la elevación o disminución en las concentraciones de algunos de los iones en el líquido extracelular puede afectar el potencial de membrana en reposo y desde luego también los potenciales de acción.
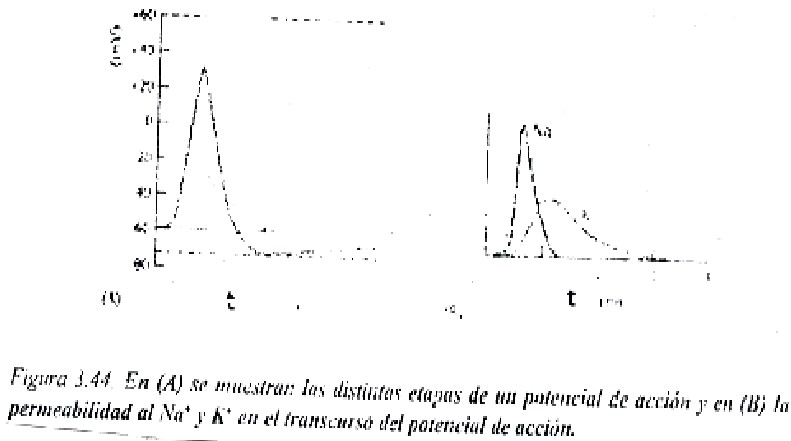
En un potencial de acción se pueden distinguir las siguientes fases (figura 3.44):
1. Fase de reposo. Es el potencial de la membrana en reposo, que existe antas de que comience el potencial de acción y que tiene valores cercanos a -70 mV.
2. Fase de despolarización. Ante un estímulo eléctrico de determinada intensidad, en algunos casos puede ser mecánico o químico, que sobrepase un límite o umbral con respecto al potencial de membrana en reposo, los canales de Na+ dependientes de voltaje se abren, lo que permite un paso súbito de estos iones hacía el interior celular. Con esto el potencial de membrana se hace más positivo y llega a invertirse la polaridad de la membrana, quedando ahora positiva por dentro y negativa por fuera. Esta despolarización brusca de la membrana se transmite a regiones vecinas, con lo que se inicia así un circulo vicioso de retroalimentación positiva: el estímulo eléctrico despolariza la membrana, se abren los canales de na+, se despolariza la región vecina de la membrana, se abren nuevos canales de Na+ , y así sucesivamente.
Por lo general, el estímulo para iniciar un potencial de acción debe elevar el potencial de membrana en reposo de 15 a 30 mV, por ejemplo, de -60 mV hasta -45 o a -30mV. Estos valores de -45mV o de -15mV serían en este caso el valor umbral para iniciar el potencial de acción (figura 3.44 ). En experimentos realizados se ha observado como la concentración de Na+ extracelular disminuye la amplitud del potencial de acción, que se recupero al elevar la concentración de nuevo en el líquido extracelular.
3. Fase de repolarización. Los canales de Na+ se cierran inmediatamente quedando inactivados y se abren entonces los canales de K+ dependientes del cambio de voltaje de la membrana (son algo distintos a los canales de K+ que dan origen al potencial de membrana en reposo y que también son conocidos como canales de escape de K+). Entonces una rápida difusión de iones potasio hacia fuera restablece el potencial de membrana en reposo.
4. Fase de hiperpolarización
Por un tiempo muy breve el potencial de membrana en reposo incluso se hace más negativo (hiperpolarización) porque algunos canales de K+ permanecen abiertos.
El potencial de acción se origina en una región muy pequeña de la membrana pero de ahí se transmite a las regiones vecinas, dando lugar a la propagación de esta señal eléctrica, porque se abren nuevos canales de sodio al cambiar la polaridad de la membrana. Una vez desencadenado el potencial de acción viaja por la membrana si las condiciones son adecuadas o no viaja en absoluto si no lo son, lo que se ha denominado como principio del todo y se aplica a todos los tejidos excitables normales.
Es importante el hecho que en una célula excitable no se puede originar un segundo potencial de acción cuando la membrana esta desmoralizada y mientras se mantengan los canales de Na+ inactivados conociéndose este momento como periodo refractario. Ni con estímulo fuerte es posible desencadenar un potencial de acción en el periodo refractario.
A largo plazo el papel de la bomba sodio-potasio, utilizando ATP, restablece los gradientes normales de los iones, pero se considera que las fibras nerviosas pueden transmitir hasta millones de impulsos antes que las diferencias de concentración hayan descendido hasta un punto en que cese la conducción del potencial de acción.
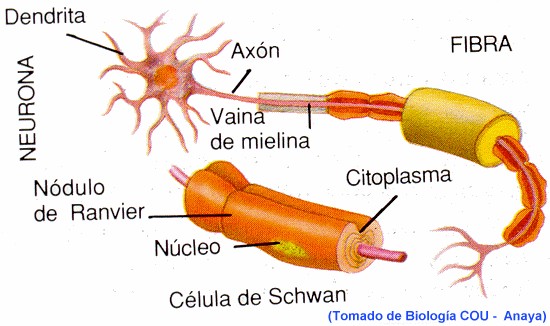
En otras células excitables como las células musculares también so originan y transmiten potenciales de acción, pero estos tienen algunas diferencias con los de las células nerviosas. En la célula muscular cardiaca por ejemplo, la membrana no se repolariza tan rápido y el potencial de acción alcanza una meseta que dura 0.2 a 0.3 s y hace que la contracción de la fibra cardiaca se prolongo por ese mismo tiempo. En las fibras nerviosas rodeadas de vainas de mielina, la transmisión del potencial de acciones muy rápida y se produce a “saltos” entre los estrechamientos conocidos como nódulos de Ranvier, y es donde único se produce en estas fibras nerviosas el intercambio de iones con el entorno.
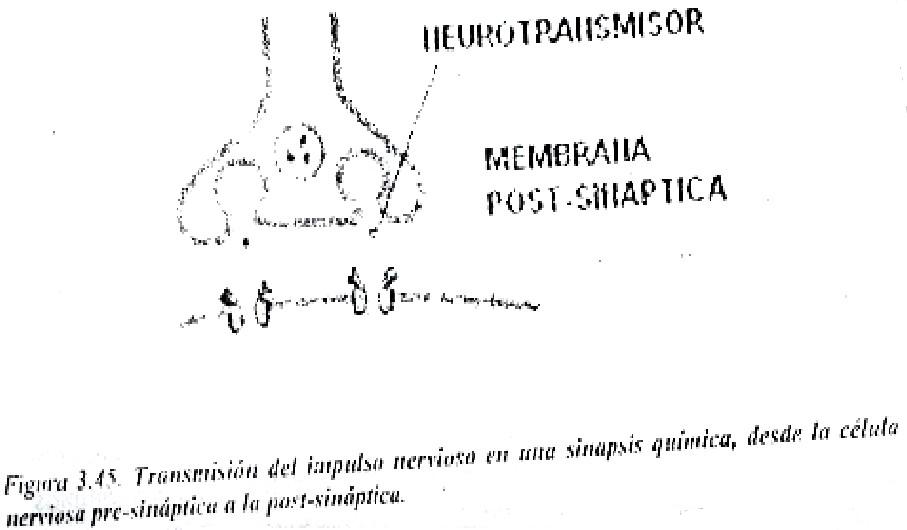
Entre una neurona y otra, la transmisión del impulso nervioso se produce como se mencionó, a través de neurotransmisores químicos frecuentemente. Al llegar un potencial de acción a la terminal pre-sináptica (figura 3.45) se abren canales de Ca2+, y como este catión está más concentrado en el medio extracelular, pasa la interior de la terminal axónica dando lugar a la liberación del neurotransmisor, como puede ser por ejemplo la acetilcolina este neurotransmisor se une a receptores específicos de la neurona post-sináptica, abriendo entonces canales de Na+ activados por ligando, con lo que se despolariza la membrana, pudiendo dar inicio, si se alcanza el valor umbral, al potencial de acción en la neurona post-sináptica.
Algunas sustancias, como la toxina de un pez conocido pez burbuja (manjar exquisito en el Japón después de que “chef” muy profesionales logran quitarle la toxina), denominada tetrodotoxina, bloque los canales de Na+ e impide la transmisión de los impulsos nerviosos. También los escorpiones tiene un tipo de toxina, la á-toxina, de naturaleza peptídico, que afectan los canales de este mismo ion, pero en este caso haciendo más lenta su inactivación. Una sustancia obtenida de las abejas, la apamina, afecta por el contrario los canales de K+ dependientes de voltaje, bloqueándolos. Estas toxinas en general han sido muy útiles en investigaciones científicas, como por ejemplo para calcular el número de canales por micrómetro cuadrado en la membrana o para descubrir que los canales de sodio están formados por una subunidad grande de unos 3000 kDa y una o varias subunidades pequeñas de 30 a 430 kda cada una. Por último, se han señalado 3 características del potencial de acción que lo hacen una onda de despolarización eléctrica de la membrana diferente a la conducción eléctrica mediante electrones en un cable eléctrico:
1. El potencial de acción no disminuye de manera apreciable con la distancia a que se transmite, ya que se renueva continuamente en cada punto a lo largo del axón.
2. El potencial de acción es un fenómeno de todo o nada. Si el estímulo es suficiente y rebasa determinado umbral se produce, y su magnitud es independiente del voltaje del estímulo. En virtud de esta característica se hace dicho que las redes neurales se parecen más a circuitos digitales que a los analógicos. Una neurona realiza o no realiza una descarga y estímulos más intensos no dan lugar a potenciales de acción mayores, si no simplemente se producen impulsos frecuentes.
3. Una vez que a pasado un potencial de acción, la región que queda detrás por encontrarse en periodo refractario es incapaz de generar un nuevo impulso durante algunos milisegundos. Por esta característica el sistema nervioso puede saturarse, pues tan solo puede manejar un número determinado de impulsos por segundo.
Comunicación e interacción con otras células.
Las células en un organismo pluricelular necesitan comunicarse e interactuar unas con otras continuamente. El proceso de comunicación celular que se estudiara con mas detalle posteriormente, se puede llevar a cabo por la existencia en la membrana plasmática de receptores, que son proteínas integrales que están presentes en todas las células de nuestro organismo. Al unirse estos receptores de manera muy específica mediante un proceso de reconocimiento molecular con señales del medio externo, se transmiten determinado cambios al interior celular que ocasionan en definitiva una respuesta celular.
Por otra parte en la membrana plasmática también se localizan proteínas que conocidas como Moléculas de Adhesión Celular (CAM por sus siglas en inglés), le permiten a las células establecer contactos con células vecinas o con la matriz extracelular. Entre ese grupo de proteínas se encuentran las Cadherinas, las Integrinas, las Selectinas y las de la familia de la Inmunoglobulinas.
En ese reconocimiento e interacción con células vecinas y con componentes de la matriz extracelular, las células se transmiten entre si señales de supervivencia o de muerte, migran en los procesos de diferenciación de los tejidos y en otros procesos como es el paso de algunos leucocitos de la sangre al espacio intersticial.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}